Population Genetic Analysis for Stock Enhancement of Silver Sea Bream (Rhabdosargus sarba) in Taiwan

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
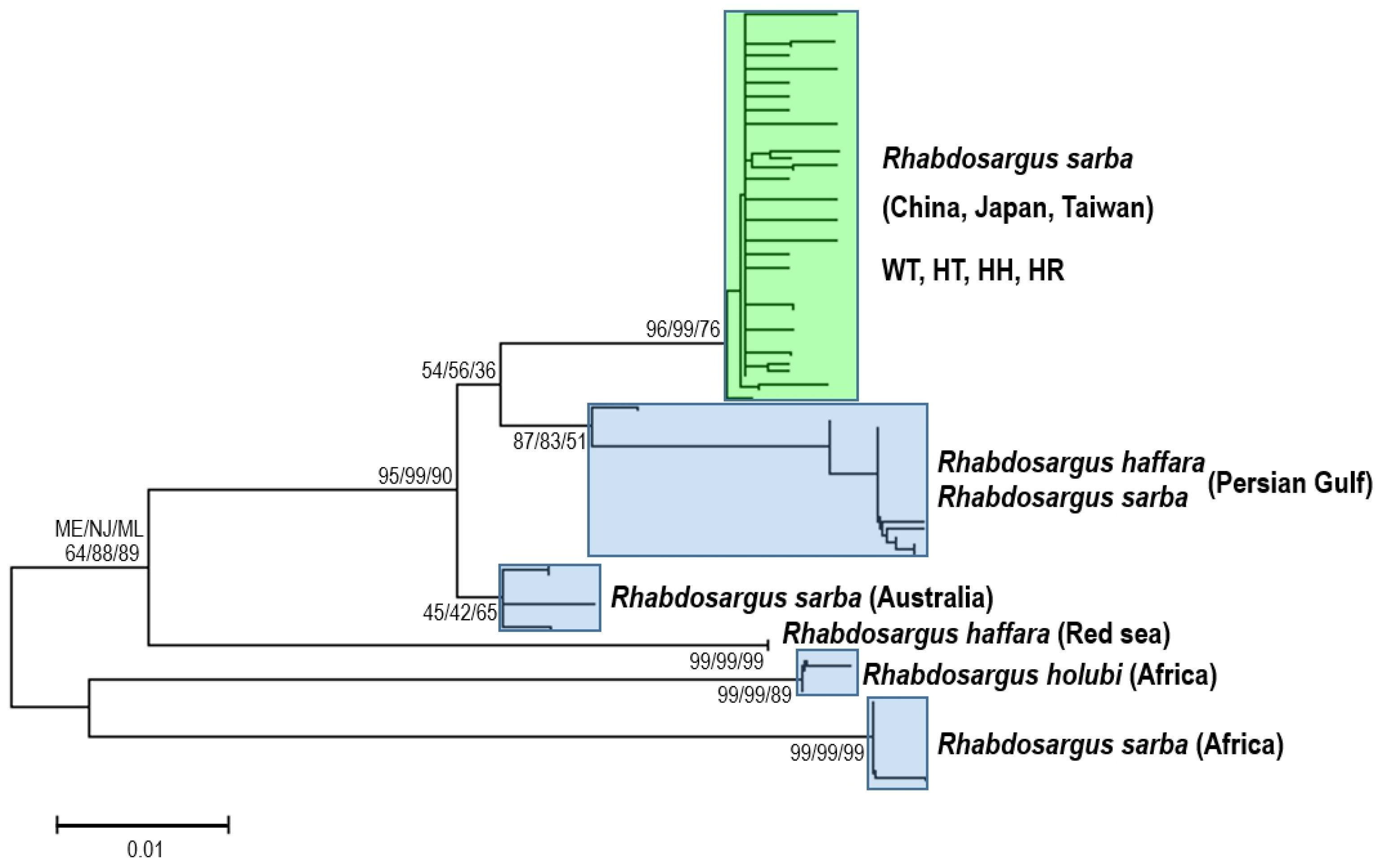
2.1. DNA Barcoding of Silver Seabream
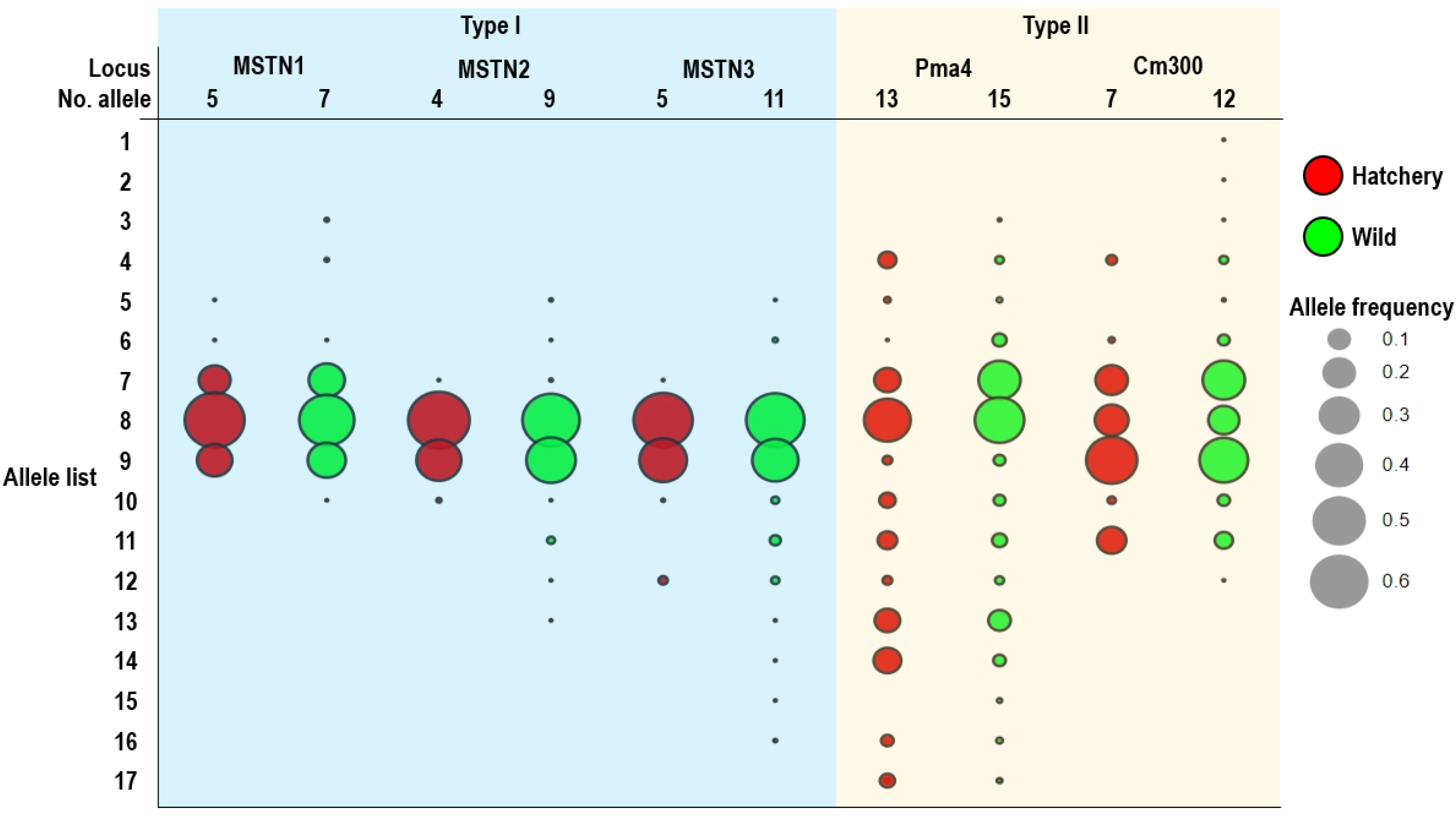
2.2. Genetic Diversity within Populations
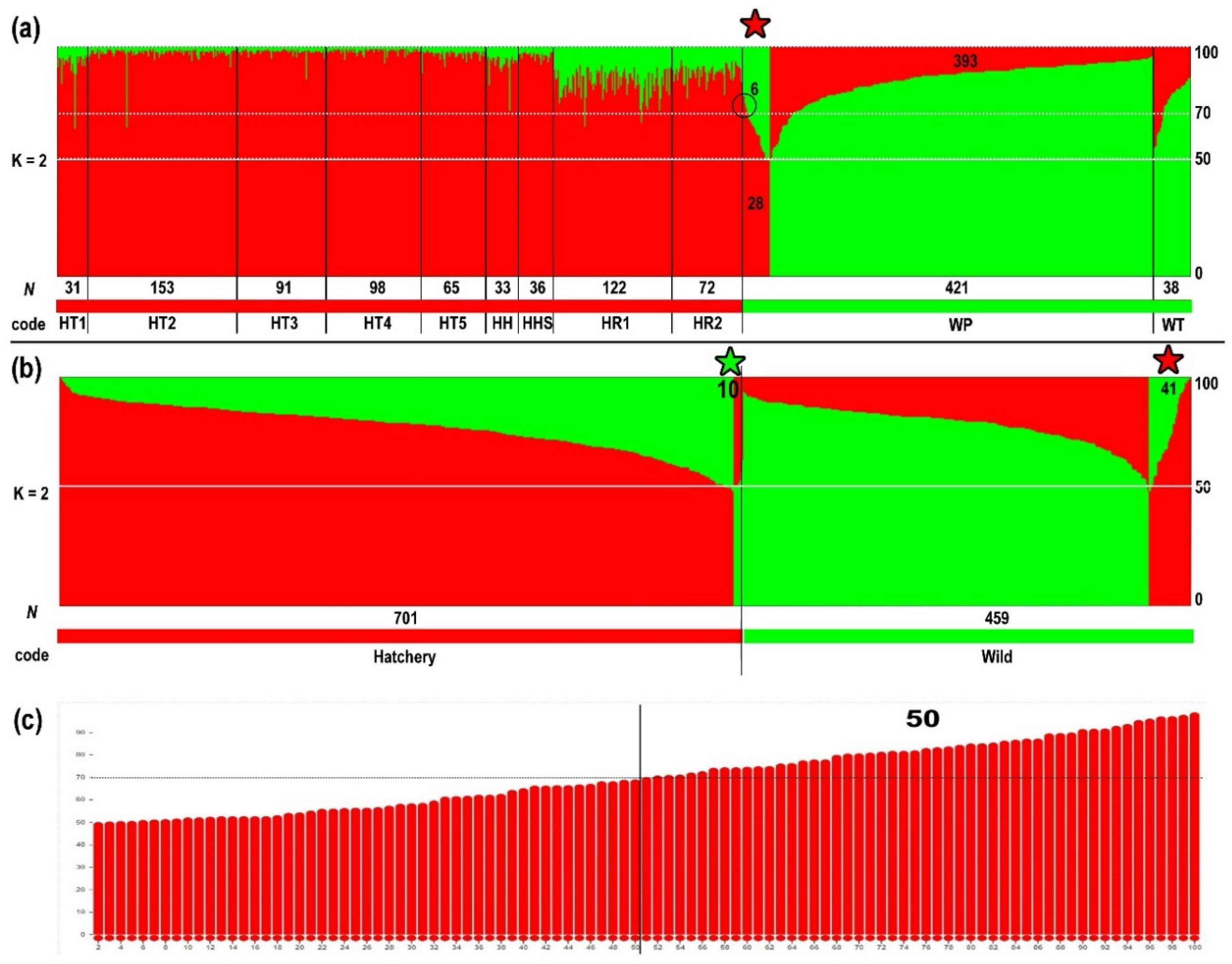
2.3. Genetic Differentiation among Populations
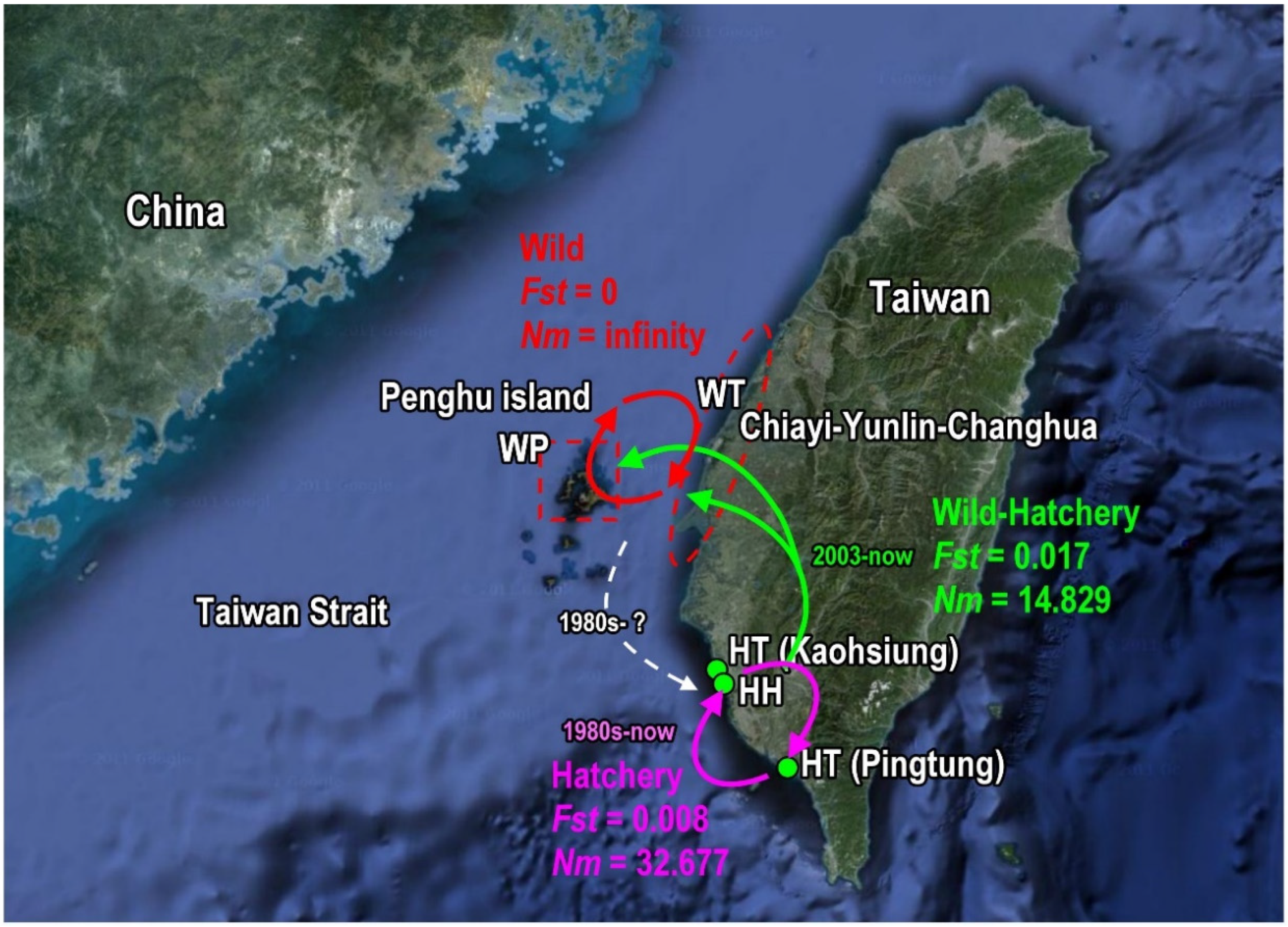
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blankenship, H.L.; Leber, K.M. A responsible approach to marine stock enhancement. Am. Fish. Soc. Symp. 1995, 15, 167–175. [Google Scholar]
- Bell, J.D.; Leber, K.M.; Blankenship, H.L.; Loneragan, N.R.; Masuda, R. A new era for restocking, stock enhancement and sea ranching of coastal fisheries resources. Rev. Fish. Sci. 2008, 16, 1–9. [Google Scholar] [CrossRef]
- Araki, H.; Schmid, C. Is hatchery stocking a help or harm? Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture 2010, 308, S2–S11. [Google Scholar] [CrossRef]
- Lorenzen, K.; Leber, K.M.; Blankenship, H.L. Responsible approach to marine stock enhancement: An update. Rev. Fish. Sci. 2010, 18, 189–210. [Google Scholar] [CrossRef]
- Liao, I.C. Status, Problems and prospects of stock enhancement in Taiwan. Hydrobiologia 1997, 352, 167–180. [Google Scholar] [CrossRef]
- Liao, I.C.; Su, M.S.; Leano, E.M. Status of research in stock enhancement and sea ranching. Rev. Fish Biol. Fisher. 2003, 13, 151–163. [Google Scholar] [CrossRef]
- Laikre, L.; Schwartz, M.K.; Waples, R.S.; Ryman, N.; Grp, G.W. Compromising genetic diversity in the wild: Unmonitored large-scale release of plants and animals. Trends Ecol. Evol. 2010, 25, 520–529. [Google Scholar] [CrossRef] [Green Version]
- Kitada, S.; Shishidou, H.; Sugaya, T.; Kitakado, T.; Hamasaki, K.; Kishino, H. Genetic effects of long-term stock enhancement programs. Aquaculture 2009, 290, 69–79. [Google Scholar] [CrossRef]
- Ward, R.D. The importance of identifying spatial population structure in restocking and stock enhancement programmes. Fish. Res. 2006, 80, 9–18. [Google Scholar] [CrossRef]
- Satake, A.; Araki, H. Stocking of captive-bred fish can cause long-term population decline and gene pool replacement: Predictions from a population dynamics model incorporating density-dependent mortality. Theor. Ecol. 2012, 5, 283–296. [Google Scholar] [CrossRef]
- Baskett, M.L.; Burgess, S.C.; Waples, R.S. Assessing strategies to minimize unintended fitness consequences of aquaculture on wild populations. Evol. Appl. 2013, 6, 1090–1108. [Google Scholar] [CrossRef] [PubMed]
- Milot, E.; Perrier, C.; Papillon, L.; Dodson, J.J.; Bernatchez, L. Reduced fitness of Atlantic salmon released in the wild after one generation of captive breeding. Evol. Appl. 2013, 6, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Naish, K.A.; Seamons, T.R.; Dauer, M.B.; Hauser, L.; Quinn, T.P. Relationship between effective population size, inbreeding and adult fitness-related traits in a steelhead (Oncorhynchus mykiss) population released in the wild. Mol. Ecol. 2013, 22, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Boscari, E.; Congiu, L. The need for genetic support in restocking activities and ex situ conservation programmes: The case of the Adriatic sturgeon (Acipenser naccarii Bonaparte, 1836) in the Ticino River Park. J. Appl. Ichthyol. 2014, 30, 1416–1422. [Google Scholar] [CrossRef]
- An, H.S.; Nam, M.M.; Myeong, J.I.; An, C.M. Genetic diversity and differentiation of the Korean starry flounder (Platichthys stellatus) between and within cultured stocks and wild populations inferred from microsatellite DNA analysis. Mol. Biol. Rep. 2014, 41, 7281–7292. [Google Scholar] [CrossRef]
- Glover, K.A.; Dahle, G.; Jørstad, K.E. Genetic identification of farmed and wild Atlantic cod, Gadus morhua, in coastal Norway. ICES J. Mar. Sci. 2011, 68, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Wringe, B.F.; Jeffery, N.W.; Stanley, R.R.E.; Hamilton, L.C.; Anderson, E.C.; Fleming, I.A.; Bradbury, I.R. Extensive hybridization following a large escape of domesticated Atlantic salmon in the Northwest Atlantic. Commun. Biol. 2018, 1, 108. [Google Scholar] [CrossRef] [PubMed]
- Šegvić-Bubić, T.; Grubišić, L.; Trumbić, Ž.; Stanić, R.; Ljubković, J.; Maršić-Lučić, J.; Katavić, I. Genetic characterization of wild and farmed European seabass in the Adriatic sea: Assessment of farmed escapees using a Bayesian approach. ICES J. Mar. Sci. 2016, 74, 369–378. [Google Scholar] [CrossRef]
- Sawayama, E.; Nakao, H.; Kobayashi, W.; Minami, T.; Takagi, M. Identification and quantification of farmed red sea bream escapees from a large aquaculture area in Japan using microsatellite DNA markers. Aquat. Living Resour. 2019, 32, 26. [Google Scholar] [CrossRef]
- Gonzalez, E.B.; Umino, T. Fine-scale genetic structure derived from stocking black sea bream, Acanthopagrus schlegelii (Bleeker, 1854), in Hiroshima Bay, Japan. J. Appl. Ichthyol. 2009, 25, 407–410. [Google Scholar] [CrossRef]
- Hunt, T.L.; Jones, P. Informing the great fish stocking debate: An Australian case study. Rev. Fish. Sci. Aquac. 2018, 26, 275–308. [Google Scholar] [CrossRef]
- Hesp, S.A.; Potter, I.C. Reproductive biology of Rhabdosargus sarba (Sparidae) in Western Australian waters, in which it is a rudimentary hermaphrodite. J. Mar. Biol. Assoc. UK 2003, 83, 1333–1346. [Google Scholar] [CrossRef] [Green Version]
- De Mitcheson, Y.S.; Liu, M. Functional hermaphroditism in teleosts. Fish Fish. 2008, 9, 1–43. [Google Scholar] [CrossRef]
- Bauchot, M.L.; Skelton, P.H. Sparidae. In Check-List of The Freshwater fishes of Africa (CLOFFA); Daget, J., Gosse, J.-P., van den Audenaerde, D.F.E., Eds.; Tervuren and ORSTOM: Paris, France, 1986; pp. 331–332. [Google Scholar]
- Frost, L.A.; Evans, B.S.; Jerry, D.R. Loss of genetic diversity due to hatchery culture practices in barramundi (Lates calcarifer). Aquaculture 2006, 261, 1056–1064. [Google Scholar] [CrossRef]
- Lind, C.E.; Evans, B.S.; Knauer, J.; Taylor, J.J.U.; Jerry, D.R. Decreased genetic diversity and a reduced effective population size in cultured silver-lipped pearl oysters (Pinctada maxima). Aquaculture 2009, 286, 12–19. [Google Scholar] [CrossRef]
- Loukovitis, D.; Sarropoulou, E.; Vogiatzi, E.; Tsigenopopulos, C.S.; Kotoulas, G.; Magoulas, A.; Chatziplis, D. Genetic variation in farmed populations of the gilthead sea bream Sparus aurata in Greece using microsatellite DNA markers. Aquac. Res. 2012, 43, 239–246. [Google Scholar] [CrossRef]
- Wang, L.; Shi, X.F.; Su, Y.Q.; Meng, Z.N.; Lin, H.R. Loss of genetic diversity in the cultured stocks of the large yellow croaker, Larimichthys crocea, revealed by microsatellites. Int. J. Mol. Sci. 2012, 13, 5584–5597. [Google Scholar] [CrossRef] [Green Version]
- Segovia-Viadero, M.; Serrao, E.A.; Canteras-Jordana, J.C.; Gonzalez-Wanguemert, M. Do hatchery-reared sea urchins pose a threat to genetic diversity in wild populations? Heredity 2016, 116, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Porta, J.; Porta, J.M.; Canavate, P.; Martinez-Rodriguez, G.; Alvarez, M.C. Substantial loss of genetic variation in a single generation of Senegalese sole (Solea senegalensis) culture: Implications in the domestication process. J. Fish Biol. 2007, 71, 223–234. [Google Scholar] [CrossRef]
- Holmer, M. Environmental issues of fish farming in offshore waters: Perspectives, concerns and research needs. Aquac. Environ. Interact. 2010, 1, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Dimitriou, E.; Katselis, G.; Moutopoulos, D.K.; Akovitiotis, C.; Koutsikopoulos, C. Possible influence of reared gilthead sea bream (Sparus aurata, L.) on wild stocks in the area of the Messolonghi lagoon (Ionian Sea, Greece). Aquac. Res. 2007, 38, 398–408. [Google Scholar] [CrossRef]
- Somarakis, S.; Pavlidis, M.; Saapoglou, C.; Tsigenopoulos, C.S.; Dempster, T. Evidence for ’escape through spawning’ in large gilthead sea bream Sparus aurata reared in commercial sea-cages. Aquacult. Environ. Interact. 2013, 3, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.M.; Fraser, D.J.; Meier, K.; Mensberg, K.-L.D. Sixty years of anthropogenic pressure: A spatio-temporal genetic analysis of brown trout populations subject to stocking and population declines. Mol. Ecol. 2009, 18, 2549–2562. [Google Scholar] [CrossRef] [PubMed]
- Kitada, S. Economic, ecological and genetic impacts of marine stock enhancement and sea ranching: A systematic review. Fish Fish. 2018, 19, 511–532. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Takagi, M.; Taniguchi, N.; Cook, D.; Doyle, R.W. Isolation and characterization of microsatellite loci from red sea bream Pagrus major and detection in closely related species. Fish. Sci. 1997, 63, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Rice, W.R. Analysing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Borrell, Y.J.; Pinera, J.A.; Sanchez Prado, J.A.; Blanco, G. Mitochondrial DNA and microsatellite genetic differentiation in the European anchovy Engraulis encrasicolus L. ICES J. Mar. Sci. 2012, 69, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. C LUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, R.K.; Ramasamy, S.; Bindroo, B.B.; Naik, V.G. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. SpringerPlus 2014, 3, 431. [Google Scholar] [CrossRef] [Green Version]
- Piry, S.; Alapetite, A.; Cornuet, J.-M.; Paetkau, D.; Baudouin, L.; Estoup, A. GeneClass2: A software for genetic assignment and first-generation migrant detection. J. Hered. 2004, 95, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Dupanloup, I.; Schneider, S.; Excoffier, L.A. Simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
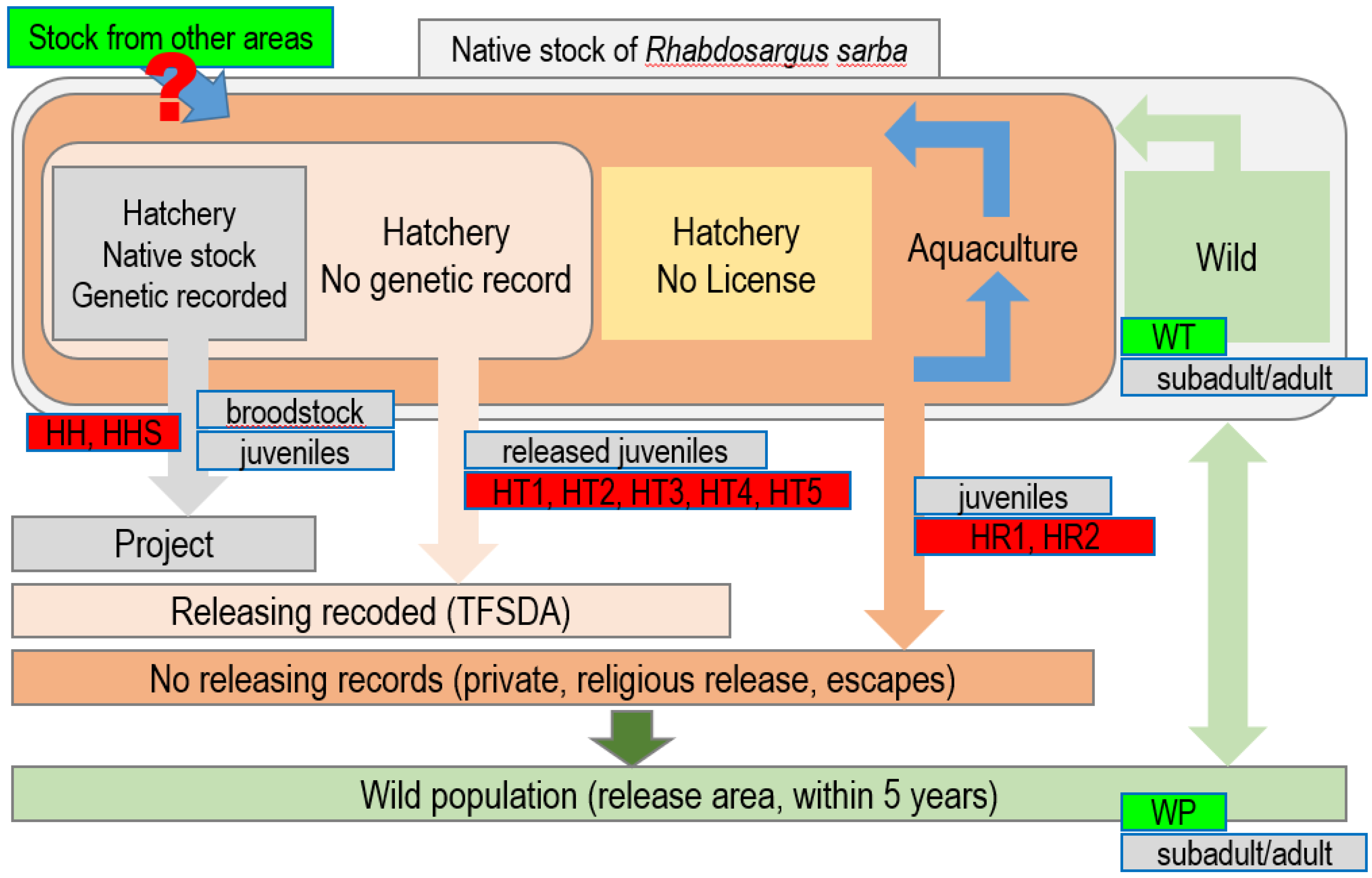{kind=link}
| Code | N | Year | Sources | Fish Types | Structure Analysis * |
|---|---|---|---|---|---|
| HT | |||||
| HT1 | 31 | 2015 | TFSDA releasing from Pingtung hatchery | Released juveniles | All samples are hatchery |
| HT2 | 153 | 2015 | All samples are hatchery | ||
| HT3 | 91 | 2015 | TFSDA releasing from Kaohsiung hatchery | All samples are hatchery | |
| HT4 | 98 | 2015 | All samples are hatchery | ||
| HT5 | 65 | 2016 | All samples are hatchery | ||
| HH | All samples are hatchery | ||||
| HH | 33 | 2015 | Juveniles from Kaohsiung hatchery broodstock | Juveniles | All samples are hatchery |
| HHS | 36 | 2015 | Broodstock of Kaohsiung hatchery | Broodstock | All samples are hatchery |
| HR | All samples are hatchery | ||||
| HR1 | 122 | 2016 | Religious releasing from Unknown hatchery | Released juveniles | All samples are hatchery |
| HR2 | 72 | 2016 | All samples are hatchery | ||
| WP | 421 | 2015 | Wild; Penghu island (releasing area) | Subadult/adult | Wild: 393 (93.35%) Hatchery: 28 (6.65%) + |
| WT | 38 | 2015 | Wild; Chiayi–Yunlin–Changhua | Subadult/adult | All samples are wild cluster |
| Population (N) | MSTN1 | MSTN2 | MSTN3 | Pma4 | Cm300 | Average | |
|---|---|---|---|---|---|---|---|
| HT1 (31) | Na | 3 | 2 | 4 | 10 | 4 | 4.6 |
| Ne | 1.805 | 1.875 | 2.051 | 5.209 | 2.580 | 2.704 | |
| Ho | 0.548 | 0.548 | 0.516 | 0.903 | 1.000 | 0.703 | |
| He | 0.446 | 0.467 | 0.512 | 0.808 | 0.612 | 0.569 | |
| FIS | −0.230 NS | −0.175 NS | −0.007 NS | −0.118 NS | −0.633 *** | −0.233 | |
| HT2 (153) | Na | 4 | 2 | 3 | 13 | 7 | 5.8 |
| Ne | 2.230 | 1.906 | 2.026 | 5.128 | 3.478 | 2.954 | |
| Ho | 0.660 | 0.516 | 0.601 | 0.797 | 0.987 | 0.712 | |
| He | 0.552 | 0.475 | 0.506 | 0.805 | 0.712 | 0.61 | |
| FIS | −0.197 ** | −0.086 NS | −0.187 NS | 0.009 NS | −0.385 *** | −0.169 | |
| HT3 (91) | Na | 3 | 3 | 5 | 12 | 6 | 5.8 |
| Ne | 2.138 | 1.650 | 1.914 | 4.051 | 3.529 | 2.656 | |
| Ho | 0.604 | 0.356 | 0.473 | 0.778 | 0.956 | 0.633 | |
| He | 0.532 | 0.394 | 0.478 | 0.753 | 0.717 | 0.575 | |
| FIS | −0.136 NS | 0.098 NS | 0.010 NS | −0.033 NS | −0.333 *** | −0.079 | |
| HT4 (98) | Na | 3 | 3 | 3 | 12 | 6 | 5.4 |
| Ne | 2.284 | 1.927 | 1.930 | 5.822 | 3.4 | 3.072 | |
| Ho | 0.694 | 0.541 | 0.469 | 0.847 | 0.980 | 0.706 | |
| He | 0.562 | 0.481 | 0.482 | 0.828 | 0.706 | 0.612 | |
| FIS | −0.234 * | −0.124 NS | 0.026 NS | −0.023 *** | −0.388 *** | −0.149 | |
| HT5 (65) | Na | 3 | 4 | 3 | 11 | 7 | 5.6 |
| Ne | 2.399 | 1.862 | 1.793 | 5.201 | 3.435 | 2.938 | |
| Ho | 0.723 | 0.477 | 0.462 | 0.828 | 0.985 | 0.695 | |
| He | 0.583 | 0.463 | 0.442 | 0.808 | 0.709 | 0.601 | |
| FIS | −0.240 * | −0.030 NS | −0.044 NS | −0.025 NS | −0.389 *** | −0.146 | |
| HH (33) | Na | 3 | 2 | 3 | 10 | 6 | 4.8 |
| Ne | 2.295 | 1.695 | 1.957 | 5.018 | 3.285 | 2.85 | |
| Ho | 0.515 | 0.333 | 0.788 | 0.909 | 1.000 | 0.709 | |
| He | 0.564 | 0.410 | 0.489 | 0.801 | 0.696 | 0.592 | |
| FIS | 0.087 NS | 0.187 NS | −0.611 ** | −0.135 NS | −0.438 *** | −0.182 | |
| HHS (36) | Na | 3 | 3 | 3 | 11 | 7 | 5.4 |
| Ne | 1.889 | 1.944 | 2.008 | 6.056 | 3.590 | 3.098 | |
| Ho | 0.528 | 0.444 | 0.472 | 0.833 | 0.972 | 0.65 | |
| He | 0.471 | 0.486 | 0.502 | 0.835 | 0.721 | 0.603 | |
| FIS | −0.121 NS | 0.085 NS | 0.059 NS | 0.002 NS | −0.348 * | −0.065 | |
| HR1 (122) | Na | 3 | 2 | 4 | 13 | 7 | 5.8 |
| Ne | 2.068 | 1.871 | 1.969 | 5.530 | 3.268 | 2.941 | |
| Ho | 0.639 | 0.475 | 0.516 | 0.820 | 0.893 | 0.669 | |
| He | 0.516 | 0.466 | 0.492 | 0.819 | 0.694 | 0.597 | |
| FIS | −0.238 ** | −0.021 NS | −0.049 *** | −0.001 *** | −0.287 *** | −0.119 | |
| HR2 (72) | Na | 4 | 2 | 2 | 12 | 7 | 5.4 |
| Ne | 2.499 | 1.714 | 1.870 | 3.370 | 3.362 | 2.563 | |
| Ho | 0.708 | 0.423 | 0.542 | 0.676 | 0.958 | 0.661 | |
| He | 0.600 | 0.417 | 0.465 | 0.703 | 0.703 | 0.577 | |
| FIS | −0.181 NS | −0.014 NS | −0.164 NS | 0.039 *** | −0.364 *** | −0.137 | |
| WP (421) | Na | 6 | 8 | 9 | 16 + | 11 | 10 |
| Ne | 2.592 | 2.022 | 2.081 | 3.567 | 3.393 | 2.731 | |
| Ho | 0.660 | 0.480 | 0.558 | 0.651 | 0.936 | 0.657 | |
| He | 0.614 | 0.506 | 0.520 | 0.720 | 0.705 | 0.613 | |
| FIS | −0.075 *** | 0.051 *** | −0.074 *** | 0.096 *** | −0.327 *** | −0.066 | |
| WT (38) | Na | 5 | 4 | 7 | 10 | 9 | 7 |
| Ne | 2.882 | 2.131 | 2.429 | 3.402 | 4.079 | 2.985 | |
| Ho | 0.711 | 0.526 | 0.447 | 0.632 | 0.895 | 0.642 | |
| He | 0.653 | 0.531 | 0.588 | 0.706 | 0.755 | 0.647 | |
| FIS | −0.088 ** | 0.008 *** | 0.240 *** | 0.105 * | −0.185 *** | 0.016 | |
| All (1160) | Na | 8 | 9 | 12 | 16 | 12 | 11.4 |
| Ne | 2.374 | 1.928 | 2.027 | 4.656 | 3.523 | 2.901 | |
| HT1 | HT2 | HT3 | HT4 | HT5 | HH | HHS | HR1 | HR2 | WP | WT | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| HT1 | - | 0.055 NS | 0.037 NS | 0.148 NS | 0.015 NS | 0.146 NS | 0.156 NS | 0.140 NS | 0.027 NS | 0.000 * | 0.012 NS |
| HT2 | 0.010 | - | 0.000 * | 0.017 NS | 0.003 NS | 0.063 NS | 0.125 NS | 0.003 NS | 0.002 NS | 0.000 * | 0.011 NS |
| HT3 | 0.012 | 0.013 | - | 0.035 NS | 0.283 NS | 0.297 NS | 0.179 NS | 0.004 NS | 0.346 NS | 0.000 * | 0.000 * |
| HT4 | 0.005 | 0.007 | 0.006 | - | 0.430 NS | 0.227 NS | 0.392 NS | 0.035 NS | 0.037 NS | 0.000 * | 0.019 NS |
| HT5 | 0.016 | 0.012 | 0.001 | 0.000 | - | 0.171 NS | 0.363 NS | 0.018 NS | 0.123 NS | 0.000 * | 0.018 NS |
| HH | 0.008 | 0.007 | 0.002 | 0.003 | 0.005 | - | 0.234 NS | 0.259 NS | 0.359 NS | 0.004 NS | 0.058 NS |
| HHS | 0.007 | 0.005 | 0.004 | 0.000 | 0.000 | 0.004 | - | 0.209 NS | 0.050 NS | 0.004 NS | 0.041 NS |
| HR1 | 0.006 | 0.010 | 0.011 | 0.006 | 0.009 | 0.002 | 0.003 | - | 0.005 NS | 0.000 * | 0.029 NS |
| HR2 | 0.015 | 0.015 | 0.001 | 0.007 | 0.004 | 0.001 | 0.010 | 0.013 | - | 0.000 * | 0.048 NS |
| WP | 0.026 | 0.023 | 0.027 | 0.016 | 0.019 | 0.016 | 0.020 | 0.015 | 0.017 | - | 0.426 NS |
| WT | 0.024 | 0.014 | 0.021 | 0.012 | 0.015 | 0.011 | 0.014 | 0.012 | 0.010 | 0.000 | - |
| Source | df | Sum of Squares | Mean Squares | Variance | % Total |
|---|---|---|---|---|---|
| Eleven populations | |||||
| Among sampling localities | 10 | 55.329 | 5.533 | 0.022 | 1% |
| Among individuals | 1149 | 1559.888 | 1.358 | 0.000 | 0% |
| Within individuals | 1160 | 1947.500 | 1.679 | 1.679 | 99% |
| Total | 2319 | 3562.716 | 1.701 | 100% | |
| Average FST value = 0.014 (p = 0 < 0.001); Nm = 17.209 | |||||
| Hatchery (9) and wild (2) | |||||
| Among sampling localities | 1 | 29.874 | 29.874 | 0.026 | 2% |
| Among individuals | 1158 | 1585.342 | 1.369 | 0.000 | 0% |
| Within individuals | 1160 | 1947.500 | 1.679 | 1.679 | 98% |
| Total | 2319 | 3562.716 | 1.705 | 100% | |
| FST value = 0.017 (p = 0 < 0.001), Nm = 14.829 | |||||
| Nine hatchery populations | |||||
| Among sampling localities | 8 | 24.252 | 3.031 | 0.011 | 1% |
| Among individuals | 692 | 899.459 | 1.300 | 0.000 | 0% |
| Within individuals | 701 | 1195.000 | 1.705 | 1.705 | 99% |
| Total | 1401 | 2118.710 | 1.716 | 100% | |
| Region Groupings | ΦCT | % Variance among Groups | |
|---|---|---|---|
| K = 2 | (HT1, HT2, HT3, HT4, HT5, HH, HHS, HR1, HR2); (WP, WT) | 0.077 * | 7.65 |
| K = 3 | (HT1, HT2, HT3, HT4, HT5, HH, HHS); (HR1, HR2); (WP, WT) | 0.075 * | 7.53 |
| K = 4 | (HT1, HT2, HT3, HT4, HT5); (HH, HHS); (HR1, HR2); (WP, WT) | 0.075 ** | 7.47 |
| K = 5 | (HT1, HT2, HT3, HT4, HT5); (HH, HHS); (HR1, HR2); (WP); (WT) | 0.073 ** | 7.30 |
| K = 6 | (HT1, HT2, HT3, HT4, HT5); (HH, HHS); (HR1); (HR2); (WP); (WT) | 0.071 ** | 7.06 |
| K = 7 | (HT1, HT2); (HT3, HT4, HT5); (HH, HHS); (HR1); (HR2); (WP); (WT) | 0.068 ** | 6.86 |
| Locus | Primer Sequences (5′–3’) | Repeat Motif | Ta (°C) | Size Range (bp) | Reference |
|---|---|---|---|---|---|
| MSTN1 | F:CACGCCATCACGGAGACGATTATG | (T)n | 59 | 248–288 | This study |
| R:CAATCCCTGACACGAATCCCTGAC | |||||
| MSTN2 | F:GCAGGCGGTTAAACATTCCTGC | (CA)n | 58 | 340–362 | This study |
| R:GGTTGGTTAACCACCGCCGTCTC | |||||
| MSTN3 | F:CTGCTTTCACATCCGGCACAGC | (CA)n | 60 | 179–207 | This study |
| R:GAACACAGAGACGACGAAGGACGAG | |||||
| Pma4 | F:GCCACCTACTGTTTCCTCAACTTCTG | (CA)n | 60 | 150–180 | [32] |
| R:GTGATTACAGTCGGGTTTGGCTG | |||||
| Cm300 | F:GAAAGATGGGTTGTGAGGGT | (CT)n | 54 | 117–139 | AB703235 |
| R:CCATCTGAACGTCTGAGCG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, T.-H.; Huang, C.-W.; Lee, H.-T.; Kuo, Y.-H.; Liu, K.-M.; Lin, C.-H.; Gong, H.-Y. Population Genetic Analysis for Stock Enhancement of Silver Sea Bream (Rhabdosargus sarba) in Taiwan. Fishes 2020, 5, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5020019
Hsu T-H, Huang C-W, Lee H-T, Kuo Y-H, Liu K-M, Lin C-H, Gong H-Y. Population Genetic Analysis for Stock Enhancement of Silver Sea Bream (Rhabdosargus sarba) in Taiwan. Fishes. 2020; 5(2):19. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5020019
Chicago/Turabian StyleHsu, Te-Hua, Chang-Wen Huang, Hung-Tai Lee, Yi-Hsuan Kuo, Kwang-Ming Liu, Cheng-Hui Lin, and Hong-Yi Gong. 2020. "Population Genetic Analysis for Stock Enhancement of Silver Sea Bream (Rhabdosargus sarba) in Taiwan" Fishes 5, no. 2: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5020019