4.1. Body Fatty Acid Composition and Apparent Retention
It has been well established in fish that the tissue fatty acid composition generally reflects dietary fatty acid composition [
30,
31,
32]. Based on the regression analysis, the higher the inclusion of AM in the diets, the higher the accumulation of DHA in the whole body of the fish. Our findings also show an increase in the whole-body n-3 fatty acid content, followed by an increase in the dietary inclusion of AM, a source of DHA. Several authors cited an accumulation of DHA in the body and muscle composition of fish, especially at a low temperature [
15,
18,
33,
34]. The DHA is considered a fatty acid of high biological value, with a close link between DHA and the composition of structural phospholipids in cells [
17]. Thus, DHA has a different catabolism from other fatty acids, being more preserved in cell phospholipids and less catabolized to produce energy [
35]. Due to its high flexibility provided by its chemical arrangement, DHA maintains a cell membrane bilayer with the balance between fluidity and rigidity needed to accommodate rapid conformational changes in cell membrane proteins. This structure brings fluidity to the membrane at low temperatures [
17]. In addition to preserving DHA, which can be modulated directly by the diet, fish also have desaturase and elongate enzymes that are responsible for desaturating LOA to ARA, LNA to EPA, and EPA to DHA. Indeed, Nile tilapia have been reported to express the genes for Δ-4 and Δ-6 desaturases, thus enabling the conversion of EPA into DHA by two different pathways, which are mediated by both desaturases [
17,
36].
On the other hand, during the β-oxidation of fatty acids, fish preferentially use SFA and MUFA, with shorter chain lengths and numbers of unsaturation, as energy sources [
37,
38]. This observation may explain the linear reduction in PAL, SFA, and MUFA in fish fed 10AM, 20AM, and 40AM, which in turn, could be preferentially degraded for energy production in order to preserve the fatty acids of greatest biological value, such as PUFAs. The decrease in ARA, n-6 PUFA, and n-6 LC-PUFA groups can be explained by the competition of desaturase and elongase enzymes, which compete for the substrates of n-3 and n-6 series fatty acids and have a greater affinity for n-3 PUFA substrates [
39]. Thus, the reduction in bioconversion from n-6 PUFA to n-6 LC-PUFA may have occurred because of the inhibition of the enzymatic activity of desaturases and elongases in the presence of higher levels of DHA in the diets [
32,
39].
In the present study, the higher concentration of DHA in the bodies of fish fed the CLO diet than in fish fed the diet containing 10AM probably occurred due to the bioconversion of its precursors (LNA, EPA, and DPA) which were detected at higher levels only in fish fed the CLO diet. Through the study of the fatty acid balance, it was possible to understand that for Nile tilapia, the bioconversion of LNA to DHA is efficient, and the biosynthesis of DHA from EPA can be more direct or faster than the production of EPA from LNA [
32]. Our study corroborates such findings, where the CLO diet contained more LNA and EPA in its composition, favoring their bioconversion to DHA.
The ARR of DHA and n-3 PUFA was influenced by the levels of dietary inclusion; the higher the dietary concentration of these fatty acids, the lower the retention. Other authors have also reported a similar pattern, where the higher the concentration of a fatty acid in the diet, the lower its relative deposition [
18,
39]. In our study, as also reported by Brignol et al. [
22] at 28 °C, the ARR of α-LNA increased with the inclusion of AM. Increasing the dietary concentration of DHA following the inclusion of AM possibly prevented the elongation and desaturation of α-LNA for the production of LC-PUFA, thus preserving and retaining α-LNA in the membranes. In addition, diets containing higher inclusions of AM contained less α-LNA content. Consequently, the lower the presence of an essential fatty acid in the diet, the higher its body retention. However, contrary to the report by Brignol et al. [
22], at 28 °C, where there was an increase in n-6 PUFA body retention with the inclusion of AM, we did not observe such a response in our study, at 22 °C. Here, PUFA n-6 retention decreased significantly with the inclusion of AM, following the body composition trend that also decreased, probably due to the competition of desaturases and elongases with the n-3 substrate and/or due to β-oxidation of the PUFA n-6, in addition to the greater conservation of the n-3 fatty acids to maintain membrane functionality at low temperatures.
When comparing the two sources of DHA, fish fed CLO showed higher retention of the following fatty acids: LOA, α-LNA, DHA, n-3 PUFA, and n-6 PUFA at 28 °C [
22]. However, in our study, at 22 °C, the retention of α-LNA and n-6 PUFA was higher in fish fed the AM-supplemented diet. The higher retention of α-LNA can be explained by the low content of this fatty acid in the diet supplemented with AM, which may also have led to an increased retention of n-6 PUFA, as there was little n-3 substrate for inhibition of the desaturases and elongases in the n-6 pathway. Despite having similar amounts of DHA, the higher DHA, total PUFA, and n-3 PUFA retention in fish fed the CLO diet was probably due to the higher synthesis of DHA from precursor fatty acids, detected only in the CLO diet [
17,
22].
4.2. Fatty Acid Composition in the Hepatopancreas
The greater the dietary inclusion of AM, the lower the amount of total lipid in the liver, in addition to EPA, ARA, ADA, PUFA n-6, and LC-PUFA n-6. However, the increasing inclusions of AM increased the composition of DHA, PUFA, LC-PUFA n-3, and the n-3:n-6 ratio. The fatty acid composition of hepatopancreas largely reflected that of the diet, which is consistent with studies in Atlantic salmon [
40].
The hepatopancreas is the main organ that regulates lipid metabolism, including both the synthesis and degradation of fatty acids, where several regulating enzymes show varied affinities for the different fatty acids available in that organ [
37,
41]. In addition, the hepatopancreas functions as an important energy reservoir, often in the form of triacylglycerols [
21]. The high intake of PUFAs (mainly EPA and DHA) prevents the accumulation of lipids by inducing lipid oxidation, [
41,
42], inhibiting lipogenic metabolism, and stimulating the synthesis of lipoproteins [
43,
44,
45,
46].
The significant increase in DHA in the hepatopancreas, about 105% between 0AM- and 40AM-fed fish, suggests the important role of this fatty acid in cell membrane function at a low temperature. Additionally, the decrease in EPA content in the hepatopancreas may be linked to the bioconversion of EPA to DHA [
17,
32]. In addition, the content of n-6 PUFA decreased, suggesting the preferential route of activity of the desaturase and elongase enzymes by the n-3 series fatty acids. Indeed, Chen et al. [
32] reported that increasing dietary inclusion of LNA in Nile tilapia raised at the optimal temperature could block or at least slow down n-6 LC-PUFA biosynthesis from LOA. Therefore, there is competition for accessing the Δ-6 desaturase and elongase between substrates of the n-3 and n-6 series. Although we have not evaluated such enzymes in our study, this substrate competition is well known for tilapia [
32].
4.3. Morphology and Histological Changes in the Intestine and Hepatopancreas
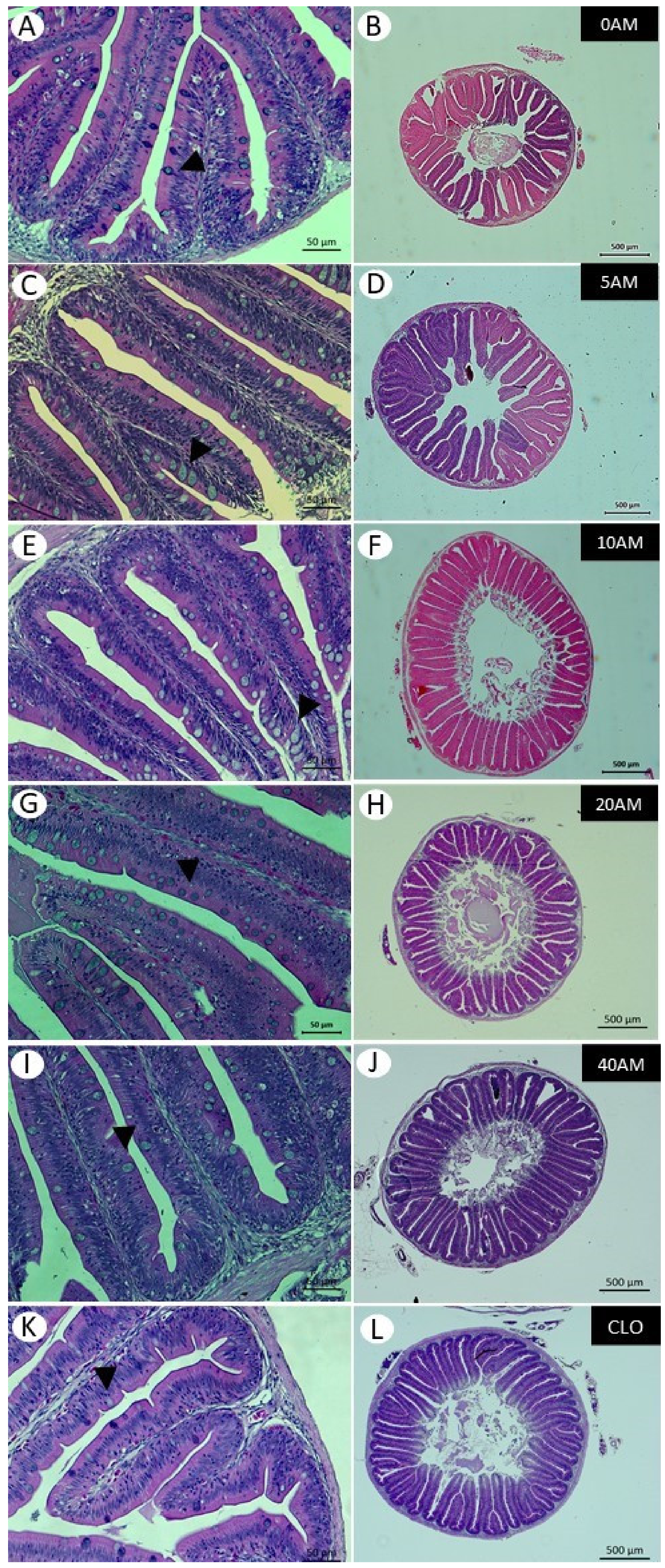
Different levels of dietary supplementation with AM affected the histology of the hepatopancreas and intestine. Fish fed the highest levels of AM showed a significant increase in the number of intestinal folds. The increase in the absorption area, caused by the increase in the number of folds, could improve the digestive and absorptive processes, suggesting a more efficient use of nutrients [
47]. Our results regarding the increase in the number of villi in fish fed the 10AM diet corroborate the findings of Nobrega et al. [
15], where fish fed the 10AM diet reached the highest weight gain. Such high growth may be associated with an increase in the nutrient absorption area. The influence of diet-derived substances on intestinal epithelial function, including barrier integrity, is likely to be important [
48].
A recent review on mammals [
48] reported that n-3 LC-PUFA, especially DHA, contributes to maintaining the integrity of the intestinal epithelial barrier by exerting anti-inflammatory effects and accelerating recovery from intestinal inflammation. However, few studies have addressed the relationship between dietary fatty acids, especially LC-PUFA n-3, and intestinal health in fish. Dietary supplementation with
Schizochytrium sp. (a DHA-producing marine heterotrophic microorganism from the same family as
Aurantiochytrium sp.) in juvenile mirror carp (
Cyprinus carpio var. specularis) resulted in a higher intestinal fold height in fish fed 30 and 60 g kg
−1 supplementation if compared to fish fed the non-supplemented diet [
49]. The dietary replacement of fish oil and fish meal by heterotrophic microorganisms results in a good response to intestinal integrity, since such microorganisms also contain additional bioactive cell wall compounds such as β-glucans, β-carotenes, flavonoids, nucleotides, and water-soluble peptides [
50,
51,
52] which can affect nutrient availability and growth performance, but also enhance the well-being of fish by improving gut health and thus nutrient assimilation and immune competence [
52].
The rising levels of AM also resulted in a significant increase in the number of goblet cells in fish fed diets 0AM to 20AM. An increase in the number of goblet cells was also reported in the intestine of Atlantic salmon, reared at an optimal temperature for the species (10.2 °C), with an increase in dietary inclusion of 6 to 15 g kg
−1 Schizochytrium sp. meal [
52]. Goblet cells produce mucus, which plays an important role in immunity. Besides serving as a mechanical barrier, making it difficult for pathogenic bacteria to adhere, mucus contains several components of the innate immune response, such as lysozymes, immunoglobulins, complement system proteins, lectins, and several other antimicrobial components [
53]. However, in fish fed the highest AM level (40AM), there was a reduction in the number of goblet cells. A previous study evaluated different dietary sources of n-3 LC-PUFA (fish oil, EPA-enriched oil, and DHA-enriched oil) for the carnivorous marine fish pompano,
Trachinotus ovatus, raised at an optimal temperature and reported that high dietary EPA or DHA levels caused a depressed expression of Muc13 mRNA in the intestines [
54]. Mucins are the major constituent of the mucous layer and are produced by goblet cells [
55]. Therefore, more research needs to be carried out to assess how dietary DHA interacts with mucus production in the intestine. Furthermore, the decrease in goblet cells does not seem to be related to inflammation in the intestine, since treatments with higher AM inclusions showed a greater number of villi, showing its positive effect on intestinal morphometry.
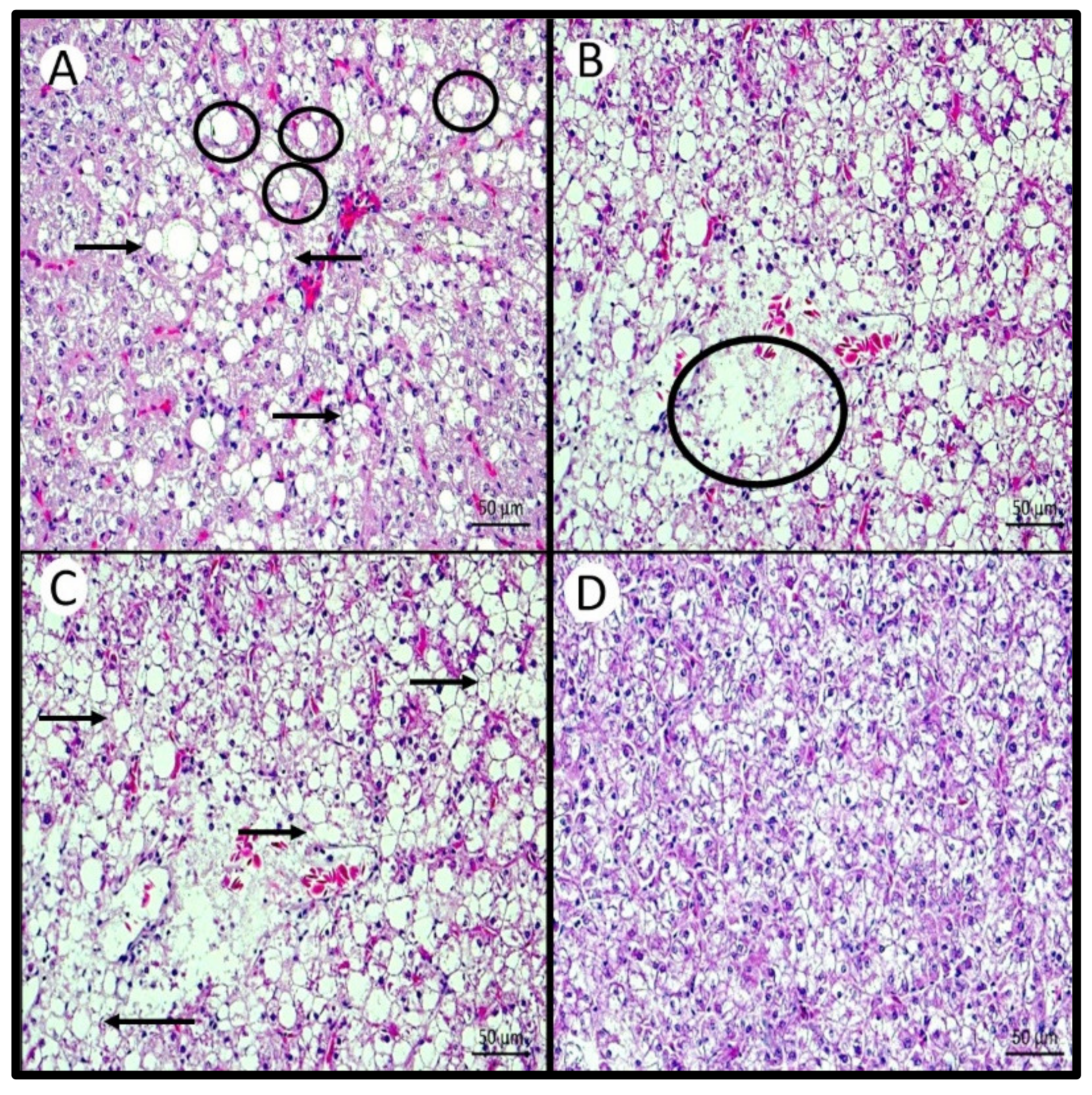
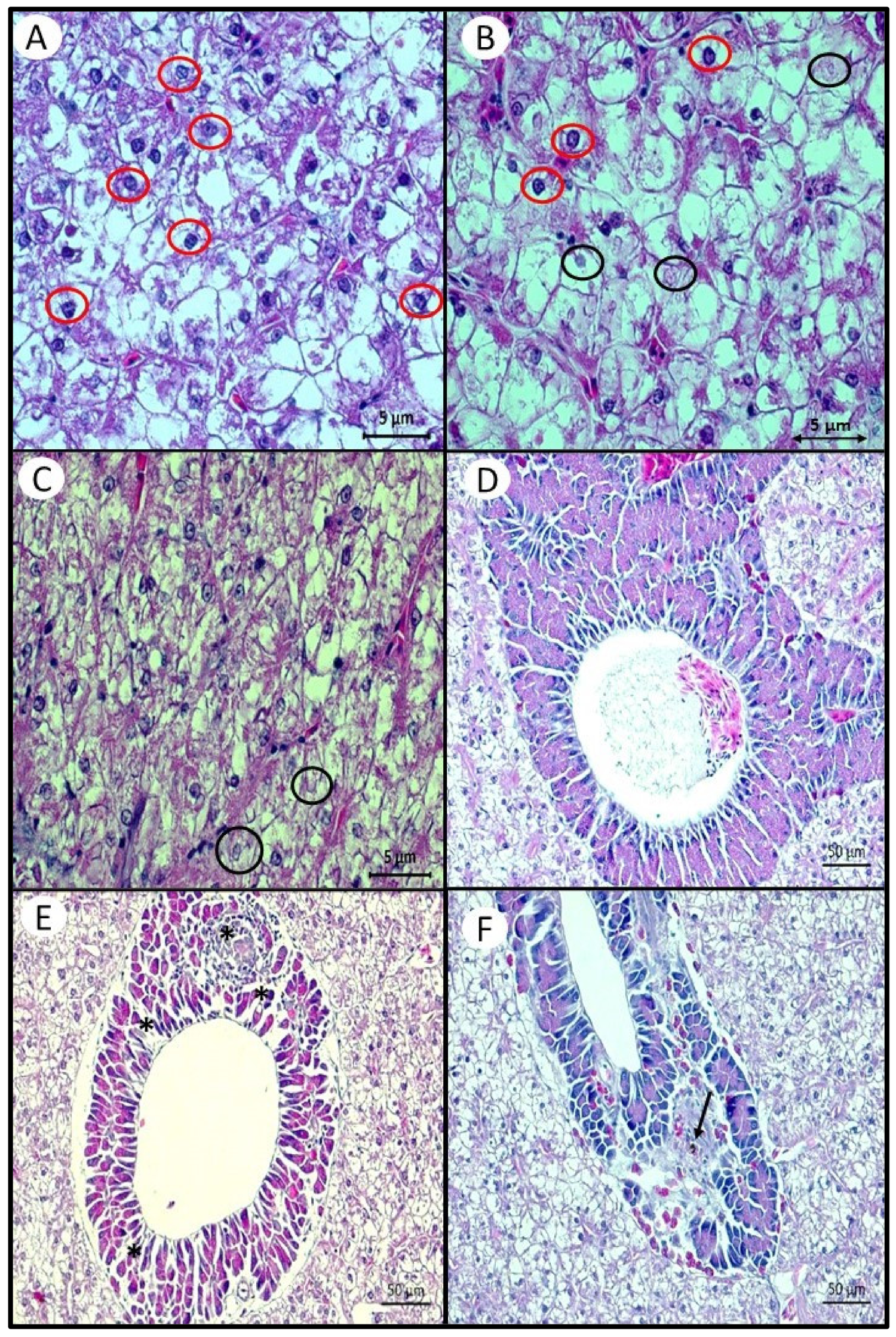
Histological analysis of the hepatopancreas showed a similar pattern to that observed in the total lipid content of this organ. Likewise, the intensity of macrosteatosis, microsteatosis, necrosis, and loss of the nucleus of hepatocytes were lower in the hepatopancreas of fish fed the highest inclusions of AM. Therefore, the highest intensity of histological and lipid composition changes in fish fed the control diet, without the inclusion of AM but with a lipid base of corn oil and swine lard, corroborates the findings for other fish species when fish oil is replaced with vegetable oils [
21,
56]. Lima de Andrade et al. [
13] also found histopathological changes in tilapia juveniles fed diets with different LOA:LNA (n6/n3 = 12.02 and n6/n3 = 3.85), at temperatures of 30 and 20 °C. The frequency of cell displacement injury was lower in fish fed the diet containing the low n6/n3. This may be associated with lower lipid deposition. In addition, the liver of fish fed n6/n3 = 3.85 showed low cytoplasmic vacuolization at both temperatures [
13]. Therefore, an imbalance in dietary fatty acids can modify the function and morphology of this organ. It is possible to establish a relationship between the type of dietary fatty acids and the appearance of steatosis, that is, LOA > LNA > oleic acid [
21].
In a three-month trial, the marine fish,
Sparus aurata, fed diets based on fish oil, canola, flaxseed, or a mix of these oils, showed uniformity in cell size and little lipid accumulation. However, fish fed soybean oil presented steatosis foci, with hepatocytes containing numerous lipid vacuoles [
21]. In another study testing rearing temperatures (optimum 12 °C and suboptimal 5 °C) and lipid sources (soy oil replacing fish oil, at 50% and 100%) in Atlantic salmon, the suboptimal temperature positively influenced the deposition of fat in liver cells and the intestines [
56]. In addition, the diet with 100% soy oil resulted in the highest accumulation of fat in the liver at 5 °C [
56]. According to the authors, the accumulation of fat at low temperatures could be explained by the reduction in the activity of the enzymes involved in the esterification of fatty acids into triacylglycerol and phospholipids for very low-density lipoprotein (VLDL) production.
On the other hand, despite appearing at a low intensity, the presence of nuclei with karyolysis and karyorrhexis was lower in fish fed the lowest level of AM (5AM). It is possible that a minimal inclusion of n-3 PUFA would be sufficient to prevent these changes. Karyolysis is the complete dissolution of chromatin in a cell that is dying due to enzymatic degradation, and is usually associated with karyorrhexis, which occurs mainly as a result of necrosis [
57]. This is the first time that such a change has been reported with an increase in dietary PUFAs. These changes have already been described, in greater intensity than found in our study, for several species of fish, when exposed to contaminating agents [
58,
59,
60].
Several studies have shown that a drop in temperature causes fasting induced by cold, heat stress, and metabolic depression in fish [
61]. To mitigate these effects, adequate nutrition is suggested in this study. We verified that AM, as a source of DHA in Nile tilapia, provides the body with the accumulation of DHA, an important fatty acid for adequate metabolic functioning of fish, when subjected to low suboptimal temperatures. In addition, increasing the inclusion of AM decreased the hepatopancreas lipid content, increased the hepatopancreas concentration of n-3 fatty acid series, and promoted significant improvements in the morphophysiology of the hepatopancreas, preventing signs of macrosteatosis, microsteatosis, and necrosis, seen in fish fed a practical diet, without any DHA supplementation. These data provide evidence for the physiological need for DHA supplementation in Nile tilapia diets at suboptimal temperatures and the potential for the development of specific winter aquafeeds for the species, with the aim of improving growth performance and physiological well-being.
In addition to its important role in fish performance and metabolism, DHA appears to play an important role in the cardiac, cardiovascular, brain, and visual functions of humans [
62]. Currently, a greater intake of n-3 fatty acids is desirable for reducing the risk of many of the highly prevalent chronic diseases in Western societies, as well as in developing countries.
Aurantiochytrium sp. meal supplementation in Nile tilapia diets can also provide a source of n-3 fatty acids for human consumption, adding value to this freshwater species.
More research should be carried out to further evaluate the effects of dietary AM supplementation in the intestinal functions of fish. We have registered a positive effect on intestinal health; however, a wide range of analyses including intestinal microbiota composition and gut-associated lymphoid tissues (GALT) immune responses would provide further understanding about the effect of such a novel additive. Furthermore, future studies should evaluate the dietary supplementation of AM in growing-out conditions to validate our findings in a controlled lab situation.


 and
and 



{kind=link}
{kind=link}
{kind=link}