
Survival and Physiological Recovery after Capture by Hookline: The Case Study of the Blackspot Seabream (Pagellus bogaraveo)

 ,
,  , , , and
, , , and
Abstract
:
1. Introduction
2. Results
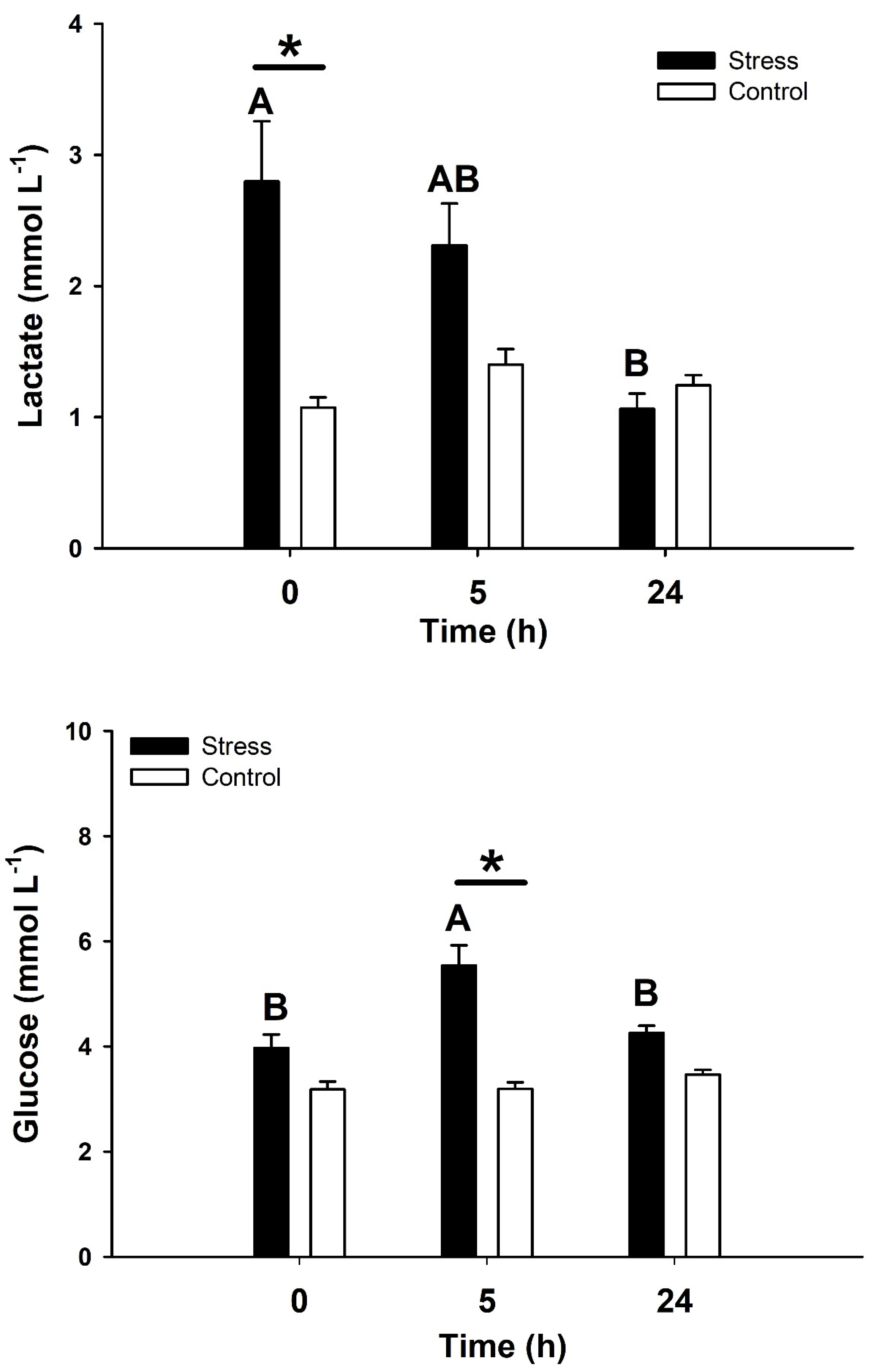
2.1. Stress Responses and Physiological Recovery in Ground Facilities
2.2. Survival Rates Onboard
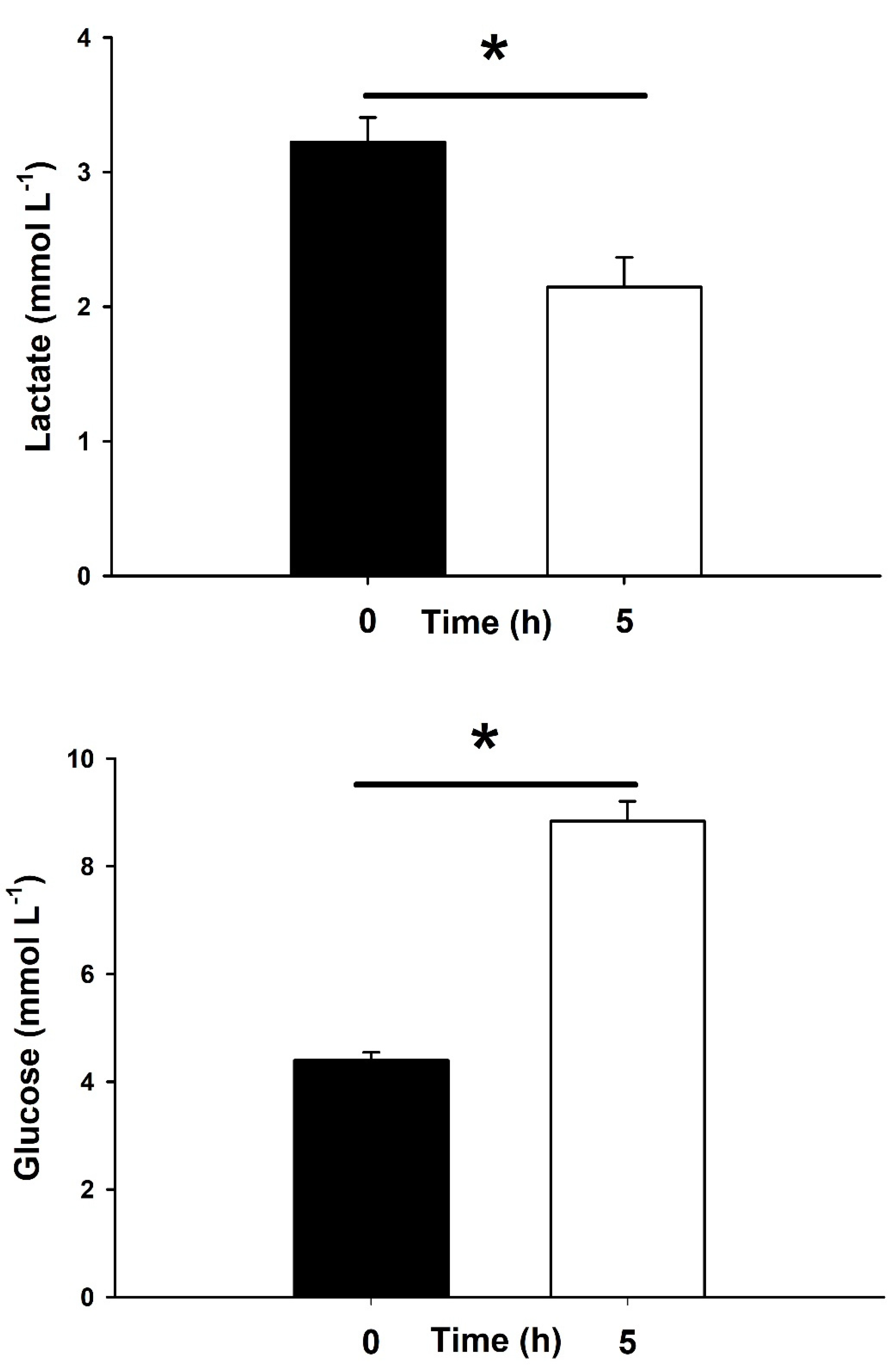
2.3. Stress Responses and Physiological Signs of Recovery Onboard
3. Discussion
3.1. Physiological Recovery
3.2. Survival Rates
4. Materials and Methods
4.1. Ethics Statement
4.2. Time-Course Responses after Acute Stress in Ground Facilities
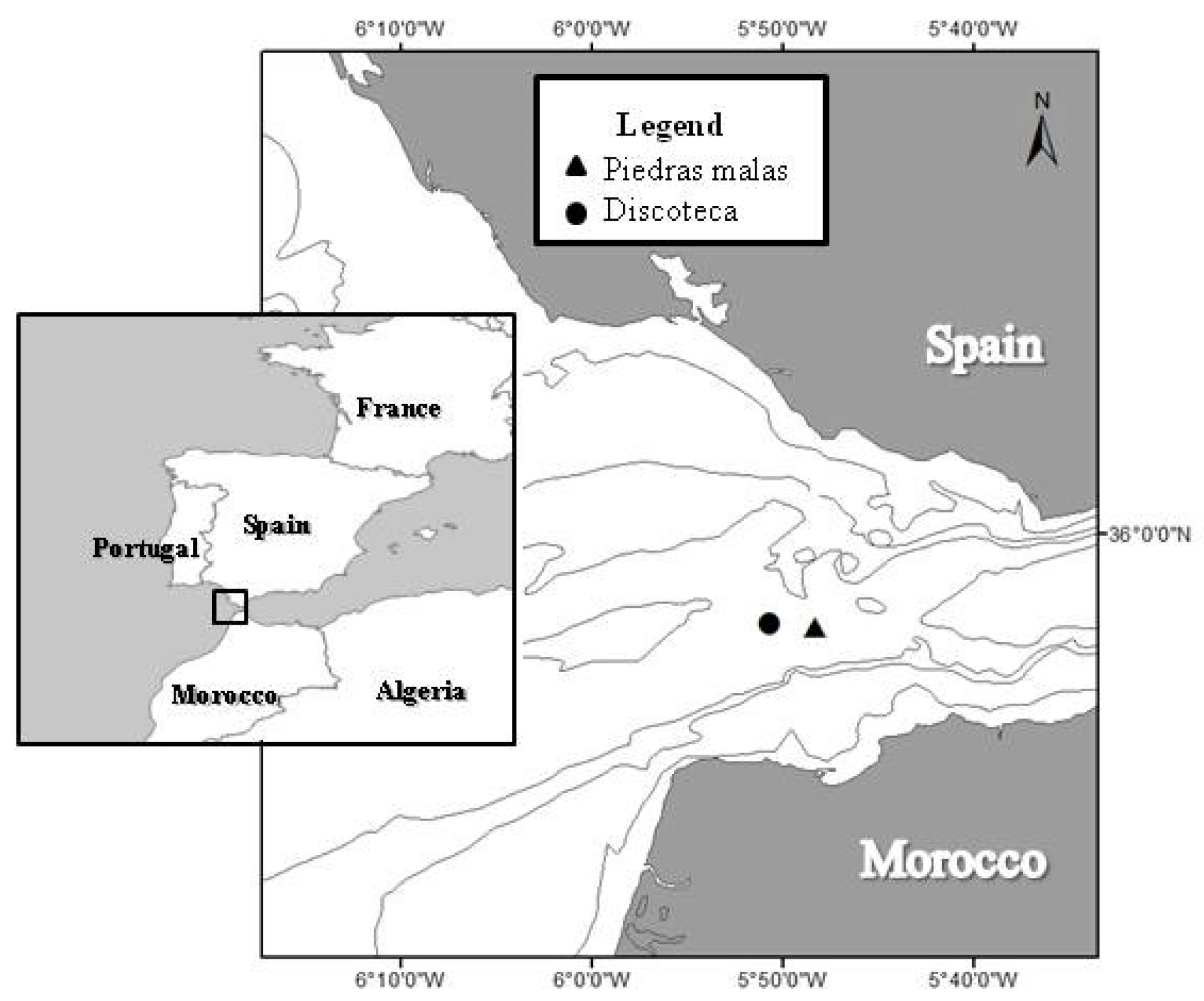
4.3. Geographical Location of the Fisheries, Vessel, and Hookline Characteristics
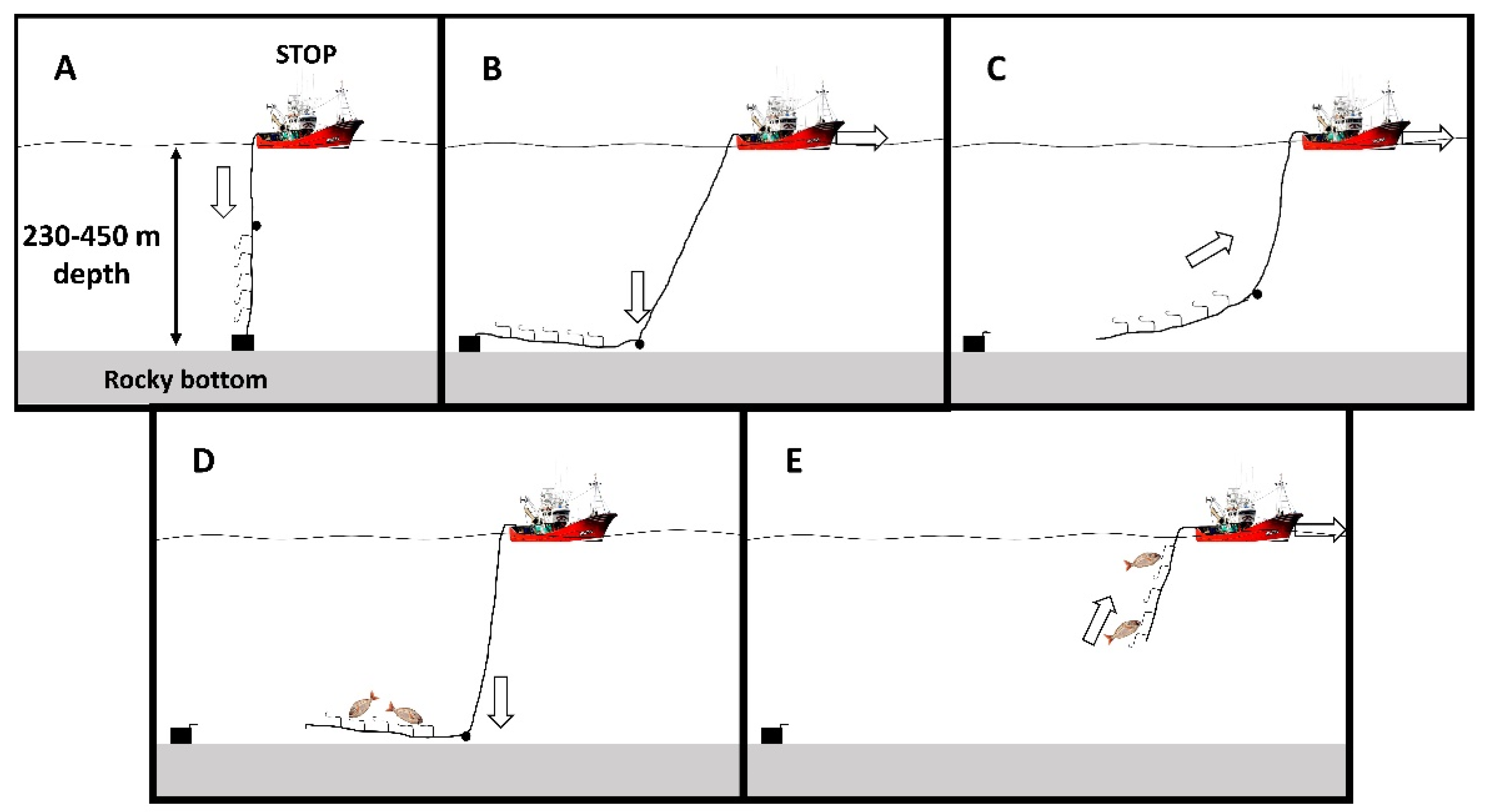
4.4. Survival Rates Onboard
4.5. Physiological Recovery Onboard
4.6. Fish Tagging Onboard
4.7. Plasma Parameters
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Beek, F.A.; Van Leeuwen, P.I.; Rijnsdorp, A.D. On the survival of plaice and sole discards in the otter-trawl and beam-trawl fisheries in the North Sea. Neth. J. Sea Res. 1990, 26, 151–160. [Google Scholar] [CrossRef]
- Ellis, J.R.; Burt, G.J.; Grilli, G.; McCully Phillips, S.R.; Catchpole, T.L.; Maxwell, D.L. At-vessel mortality of skates (Rajidae) taken in coastal fisheries and evidence of longer-term survival. J. Fish Biol. 2018, 92, 1702–1719. [Google Scholar] [CrossRef]
- Broadhurst, M.K.; Suuronen, P.; Hulme, A. Estimating collateral mortality from towed fishing gear. Fish Fish. Ser. 2006, 7, 180–218. [Google Scholar] [CrossRef]
- Methling, C.; Skov, P.V.; Madsen, N. Reflex impairment, physiological stress, and discard mortality of European plaice Pleuronectes platessa in an otter trawl fishery. ICES J. Mar. Sci. 2017, 496, 207–218. [Google Scholar] [CrossRef]
- Morfin, M.; Méhault, S.; Benôit, H.P.; Kopp, D. Narrowing down the number of species requiring detailed study as candidates for the EU Common Fisheries Policy discard ban. Mar. Policy 2017, 77, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Veiga, P.; Pita, C.; Rangel, M.; Goncalves, J.M.S.; Campos, A.; Fernandes, P.G.; Sala, A.; Virgili, M.; Lucchetti, A.; Brcic, J.; et al. The EU landing obligation and European small-scale fisheries: What are the odds for success? Mar. Policy 2016, 64, 64–71. [Google Scholar] [CrossRef]
- Uhlmann, S.S.; Theunynck, R.; Ampe, B.; Desender, M.; Soetaert, M.; Depestele, J. Injury, reflex impairment, and survival of beam-trawled flatfish. ICES J. Mar. Sci. 2016, 73, 1244–1254. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, A.J.; Serafy, J.E.; Cooke, S.J.; Hammerschlag, N. Physiological stress response, reflex impairment, and survival of five sympatric shark species following experimental capture and release. Mar. Ecol. Prog. Ser. 2014, 496, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Humborstad, O.B.; Breen, M.; Davis, M.W.; Lokkeborg, S.; Mangor-Jensen, A.; Midling, K.O.; Olsen, R.E. Survival and recovery of longline- and pot-caught cod (Gadus morhua) for use in capture-based aquaculture (CBA). Fish. Res. 2016, 174, 103–108. [Google Scholar] [CrossRef]
- Wilson, S.M.; Raby, G.D.; Burnett, N.J.; Hinch, S.G.; Cooke, S.J. Looking beyond the mortality of bycatch: Sublethal effects of incidental capture on marine animals. Biol. Conserv. 2014, 171, 61–72. [Google Scholar] [CrossRef]
- Frick, L.H.; Reina, R.D.; Walker, T.I. Stress related physiological changes and post-release survival of Port Jackson sharks (Heterodontus portusjacksoni) and gummy sharks (Mustelus antarcticus) following gill-net and longline capture in captivity. J. Exp. Mar. Biol. Ecol. 2010, 385, 29–37. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Jerez-Cepa, I.; Ruiz-Jarabo, I. Physiology: An Important Tool to Assess the Welfare of Aquatic Animals. Biology 2021, 10, 61. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Skrzynska, A.K.; Maiorano, E.; Bastaroli, M.; Naderi, F.; Miguez, J.M.; Martinez-Rodriguez, G.; Mancera, J.M.; Martos-Sitcha, J.A. Impact of air exposure on vasotocinergic and isotocinergic systems in gilthead sea bream (Sparus aurata): New insights on fish stress response. Front. Physiol. 2018, 9, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costas, B.; Conceicao, L.; Aragao, C.; Martos, J.A.; Ruiz-Jarabo, I.; Mancera, J.; Afonso, A. Physiological responses of Senegalese sole (Solea senegalensis Kaup, 1858) after stress challenge: Effects on non-specific immune parameters, plasma free amino acids and energy metabolism. Aquaculture 2011, 316, 68–76. [Google Scholar] [CrossRef]
- Ruiz-Jarabo, I.; Amanajas, R.D.; Baldisserotto, B.; Mancera, J.M.; Val, A.L. Tambaqui (Colossoma macropomum) acclimated to different tropical waters from the Amazon basin shows specific acute-stress responses. Comp. Biochem. Physiol. A 2020, 245, 110706. [Google Scholar] [CrossRef] [PubMed]
- Laiz-Carrion, R.; Guerreiro, P.M.; Fuentes, J.; Canario, A.V.M.; Martin Del Rio, M.P.; Mancera, J.M. Branchial osmoregulatory response to salinity in the gilthead sea bream, Sparus auratus. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303A, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Iwama, G.K.; Afonso, L.O.; Vijayan, M.M. Stress in fish. Ann. N. Y. Acad. Sci. 1998, 851, 304–310. [Google Scholar] [CrossRef]
- Skomal, G.B.; Mandelman, J.W. The physiological response to anthropogenic stressors in marine elasmobranch fishes: A review with a focus on the secondary response. Comp. Biochem. Physiol. A 2012, 162, 146–155. [Google Scholar] [CrossRef]
- Sangiao-Alvarellos, S.; Arjona, F.J.; Martin del Rio, M.P.; Miguez, J.M.; Mancera, J.M.; Soengas, J.L. Time course of osmoregulatory and metabolic changes during osmotic acclimation in Sparus auratus. J. Exp. Biol. 2005, 208, 4291–4304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Arjona, F.J.; Vargas-Chacoff, L.; Ruiz-Jarabo, I.; Gonçalves, O.; Pâscoa, I.; Martín del Río, M.P.; Mancera, J.M. Tertiary stress responses in Senegalese sole (Solea senegalensis Kaup, 1858) to osmotic challenge: Implications for osmoregulation, energy metabolism and growth. Aquaculture 2009, 287, 419–426. [Google Scholar] [CrossRef]
- Rotllant, J.; Ruane, N.M.; Dinis, M.T.; Canario, A.V.; Power, D.M. Intra-adrenal interactions in fish: Catecholamine stimulated cortisol release in sea bass (Dicentrarchus labrax L.). Comp. Biochem. Physiol. A 2006, 143, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Imbrogno, S.; Cerra, M.C. Hormonal and autacoid control of cardiac function. In The Cardiovascular System: Morphology, Control and Function; Gamperl, A.K., Gillis, T.E., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: New York, NY, USA, 2017; Volume 36, pp. 265–315. [Google Scholar]
- Sánchez-Leal, R.F.; Bellanco, M.J.; Fernández-Salas, L.M.; García-Lafuentes, J.; Gasser-Rubinat, M.; González-Pola, C.; Hernández-Molina, F.J.; Pelegrí, J.L.; Peliz, A.; Relvas, P.; et al. The Mediterranean Overflow in the Gulf of Cadiz: A rugged journey. Sci. Adv. 2018, 3, eaao0609. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Estrada, J.C.; Gil-Herrera, J.; Pulido-Clavo, I.; Czerwinski, I.A. Is it possible to differentiate between environmental and fishery effects on abundance-biomass variation? A case study of blackspot seabream (Pagellus bogaraveo) in the Strait of Gibraltar. Fish. Oceanogr. 2017, 26, 455–475. [Google Scholar] [CrossRef]
- ICES. Report of the Working Group on the Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP); ICES: Copenhagen, Denmark, 2012; p. 929. [Google Scholar]
- Silva, L.; Gil, J.; Sobrino, J. Definition of fleet components in the Spanish artisanal fishery of the Gulf of Cádiz (SW Spain ICES division IXa). Fish. Res. 2002, 59, 117–128. [Google Scholar] [CrossRef]
- Higgins, R.M.; Diogo, H.; Isidro, E.J. Modelling growth in fish with complex life histories. Rev. Fish Biol. Fish. 2015, 25, 449–462. [Google Scholar] [CrossRef]
- IEO. Estudios de Marcado y Recaptura de Especies Marinas; Servicios Periodísticos, S.L.: Madrid, Spain, 2009; Volume 2, p. 263. [Google Scholar]
- Wunderink, Y.S.; Engels, S.; Halm, S.; Yufera, M.; Martinez-Rodriguez, G.; Flik, G.; Klaren, P.H.M.; Mancera, J.M. Chronic and acute stress responses in Senegalese sole (Solea senegalensis): The involvement of cortisol, CRH and CRH-BP. Gen. Comp. Endocrinol. 2011, 171, 203–210. [Google Scholar] [CrossRef]
- Frick, L.H.; Walker, T.I.; Reina, R.D. Immediate and delayed effects of gill-net capture on acid-base balance and intramuscular lactate concentration of gummy sharks, Mustelus antarcticus. Comp. Biochem. Physiol. A 2012, 162, 88–93. [Google Scholar] [CrossRef]
- McArley, T.J.; Herbert, N.A. Mortality, physiological stress and reflex impairment in sub-legal Pagrus auratus exposed to simulated angling. J. Exp. Mar. Biol. Ecol. 2014, 461, 61–72. [Google Scholar] [CrossRef]
- McCormick, S.D. The Hormonal Control of Osmoregulation in Teleost Fish. In Encyclopedia of Fish Physiology: From Genome to Environment; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 2, pp. 1466–1473. [Google Scholar]
- Laiz-Carrion, R.; Martin Del Rio, M.P.; Miguez, J.M.; Mancera, J.M.; Soengas, J.L. Influence of cortisol on osmoregulation and energy metabolism in gilthead seabream Sparus aurata. J. Exp. Zool. Part A Comp. Exp. Biol. 2003, 298A, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Guillen, C.; Yufera, M.; Engrola, S. Daily feeding and protein metabolism rhythms in Senegalese sole post-larvae. Biol. Open 2017, 6, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Revill, A.S.; Dulvy, N.K.; Holst, R. The survival of discarded lesser-spotted dogfish (Scyliorhinus canicula) in the Western English Channel beam trawl fishery. Fish. Res. 2005, 71, 121–124. [Google Scholar] [CrossRef]
- Barría, C.; Navarro, J.; Coll, M. Trophic habits of an abundant shark in the northwestern Mediterranean Sea using an isotopic non-lethal approach. Estuar. Coast. Shelf Sci. 2018, 207, 383–390. [Google Scholar] [CrossRef]
- Guijarro, B.; Quetglas, A.; Moranta, J.; Ordines, F.; Valls, M.; González, N.; Massutí, E. Inter- and intra-annual trends and status indicators of nektobenthic elasmobranchs off the Balearic Islands (northwestern Mediterranean). Sci. Mar. 2012, 76, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Brooks, E.J.; Mandelman, J.W.; Sloman, K.A.; Liss, S.; Danylchuk, A.J.; Cooke, S.J.; Skomal, G.B.; Philipp, D.P.; Sims, D.W.; Suski, C.D. The physiological response of the Caribbean reef shark (Carcharhinus perezi) to longline capture. Comp. Biochem. Physiol. A 2012, 162, 94–100. [Google Scholar] [CrossRef]
- Ruiz-Jarabo, I.; Klaren, P.H.M.; Louro, B.; Martos-Sitcha, J.A.; Pinto, P.I.S.; Vargas-Chacoff, L.; Flik, G.; Martínez-Rodríguez, G.; Power, D.M.; Mancera, J.M.; et al. Characterization of the peripheral thyroid system of gilthead seabream acclimated to different ambient salinities. Comp. Biochem. Physiol. A 2017, 203, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Chacoff, L.; Saavedra, E.; Oyarzún, R.; Martínez-Montaño, E.; Pontigo, J.P.; Yáñez, A.; Ruiz-Jarabo, I.; Mancera, J.M.; Ortiz, E.; Bertrán, C. Effects on the metabolism, growth, digestive capacity and osmoregulation of juvenile of Sub-Antarctic Nototheniod fish Eleginops maclovinus acclimated at different environmental salinities. Fish Physiol. Biochem. 2015, 41, 1369–1381. [Google Scholar] [CrossRef]
- Pujante, I.M.; Martos-Sitcha, J.A.; Moyano, F.J.; Ruiz-Jarabo, I.; Martinez-Rodriguez, G.; Mancera, J.M. Starving/re-feeding processes induce metabolic modifications in thick-lipped grey mullet (Chelon labrosus, Risso 1827). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 180, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Laiz-Carrion, R.; Viana, I.R.; Cejas, J.; Ruiz-Jarabo, I.; Jerez, S.; Martos, J.A.; Almansa, E.; Mancera, J.M. Influence of food deprivation and high stocking density on energetic metabolism and stress response in red porgy, Pagrus pagrus L. Aquac. Int. 2012, 20, 585–599. [Google Scholar] [CrossRef]
- Herrera, M.; Ruiz-Jarabo, I.; Hachero, I.; Vargas-Chacoff, L.; Amo, A.; Mancera, J. Stocking density affects growth and metabolic parameters in the brill (Scophthalmus rhombus). Aquac. Int. 2012, 20, 1041–1052. [Google Scholar] [CrossRef]
- Avila, P.R.; Marques, S.O.; Luciano, T.F.; Vitto, M.F.; Engelmann, J.; Souza, D.R.; Pereira, S.V.; Pinho, R.A.; Lira, F.S.; De Souza, C.T. Resveratrol and fish oil reduce catecholamine-induced mortality in obese rats: Role of oxidative stress in the myocardium and aorta. Braz. J. Nutr. 2013, 110, 1580–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, M.F. Sudden cardiac death: The lost fatty acid hypothesis. J. Assoc. Physicians 2006, 99, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Louison, M.J.; Adhikari, S.; Stein, J.A.; Suski, C.D. Hormonal responsiveness to stress is negatively associated with vulnerability to angling capture in fish. J. Exp. Biol. 2017, 220, 2529–2535. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Gomez, M.L.; Huntingford, F.A.; Overli, O.; Thornqvist, P.O.; Hoglund, E. Response to environmental change in rainbow trout selected for divergent stress coping styles. Physiol. Behav. 2011, 102, 317–322. [Google Scholar] [CrossRef]
- Gesto, M.; Hernandez, J.; Lopez-Patino, M.A.; Soengas, J.L.; Miguez, J.M. Is gill cortisol concentration a good acute stress indicator in fish? A study in rainbow trout and zebrafish. Comp. Biochem. Physiol. A 2015, 188, 65–69. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Group | 0 h | 5 h | 24 h |
|---|---|---|---|---|
| TAG (mmol L−1) | Control | 0.75 ± 0.04 | 1.01 ± 0.16 | 0.74 ± 0.12 |
| Stress | 0.79 ± 0.09 | 0.64 ± 0.08 | 0.65 ± 0.07 | |
| Proteins (mg dL−1) | Control | 27.5 ± 1.4 | 25.7 ± 0.8 | 27.5 ± 1.3 |
| Stress | 28.1 ± 1.0 | 23.6 ± 0.8 | 24.6 ± 1.5 | |
| Osmolality (mOsm kg−1) | Control | 264 ± 4 * | 267 ± 1 | 263 ± 1 |
| Stress | 291 ± 3 A | 260 ± 2 B | 257 ± 2 B |
| Parameter | 0 h | 5 h |
|---|---|---|
| TAG (mmol L−1) | 1.79 ± 0.18 | 1.65 ± 0.18 |
| Proteins (mg dL−1) | 26.6 ± 0.47 | 21.5 ± 0.4 * |
| Osmolality (mOsm kg−1) | 302 ± 3 | 291 ± 3 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Jarabo, I.; Fernández-Castro, M.; Jerez-Cepa, I.; Barragán-Méndez, C.; Pérez, M.; Pérez, E.; Gil, J.; Canoura, J.; Farias, C.; Mancera, J.M.; et al. Survival and Physiological Recovery after Capture by Hookline: The Case Study of the Blackspot Seabream (Pagellus bogaraveo). Fishes 2021, 6, 64. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040064
Ruiz-Jarabo I, Fernández-Castro M, Jerez-Cepa I, Barragán-Méndez C, Pérez M, Pérez E, Gil J, Canoura J, Farias C, Mancera JM, et al. Survival and Physiological Recovery after Capture by Hookline: The Case Study of the Blackspot Seabream (Pagellus bogaraveo). Fishes. 2021; 6(4):64. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040064
Chicago/Turabian StyleRuiz-Jarabo, Ignacio, Miriam Fernández-Castro, Ismael Jerez-Cepa, Cristina Barragán-Méndez, Montse Pérez, Evaristo Pérez, Juan Gil, Jesús Canoura, Carlos Farias, Juan Miguel Mancera, and et al. 2021. "Survival and Physiological Recovery after Capture by Hookline: The Case Study of the Blackspot Seabream (Pagellus bogaraveo)" Fishes 6, no. 4: 64. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040064