
Adolescence and Postpartum: Two Life Periods to Deepen Our Understanding of the Complexity of Female Rat Sexual Behavior


Physiology and Nutrition Department, Facultad de Ciencias, Universidad de la Republica, Montevideo 11400, Uruguay
Sexes 2022, 3(2), 282-297; https://0-doi-org.brum.beds.ac.uk/10.3390/sexes3020022
Submission received: 9 April 2022
/
Accepted: 13 May 2022
/
Published: 23 May 2022
(This article belongs to the Section Biological and Psychosocial Basis Underlying Sexual Response and Differences)
Abstract
:The sexual behavior of the female rat has been extensively studied and used as a model for human female sexuality. The development of models that focus on sexual interaction from the female’s perspective has demonstrated the complexity of this behavior, in which motivational and affective aspects have great relevance. This review describes, from a historical perspective, some of the studies that have shed light on this complexity and presents the potential of two periods in the female’s life to expand our understanding of sexual behavior: adolescence and postpartum estrus. Cycling adolescent female rats can express sexual behavior even though some of the underlying circuitry is still developing. Therefore, this is an ideal time for studying the ontogeny of sexual behavior, the interaction of sexual and social motivations, and the neuroendocrine changes that lead to adult behavior. Female rats also show a fertile estrus a few hours after parturition. During this period, rats are simultaneously sexually and maternally motivated, making postpartum estrus an excellent model for exploring the interaction of these two social motivations. What we know so far about female sexual behavior in these unique periods of life reaffirms its richness and shows that much remains to be known.
1. Introduction
In the last decades, our conceptualization of the sexual behavior of the female rat has shifted from a passive and stereotyped response to an active and complex behavior, in which the interaction between affective, motivational, and cognitive processes determines female’s behavioral output. This vision has allowed the use of female rat sexual behavior, in particular sexual motivation, to model different aspects of human female sexuality (for revision, see [1,2,3,4]). Novel approaches and the development of new models to study female sexual behavior continue to show its richness and complexity. This review aims to contribute by presenting two unique life-history contexts in which sexual behavior is expressed: adolescence and postpartum estrus. Sexual behavior of females during these periods has been understudied but has the potential to expand our knowledge about this fascinating affiliative behavior. The two first sections of the review summarize central concepts of female rat sexual behavior with a historical perspective. These sections are based on hallmark studies that have advanced the understanding of the sexual behavior of female rats and on reviews of the last years on the subject. Then, based on an extensive review of the literature on adolescence and postpartum estrus in the rat, it describes sexual behavior during these periods and discusses the importance of further study.
2. Is Female Rat Sexual Behavior a Stereotyped and Passive Response?
Sexual behavior of the female rat is relatively easy to study because it compromises the expression of stereotype and characteristic behavioral units, and in fact, it has been extensively investigated over the last one hundred years. Why, then, is there still so much we do not know about this behavior?
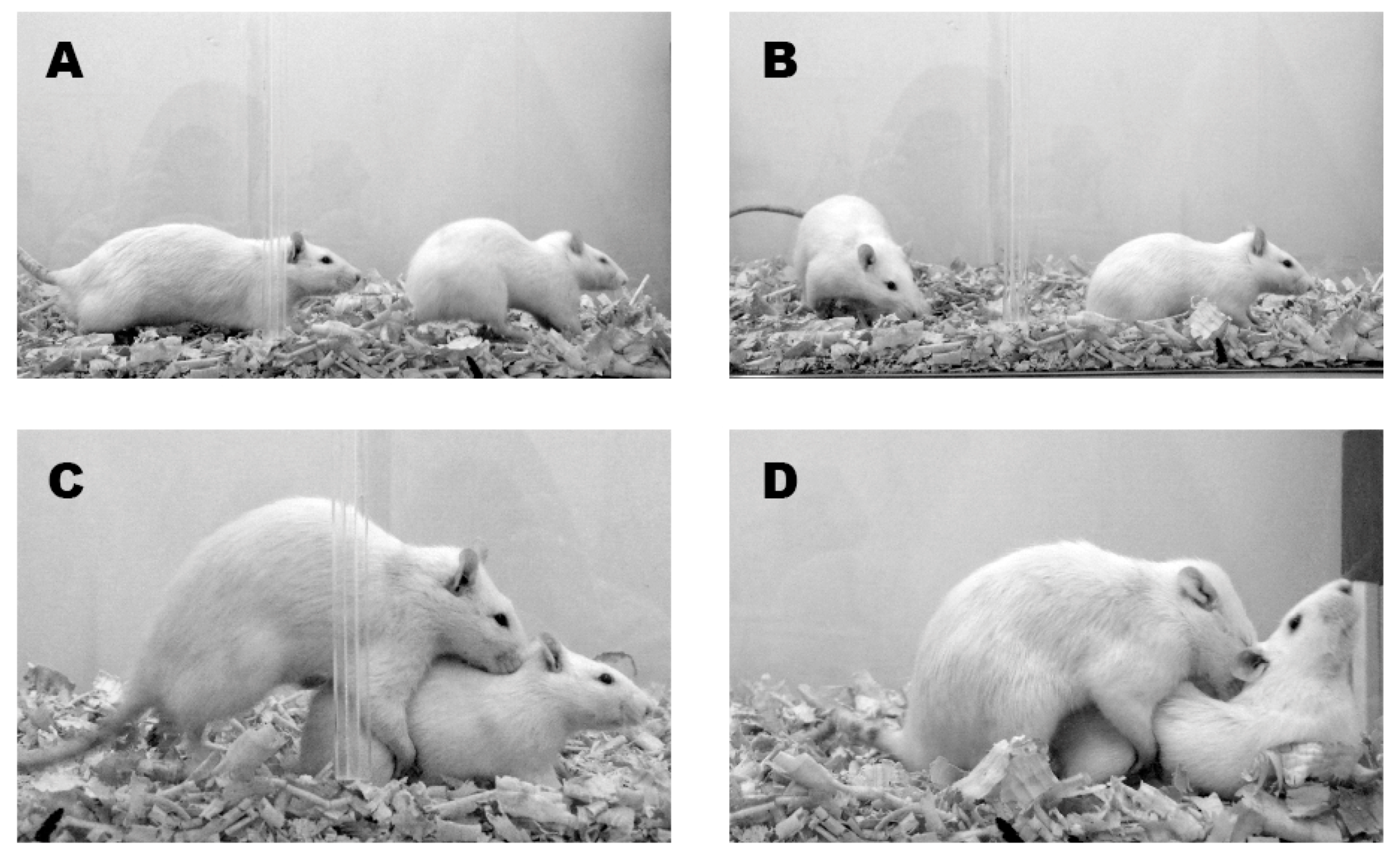
During a typical copulatory bout, sexually active female rats display a series of behaviors that include solicitations, hops and darts, ear wiggling, and presentation postures, which are displays that have been collectively called proceptive [5] or paracopulatory [6] behaviors (Figure 1A–C). These behaviors usually promote male chasing and mounting. Female rats display lordosis in response to mount (Figure 1D), a corporal posture characterized by the dorsiflexion of the spine that elevates the rump and anogenital region, allowing penile intromission [7].
While the expression of proceptive components is variable and has been proposed to reflect the female’s motivational state, the adoption of the lordosis posture is more reflexive and has been used as an index of female sexual receptivity to male mounting [5,8,9]. Probably because lordosis can be measured and quantified in a precise and unequivocal way, this reflexive response has been historically studied as a hallmark of female rat sexual behavior. Donald Pfaff and colleagues carefully studied lordosis in the rat, identifying the somatosensory stimulation that induces it and the neuroendocrine mechanisms that underlie this response. Lordosis is primarily triggered by the stimulation of the flanks and the perineal region of the female during mounting [8,9] and is strictly dependent on sexual steroid action, mainly estrogen, on several regions of the nervous system collectively known as the lordosis behavior circuit [10,11,12,13]. Thus, several hypothalamic nucleus and areas rich in ovarian steroids receptors and densely connected between each other, including the ventromedial nucleus of the hypothalamus (VMN), the anterior hypothalamus, the arcuate nucleus, and the medial preoptic area (mPOA), control this supraspinal reflex. The VMN, considered the “heart” of female rat sexual behavior, is the final common projection from this integrative circuit to lower brainstem and spinal cord nuclei that complete the lordosis behavior circuit (for an extensive revision of lordosis behavior circuit, see [14,15,16]).
The study of female sexual behavior, particularly their lordosis response, under standard laboratory conditions significantly advanced our knowledge of this behavior. However, interpreting this social affiliative behavior mainly based on lordosis analysis may lead to viewing it as more rigid and passive than it actually is [6,17]. As early as 1976, in his seminal paper on the sexual behavior of female mammals, Frank Beach [5] wrote:
Proceptive behavior is functionally as important as other patterns traditionally termed “receptive”, but the female’s tendency to display appetitive responses finds little opportunity for expression in laboratory experiments which focus exclusively upon her receptive behavior, or upon the male’s execution of this coital pattern.
3. Adding Complexity to the Sexual Behavior of Females: The Role of Sexual Motivation
One of the first approaches employed to better understand the richness of female sexual behavior was developed by Pierce and Nuttal (1961) and consisted ofadapting a sexual arena (either a circular or rectangular enclosure) into two compartments connected by a hole through which only the female could pass [18]. Under this experimental condition, the female can pace the rhythm of sexual stimulation she receives by escaping and re-entering the male compartment [18,19]. Extensive work by Mary Erskine and other researchers made it possible to determine the factors that lead the female to perform a specific pattern of approach/withdrawal from the male. It also established the importance of the rhythm of sexual stimulation to trigger the neuroendocrine response necessary to sustain gestation and generate a positive affective state associated with mating [19,20,21,22,23,24].
To further challenge the constraints of standard laboratory mating conditions that usually include interacting with one male in a small enclosure, McClintock and Adler studied the sexual pattern of interaction of a female and a male confined in a large semi-natural enclosure with several chambers and burrows. These authors reported fewer intromissions and a greater inter-intromission interval in pair copulation in this environment relative to standard laboratory conditions [25]. Moreover, in a series of studies in which they analyzed mating in groups in these semi-natural enclosures, they observed that females and males copulate with several individuals within a reproductive bout and that both sexes differ in the sequence and temporal pattern of copulation. Because mating is promiscuous, and females also regulate its rhythm, unlike males, females receive intromissions and ejaculations in a random order and show larger inter-intromission intervals than males. Thus, females copulating in groups received fewer intromissions at longer inter-intromission intervals than females mating in pairs in the same environment [26,27]. These studies demonstrate the importance of both the physical and social environment for the expression of sexual behavior in the female rat. Interestingly, in both types of sexual interactions in semi-natural enclosures, females display a particular behavioral pattern consisting of approach toward, orientation to, and withdrawal from the male, which is usually followed by males mounting. This behavioral pattern, called sexual solicitation, proposedly reflects the active female role in mating, as females that solicited more often received more intromissions and ejaculations [26,27,28]. These authors also described that during group mating, females compete with each other for the opportunity to mate with a male as his ejaculation approaches [27]. In recent years, there has been a resurgence in the use of this model analyzing the importance of behaviors displayed by the female and the male in mating dynamics [29,30,31,32,33]. For example, it has been shown that female rats living in a semi-natural enclosure present an abrupt transition from non-receptivity to receptivity, which is characterized by a drastic increase in the expression of lordosis and paracopulatory behaviors from very low levels prior to non-receptivity to very high levels during behavioral estrus. These high levels are sustained through estrus and sharply drop again at its end [31].
In the last decades, other models with greater ecological validity have been developed than standard laboratory mating, including the bilevel chamber with one male [34,35,36] and mate-choice models with two or more males [37,38]. Results from Fay Guarraci’s laboratory have shown that although mating in the female rat is promiscuous, she exhibits preference for copulating with a particular male. In a mate-choice test where female rats can interact freely with either of two male stimuli, they spend more time and make more visits to one male over another [38,39,40].Together, the use of these models has demonstrated two main aspects of female sexual behavior: (1) the context of sexual interaction conditions female performance, and (2) females have an active role in copulation, and therefore, their motivational state strongly influences mating rhythm (see Table 1 for a summary).
Female behavior during a sexual interaction provides clues as to sexual motivation; however, specific models are required to separate the analysis of the motivational state from the execution of the behavior [17,41]. Motivation-assessment tests based on learning tasks (e.g., lever-pressing, place preference conditioning, or nose poking) [23,42,43] and on unconditioned approaching behaviors (e.g., partner preference with or without physical contact or multiple partner choice arena) [37,44,45] have provided further evidence of the motivational nature of female sexual behavior [4,17] (Table 1). For example, sexual preference tests—or partner preference or sexual incentive tests—are among the most commonly unconditional tests used. They evaluate the approach behavior of the female to a sexual stimulus (usually a sexually experienced male) in relation to a stimulus with less or no sexual incentive value (usually castrated male or sexually active female) [4,44,46]. These tests are performed in different apparatuses: an open field with attached boxes containing the stimulus animals [47], a camera divided into three compartments with the stimuli positioned in the extremes [48], or a Y-shaped maze with choice compartments at the end of the arms ([49], described in the next section). The central feature of all these tests is that the female can see, hear, and smell the stimuli but does not have physical access to them in order to evaluate the incentive value of the male without interference from the sexual experience, although allowing interaction is also used to study other aspects of female behavior [48]. These models made it possible to establish the role of the endocrine profile and previous sexual experience in the regulation of female sexual motivation (for examples, see [44,47,48,49,50,51]).
Interestingly, Paredes and Alonso (1997) explored the role of female´s ability to pace copulatory rhythm on its reinforcing value by assessing conditioned place preference (CPP) for context paired with pace and non-pace mating experiences. In their experimental conditions, the authors observed that only self-paced mating generated CPP, similar to that induced by morphine administration [23]. It seems that the distributed sexual stimulation received by the female rather than her ability to control the rhythm of mating is the main factor of the pace mating experience responsible for the induction of a positive hedonic affective state. This conclusion is based on the fact that distributed artificial vaginocervical simulation [52] and distributed clitoral stimulation [53]—but not social interaction with a male or continuous clitoral stimulation—induce CPP in sexually active females. It has been proposed that the physiological and behavioral changes associated with distributed clitoris stimulation (including cervico-uterine reflexes, 50 kHz ultrasonic vocalizations (USV) emission and long-term behavioral changes that depend on a reward state) are consistent with the induction of an orgasm-like response in the female rat [54].
These voluntary and flexible aspects of the female’s sexual behavior, which depend on her motivational state, have been proposed to be regulated by a limbic-hypothalamic net of areas and its upstream connections with the mesolimbic dopaminergic system. In particular, a major role has been attributed to the mPOA, the VMN, and the medial amygdala in assigning sexual incentive or motivational value to a male. Lesion or inactivation of these closely connected areas reduces the expression of paracopulatory behaviors and/or the preference for male or male-related cues in sexually active females [55,56,57,58,59,60]. In addition, the local administration of naloxone (an opioid antagonist) to these areas prevents the formation of CPP associated with pace mating [61]. It has been postulated that mPOA acts as a connection node of this limbic-hypothalamic circuitry with the mesolimbic dopaminergic system, which is considered a general motivational system that promotes behavioral activation (for extensive revision, see [14,15,62,63,64,65] and, on the other hand, [66]).
The background presented in these first sections reveals the complexity of the control of sexual behavior in the female rat (Table 1) and encourages further exploration. As Blaustein and Erskine (2002) [6] pointed out:
As studies of brain and behavior become more sophisticated, it has become clear that our understanding of the neurobiological mechanisms controlling behavior will be significantly deepened if more naturalistic aspects of the behavior are incorporated into our studies.
Under this framework, it is important to note that most of the studies presented here were conducted on adult rats and, more often, in ovariectomized females treated with estradiol and progesterone to induce behavioral estrus. Although the hormonal induction of estrus evokes a behavioral profile similar to that observed in natural estrus and homogenizes the hormonal background between females, it is still an artificial state. Moreover, throughout their reproductive life, naturally cycling females may express sexual behavior at different life-history contexts, which can vary significantly from one another, making it difficult to generalize conclusions obtained in studies with ovariectomized adult rats. For example, female rats begin to exhibit sexual behavior during adolescence, around postnatal day 40 [67,68], a developmental period when the neural circuits that control this behavior are still immature [69,70]. Another reproductive context in which females are able to display sexual behavior is the postpartum estrus that occurs some hours after delivery [71,72,73]. During this brief period of time, rats are sexually and maternally motivated at the same time. What do adolescence and postpartum estrus have to tell us about the sexual behavior and motivation of the female rat?

{kind=link}
{kind=link}
Table 1.
Behavior of sexually active female rats under different testing conditions and its contribution to the understanding of sexual behavior.
Table 1.
Behavior of sexually active female rats under different testing conditions and its contribution to the understanding of sexual behavior.
| Testing Conditions | Female Behavior | Contributions |
|---|---|---|
| Sexual interaction tests | ||
| Standard laboratory conditions | ||
| - One female and one male in small enclosure | Lordosis and paracopulatory behaviors | Endocrine regulation. Lordosis behavior circuitry. for revision see [5,6,15,16,17] |
| Adding complexity to physical environment | ||
| - Paced-mating chamber - Bilevel chamber - Semi-natural enclosure | Lordosis and other paracopulatory displays Pacing the rhythm of mating | Active role of the female. Importance of the stimulation received. Significance of the context. Broadening of endocrine and contextual modulation. Limbic-hypothalamic control. for revision see [1,4,6,17,19,28] |
| Adding complexity to social environment | ||
| - Mate-choice test with two or more males - Group mating in semi-natural enclosure | Mating preference Female competition | |
| Sexual motivation tests | ||
| Unconditioned tests | ||
| - Sexual preference tests - USV emission test * | Approach behavior | Male sexual incentive value. Endocrine and experiential regulation. Importance of genitosensory stimulation. Sexual motivation circuitry. for revision see [1,4,44,46,54,64,65] |
| Conditioned tests | ||
| - Conditioned place preference - Operant conditioning | Reinforced behaviors | |
*: this test has been proposed as a model to evaluate the sexual motivation of females (McGinnis et al., 2003, [74]); however, its validity has yet to be fully tested.
4. Adolescence: The Developmental Period When the Female’s Sexual Behavior Emerges
Adolescence represents a transition period between infancy and adulthood, which is characterized by changes in cognitive functions as well as motivational and affective processes that lead to the behavioral profile typical of adulthood [70,75]. In the rat, this period can be delimited between days 21 and 60 of postnatal life and further subdivided into early, middle, and late adolescence according to developmental milestones [67]. Adolescent rats, like other mammals, exhibit increased risk-taking behaviors, novelty seeking, and exploration as well as high expression of peer social interactions (for revision, see [70,76,77]). This unique behavioral profile is the consequence of the particular state of maturation of different neural systems. For example, the mesocorticolimbic dopamine system is still maturing during adolescence [78,79,80], and the medial prefrontal cortex undergoes a major reorganization, which includes its connectivity with subcortical regions [69]. While some of these maturation changes are independent of sexual steroids, others are modulated by these hormones, which thus influence the development of individuals in a way that Cherly Sisk defined as “…second wave of steroid-dependent neural organization during puberty and adolescence…” [75].
Regarding reproductive maturation, female sexual behavior begins expression during middle adolescence, after the activation of the hypothalamus–pituitary–gonadal (HPG) axis [67,68,81], which is manifested externally by vaginal opening occurring around postnatal days 34 to 39. Adolescent females can display lordosis the night of vaginal opening [82]; however, this response has been more often reported some days later, between postnatal days 40 to 45 [68,83,84]. Mating during this period is efficient, as adolescent rats can become pregnant and give birth to a viable litter [82,85].
Although adolescent rats are able to express sexual behavior, studies on prepubertally ovariectomized females suggest that this behavior is not fully developed at middle adolescence. When estradiol and progesterone are periodically administered to these females, a progressive increase in the display of proceptive and receptive components is observed after subsequent hormonal administrations until the sixth exogenously induced cycle, when the expression stabilizes [86]. Moreover, the dose of steroids necessary to achieve maximal proceptivity in adolescents [86] is higher than that required in adults [87]. These studies, together with that of Sodersten (1975), indicate that prepubertally ovariectomized rats have reduced sensitivity to the effects of estradiol and progesterone and that these effects are enhanced by repetitive exposure [68].
The study of the maturation of female rat sexual behavior throughout adolescence, to the best of my knowledge, did not advance much from these initial studies. Recently, a study from our laboratory found that naturally cycling mid-adolescent rats (30–43 postnatal days) show similar expression of proceptive behaviors and maximal lordosis quotient when compared to that displayed by late-adolescent (59–53 postnatal days) and adult (90–110 postnatal days) females [88]. However, this adult-like sexual response of mid-adolescent rats, unlike adult females, was accompanied by play behavior and high social exploration. The expression of these typical adolescent behaviors [70,77] concomitant with sexual responses results in a unique behavioral profile of young adolescent females during copulation and suggests high social motivation.
Despite the lack of differences in proceptive responses and lordosis quotient between these age-groups, the intensity of the lordosis response (how pronounced the dorsiflexion is [89]) was reduced in mid-adolescent females [88]. As this variable is sensitive to hormonal treatments that modify the sexual motivation of females [89,90,91,92], we evaluated the incentive value of the male for adolescent and adult rats in two motivational tests: a male vs. female preference task [45] (Figure 2A) and 50 kHz USV emission after a brief sexual vs. social interaction [74] (Figure 2B). Interestingly, mid-adolescent rats exhibited a reduced preference for the male compared to adults, and unlike females in later developmental stages, did not perform a greater number of attempts to access the male in this test [45,88]. In addition, these young sexually active females did not increase the emission of 50 kHz USV after a brief interaction with a male relative to an interaction with a female, as late-adolescent and adult rats did [88]. These results indicate that sexual motivation at mid-adolescence is low and increases in late adolescence to the same levels observed in adulthood.
The increment in the incentive value of the male throughout adolescence seems to be selective for sexual—rather than social—motivation because results from our laboratory and those of others indicate that social motivation is high in middle adolescence and decreases towards adulthood [77,88,93]. Thus, an opposite time pattern of sexual and socialmotivations develops throughout adolescence, with late-adolescent females exhibiting high levels of both sexual and social responses [88]. The high social interest of middle-and late-adolescent rats opens the possibility that the incentive value of a male partially depends on social motivation during this developmental period. Similarly, a recent study in humans indicates that the sexual pleasure reported by adolescents between ages 14 and 17 years is associated to socioemotional intimacy and desire rather than to specific sexual behaviors and orgasm experienced. The authors of this study proposed that for adolescents, relational intimacy and desire are primary motives for engaging in partnered sexual experiences [94]. This possible “intrusion” of social motivation into sexual experience in both rat and human adolescents reinforces the importance of delving deeper into the factors that modulate the expression of sexual motivation in adolescent rats as well as the associated affective response. What role does social motivation play in the approach behavior of sexually active adolescent rats to a male? Do they prefer to interact with a male over a peer? How reinforcing is sexual interaction relative to a social one for adolescent females? These questions call for further study of female sexual behavior during this period of development.
It is to be expected and is supported by the literature that the increment in sexual steroid levels that occurs during puberty plays a central role in the maturation of female sexual behavior [75,95,96,97,98]. In this line of thought, modifying the endocrine profile to which female rats are exposed during adolescence and early adulthood changes their sexual preference when adults [99]. Moreover, as previously mentioned, a gradual increase in the sensitivity of the nervous system to the sexual behavior-inducing effect of estrogen seems to occur throughout adolescence [68,86]. Based on this idea and our behavioral results, we are currently exploring the role of repeated exposure to estrogen, which naturally occurs after the activation of the HPG axis, as part of the neuroendocrine mechanisms that mediate sexual motivation increment.
In summary, although female rats can display sexual behavior early after vaginal opening, the maturity of the full sexual behavior repertoire, including sexual motivation, is not complete [68,86,88,100]. The intermediate behavioral profile of late-adolescent females in sexual and social interactions [88] suggests that this maturation develops gradually; therefore, adolescence should be studied at different timepoints for the full comprehension of the dynamic nature of behavioral development. Changes in sexual motivation throughout adolescence are likely to be mediated by the effect of gonadal steroids in sexual circuitry; however, the involvement of non-hormone-dependent maturation processes (as have been reported in the mesolimbic dopaminergic system [101]) as well as the interaction between hormone-dependent and -independent mechanisms cannot be ruled out. We know much about the neuroendocrine regulation of sexual behavior in the adult female; yet, much remains to be understood about how the expression of this behavior matures. Moreover, both middle and late adolescence represent unique periods of the female life cycle for studying how social and sexual motivations interact to shape the behavioral output of individuals when both motivations are high.
5. Postpartum Estrus: A Unique Reproductive Period When Sexual and Maternal Motivations Converge
Female rats, like many other mammals, have the capacity to conceive rapidly after delivering their young due to the existence of a postpartum estrus (PPE) within the first 24 h following parturition [71,72,102]. This is a common physiological phenomenon since all, or almost all, female rats present it ([71,103,104], observations from our lab). If a female becomes pregnant during the PPE, she will gestate one litter while nursing another, and after the second parturition, if the older litter remains in the nest, she will simultaneously raise successive litters with different physiological demands and behavioral capabilities. Raising overlapping litters has been extensively studied by Natalia Uriarte’s laboratory as a model of maternal flexibility in the rat [105,106,107,108].
The regulation of the timing of the behavioral estrus and ovulation after parturition differs from its regulation during estrus cycle [104,109,110,111]. For example, the luteinizing hormone (LH) preovulatory peak has been reported to occur earlier and culminate later in postpartum when compared to proestrus [112]. Moreover, the time of LH surge in the postpartum has been proposed to depend on the vaginocervical stimulation that occurs during labor [109,113]. Accordingly, the timing of PPE depends on circadian signals, as has been established in cycling females [114], but is also determined by signals related to parturition. Thus, the time of delivery is the best predictor of behavioral estrus in postpartum rats [71,72,104,115]. When tested in a sexual arena with a male, females that give birth early in the light phase begin to display lordosis responses approximately 6 h after parturition and exhibit the highest sexual receptivity between 9 and 15 h. Although the maximal expression of proceptive and receptive components as well as minimal rejection of the male occurs around 12 h after delivery, females may show lordosis in response to mounting between 6 to 18 h postpartum [71,115]. In accordance, studies in semi-natural environments show that most PPE rats leave the nest and mate approximately 10 h after parturition [72,73,116]. However, if females deliver close to the onset of the dark phase, then PPE is delayed to the second postpartum day, stressing the interaction between circadian and delivery-related signals in its control [103,113]. To the best of my knowledge, the neuroendocrine mechanisms that trigger PPE in the female rat, including the dual dependence on parturition and circadian signals, are still unclear.
The general expression of sexual behavior of PPE rats as well as the structure and efficiency of mating do not differ from those observed in the sexual interaction of late proestrus cycling females. Copulatory time in semi-natural environments, however, has been reported to be reduced during PPE either by decreasing the number of intromissions [72] or by reducing inter-intromission intervals [116]. The similarity in the behavioral profile of PPE and cycling females may respond to similar changes in sexual steroids levels—estrogen peak followed by progesterone peak—that precede both behavioral estrus. Thus, Connors and Davis (1980) reported that estrogen plasma levels that increase during parturition [117] decrease after it (although Carrillo-Martínez et al., 2011, reported a slight augment at 9 h postpartum [115]), while progesterone levels peak at 9 to 12 h postpartum, coinciding with the period of maximal sexual activity [104]. In accordance, Sodersten and Eneroth (1981) found a close relationship between the preovulatory peak of progesterone and the onset of sexual receptivity in cycling females [118]. In addition, the action of progesterone on its intracellular receptor is necessary for the expression of PPE sexual behavior [115,119], which is in line with what has been observed in cycling rats [120].
The connection between the time of parturition, when maternal behavior is first displayed, and the time of behavioral estrus reflects the coordination of these two main reproductive behaviors. Like sexual behavior, maternal behavior is highly motivated, as maternal females spend time and make effort to get access to and interact with pups (for revision, see [121,122]). Thus, lactating rats exhibit CPP to a context paired with pups [123] and learn to bar press in order to gain access to them [124]. Remarkably, during PPE, as females are sexually and maternally motivated, both the male and the pups acquire incentive value simultaneously, converting this period into an excellent model for exploring how these two strong social motivations interact. This interaction was firstly studied by Gilbert and colleagues, who observed that when parturient females are housed with a male in large semi-natural enclosures, they will exhibit both mating and maternal care during PPE but in separated locations and alternating between these behaviors [72]. During the sexual interaction, PPE rats spent little time with their litters and do so mainly in the post-ejaculatory intervals even if the nest area is disturbed in an attempt to promote their return to the nest [73]. This means that once sexual interaction has begun, an ejaculatory series will proceed until its conclusion before the female returns to take care of her litter. Therefore, under semi-natural conditions, it seems that sexual behavior dominates maternal care.
Interestingly, sexual interaction in a semi-natural enclosure is not accompanied by female aggressive responses [72], whereas PPE rats removed from the home cage and immediately tested with a male always show rejection even during the period of maximal sexual activity [71]. Considering the importance of the context in which mating occurs for the female [19,125], we explored how PPE rats respond to a male in their home cages when pups are present. This question was particularly intriguing for us considering that postpartum rats display intense maternal aggression toward an intruder in their home cages, especially during the first postpartum week [126,127,128]. While sexually active females during late proestrus display active sexual solicitations to the male, lactating rats (tested out of the PPE period) show an intense agonistic response towards a male while in their nest; these opposite responses evoke from the male either approaches and mounts [5,34] or escapes and submissions [128]. Astonishingly, when PPE rats were simultaneously exposed to their pups and a male in their home cages, they exhibited a merge of sexual and aggressive responses towards males with rapid transitions between hops and darts and aggressive components. This result indicates that the male can be perceived as an ambivalent stimulus for these females, who consequently will co-express both behaviors [129]. Moreover, the latencies to display sexual solicitation or aggressive response as well as their intensities did not differ when PPE females were co-expressing both behaviors in their home cages from those exhibited when sexual behavior and maternal aggression were independently expressed either by placing PPE rats with a male in a new cage or by placing a female intruder in their home cages [129]. The high levels of sexual behavior and maternal aggression displayed when co-expressed indicate the activation of both motivational systems without mutual inhibition. Even when maternal behavior was stimulated, the observed increment of aggression toward the male did not affect the display of hops and darts, suggesting that, at least partially, there is an independent regulation of both motivations when co-activated [129]. Interestingly, males copulating with females in PPE in their home cages did not modify their ejaculation latency (approximately 8–10 min employing vigorous males) but did increase their latency to mounting and reduced the number of mounts required to ejaculate [129]. This increment in copulation efficiency could represent a strategy used by males to cope with the aggression displayed by these sexually active mothers.
If the PPE female co-expresses sexual behavior and maternal aggression toward the male in her home cage with her pups, do these two social stimuli—male and pups—have the same incentive value for her? To answer this question, we assessed the relative incentive value of a male and pups for PPE rats in a male vs. pups preference task (same model presented in Figure 2A) and compared it to that of sexually active cycling mothers who did not undergo the endocrine changes of gestation (and therefore exhibit deficient maternal behavior [130]) and to that of PPE rats with previous reproductive experience (who have stronger maternal motivation [131]). While maternal virgin rats in late proestrus chose the male, PPE females preferred the pups, and this preference was stronger in multiparous rats [49,132]. Furthermore, if the maternal motivation of primiparous PPE rats was reduced (by limiting the time of interaction between mothers and their litters), they showed a strong preference for the male as opposed to the clear preference for the pups of undisturbed PPE females [132]. Together, these results indicate that the incentive value of pups, which is modulated by the endocrine changes of gestation and parturition and by previous and recent maternal experiences, influences the value of the male for sexually active mothers. We do not yet know whether increasing the incentive value of a male affects or modifies the strong value of pups for PPE rats.
Partial overlap of the neural circuits that regulate sexual and maternal behaviors have been proposed [62,63], as active components of both affiliative behaviors are regulated by the mPOA and dopamine neurotransmission in it (for sexual behavior, see [133,134,135], and for maternal behavior, see [136,137]) and by the interaction of this area with the mesolimbic dopamine system (for sexual, see [62,65] and, on the other hand [66], and for maternal, see [138,139]). As a system that promotes behavioral activation, the mesolimbic dopaminergic system has been implicated in mediating effort-related decision-making between two reinforcers [140,141,142]. In our preference task between two social reinforcers without physical access to them, PPE females not only spend more time with the pups but also make more attempts to gain access to them, which is an effort-related response [49,132]. Interestingly, antagonizing dopaminergic neurotransmission diminishes the preference for the pups over a male of PPE rats but does not affect the preference for the pups over a neutral social stimulus and for the male over a neutral social stimulus [143]. This work points to a role for dopaminergic activity in assigning greater incentive value to the pups relative to a male when both reinforcers are inaccessible and compete with each other. We are currently further exploring the role of the dopaminergic system in the interaction between these two social motivations during the PPE.
The evidence obtained so far reveals that the response of the PPE female to a sexually active male drastically changes according to the situation. She courts and attacks the male at the same time when she is in her home cage with the pups [129]; however, she courts—without aggression—when able to copulate far from the nest [72] or in a new environment [129,132], and she even ignores the male and chooses the pups in a preference test without physical access to them [132,143]. In conclusion, the incentive value of the male and the pups for the PPE rat influence one another and change depending upon the context, broadening her spectrum of possible behavioral outcomes. In addition, the male vs. pups preference task was sensitive enough to detect motivational differences between primiparous and multiparous PPE rats and between postparturient females with limited and continuous mother–infant interaction, which did not emerge in the maternal behavior home cage test [132]. This study highlights the importance of confronting motivations for their better understanding and advocates for the use of new and challenging models to unveil the complexity of motivated behaviors.
6. Conclusions and Final Considerations
The study of sexual behavior and motivation in the female rat has lagged in comparison to that of the male rat; however, the development of new and more complex models established female rats’ active role in mating as well as the strong motivational basis of this behavior (Table 1). This review does not pretend to be exhaustive since sexual behavior in the female rat is studied in multiple models, contexts, and physiological conditions in addition to the ones mentioned above (e.g., [50,144,145,146,147]) but rather aims to introduce two life periods that have been understudied and that possess the potential to allow us to further deepen our understanding of this behavior.
The naturally cycling adolescent female offers the opportunity to characterize the neuroendocrine mechanisms responsible for the gradual maturation of female’s sexual repertoire. Furthermore, the development of sexual behavior during adolescence, a period of great neural plasticity, constitutes an excellent model for assessing the organizational effects of sexual steroids on the central nervous system beyond the perinatal period. Following another line of analysis, studying the interaction of social and sexual motivations in adolescent proestrous females may help us to understand the contribution of high social motivation, which is typical of this developmental period, in their response to males. Similarly, PPE generates a unique physiological context to confront two of the most important social reinforcers in a female’s life: her offspring and a male. Female rat behavior in these two life-history contexts makes us realize that an individual’s motivation should not be understood as a static state associated with an internal condition but rather as a dynamic state that changes according to the context and the coexistence of other motivations. The unique developmental and physiological characteristics of adolescence and postpartum periods open the door to study the neuroendocrine and contextual regulation of sexual behavior outside the classically studied periods.
Funding
This work was supported by CSIC I+D grant 2020 to D.A. and by PEDECIBA, SNI-ANII, and DT CSIC-UdelaR funding to D.A.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to thank Natalia Uriarte for her thoughtful and careful reading of the manuscript, and Elisa Hernandez for the beautiful drawings. I am deeply grateful to Annabel Ferreira for encouraging me to question current paradigms.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ventura-Aquino, E.; Paredes, R.G. Sexual behavior in rodents: Where do we go from here? Horm. Behav. 2020, 118, 104678. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G.; Kippin, T.E.; Coria-Avila, G. What Can Animal Models Tell Us about Human Sexual Response? Annu. Rev. Sex Res. 2003, 14, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Le Moëne, O.; Ågmo, A. Modeling Human Sexual Motivation in Rodents: Some Caveats. Front. Behav. Neurosci. 2019, 13, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarraci, F.A.; Frohardt, R.J. “What a Girl Wants”: What Can We Learn From Animal Models of Female Sexual Motivation? Front. Behav. Neurosci. 2019, 13, 216. [Google Scholar] [CrossRef]
- Beach, F.A. Sexual attractivity, proceptivity, and receptivity in female mammals. Horm. Behav. 1976, 7, 105–138. [Google Scholar] [CrossRef]
- Blaustein, J.D.; Erskine, M.S. 2-Feminine Sexual Behavior: Cellular Integration of Hormonal and Afferent Information in the Rodent Forebrain. In Hormones, Brain and Behavior; Pfaff, D.W., Arnold, A.P., Fahrbach, S.E., Etgen, A.M., Rubin, R.T., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 139–214. [Google Scholar]
- Pfaff Donald, W. Luteinizing Hormone-Releasing Factor Potentiates Lordosis Behavior in Hypophysectomized Ovariectomized Female Rats. Science 1973, 182, 1148–1149. [Google Scholar] [CrossRef]
- Kow, L.-M.; Pfaff, D.W. Sensory requirements for the lordosis reflex in female rats. Brain Res. 1976, 101, 47–66. [Google Scholar] [CrossRef]
- Pfaff, D.; Montgomery, M.; Lewis, C. Somatosensory determinants of lordosis in female rats: Behavioral definition of the estrogen effect. J. Comp. Physiol. Psychol. 1977, 91, 134–145. [Google Scholar] [CrossRef]
- Pfaff, D.W.; Kow, L.M.; Loose, M.D.; Flanagan-Cato, L.M. Reverse engineering the lordosis behavior circuit. Horm. Behav. 2008, 54, 347–354. [Google Scholar] [CrossRef]
- Kow, L.-M.; Pfaff, D.W. The membrane actions of estrogens can potentiate their lordosis behavior-facilitating genomic actions. Proc. Natl. Acad. Sci. USA 2004, 101, 12354. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, D.W. Estrogens and brain function; Springer: Berlin/Heidelberg, Germany, 1979. [Google Scholar]
- Pfaff, D.W.; Vasudevan, N.; Kia, H.K.; Zhu, Y.-S.; Chan, J.; Garey, J.; Morgan, M.; Ogawa, S. Estrogens, brain and behavior: Studies in fundamental neurobiology and observations related to women’s health. J. Steroid Biochem. Mol. Biol. 2000, 74, 365–373. [Google Scholar] [CrossRef]
- Micevych, P.E.; Meisel, R.L. Integrating neural circuits controlling female sexual behavior. Front. Syst. Neurosci. 2017, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G.; Jones, S.L.; Flanagan-Cato, L.M.; Blaustein, J.D. Female sexual behavior. Knobil Neill’s Physiol. Reprod. 2015, 2, 2287–2370. [Google Scholar]
- Pfaff, D. Cellular and molecular mechanisms of female rproductive behaviors. Physiol. Reprod. 1994, 2, 107–220. [Google Scholar]
- Ventura-Aquino, E.; Paredes, R.G. Animal Models in Sexual Medicine: The Need and Importance of Studying Sexual Motivation. Sex. Med. Rev. 2017, 5, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Peirce, J.T.; Nuttall, D.L. Self-paced sexual behavior in the female rat. J. Comp. Physiol. Psychol. 1961, 54, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Erskine, M.S. Solicitation behavior in the estrous female rat: A review. Horm. Behav. 1989, 23, 473–502. [Google Scholar] [CrossRef]
- Coopersmith, C.; Candurra, C.; Erskine, M.S. Effects of paced mating and intromissive stimulation on feminine sexual behavior and estrus termination in the cycling rat. J. Comp. Physiol. Psychol. 1996, 110, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Erskine, M.S. Pelvic and pudendal nerves influence the display of paced mating behavior in response to estrogen and progesterone in the female rat. Behav. Neurosci. 1992, 106, 690–697. [Google Scholar] [CrossRef]
- Martínez, I.; Paredes, R.G. Only Self-Paced Mating Is Rewarding in Rats of Both Sexes. Horm. Behav. 2001, 40, 510–517. [Google Scholar] [CrossRef]
- Paredes, R.G.; Alonso, A. Sexual behavior regulated (paced) by the female induces conditioned place preference. Behav. Neurosci. 1997, 111, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Coopersmith, C.; Erskine, M.S. Influence of paced mating and number of intromissions on fertility in the laboratory rat. Reproduction 1994, 102, 451–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClintock, M.K.; Adler, N.T. The Role of the Female During Copulation in Wild and Domestic Norway Rats (Rattus Norvegicus). Behaviour 1978, 67, 67–95. [Google Scholar] [CrossRef]
- McClintock, M.K.; Anisko, J.J. Group mating among Norway rats I. Sex differences in the pattern and neuroendocrine consequences of copulation. Anim. Behav. 1982, 30, 398–409. [Google Scholar] [CrossRef]
- McClintock, M.K.; Anisko, J.J.; Adler, N.T. Group mating among Norway rats II. The social dynamics of copulation: Competition, cooperation, and mate choice. Anim. Behav. 1982, 30, 410–425. [Google Scholar] [CrossRef]
- McClintock, M.K. Group mating in the domestic rat as a context for sexual selection: Consequences for the analysis of sexual behavior and neuroendocrine responses. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 1984; Volume 14, pp. 1–50. [Google Scholar]
- Bergheim, D.; Chu, X.; Ågmo, A. The function and meaning of female rat paracopulatory (proceptive) behaviors. Behav. Process. 2015, 118, 34–41. [Google Scholar] [CrossRef]
- Chu, X.; Ågmo, A. Sociosexual behaviours in cycling, intact female rats (Rattus norvegicus) housed in a seminatural environment. Behaviour 2014, 151, 1143–1184. [Google Scholar] [CrossRef]
- Chu, X.; Ågmo, A. Sociosexual behaviors during the transition from non-receptivity to receptivity in rats housed in a seminatural environment. Behav. Process. 2015, 113, 24–34. [Google Scholar] [CrossRef]
- Chu, X.; Snoeren, E.; Ågmo, A. Functions of vocalization in sociosexual behaviors in rats (Rattus norvegicus) in a seminatural environment. J. Comp. Psychol. 2017, 131, 10. [Google Scholar] [CrossRef] [Green Version]
- Le Moëne, O.; Hernández-Arteaga, E.; Chu, X.; Ågmo, A. Rapid changes in sociosexual behaviors around transition to and from behavioral estrus, in female rats housed in a seminatural environment. Behav. Process. 2020, 174, 104101. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Smith, W.J.; Coopersmith, C.B. Appetitive and Consummatory Sexual Behaviors of Female Rats in Bilevel Chambers: I. A Correlational and Factor Analysis and the Effects of Ovarian Hormones. Horm. Behav. 1999, 35, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G.; Mendelson, S.D.; Phillips, A.G. A correlational and factor analysis of anticipatory and consummatory measures of sexual behavior in the male rat. Psychoneuroendocrinology 1990, 15, 329–340. [Google Scholar] [CrossRef]
- Mendelson, S.D.; Gorzalka, B.B. An improved chamber for the observation and analysis of the sexual behavior of the female rat. Physiol. Behav. 1987, 39, 67–71. [Google Scholar] [CrossRef]
- Ferreira-Nuño, A.; Morales-Otal, A.; Paredes, R.G.; Velázquez-Moctezuma, J. Sexual behavior of female rats in a multiple-partner preference test. Horm. Behav. 2005, 47, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.L.; Diehl, A.; Joyce, E.; Cohn, J.; Lopez, J.; Guarraci, F.A. “Some guys have all the luck”: Mate preference influences paced-mating behavior in female rats. Physiol. Behav. 2007, 90, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Winland, C.; Bolton, J.L.; Ford, B.; Jampana, S.; Tinker, J.; Frohardt, R.J.; Guarraci, F.A.; Zewail-Foote, M. “Nice guys finish last”: Influence of mate choice on reproductive success in Long–Evans rats. Physiol. Behav. 2012, 105, 868–876. [Google Scholar] [CrossRef]
- Zewail-Foote, M.; Diehl, A.; Benson, A.; Lee, K.H.; Guarraci, F.A. Reproductive success and mate choice in Long–Evans rats. Physiol. Behav. 2009, 96, 98–103. [Google Scholar] [CrossRef]
- Berridge, K.C. Motivation concepts in behavioral neuroscience. Physiol. Behav. 2004, 81, 179–209. [Google Scholar] [CrossRef]
- Bermant, G. Response Latencies of Female Rats during Sexual Intercourse. Science 1961, 133, 1771–1773. [Google Scholar] [CrossRef]
- Uphouse, L.; Pinkston, J.; Baade, D.; Solano, C.; Onaiwu, B. Use of an operant paradigm for the study of antidepressant-induced sexual dysfunction. Behav. Pharmacol. 2015, 26, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Ågmo, A. Unconditioned sexual incentive motivation in the male Norway Rat (Rattus norvegicus). J. Comp. Psychol. 2003, 117, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Agrati, D.; Machado, L.; Delgado, H.; Uriarte, N.; Zuluaga, M.J.; Ferreira, A. Sexual behaviour of the female rat during late adolescence: Effect of chronic cocaine treatment. Behav. Pharmacol. 2019, 30, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.G. Evaluating the neurobiology of sexual reward. ILAR J. 2009, 50, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuszczyk, J.V.; Larsson, K. Role of androgen, estrogen and sexual experience on the female rat’s partner preference. Physiol. Behav. 1991, 50, 139–142. [Google Scholar] [CrossRef]
- Clark, A.S.; Kelton, M.C.; Guarraci, F.A.; Clyons, E.Q. Hormonal status and test condition, but not sexual experience, modulate partner preference in female rats. Horm. Behav. 2004, 45, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Agrati, D.; Fernández-Guasti, A.; Ferreira, A. The reproductive stage and experience of sexually receptive mothers alter their preference for pups or males. Behav. Neurosci. 2008, 122, 998. [Google Scholar] [CrossRef]
- Guarraci, F.A.; Gonzalez, C.M.F.; Lucero, D.; Davis, L.K.; Meerts, S.H. Sexual Behavior is Enhanced by Regular, Repeated Mating from Young Adulthood to Middle Age in Female Long-Evans Rats. Curr. Aging Sci. 2020, 13, 169–177. [Google Scholar] [CrossRef]
- Paredes-Ramos, P.; Miquel, M.; Manzo, J.; Pfaus, J.G.; López-Meraz, M.L.; Coria-Avila, G.A. Tickling in juvenile but not adult female rats conditions sexual partner preference. Physiol. Behav. 2012, 107, 17–25. [Google Scholar] [CrossRef]
- Meerts, S.H.; Clark, A.S. Artificial vaginocervical stimulation induces a conditioned place preference in female rats. Horm. Behav. 2009, 55, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Parada, M.; Chamas, L.; Censi, S.; Coria-Avila, G.; Pfaus, J.G. Clitoral stimulation induces conditioned place preference and Fos activation in the rat. Horm. Behav. 2010, 57, 112–118. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Scardochio, T.; Parada, M.; Gerson, C.; Quintana, G.R.; Coria-Avila, G.A. Do rats have orgasms? Socioaffect. Neurosci. Psychol. 2016, 6, 31883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemens, L.G.; Yang, L.-Y. MPOA lesions affect female pacing of copulation in rats. Behav. Neurosci. 2000, 114, 1191. [Google Scholar] [CrossRef]
- Xiao, K.; Kondo, Y.; Sakuma, Y. Differential regulation of female rat olfactory preference and copulatory pacing by the lateral septum and medial preoptic area. Neuroendocrinology 2005, 81, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Sakuma, Y. The Medial Amygdala Controls Coital Access of Female Rats: A Possible Involvement of Emotional Responsiveness. Jpn. J. Physiol. 2005, 55, 0601160021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarraci, F.A.; Megroz, A.B.; Clark, A.S. Paced mating behavior in the female rat following lesions of three regions responsive to vaginocervical stimulation. Brain Res. 2004, 999, 40–52. [Google Scholar] [CrossRef]
- Clark, A.S.; Pfeifle, J.K.; Edwards, D.A. Ventromedial hypothalamic damage and sexual proceptivity in female rats. Physiol. Behav. 1981, 27, 597–602. [Google Scholar] [CrossRef]
- Emery, D.E.; Moss, R.L. Lesions confined to the ventromedial hypothalamus decrease the frequency of coital contacts in female rats. Horm. Behav. 1984, 18, 313–329. [Google Scholar] [CrossRef]
- García-Horsman, S.P.; Ǻgmo, A.; Paredes, R.G. Infusions of naloxone into the medial preoptic area, ventromedial nucleus of the hypothalamus, and amygdala block conditioned place preference induced by paced mating behavior. Horm. Behav. 2008, 54, 709–716. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Manzo, J.; Garcia, L.I.; Carrillo, P.; Miquel, M.; Pfaus, J.G. Neurobiology of social attachments. Neurosci. Biobehav. Rev. 2014, 43, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Stolzenberg, D.S.; Numan, M. Hypothalamic interaction with the mesolimbic DA system in the control of the maternal and sexual behaviors in rats. Neurosci. Biobehav. Rev. 2011, 35, 826–847. [Google Scholar] [CrossRef]
- Ventura-Aquino, E.; Portillo, W.; Paredes, R.G. Sexual motivation: A comparative approach in vertebrate species. Curr. Sex. Health Rep. 2018, 10, 114–123. [Google Scholar] [CrossRef]
- Pfaus, J.G. REVIEWS: Pathways of Sexual Desire. J. Sex. Med. 2009, 6, 1506–1533. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.G.; Ågmo, A. Has dopamine a physiological role in the control of sexual behavior?: A critical review of the evidence. Prog. Neurobiol. 2004, 73, 179–225. [Google Scholar] [CrossRef] [PubMed]
- Holder, M.K.; Blaustein, J.D. Puberty and adolescence as a time of vulnerability to stressors that alter neurobehavioral processes. Front. Neuroendocrinol. 2014, 35, 89–110. [Google Scholar] [CrossRef] [Green Version]
- Södersten, P. Receptive behavior in developing female rats. Horm. Behav. 1975, 6, 307–317. [Google Scholar] [CrossRef]
- Brenhouse, H.C.; Andersen, S.L. Developmental trajectories during adolescence in males and females: A cross-species understanding of underlying brain changes. Neurosci. Biobehav. Rev. 2011, 35, 1687–1703. [Google Scholar] [CrossRef] [Green Version]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- Connor, J.R.; Davis, H.N. Postpartum Estrus in Norway Rats. I. Behavior. Biol. Reprod. 1980, 23, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, A.N.; Pelchat, R.J.; Adler, N.T. Postpartum copulatory and maternal behaviour in Norway rats under seminatural conditions. Anim. Behav. 1980, 28, 989–995. [Google Scholar] [CrossRef]
- Gilbert, A.N.; Pelchat, R.J.; Adler, N.T. Sexual and maternal behaviour at the postpartum oestrus: The role of experience in time-sharing. Anim. Behav. 1984, 32, 1045–1053. [Google Scholar] [CrossRef]
- McGinnis, M.Y.; Vakulenko, M. Characterization of 50-kHz ultrasonic vocalizations in male and female rats. Physiol. Behav. 2003, 80, 81–88. [Google Scholar] [CrossRef]
- Schulz, K.M.; Sisk, C.L. The organizing actions of adolescent gonadal steroid hormones on brain and behavioral development. Neurosci. Biobehav. Rev. 2016, 70, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, M.; Romeo, R.D.; Andersen, S.L. Neurobiology of the development of motivated behaviors in adolescence: A window into a neural systems model. Pharmacol. Biochem. Behav. 2009, 93, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. The ontogeny of play in rats. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 1981, 14, 327–332. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Varlinskaya, E.I.; Spear, L.P. Motivational systems in adolescence: Possible implications for age differences in substance abuse and other risk-taking behaviors. Brain Cogn. 2010, 72, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Simon, N.W.; Moghaddam, B. Neural processing of reward in adolescent rodents. Dev. Cogn. Neurosci. 2015, 11, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Teicher, M.H.; Andersen, S.L.; Polcari, A.; Anderson, C.M.; Navalta, C.P. Developmental neurobiology of childhood stress and trauma. Psychiatr. Clin. 2002, 25, 397–426. [Google Scholar] [CrossRef]
- Sisk, C.L.; Foster, D.L. The neural basis of puberty and adolescence. Nat. Neurosci. 2004, 7, 1040–1047. [Google Scholar] [CrossRef]
- Hashizume, K.; Ōhashi, K. Timing of sexual receptivity and the release of gonadotrophins during puberty in female rats. Reproduction 1984, 72, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S. Mounting behavior and receptive behavior in developing female rats and the effect of social isolation. Physiol. Behav. 1977, 19, 749–752. [Google Scholar] [CrossRef]
- Meaney, M.J.; Stewart, J. A descriptive study of social development in the rat (Rattus norvegicus). Anim. Behav. 1981, 29, 34–45. [Google Scholar] [CrossRef]
- Hashizume, K.; Ohashi, K.; Hamajima, F. Adolescent pregnancy and growth of progeny in rats. Physiol. Behav. 1991, 49, 367–371. [Google Scholar] [CrossRef]
- Hliňák, Z. Estradiol plus progesterone treatment and precopulatory behavior in prepubertally ovariectomized female rats: Dose-response relationships. Horm. Behav. 1986, 20, 263–269. [Google Scholar] [CrossRef]
- Hlinák, Z.; Madlafousek, J. Importance of Female’s Precopulatory Behaviour for the Primary Initiation of Male’s Copulatory Behaviour in the Laboratory Rat. Behaviour 1983, 86, 237–248. [Google Scholar] [CrossRef]
- Armas, M.; Marín, G.; Uriarte, N.; Agrati, D. Increase in sexual motivation throughout adolescence in the cycling female rat. Dev. Psychobiol. 2021, 63, e22162. [Google Scholar] [CrossRef]
- Hardy, D.F.; DeBold, J.F. Effects of coital stimulation upon behavior of the female rat. J. Comp. Physiol. Psychol. 1972, 78, 400–408. [Google Scholar] [CrossRef]
- Sánchez Montoya, E.L.; Hernández, L.; Barreto-Estrada, J.L.; Ortiz, J.G.; Jorge, J.C. The Testosterone Metabolite 3α-Diol Enhances Female Rat Sexual Motivation When Infused in the Nucleus Accumbens Shell. J. Sex. Med. 2010, 7, 3598–3609. [Google Scholar] [CrossRef] [Green Version]
- Hardy, D.F.; DeBold, J.F. Effects of repeated testing on sexual behavior of the female rat. J. Comp. Physiol. Psychol. 1973, 85, 195. [Google Scholar] [CrossRef]
- Holder, M.K.; Mong, J.A. Methamphetamine enhances paced mating behaviors and neuroplasticity in the medial amygdala of female rats. Horm. Behav. 2010, 58, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Douglas, L.A.; Varlinskaya, E.I.; Spear, L.P. Rewarding properties of social interactions in adolescent and adult male and female rats: Impact of social versus isolate housing of subjects and partners. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2004, 45, 153–162. [Google Scholar] [CrossRef]
- Beckmeyer, J.J.; Herbenick, D.; Fu, T.-C.; Dodge, B.; Fortenberry, J.D. Pleasure During Adolescents’ Most Recent Partnered Sexual Experience: Findings from a U.S. Probability Survey. Arch. Sex. Behav. 2021, 50, 2423–2434. [Google Scholar] [CrossRef] [PubMed]
- Olster, D.H.; Blaustein, J.D. Development of progesterone-facilitated lordosis in female guinea pigs: Relationship to neural estrogen and progestin receptors. Brain Res. 1989, 484, 168–176. [Google Scholar] [CrossRef]
- Bakker, J.; Honda, S.-I.; Harada, N.; Balthazart, J. The Aromatase Knock-Out Mouse Provides New Evidence That Estradiol Is Required during Development in the Female for the Expression of Sociosexual Behaviors in Adulthood. J. Neurosci. 2002, 22, 9104. [Google Scholar] [CrossRef]
- Schulz, K.M.; Sisk, C.L. Pubertal hormones, the adolescent brain, and the maturation of social behaviors: Lessons from the Syrian hamster. Mol. Cell. Endocrinol. 2006, 254–255, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Snoeren, E.; Södersten, P.; Ågmo, A. Sexual incentive motivation and male and female copulatory behavior in female rats given androgen from postnatal day 20. Physiol. Behav. 2021, 237, 113460. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, F.H.; Muntjewerff, J.-W.; Louwerse, A.L.; van de Poll, N.E. Sexual behavior and sexual orientation of the female rat after hormonal treatment during various stages of development. Horm. Behav. 1988, 22, 100–115. [Google Scholar] [CrossRef]
- Hlinák, Z. Oestradiol and progesterone treatment and precopulatory behaviour in female rats ovariectomized at different ages. Physiol. Bohemoslov. 1985, 34, 373–380. [Google Scholar]
- Andersen, S.L.; Thompson, A.P.; Krenzel, E.; Teicher, M.H. Pubertal changes in gonadal hormones do not underlie adolescent dopamine receptor overproduction. Psychoneuroendocrinology 2002, 27, 683–691. [Google Scholar] [CrossRef]
- Dewsbury, D.A. Modes of estrus induction as a factor in studies of the reproductive behavior of rodents. Neurosci. Biobehav. Rev. 1990, 14, 147–155. [Google Scholar] [CrossRef]
- Blandau, R.J.; Soderwall, A.L. Post-parturitional heat and the time of ovulation in the albino rat. Data on parturition. Anat. Rec. 1941, 81, 419–431. [Google Scholar] [CrossRef]
- Connor, J.R.; Davis, H.N. Postpartum Estrus in Norway Rats. II. Physiology. Biol. Reprod. 1980, 23, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Ferreño, M.; Pose, S.; Agrati, D.; Zuluaga, M.J.; Ferreira, A.; Uriarte, N. Incentive value of newborn pups relative to juveniles for mother rats raising overlapping litters. Behav. Process. 2018, 157, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Pose, S.; Zuluaga, M.J.; Ferreño, M.; Agrati, D.; Bedó, G.; Uriarte, N. Raising overlapping litters: Differential activation of rat maternal neural circuitry after interacting with newborn or juvenile pups. J. Neuroendocrinol. 2019, 31, e12701. [Google Scholar] [CrossRef]
- Uriarte, N.; Ferreira, A.; Rosa, X.F.; Lucion, A.B. Effects of litter-overlapping on emotionality, stress response, and reproductive functions in male and female rats. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2009, 51, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, N.; Ferreira, A.; Rosa, X.F.; Sebben, V.; Lucion, A.B. Overlapping litters in rats: Effects on maternal behavior and offspring emotionality. Physiol. Behav. 2008, 93, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- HAszuME, K.; SUGAwARA, S. Studies on the Reproductive Phenomena in the Post-partum Rat II. Ovulating Hormone Release in the Post-partum Ovulation in Rat. Tohoku J. Agric. Res. 1974, 24, 123–127. [Google Scholar]
- Ying, S.-Y.; Gove, S.; Fang, V.S.; Greep, R.O. Ovulation in Postpartum Rats1. Endocrinology 1973, 92, 108–116. [Google Scholar] [CrossRef]
- Takiguchi, S.; Sugino, N.; Esato, K.; Karube-Harada, A.; Sakata, A.; Nakamura, Y.; Ishikawa, H.; Kato, H. Differential regulation of apoptosis in the corpus luteum of pregnancy and newly formed corpus luteum after parturition in rats. Biol. Reprod. 2004, 70, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.C.; Schwartz, N.B. Timing of Post-partum Ovulation in the Rat1. Endocrinology 1965, 76, 620–625. [Google Scholar] [CrossRef]
- Fox, S.R.; Susan Smith, M. Postpartum Preovulatory Surge of Gonadotropin Secretion in the Rat May Be Initiated by the Labor Process1. Biol. Reprod. 1984, 31, 619–626. [Google Scholar] [CrossRef] [Green Version]
- de la Iglesia, H.O.; Schwartz, W.J. Minireview: Timely ovulation: Circadian regulation of the female hypothalamo-pituitary-gonadal axis. Endocrinology 2006, 147, 1148–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrillo-Martínez, G.E.; Gómora-Arrati, P.; González-Arenas, A.; Morimoto, S.; Camacho-Arroyo, I.; González-Flores, O. Role of progesterone receptors during postpartum estrus in rats. Horm. Behav. 2011, 59, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Hedricks, C.; McClintock, M.K. The timing of mating by postpartum estrous rats. Z. Tierpsychol. 1985, 67, 1–16. [Google Scholar] [CrossRef]
- Yoshida, T.; SUZUKI, H.; HATTORI, Y.; NODA, K. Hormonal changes around the parturition in rats. Tohoku J. Exp. Med. 1981, 135, 87–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodersten, P.; Eneroth, P. Serum levels of oestradiol-17β and progesterone in relation to sexual receptivity in intact and ovariectomized rats. J. Endocrinol. 1981, 89, 45–54. [Google Scholar] [CrossRef]
- Carrillo-Martínez, G.E.; Gómora-Arrati, P.; González-Arenas, A.; Roldán-Roldán, G.; González-Flores, O.; Camacho-Arroyo, I. Effects of RU486 in the expression of progesterone receptor isoforms in the hypothalamus and the preoptic area of the rat during postpartum estrus. Neurosci. Lett. 2011, 504, 127–130. [Google Scholar] [CrossRef]
- Telleria, C.M.; Mezzadri, M.R.; Deis, R.P. Fertility impairment after mifepristone treatment to rats at proestrus: Actions on the hypothalamic-hypophyseal-ovarian axis. Contraception 1997, 56, 267–274. [Google Scholar] [CrossRef]
- Ferreira, A.; Agrati, D.; Uriarte, N.; Pereira, M.; Zuluaga, M.J. The rat as a model for studying maternal behavior. In Behavioral Animal Models; Research Signpost: Kerala, India, 2012; pp. 73–88. [Google Scholar]
- Pereira, M.; Ferreira, A. Neuroanatomical and neurochemical basis of parenting: Dynamic coordination of motivational, affective and cognitive processes. Horm. Behav. 2016, 77, 72–85. [Google Scholar] [CrossRef]
- Fleming, A.S.; Korsmit, M.; Deller, M. Rat pups are potent reinforcers to the maternal animal: Effects of experience, parity, hormones, and dopamine function. Psychobiology 1994, 22, 44–53. [Google Scholar] [CrossRef]
- Lee, A.; Clancy, S.; Fleming, A.S. Mother rats bar-press for pups: Effects of lesions of the mpoa and limbic sites on maternal behavior and operant responding for pup-reinforcement. Behav. Brain Res. 1999, 100, 15–31. [Google Scholar] [CrossRef]
- Paredes, R.G.; Vazquez, B. What do female rats like about sex? Paced mating. Behav. Brain Res. 1999, 105, 117–127. [Google Scholar] [CrossRef]
- Erskine, M.S.; Barfield, R.J.; Goldman, B.D. Intraspecific fighting during late pregnancy and lactation in rats and effects of litter removal. Behav. Biol. 1978, 23, 206–218. [Google Scholar] [CrossRef]
- Ferreira, A.; Hansen, S. Sensory control of maternal aggression in Rattus norvegicus. J. Comp. Psychol. 1986, 100, 173. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, R.M.M.; Ferreira, A.; Agrati, D. Sensory, hormonal, and neural basis of maternal aggression in rodents. Neurosci. Aggress. 2014, 17, 111–130. [Google Scholar] [CrossRef]
- Agrati, D.; Fernández-Guasti, A.; Ferreno, M.; Ferreira, A. Coexpression of sexual behavior and maternal aggression: The ambivalence of sexually active mother rats toward male intruders. Behav. Neurosci. 2011, 125, 446. [Google Scholar] [CrossRef]
- Pereira, M.; Ferreira, A. Demanding pups improve maternal behavioral impairments in sensitized and haloperidol-treated lactating female rats. Behav. Brain Res. 2006, 175, 139–148. [Google Scholar] [CrossRef]
- Bridges, R.S. Long-term alterations in neural and endocrine processes induced by motherhood in mammals. Horm. Behav. 2016, 77, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Agrati, D.; Ferreño, M.; Marin, G.; Uriarte, N.; Zuluaga, M.J.; Fernández-Guasti, A.; Ferreira, A. Previous and recent maternal experiences modulate pups’ incentive value relative to a male without affecting maternal behavior in postpartum estrous rats. J. Physiol. 2016, 110, 140–148. [Google Scholar] [CrossRef]
- Graham, M.D.; Pfaus, J.G. Differential regulation of female sexual behaviour by dopamine agonists in the medial preoptic area. Pharmacol. Biochem. Behav. 2010, 97, 284–292. [Google Scholar] [CrossRef]
- Graham, M.D.; Pfaus, J.G. Differential effects of dopamine antagonists infused to the medial preoptic area on the sexual behavior of female rats primed with estrogen and progesterone. Pharmacol. Biochem. Behav. 2012, 102, 532–539. [Google Scholar] [CrossRef]
- Matuszewich, L.; Lorrain, D.S.; Hull, E.M. Dopamine release in the medial preoptic area of female rats in response to hormonal manipulation and sexual activity. Behav. Neurosci. 2000, 114, 772. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Numan, M.J.; Pliakou, N.; Stolzenberg, D.S.; Mullins, O.J.; Murphy, J.M.; Smith, C.D. The effects of D1 or D2 dopamine receptor antagonism in the medial preoptic area, ventral pallidum, or nucleus accumbens on the maternal retrieval response and other aspects of maternal behavior in rats. Behav. Neurosci. 2005, 119, 1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.M.; Lonstein, J.S. Dopamine d1 and d2 receptor antagonism in the preoptic area produces different effects on maternal behavior in lactating rats. Behav. Neurosci. 2005, 119, 1072. [Google Scholar] [CrossRef] [PubMed]
- Mileva-Seitz, V.; Afonso, V.M.; Fleming, A.S. Dopamine: Another “magic bullet” for caregiver responsiveness? In Evolution, Early Experience and Human Development: From Research to Practice and Policy; Oxford University Press: New York, NY, USA, 2013; pp. 152–178. [Google Scholar]
- Numan, M.; Stolzenberg, D.S. Medial preoptic area interactions with dopamine neural systems in the control of the onset and maintenance of maternal behavior in rats. Front. Neuroendocrinol. 2009, 30, 46–64. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M. Motivational views of reinforcement: Implications for understanding the behavioral functions of nucleus accumbens dopamine. Behav. Brain Res. 2002, 137, 3–25. [Google Scholar] [CrossRef]
- Hosking, J.G.; Floresco, S.B.; Winstanley, C.A. Dopamine antagonism decreases willingness to expend physical, but not cognitive, effort: A comparison of two rodent cost/benefit decision-making tasks. Neuropsychopharmacology 2015, 40, 1005–1015. [Google Scholar] [CrossRef]
- Correa, M.; Pardo, M.; Bayarri, P.; López-Cruz, L.; San Miguel, N.; Valverde, O.; Ledent, C.; Salamone, J.D. Choosing voluntary exercise over sucrose consumption depends upon dopamine transmission: Effects of haloperidol in wild type and adenosine A2AKO mice. Psychopharmacology 2016, 233, 393–404. [Google Scholar] [CrossRef]
- Ferreño, M.; Uriarte, N.; Zuluaga, M.J.; Ferreira, A.; Agrati, D. Dopaminergic activity mediates pups’ over male preference of postpartum estrous rats. Physiol. Behav. 2018, 188, 134–139. [Google Scholar] [CrossRef]
- Hernández-Munive, A.K.; Rebolledo-Solleiro, D.; Fernández-Guasti, A. Reduced sexual motivation of diabetic female rats: Restoration with insulin. Horm. Behav. 2021, 132, 104992. [Google Scholar] [CrossRef]
- Ventura-Aquino, E.; Fernández-Guasti, A. Reduced proceptivity and sex-motivated behaviors in the female rat after repeated copulation in paced and non-paced mating: Effect of changing the male. Physiol. Behav. 2013, 120, 70–76. [Google Scholar] [CrossRef]
- Guarraci, F.A.; Bolton, J.L. “Sexy stimulants”: The interaction between psychomotor stimulants and sexual behavior in the female brain. Pharmacol. Biochem. Behav. 2014, 121, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.L.; Ismail, N.; Pfaus, J.G. Facilitation of sexual behavior in ovariectomized rats by estradiol and testosterone: A preclinical model of androgen effects on female sexual desire. Psychoneuroendocrinology 2017, 79, 122–133. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The images show a sequence of female proceptive behaviors that promotes male chasing (panels (A–C)) and male mounting, with the consequent display of lordosis by the female (panel (D)).
Figure 1.
The images show a sequence of female proceptive behaviors that promotes male chasing (panels (A–C)) and male mounting, with the consequent display of lordosis by the female (panel (D)).

Figure 2.
Representation of the male vs. female preference model used (panel (A)) and of the 50 kHz ultrasonic vocalization emission test after a brief sexual (with a male) or social (with a female) interaction without physical contact (panel (B)).
Figure 2.
Representation of the male vs. female preference model used (panel (A)) and of the 50 kHz ultrasonic vocalization emission test after a brief sexual (with a male) or social (with a female) interaction without physical contact (panel (B)).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Agrati, D. Adolescence and Postpartum: Two Life Periods to Deepen Our Understanding of the Complexity of Female Rat Sexual Behavior. Sexes 2022, 3, 282-297. https://0-doi-org.brum.beds.ac.uk/10.3390/sexes3020022
AMA Style
Agrati D. Adolescence and Postpartum: Two Life Periods to Deepen Our Understanding of the Complexity of Female Rat Sexual Behavior. Sexes. 2022; 3(2):282-297. https://0-doi-org.brum.beds.ac.uk/10.3390/sexes3020022
Chicago/Turabian StyleAgrati, Daniella. 2022. "Adolescence and Postpartum: Two Life Periods to Deepen Our Understanding of the Complexity of Female Rat Sexual Behavior" Sexes 3, no. 2: 282-297. https://0-doi-org.brum.beds.ac.uk/10.3390/sexes3020022