Immunofluorescence Evaluation of Myf5 and MyoD in Masseter Muscle of Unilateral Posterior Crossbite Patients

 , ,
, , 
 , ,
, , 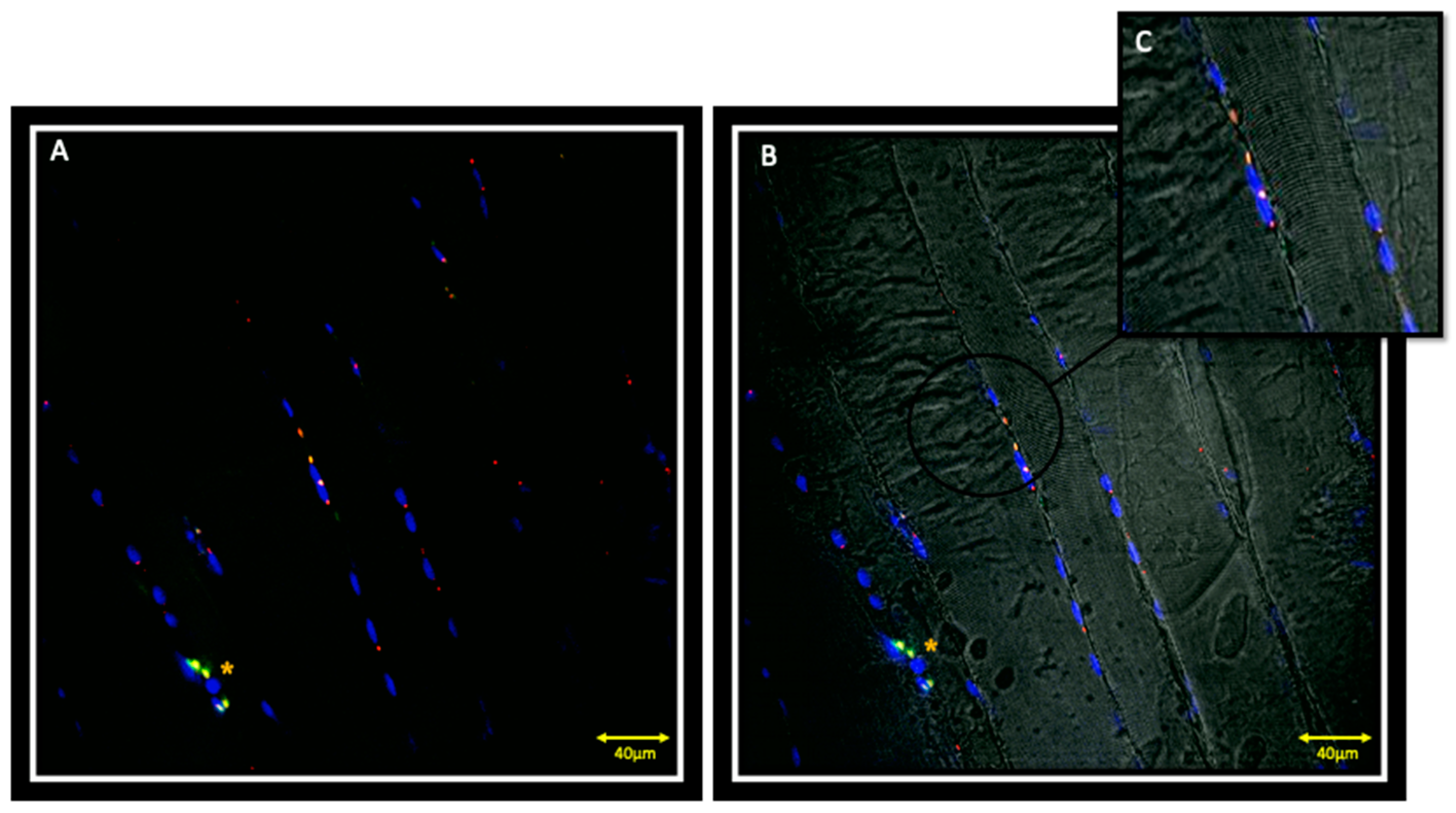{kind=link}
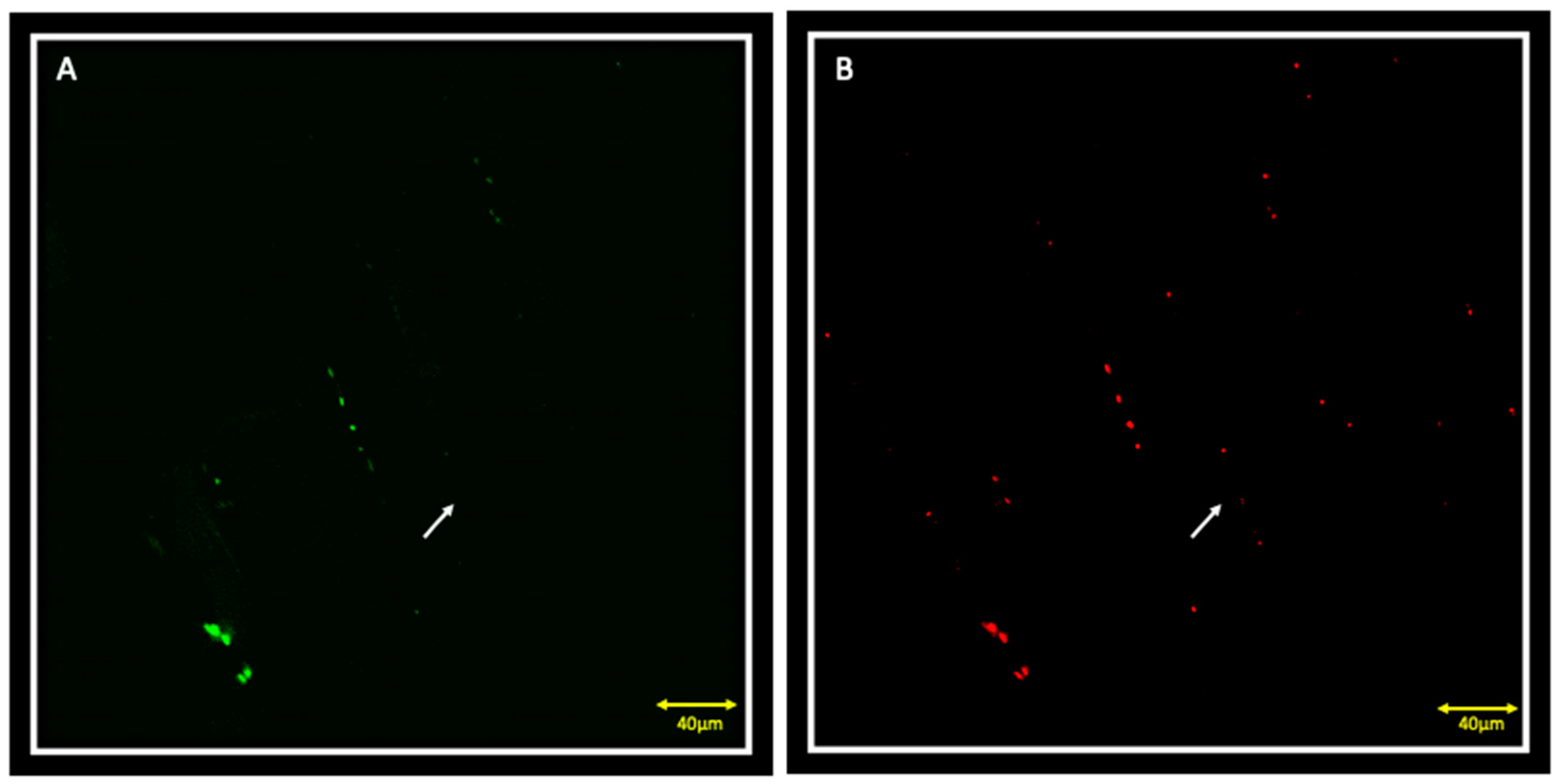{kind=link}
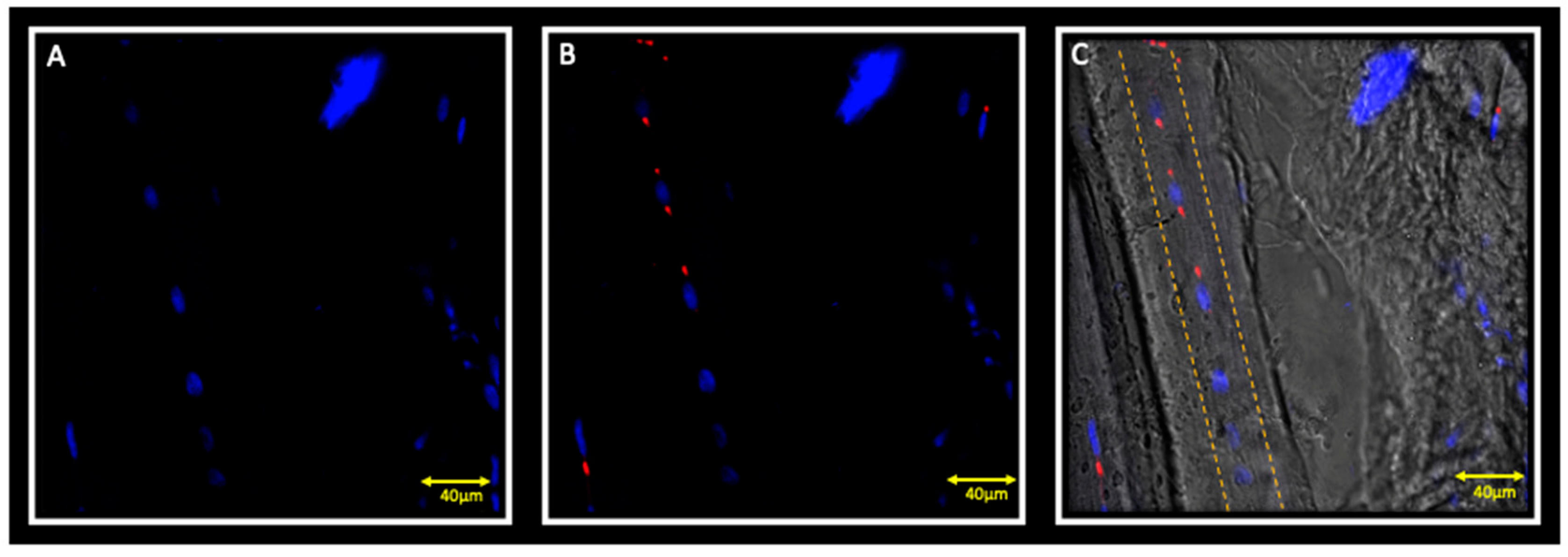{kind=link}
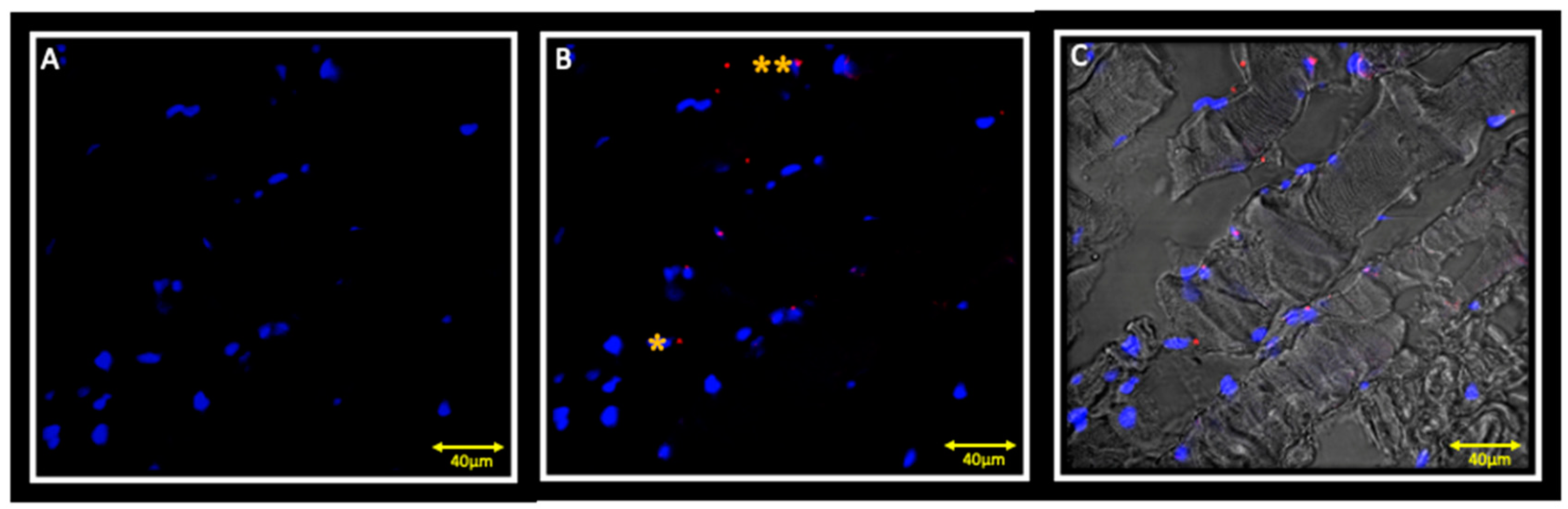{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Ethics
2.2. Muscle Biopsies
2.3. Immunofluorescence
2.4. Statistical Analysis
3. Results
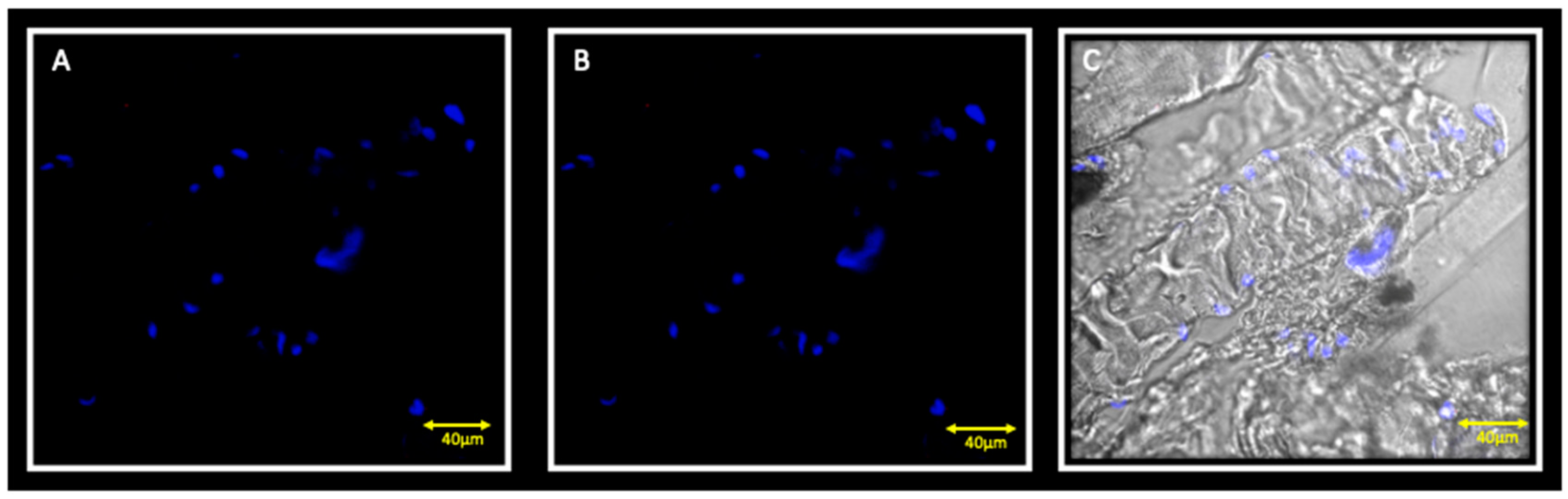
3.1. Immunofluorescence and Contralateral Muscle
3.2. Immunofluorescence and Ipsilateral Muscle
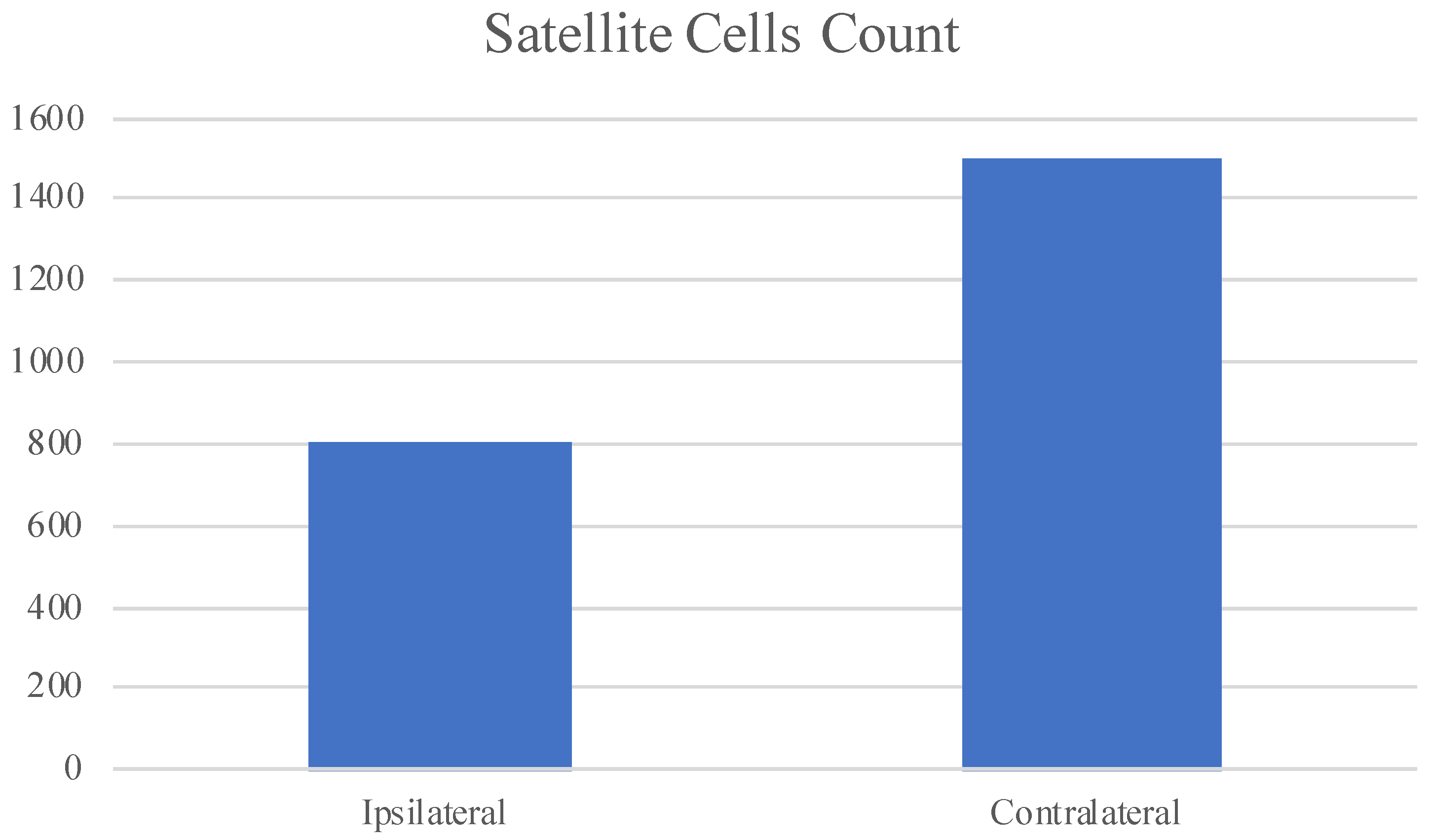
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Piancino, M.G.; Farina, D.; Talpone, F.; Merlo, A.; Bracco, P. Muscular activation during reverse and non-reverse chewing cycles in unilateral posterior crossbite. Eur. J. Oral. Sci. 2009, 117, 122–128. [Google Scholar] [CrossRef]
- Piancino, M.G.; Comino, E.; Talpone, F.; Vallelonga, T.; Frongia, G.; Bracco, P. Reverse-sequencing chewing patterns evaluation in anterior versus posterior unilateral crossbite patients. Eur. J. Orthod. 2012, 34, 536–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilding, R.J.; Lewin, A. The determination of optimal human jaw movements based on their association with chewing performance. Arch. Oral Biol. 1994, 39, 333–343. [Google Scholar] [CrossRef]
- Throckmorton, G.S.; Buschang, P.H.; Hayasaki, H.; Pinto, A.S. Changes in the masticatory cycle following treatment of posterior unilateral crossbite in children. Am. J. Orthod. Dentofac. Orthop. 2001, 120, 521–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bani, D.; Bani, T.; Bergamin, M. Morphologic and biochemical changes of the masseter muscles induced by occlusal wear: Studies in a rat model. J. Dent. Res. 1999, 78, 1735–1744. [Google Scholar] [CrossRef] [PubMed]
- Becht, M.P.; Mah, J.; Martin, C.; Razmus, T.; Gunel, E.; Ngan, P. Evaluation of masseter muscle morphology in different types of malocclusions using cone beam computed tomography. Int. Orthod. 2014, 2, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.S.; Pereira, C.L.; Issa, J.P.; Luiz, K.G.; Guimarães, E.A.; Gerlach, R.F. Histological, histochemical, and protein changes after induced malocclusion by occlusion alteration of Wistar rats. Biomed. Res. Int. 2014, 2014, 563463. [Google Scholar]
- Cutroneo, G.; Piancino, M.G.; Ramieri, G.; Bracco, P.; Vita, G.; Isola, G.; Vermiglio, G.; Favaloro, A.; Anastasi, G.; Trimarchi, F. Expression of muscle-specific integrins in masseter muscle fibers during malocclusion disease. Int. J. Mol. Med. 2012, 30, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Cutroneo, G.; Vermiglio, G.; Centofanti, A.; Rizzo, G.; Runci, M.; Favaloro, A.; Piancino, M.G.; Bracco, P.; Ramieri, G.; Bianchi, F.; et al. Morphofunctional Compensation of Masseter Muscles in Unilateral Posterior Crossbite Patients. Eur. J. Histochem. 2016, 60, 2605. [Google Scholar] [CrossRef]
- Jaffredo, T.; Horwitz, A.F.; Buck, C.A.; Rong, P.M.; Dieterlen-Lievre, F. Myoblast migration specifically inhibited in the chick embryo by grafted CSAT hybridoma cells secreting an anti- integrin antibody. Development 1988, 103, 431–446. [Google Scholar]
- Menko, A.S.; Boettiger, D. Occupation of the extracellular matrix receptor, integrin, is a control point for myogenic differentiation. Cell 1987, 51, 51–57. [Google Scholar] [CrossRef]
- Anastasi, G.; Cutroneo, G.; Sidoti, A.; Santoro, G.; D’Angelo, R.; Rizzo, G.; Rinaldi, C.; Giacobbe, O.; Bramanti, P.; Navarra, G.; et al. Sarcoglycans subcomplex in normal human smooth muscle: An immunohistochemical and molecular study. Int. J. Mol. Med. 2005, 48, 37–44. [Google Scholar] [CrossRef]
- Anastasi, G.; Cutroneo, G.; Rizzo, G.; Favaloro, A. Sarcoglycan subcomplex in normal and pathological human muscle fibers. Eur. J. Histochem. 2007, 51, 29–33. [Google Scholar] [PubMed]
- Di Mauro, D.; Gaeta, R.; Arco, A.; Milardi, D.; Lentini, S.; Runci, M.; Rizzo, G.; Magaudda, L. Distrubution of costameric proteins in normal human ventricular and atrial cardiac muscle. Folia Histochem. Cytobiol. 2009, 47, 605–608. [Google Scholar] [PubMed] [Green Version]
- Cutroneo, G.; Centofanti, A.; Speciale, F.; Rizzo, G.; Favaloro, A.; Santoro, G.; Bruschetta, D.; Milardi, D.; Micali, A.; Di Mauro, D.; et al. Sarcoglycan complex in masseter and sternocleidomastoid muscles of baboons: An immunohistochemical study. Eur. J. Histochem. 2015, 59, 2509. [Google Scholar] [CrossRef] [Green Version]
- Ventura Spagnolo, E.; Mondello, C.; Di Mauro, D.; Vermiglio, G.; Asmundo, A.; Filippini, E.; Alibrandi, A.; Rizzo, G. Analysis on sarcoglycans expression as markers of septic cardiomyopathy in sepsis-related death. J. Leg. Med. 2018, 132, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.N.; Tajbakhsh, S.; Mouly, V.; Cossu, G.; Buckingham, M.; Butler-Browne, G.S. In vivo satellite cell activation via Myf5 and MyoD in regenerating mouse. J. Cell Sci. 1999, 112, 2895–2901. [Google Scholar]
- Bruschetta, D.; Anastasi, G.; Andronaco, V.; Cascio, F.; Rizzo, G.; Di Mauro, D.; Bonanno, L.; Izzo, V.; Buda, D.; Vermiglio, G.; et al. Human calf muscles changes after strength training as revealed by diffusion tensor imaging. J. Sports Med. Phys. Fit. 2019, 59, 853–860. [Google Scholar] [CrossRef]
- Boyd, S.B.; Gonyea, W.J.; Legan, H.L.; Bell, W.H. Masseter muscle adaptation following surgical correction of vertical maxillary excess. J. Oral Maxillofac. Surg. 1989, 47, 953–962. [Google Scholar] [CrossRef]
- De Ponte, F.S.; Falzea, R.; Runci, M.; Nastro Siniscalchi, E.; Lauritano, F.; Bramanti, E.; Cervino, G.; Cicciu, M. Histomorhological and clinical evaluation of maxillary alveolar ridge reconstruction after craniofacial trauma by applying combination of allogeneic and autogenous bone graft. Chin. J. Traumatol. 2017, 20, 14–17. [Google Scholar] [CrossRef]
- De Ponte, F.S.; Favaloro, A.; Siniscalchi, E.N.; Centofanti, A.; Runci, M.; Cutroneo, G.; Catalfamo, L. Sarcoglycans and integrins in bisphosphonate treatment: Immunohistochemical and scanning electron microscopy study. Oncol. Rep. 2013, 30, 2639–2646. [Google Scholar] [CrossRef]
- De Ponte, F.S.; Cutroneo, G.; Falzea, R.; Rizzo, G.; Catalfamo, L.; Favaloro, A.; Vermiglio, G.; Runci, M.; Centofanti, A.; Anastasi, G. Histochemical and morphological aspects of fresh frozen bone: A preliminary study. Eur. J. Histochem. 2016, 60, 2642. [Google Scholar] [CrossRef]
- De Ponte, F.S.; Catalfamo, L.; Micali, G.; Runci, M.; Cutroneo, G.; Vermiglio, G.; Centofanti, A.; Rizzo, G. Effect of bisphosphonates on the mandibular bone and gingival epithelium of rats without tooth extraction. Exp. Med. 2016, 11, 1678–1684. [Google Scholar] [CrossRef] [Green Version]
- Irrera, N.; Arcoraci, V.; Mannino, F.; Vermiglio, G.; Pallio, G.; Minutoli, L.; Bagnato, G.; Anastasi, G.; Mazzon, E.; Bramanti, P.; et al. Activation of A2A Receptor by PDRN Reduces Neuronal Damage and Stimulates WNT/β-CATENIN Driven Neurogenesis in Spinal Cord Injury. Front. Pharmacol. 2018, 9, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Militi, A.; Cutroneo, G.; Favaloro, A.; Matarese, G.; Di Mauro, D.; Lauritano, F.; Centofanti, A.; Cervino, G.; Nicita, F.; Bramanti, A.; et al. An immunofluorescence study on VEGF and extracellular matrix proteins in human periodontal ligament during tooth movement. Heliyon 2019, 5, e02572. [Google Scholar] [CrossRef] [Green Version]
- Cuccia, A.M.; Caradonna, C.; Caradonna, D.; Anastasi, G.; Milardi, D.; Favaloro, A.; De Pietro, A.; Angileri, T.M.; Caradonna, L.; Cutroneo, G. The arterial blood supply of the temporomandibular joint: An anatomical study and clinical implications. Imaging Sci. Dent. 2013, 43, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Watkins, S.C.; Cullen, M.J. A quantitative comparison of satellite cell ultrastructure in Duchenne muscular dystrophy, polymyositis, and normal controls. Muscle Nerve 1986, 9, 724–730. [Google Scholar] [CrossRef]
- Bischoff, R.; Heintz, C. Enhancement of skeletal muscle regeneration. Dev. Dyn. 1994, 201, 41–54. [Google Scholar] [CrossRef]
- Tajbakhsh, S.; Buckingham, M. The birth of muscle progenitor cells in the mouse: Spatiotemporal considerations. Curr. Top. Dev. Biol. 2000, 48, 225–268. [Google Scholar]
- Grounds, M.D.; Garrett, K.L.; Lai, M.C.; Wright, W.E.; Beilharz, M.W. Identification of skeletal muscle precursor cells in vivo by use of MyoD1 and myogenin probes. Cell Tissue Res. 1992, 267, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Yablonka-Reuveni, Z.; Rivera, A.J. Temporal expression of regulatory and structural muscle proteins during myogenesis of satellite cells on isolated adult rat fibers. Dev. Biol. 1994, 164, 588–603. [Google Scholar] [CrossRef] [Green Version]
- Zammit, P.S.; Heslop, L.; Hudon, V.; Rosenblatt, J.D.; Tajbakhsh, S.; Buckingham, M.E.; Beauchamp, J.R.; Partridge, T.A. Kinetics of myoblast proliferation show that resident satellite cells are competent to fully regenerate skeletal muscle fibers. Exp. Cell Res. 2002, 281, 39–49. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vermiglio, G.; Centofanti, A.; Ramieri, G.; Tepedino, M.; Runci Anastasi, M.; Micali, A.G.; Arco, A.; Piancino, M.G. Immunofluorescence Evaluation of Myf5 and MyoD in Masseter Muscle of Unilateral Posterior Crossbite Patients. J. Funct. Morphol. Kinesiol. 2020, 5, 80. https://0-doi-org.brum.beds.ac.uk/10.3390/jfmk5040080
Vermiglio G, Centofanti A, Ramieri G, Tepedino M, Runci Anastasi M, Micali AG, Arco A, Piancino MG. Immunofluorescence Evaluation of Myf5 and MyoD in Masseter Muscle of Unilateral Posterior Crossbite Patients. Journal of Functional Morphology and Kinesiology. 2020; 5(4):80. https://0-doi-org.brum.beds.ac.uk/10.3390/jfmk5040080
Chicago/Turabian StyleVermiglio, Giovanna, Antonio Centofanti, Guglielmo Ramieri, Michele Tepedino, Michele Runci Anastasi, Antonio Girolamo Micali, Alba Arco, and Maria Grazia Piancino. 2020. "Immunofluorescence Evaluation of Myf5 and MyoD in Masseter Muscle of Unilateral Posterior Crossbite Patients" Journal of Functional Morphology and Kinesiology 5, no. 4: 80. https://0-doi-org.brum.beds.ac.uk/10.3390/jfmk5040080