
Proton-Cluster-Beam Lethality and Mutagenicity in Bacillus subtilis Spores
by
Yoshihiro Hase
1,2,*, 
Katsuya Satoh
1,2, 
Atsuya Chiba
1,
Yoshimi Hirano
1,
Kengo Moribayashi
2 and
Kazumasa Narumi
1 1
Takasaki Advanced Radiation Research Institute, National Institutes for Quantum and Radiological Science and Technology (QST), 1233 Watanuki, Takasaki 370-1292, Gunma, Japan
2
Institute for Quantum Life Science, National Institutes for Quantum and Radiological Science and Technology (QST), 8-1-7 Umemidai, Kizugawa 619-0215, Kyoto, Japan
*
Author to whom correspondence should be addressed.
Quantum Beam Sci. 2021, 5(3), 25; https://0-doi-org.brum.beds.ac.uk/10.3390/qubs5030025
Submission received: 26 April 2021
/
Revised: 25 August 2021
/
Accepted: 25 August 2021
/
Published: 28 August 2021
(This article belongs to the Special Issue Swift Cluster Ion Beams: Basic Processes and Applications)
Abstract
:The unique energy transfer characteristics of swift cluster ions have attracted the attention of many researchers working on the analysis or processing of material surfaces, but the effects on living organisms remain unclear. We irradiated B. subtilis spores with monomer and cluster proton beams and examined their lethality; the 2 MeV H2+ shows a clearly lower lethality than 340 keV H+, even though both have a comparable linear energy transfer. The 2 MeV H2+ dissociates into a pair of 1 MeV H+ by losing the bonding electrons at the target surface. The estimated internuclear distance and the radial dose distribution suggest that the spread of deposited total energy over two areas separated by just several nanometers greatly diminishes beam lethality and that the energy density in the very center of the trajectory, possibly within a 1 nm radius, has a great impact on lethality. We also performed a whole genome resequencing of the surviving colonies to compare the molecular nature of mutations but failed to find a clear difference in overall characteristics. Our results suggest that cluster beams may be a useful tool for understanding biological effects of high linear energy transfer radiation.
1. Introduction
Mutagenesis with high linear energy transfer (LET) radiations have been shown to be an effective means to generate new varieties in plants and microorganisms [1,2,3]. The high biological effects of high LET radiation attribute to the dense ionization and excitation along the tracks, as well as the severe and multiple DNA damages generated thereby [4,5,6]. In order to develop more efficient mutagenesis techniques, we need to deepen our understanding of the radiation quality dependence. Cluster ion beams are unique quantum beams and particular attention has been paid to their irradiation effects incurred from the very high energy densities at the surfaces of their targets [7,8,9]. This unique energy deposition is largely attributed to the incidence of multiple atoms that are in close proximity. The cluster ions dissociate into monomer ions by losing bonding electrons through the collision with the atoms at the target surface, then the interatomic distance broadens within the target due to the Coulomb repulsion as well as multiple scattering. Therefore, the cluster ions could produce a unique track structure within the target, which cannot be achieved by conventional monomer ion beams, depending on the cluster size and internuclear distance. This may help our understanding of the relationship of track structure to biological effects. Moreover, the cluster ions may induce different mutations at the molecular level. Therefore, we have developed an experimental method using bacterial spores as a model organism [10]. Bacillus subtilis spores have a high tolerance to vacuums and thus can be irradiated in a vacuum chamber [11,12]. They are also sufficiently thin for homogeneous irradiation with a limited penetration to cluster ions of energy of a few MeV [13]. However, when we used 2 MeV/atom carbon ions (2 MeV C+, 4 MeV C2+ and 6 MeV C3+), no difference was observed in lethality [10]. Further investigations using monomer ion beams with a wide range of LET showed that their lethality reached a maximum at around 156 keV/µm and decreased with a further increase in LET. This result suggests that even a single particle of 2 MeV C+ (1141 keV/µm) deposits an energy sufficient to kill a spore, thereby explaining why no difference was observed among the 2 MeV C+, 4 MeV C2+ and 6 MeV C3+. Therefore, cluster ion beams with an LET less than 156 keV/µm should be used to examine cluster ion-specific irradiation effects. In extending our previous study, we examine here the lethality and mutagenicity in B. subtilis spores using proton cluster beams.
2. Materials and Methods
2.1. Sample Preparation
The spore samples were prepared as previously reported [10]. A fresh spore suspension solution of B. subtilis (strain 168) was prepared in sterile water within one week before the irradiation experiments. A 2.7 µL spore suspension solution containing 2 × 105 spores was aliquoted onto small pieces (approximately 8 mm square) of silicon wafer (SI-500452, The Nilaco Corporation, Tokyo, Japan) and then placed in a sterile 12-well culture plates. After freeze-drying using a freeze drier (FDU-2200, EYLA, Tokyo, Japan), the 12-well plates were placed on ice for 3 to 5 min until water vapor began to condense on the surface. Then, the samples were air dried on a clean bench to spread the spores out to form a near monolayer. The diameter of the spore spots was less than 4.0 mm. All samples were prepared on the day before irradiation.
2.2. Irradiation
Under vacuum conditions (<1 × 10−5 Pa), the spore samples were irradiated with monomer and cluster proton beams (see Table 1 and Figure 1 for characteristics). We used the irradiation chamber connected to the 3 MV single-ended accelerator and the 400 keV ion implanter at the Takasaki Advanced Radiation Research Institute, National Institutes for Quantum and Radiological Science and Technology (Takasaki, Gunma, Japan) [14]. The beam size was adjusted with an aperture of 4.0 mm diameter. The beam current was monitored using a non-suppressed deep Faraday cup (20 mm diameter × 300 mm depth); the intensity was adjusted using a beam chopper installed in the beam line.
2.3. Survival Assay
The survival fraction of irradiated spores was determined as previously reported [10]. The irradiated spores on each silicon wafer were corrected in 1 mL of PBT buffer (10 mM sodium phosphate, 0.01% Tween 20, pH 7.0) by pipetting several times. The serial dilutions (10-, 100- and 1000-fold) of the spore suspension solution were prepared with PBT buffer and 100 µL samples were spread on LB plates (Luria-Bertani broth solidified with 1.5% agar). After an overnight incubation at 37 °C, the colony forming units (CFUs), i.e., the number of viable spores in a sample, were determined. The surviving fraction was defined as the ratio of the mean CFU of irradiated samples to the mean CFU of the mock irradiated control. Data from five silicon wafers were used for each data point.
2.4. Calculation of Physical Parameters
The penetration range, surface LET and lateral straggling of protons in the spore were calculated using the SRIM code [15]. The relationship between LET and penetration range was calculated using the IRAC M code [16]. The density and thickness of spores were assumed to be 1.52 g/cm3 and 0.48 µm, respectively [13,17]. Because the elemental composition of B. subtilis spores has not been reported, the elemental composition of dry yeast (C40.14, H75.24, O21.16, N4.72, K0.50, P0.46, Mg0.12, S0.02) was used [18]. The absorbed dose was calculated from the formula: Dose (Gy) = 1.6 × 10−9 × surface LET (keV/µm) × Fluence (particles/cm2).
The radial dose distribution was calculated as described elsewhere [19]. In brief, water molecules are located in a rectangular target so as to obtain the liquid water density. When an ion passes through this target, water molecular ions and secondary electrons are created in accordance with cross sections of the incident ion impact ionization or stopping powers using the binary collision model. The secondary electrons produced here are moved in the target until the secondary electrons have little energy to induce ionization and excitation of the water molecules. Then, the radial dose is simulated from the energy deposition of the secondary electrons to the target. The Coulomb interactions between the individual secondary electrons and individual charged particles (molecular ions or the other secondary electrons) are also considered in this simulation.
3. Results and Discussion
3.1. Lethality of Cluster Ions
Four different proton beams were used (Table 1): the 1 MeV H+ and 2 MeV H2+ beams both have the same energy per atom (1 MeV/atom), the 340 keV H+ beam having a LET value comparable to that of 2 MeV H2+ per particle and the 500 keV H+ beam having an intermediate LET value between 340 KeV H+ and 1 MeV H+. The surviving fraction of the irradiated spores are plotted in Figure 2. The lethality of the three monomer proton beams depends on the LET value, with 340 keV H+ producing the highest effect. This is consistent with the observed LET dependency in B. subtills spores exposed to various kinds of heavy ions with the LET range of 2–200 keV/µm [20]. The protons are the majority of high-energy charged particles in space environment; the 218 MeV proton (0.4 keV/µm) was shown to reduce the surviving fraction to 0.01 at around 2.5 kGy [21]. Though the LET value of 218 MeV proton is very low, the lethal effect per dose is higher than that of proton beams used in this study. This could be due to that they irradiated air dried spores at atmospheric condition, whereas we irradiated freeze dried spores in a vacuum chamber. The killing effect has also been examined for 10 to 200 keV electron beams at atmospheric condition for food decontamination; around 3 to 6 kGy was required to reduce the surviving fraction to 0.01 [22].
The 2 MeV H2+ showed a higher lethality per particle than 1 MeV H+ (Figure 2a; t test, p < 0.01 at 5 × 1010 particles/cm2). This is conceivable because twice the number of protons were bombarded per particle of 2 MeV H2+ compared with that of 1 MeV H+. In contrast, no significant difference was observed in the lethality per atom at data points that can be compared directly (Figure 2b; p = 0.122 at 1 × 1011 atoms/cm2). Further experiments are required to determine whether there is a difference in this instance.
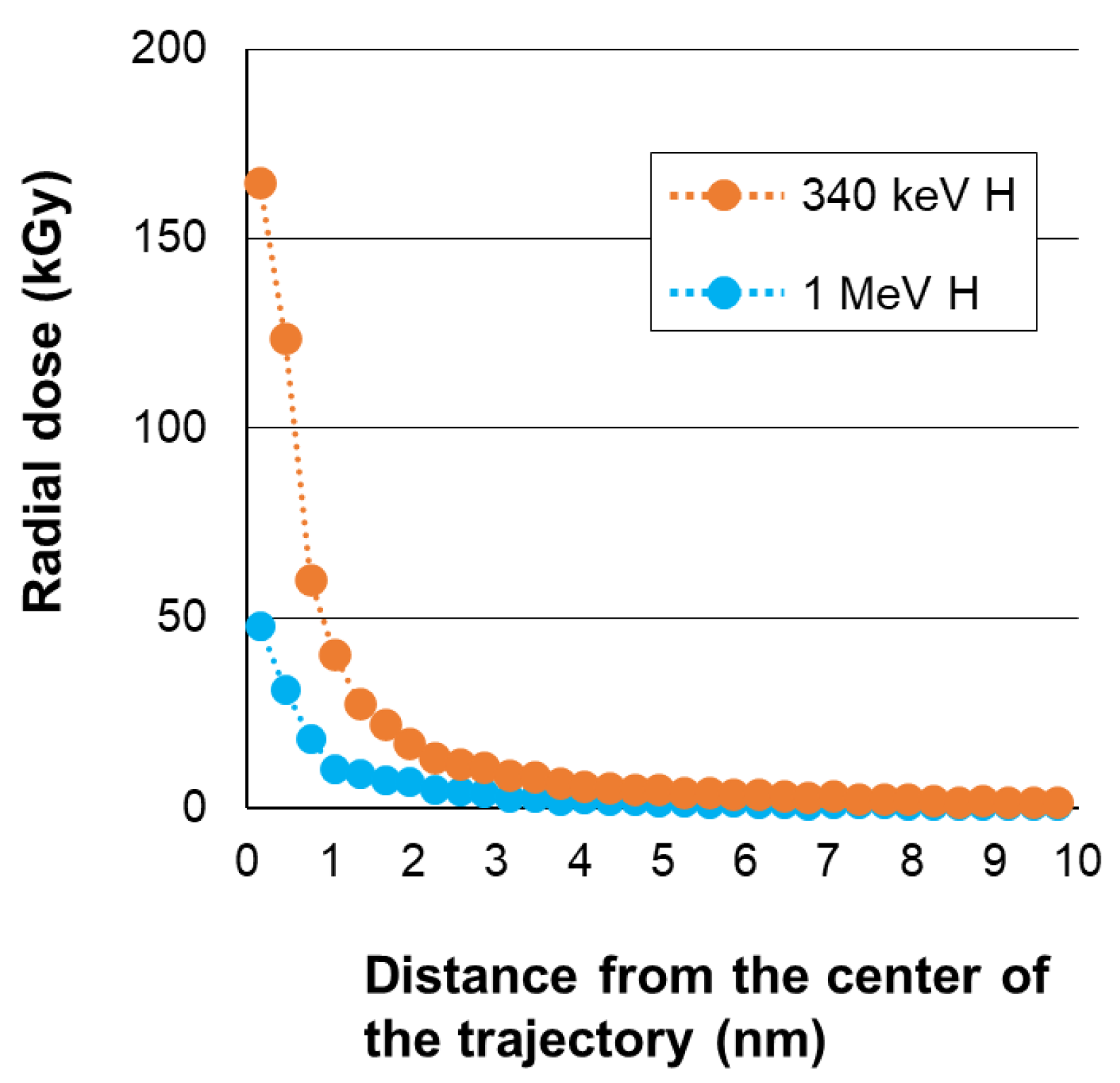
The 2 MeV H2+ exhibits a clear lower lethality than 340 keV H+, even though both have a similar LET value per particle (Figure 2a,c). This arises from the difference in the spatial energy deposition profile. The 2 MeV H2+ molecule dissociates into a pair of 1 MeV H+ by losing the bonding electrons at the surface of the target. We considered the internuclear distance based on the Coulomb repulsion and multiple scattering as with our previous study [10]. The internuclear distance was thought to be mostly within several nanometers while passing through the spore (Figure 3). This suggests that a single atom of 340 keV H+ has a higher lethality than a pair of 1 MeV H+ spaced within several nanometers. The 340 keV H+ shows a greater increase of LET than 1 MeV H+ in the depth direction of the spores (Figure 1). However, it is less than 10% and, therefore, we believe the depth distribution of the deposited energy is not a major cause of the difference of the lethality between 340 keV H+ and 1 MeV H+. The radial dose distribution calculated assuming water as a target suggests that both the 340 keV H+ and the 1 MeV H+ beams give a high dose, particularly within a 1 nm radius from the center of the trajectory; moreover, the dose in this area is more than three times higher in 340 keV H+ than in 1 MeV H+ (Figure 4). These facts suggest that energy density, rather than total energy deposited in a nanometer area contributes to lethality. The significance of energy density in regard to cell killing is well supported by experiments using monomer ions of various nuclear species with different energies [23,24]. However, our result first demonstrates experimentally that the split of deposited total energy into two places separated only by up to several nanometers diminishes lethality considerably. Our results corroborate the notion that the energy density in the very center of the trajectory, possibly within a 1 nm radius, has an immense impact on lethality.
3.2. Mutagenicity of Cluster Ions
The occurrence of multiple damage within one to two helical turns of DNA is defined as a clustered DNA damage and is believed to be a major cause of significant biological effects from high LET radiations [4,5,6]. As described above, the internuclear distance of 2 MeV H2+ is approximately within several nanometers when passing through the spore, the distance being comparable to one to two helical turns of DNA. The internuclear distance of 2 MeV H2+ is shorter than the mean distance of randomly radiated monomer 1 MeV H+ ions (~32 nm on average at a fluence of 1 × 1011 atoms/cm2). The question then is whether 2 MeV H2+ and 1 MeV H+ may have different mutagenicity. We performed the whole genome sequencing analysis of the surviving colonies; however, we failed to find a clear difference in the molecular nature of the mutation between 1 MeV H+ and 2 MeV H2+ (Supplementary Figures S1–S3 and Tables S1–S3). The number of mutation events observed in each surviving colony was around two to five after the irradiation at a fluence of 5 × 1010 or 1 × 1011 atoms/cm2 (500 or 1000 atoms/µm2). Given that only a minor fraction of incident atoms is responsible for the mutagenesis, the mutagenicity of cluster ions is likely to be similar with that of monomer ions.
4. Conclusions
We showed that cluster and monomer ions have different lethality and results from our experiments suggested that the energy density in the very center of the radiation track affects the lethality immensely. Ion beams of different cluster sizes may become useful tools for understanding biological effects of high-LET radiations.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/qubs5030025/s1, Figure S1: Relationship between surviving fraction and total mutation frequency, Figure S2: Spectra of single-base substitutions, Figure S3: Schematic representation of the predicted overall structure of SVs, Table S1: Frequency of each mutation type, Table S2: Distribution of deletion size, Table S3: Predicted SVs.
Author Contributions
Y.H. (Yoshihiro Hase) and K.S. designed the experiments; A.C. established the irradiation system; A.C., Y.H. (Yoshimi Hirano) and K.N. contributed to the irradiation experiment; Y.H. (Yoshihiro Hase) and K.S. performed survival assay and whole genome sequencing; K.M. performed simulation on radial dose; Y.H. (Yoshihiro Hase) and K.S. interpreted the data; Y.H. (Yoshihiro Hase) wrote the first draft; all authors approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by JSPS KAKENHI Grants JP17K05133 and JP20K12489.
Acknowledgments
The authors thank the technical staff of the electrostatic accelerators and N. Shikazono (QST) for their support in conducting this work, and Y. Yokota (QST) for his critical comments on the manuscript. We thank Edanz Group for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jo, Y.; Kim, J.B. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses in plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Li, W.; Chen, J. Recent advances of microbial breeding via heavy-ion mutagenesis at IMP. Lett. Appl. Microbiol. 2017, 65, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Oono, Y. Studies on application of ion beam breeding to industrial microorganisms at TIARA. Quantum Beam Sci. 2019, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Goodhead, D.T. Initial events in the cellular effects of ionizing radiations: Clustered damage in DNA. Int. J. Radiat. Biol. 1994, 65, 7–17. [Google Scholar] [CrossRef]
- Sage, E.; Shikazono, N. Radiation-induced clustered DNA lesions: Repair and mutagenesis. Free Radic. Biol. Med. 2017, 107, 125–135. [Google Scholar] [CrossRef]
- Mavragani, I.V.; Nikitaki, Z.; Kalospyros, S.A.; Georgakilas, A.G. Ionizing radiation and complex DNA damage: From prediction to detection challenges and biological significance. Cancers 2019, 11, 1789. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, A.; Jaskierowicz, G.; Della-Negra, S. Latent track formation in silicon irradiation by 30 MeV fullerenes. Nucl. Instr. Meth. Phys. Res. B 1998, 146, 302–308. [Google Scholar] [CrossRef]
- El-Said, A.S. Tracks of 30-MeV C60 clusters in yttrium iron garnet studied by scanning force microscopy. Nucl. Instr. Meth. Phys. Res. B 2009, 267, 953–956. [Google Scholar] [CrossRef] [Green Version]
- Koide, T.; Saitoh, Y.; Sakamaki, M.; Amemiya, K.; Iwase, A.; Matsui, T. Change in magnetic and structural properties of FeRh thin films by gold cluster ion beam irradiation with the energy of 1.67 MeV/atom. J. Appl. Phys. 2014, 115, 17B722. [Google Scholar] [CrossRef]
- Hase, Y.; Satoh, K.; Chiba, A.; Hirano, Y.; Tomita, S.; Saito, Y.; Narumi, I. Experimental study on biological effect of cluster ion beams in Bacillus subtilis spores. Quantum Beam Sci. 2019, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef]
- Carrera, M.; Zandomeni, R.O.; Fitzgibbon, J.; Sagripanti, J.-L. Difference between the spore size of Bacillus anthracis and other Bacillus species. J. App. Microbiol. 2007, 102, 303–312. [Google Scholar] [CrossRef]
- Kurashima, S.; Satoh, T.; Saitoh, Y.; Yokota, W. Irradiation Facilities of the Takasaki Advanced Radiation Research Institute. Quantum Beam Sci. 2017, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, J.F.; Ziegler, M.D.; Biersack, J.P. SRIM—The stopping and range of ions in matter. Nucl. Instr. Meth. Phys. Res. B 2010, 268, 1818–1823. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Fukuda, K.; Nishimura, K.; Watanabe, H.; Yamano, N. IRAC M: A Code System to Calculate Induced Radioactivity Produced by Ions and Neutrons. JAERI-Data/Code 97-019; Japan Atomic Energy Research Institute: Ibaraki, Japan, 1997. [Google Scholar]
- Carrera, M.; Zandomeni, R.O.; Sagripanti, J.-L. Wet and dry density of Bacillus anthracis and other Bacillus species. J. App. Microbiol. 2008, 105, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, T.; Fujimoto, S.; Saga, K.; Minowa, T. Process Simulation of Yeast Cultivation and Ethanol Fermentation in Bio-ethanol Production. Energy Resour. 2010, 31, 335–340. (In Japanese) [Google Scholar]
- Moribayashi, K. Application of atomic and molecular data for plasma production and cancer therapy by heavy particle irradiation. Jpn. J. Appl. Phys. 2020, 59, SH0801. [Google Scholar] [CrossRef]
- Moeller, R.; Reitz, G.; Berger, T.; Okayasu, R.; Nicholson, W.L.; Horneck, G. Astrobiological aspects of the mutagenesis of cosmic radiation on bacterial spores. Astrobiology 2010, 10, 509–521. [Google Scholar] [CrossRef]
- Moeller, R.; Reitz, G.; Li, Z.; Klein, S.; Nicholson, W.L. Multifractional resistance of Bacillus subtilis spores to high-energy proton radiation: Role of spore structural components and the homologous recombination and non-homologous end joining DNA repair pathways. Astrobiology 2012, 12, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huber, N.; Moeller, R.; Stülke, J.; Dubovcova, B.; Akepsimaidis, G.; Meneses, N.; Drissner, D.; Mathys, A. Role of DNA repair in Bacillus subtilis spore resistance to high energy and low energy electron beam treatments. Food Microbiol. 2020, 87, 103353. [Google Scholar] [CrossRef]
- Furusawa, Y.; Fukutsu, K.; Aoki, M.; Itsukaichi, H.; Eguchi-Kasai, K.; Ohara, H.; Yatagai, F.; Kanai, T.; Ando, K. Inactivation of aerobic and hypoxic cells from three different cell lines by accelerated 3He-, 12C- and 20Ne-ion beams. Radiat. Res. 2000, 154, 485–496. [Google Scholar] [CrossRef]
- Jones, B.; Hill, M.A. The Physical separation between the LET associated with the ultimate relative biological effect (RBE) and the maximum LET in a proton or ion beam. Biomed. Phys. Eng. Express 2020, 6, 055001. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relationship between LET and penetration range. The values were calculated assuming dried bacteria.
Figure 1.
Relationship between LET and penetration range. The values were calculated assuming dried bacteria.

Figure 2.
Surviving fraction of irradiated B. subtilis spores. Surviving fraction relative to that of mock irradiated spores are shown as a function of particles/cm2 (a), atoms/cm2 (b) and kGy (c). Data points represent mean ± standard errors of three to six independent experiments. An exponential regression was used to fit the data.
Figure 2.
Surviving fraction of irradiated B. subtilis spores. Surviving fraction relative to that of mock irradiated spores are shown as a function of particles/cm2 (a), atoms/cm2 (b) and kGy (c). Data points represent mean ± standard errors of three to six independent experiments. An exponential regression was used to fit the data.

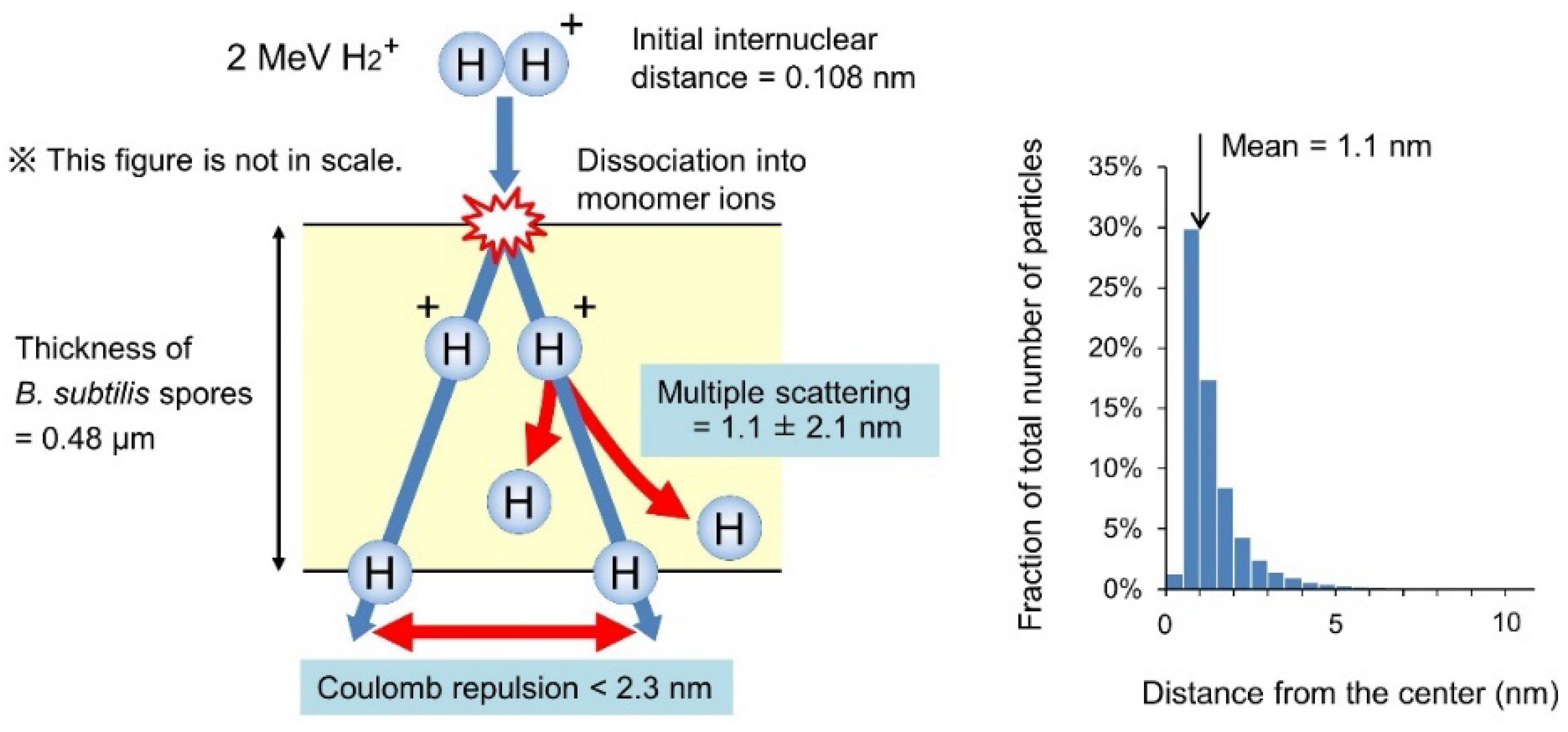
Figure 3.
Estimation of the internuclear distance between the two protons of a 2 MeV H2+ ion on passing through the B. subtilis spore. A 2 MeV H2+ ion with an initial internuclear distance of 0.108 nm incident into the spore of 0.48 µm thickness is assumed; the ion then dissociates into a couple of 1 MeV H+ at the surface by losing bonding electrons through inelastic collisions with atoms. If a 2 MeV H2+ dissociates into a horizontal direction and a pair of 1 MeV H+ travel 0.48 µm in the vacuum, the internuclear distance broadens to 2.3 nm by Coulomb repulsion. The actual internuclear distance is believed to be less than 2.3 nm, because the Coulomb repulsion is partially shielded in the spores and two protons of a 2 MeV H2+ are not always aligned in a horizontal direction. In addition, the trajectory of the 1 MeV H+ deviates by multiple scattering from the center of the trajectory by 1.1 ± 2.1 nm (mean ± standard deviation) while passing through the spore. The graph on the right indicates the distribution of horizontal deviation from the center of the trajectory for the 1 MeV H+ ions at the bottom surface of the target calculated using the SRIM code [15]. It suggests that the internuclear distance between the two protons of the 2 MeV H2+ ion is mostly within few nanometers while passing through a spore.
Figure 3.
Estimation of the internuclear distance between the two protons of a 2 MeV H2+ ion on passing through the B. subtilis spore. A 2 MeV H2+ ion with an initial internuclear distance of 0.108 nm incident into the spore of 0.48 µm thickness is assumed; the ion then dissociates into a couple of 1 MeV H+ at the surface by losing bonding electrons through inelastic collisions with atoms. If a 2 MeV H2+ dissociates into a horizontal direction and a pair of 1 MeV H+ travel 0.48 µm in the vacuum, the internuclear distance broadens to 2.3 nm by Coulomb repulsion. The actual internuclear distance is believed to be less than 2.3 nm, because the Coulomb repulsion is partially shielded in the spores and two protons of a 2 MeV H2+ are not always aligned in a horizontal direction. In addition, the trajectory of the 1 MeV H+ deviates by multiple scattering from the center of the trajectory by 1.1 ± 2.1 nm (mean ± standard deviation) while passing through the spore. The graph on the right indicates the distribution of horizontal deviation from the center of the trajectory for the 1 MeV H+ ions at the bottom surface of the target calculated using the SRIM code [15]. It suggests that the internuclear distance between the two protons of the 2 MeV H2+ ion is mostly within few nanometers while passing through a spore.

Figure 4.
Calculated radial dose distributions for the 340 keV H+ and 1 MeV H+ proton beams.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the monomer and cluster proton beams used.
| Beam | Surface LET (keV/µm) | Penetration Range (µm) |
|---|---|---|
| 1 MeV H+ | 37.7 | 16.7 |
| 2 MeV H2+ | 37.7 × 2 (75.4) * | 16.7 * |
| 500 keV H+ | 58.9 | 5.8 |
| 340 keV H+ | 75.0 | 3.4 |
The LET values and penetration ranges were calculated assuming dried bacteria. * Values assuming no interaction between the two protons of the H2+ ion.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hase, Y.; Satoh, K.; Chiba, A.; Hirano, Y.; Moribayashi, K.; Narumi, K. Proton-Cluster-Beam Lethality and Mutagenicity in Bacillus subtilis Spores. Quantum Beam Sci. 2021, 5, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs5030025
AMA Style
Hase Y, Satoh K, Chiba A, Hirano Y, Moribayashi K, Narumi K. Proton-Cluster-Beam Lethality and Mutagenicity in Bacillus subtilis Spores. Quantum Beam Science. 2021; 5(3):25. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs5030025
Chicago/Turabian StyleHase, Yoshihiro, Katsuya Satoh, Atsuya Chiba, Yoshimi Hirano, Kengo Moribayashi, and Kazumasa Narumi. 2021. "Proton-Cluster-Beam Lethality and Mutagenicity in Bacillus subtilis Spores" Quantum Beam Science 5, no. 3: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs5030025