Pathogenic Leptospira in Commensal Small Mammals from the Extensively Urbanized Coastal Benin


Abstract
:1. Introduction
2. Material and Methods
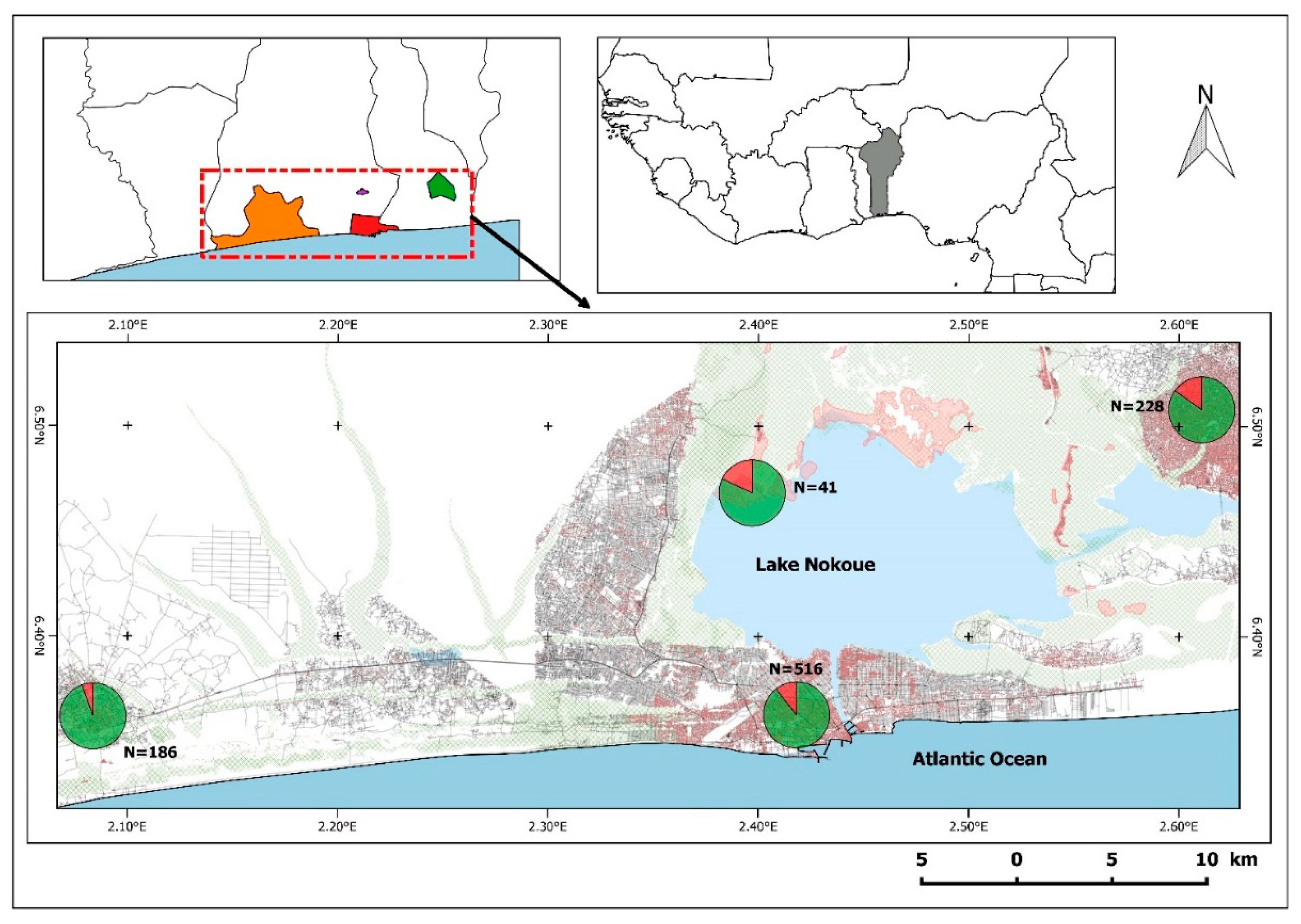
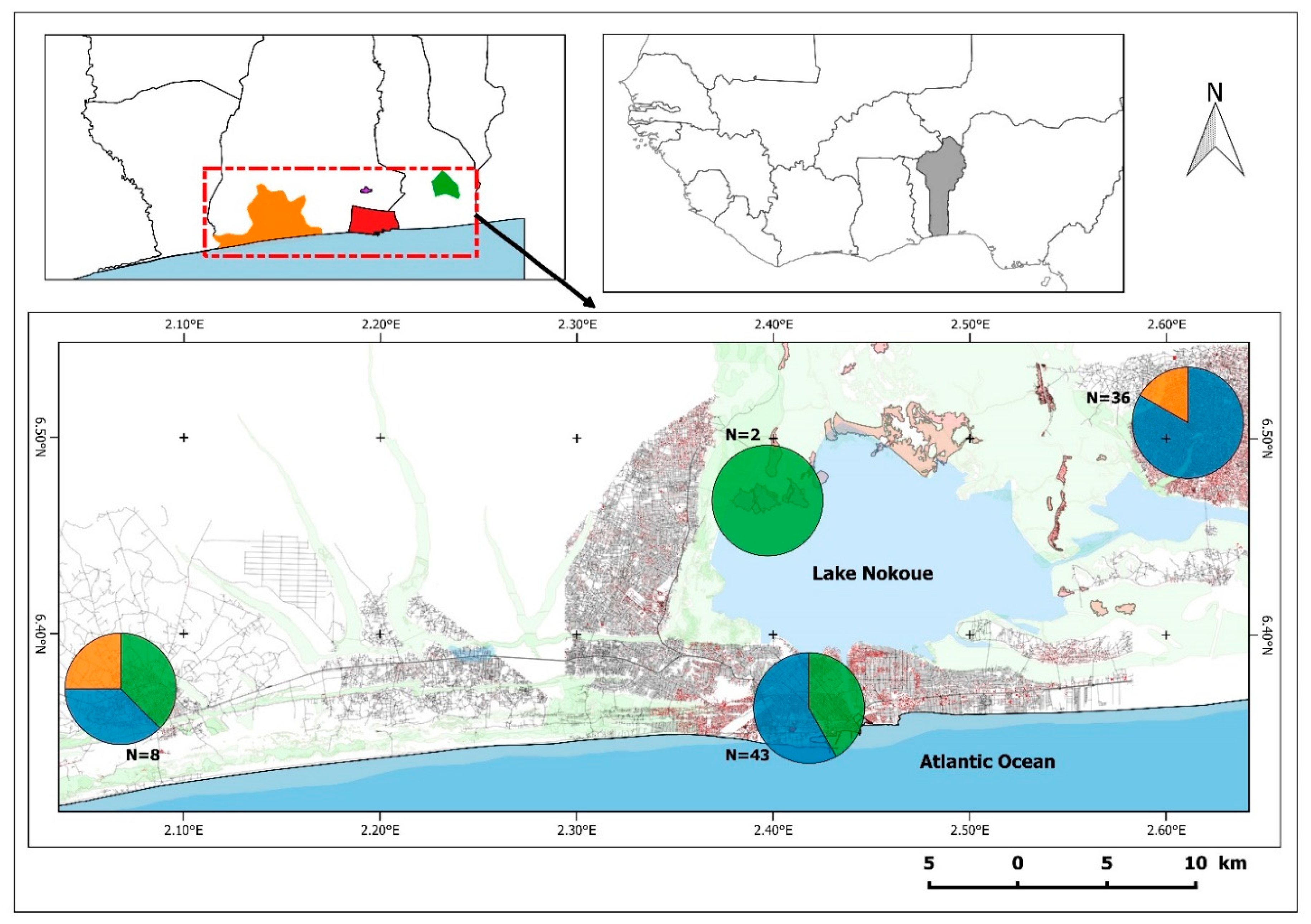
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global morbidity and mortality of leptospirosis: A systematic review. PLoS Trop. Neg. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [PubMed]
- Bertherat, E.; Renaut, A.; Nabias, R.; Dubreuil, G.; Georges-Courbot, M.C. Leptospirosis and Ebola virus infection in five gold-panning villages in northeastern Gabon. Am. J. Trop. Med. Hyg. 1999, 60, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Mattar, S.; Tique, V.; Miranda, J.; Montes, E.; Garzon, D. Undifferentiated tropical febrile illness in Cordoba, Colombia: Not everything is dengue. J. Infect. Public Health 2017, 10, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.M.; Docha de Sa, D.A.; Turra, T.Z.; Ferreira Borges, N.M.; Nascimento, U.M.; Damasceno, E.A. Hantavirus pulmonary syndrome in Brasilia periphery: A diagnostic challenge. J. Infect. Dev. Ctries 2009, 3, 639–643. [Google Scholar] [CrossRef]
- World Health Organization. Investigation of Yellow Fever Epidemics in Africa; Field Guide; WHO/HSE/ EPR/2008.5; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Allan, K.J.; Biggs, H.M.; Halliday, J.E.B.; Kazwala, R.R.; Maro, V.P.; Cleaveland, S.; Crump, J.A. Epidemiology of leptospirosis in Africa: A systematic review of a neglected zoonosis and a paradigm for ‘One Health’ in Africa. PLoS Trop. Neg. Dis. 2015, 9, e0003899. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A. Animal leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar] [PubMed]
- Ko, A.I.; Reis, M.G.; Ribeiro Dourado, C.M.; Johnson, W.D.; Riley, L.W.; The Salvador Leptospirosis Study Group. Urban epidemic of severe leptospirosis in Brazil. Lancet 1999, 354, 820–825. [Google Scholar] [CrossRef]
- Cornwall, W. A plague of rats. Science 2016, 352, 912–915. [Google Scholar] [CrossRef]
- Lau, C.L.; Smythe, L.D.; Craig, S.B.; Weinstein, P. Climate change, flooding, urbanisation and leptospirosis: Fuelling the fire? Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 631–638. [Google Scholar] [CrossRef] [PubMed]
- De Vries, S.G.; Visser, B.J.; Nagel, I.M.; Goris, M.G.A.; Hartskeerl, R.A.; Grobush, M.P. Leptospirosis in Sub-Saharan Africa: A systematic review. Int. J. Infect. Dis. 2014, 28, 47–64. [Google Scholar] [CrossRef] [PubMed]
- ONU Habitat. L’état des Villes Africaines: Réinventer la Transition Urbaine; ONU Habitat: Nairobi, Kenya, 2014. [Google Scholar]
- Dobigny, G.; Gauthier, P.; Houéménou, G.; Choplin, A.; Dossou, H.J.; Badou, S.; Etougbétché, J.; Bourhy, P.; Koffi, S.; Durski, K.N.; et al. Leptospirosis and extensive urbanization in West Africa: A neglected and underestimated threat? Urban Sci. 2018, 2, 29. [Google Scholar] [CrossRef]
- Direction des Études Démographiques. In RGPH4: Que Retenir des Effectifs de la Population en 2013? République du Bénin, Institut National de Statistique et de l’Analyse Économique: Cotonou, Benin, 2015.
- Houemenou, G.; Ahmed, A.; Libois, R.; Hartskeerl, R.A. Leptospira spp. prevalence in small mammal populations in Cotonou, Benin. ISRN Epidemiol. 2013. [Google Scholar] [CrossRef]
- Bello, C.I. Prévalence et Facteurs Associés de la Séropositivité à la Leptospirose dans Certains Quartiers de Cotonou (Bénin). Ph.D. Thesis, Abomey-Calavi University, Cotonou, Benin, 2011. [Google Scholar]
- Houéménou, G. Les Petits Mammifères de la Ville de Cotonou (Bénin): Peuvent-ils Constituer un Risque pour la Santé Humaine? Ph.D. Thesis, Liège University, Liège, Belgium, 2013. [Google Scholar]
- Houngbo, P.T.; N’Gouizé, J. Premiers Résultats de Dépistage Sérologique de la Leptospirose à Cotonou et Ses Environs; DIT Report, TBH/ABM, Cotonou, Benin; CPU/UNB: Fredericton, NB, Canada; Saint John, NB, Canada, 1995. [Google Scholar]
- Artige, E. Collections Taxonomiques, Collections Vivantes et Ressources Génétiques pour la Biodiversité; Technical and Research Document; Agropolis International: Montpellier, France, 2013; ISSN 1628-4240. [Google Scholar]
- Dobigny, G.; Garba, M.; Tatard, C.; Loiseau, A.; Galan, M.; Kadaouré, I.; Rossi, J.P.; Picardeau, M.; Bertherat, E. Urban market gardening and rodent-borne pathogenic Leptospira in arid zones: A case study in Niamey, Niger. PLoS Trop. Neg. Dis. 2015, 9, e4097. [Google Scholar] [CrossRef] [PubMed]
- R Project for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 1 April 2018).
- Meerburg, B.; Singleton, G.R.; Kijlstra, H. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Rahelinirina, S.; Leon, A.; Hartskeerl, R.A.; Sertour, N.; Ahmed, A.; Raharimanana, C.; Ferquel, E.; Garnier, M.; Chartier, L.; Duplantier, J.M.; et al. First isolation and direct evidence for the existence of large small mammal reservoirs of Leptospira sp. in Madagascar. PLoS ONE 2010, 5, e14111. [Google Scholar] [CrossRef] [PubMed]
- Maciel, E.A.P.; de Carvalho, A.L.F.; Nascimento, S.F.; de Matos, R.B.; Gouveia, E.L.; Reis, M.G.; Ko, A.I. Household transmission of Leptospira infection in urban slum communities. PLoS Trop. Neg. Dis. 2008, 2, e154. [Google Scholar] [CrossRef] [PubMed]
- Hagan, J.E.; Moraga, P.; Costa, F.; Capian, N.; Ribeiro, F.S.; Wunder, E.A.; Felzemburgh, R.D.M.; Reis, R.B.; Nery, N.; Sanata, F.S.; et al. Spatio-temporal determinants of urban leptospirosis transmission: Four-year prospective cohort study of slum residents in Brazil. PLoS Trop. Neg. Dis. 2016, 10, e0004275. [Google Scholar] [CrossRef]
- Himsworth, C.G.; Bidulka, J.; Parsons, K.L.; Feng, A.Y.T.; Tang, P.; Jardine, C.M.; Kerr, T.; Mak, S.; Robinson, J.; Patrick, D.M. Ecology of Leptospira interrogans in Norway rats (Rattus norvegicus) in an inner-city neighborhood of Vancouver, Canada. PLoS Trop. Neg. Dis. 2013, 7, e2270. [Google Scholar] [CrossRef]
- Tacunduva da Faria, M.; Calderwood, M.S.; Athanazio, D.A.; McBride, A.J.A.; Harstkerl, R.A.; Pereira, M.M.; Ko, A.I.; Reis, M.G. Carriage of Leptospira interrogans among domestic rats from an urban setting endemic for leptospirosis in Brazil. Acta Trop. 2008, 108, 1–5. [Google Scholar] [CrossRef]
- Taylor, P.J.; Arntzen, L.; Hayter, M.; Iles, M.; Frean, J.; Belmain, S. Understanding and managing sanitary risks due to rodent zoonoses in an African city: Beyond the Boston model. Integr. Zool. 2008, 3, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Halliday, J.E.B.; Knobel, D.L.; Allan, K.J.; Bronsvoort, B.M.; Handel, I.; Agwanda, I.; Agwanda, B.; Cutler, S.J.; Olack, B.; Ahmed, A.; et al. Urban leptospirosis in Africa: A cross-sectional survey of Leptospira infection in rodents in the Kibera urban settlement, Nairobi, Kenya. Am. J. Trop. Med. Hyg. 2013, 89, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.; ter Meulen, A.; Fichet-Calvet, E.; Koivogui, L.; Sylla, O.; Goris, M.; Hartskeerl, R.A.; ter Meulen, J. Seroprevalence and reservoirs of leptospirosis in Conakry (Guinea). Int. J. Med. Microbiol. 2007, 297, 147–148. [Google Scholar] [CrossRef]
- Bonfoh, B.; Mwachui, M.; Akpatou, B.; Zinsstag, J. Combined assessment of leptospirosis exposure in humans and rodents in Abidjan, Côte d’Ivoire. Int. J. Infect. Dis. 2014, 21, 220. [Google Scholar] [CrossRef]
- Dietrich, M.; Wilkinson, D.A.; Soarimalala, V.; Goodman, S.M.; Dellagi, K.; Tortosa, P. Diversification of an emerging pathogen in a biodiversity hotspot: Leptospira in endemic small mammals of Madagascar. Mol. Ecol. 2014, 23, 2783–2796. [Google Scholar] [CrossRef] [PubMed]
- Guernier, V.; Richard, V.; Nhan, T.; Rouault, E.; Tessier, A.; Musso, D. Leptospira diversity in animals and humans in Tahiti, French Polynesia. PLoS Trop. Neg. Dis. 2017, 11, e0005676. [Google Scholar] [CrossRef] [PubMed]
- Cosson, J.F.; Picardeau, M.; Mielcarek, M.; Tatard, C.; Chaval, Y.; Suputtamongkol, Y.; Buchy, P.; Jittapalapong, S.; Herbreteau, V.; Morand, S. Epidemiology of Leptospira transmitted by rodents in Southeast Asia. PLoS Trop. Neg. Dis. 2014, 8, e2902. [Google Scholar] [CrossRef] [PubMed]
- Houéménou, G.; Kassa, B.; Libois, R. Ecologie, diversité spécifique et abundance des petits mammifères de la ville de Cotonou au Bénin (Afrique de l’Ouest). Int. J. Biol. Chem. Sci. 2014, 8, 1202–1213. [Google Scholar] [CrossRef]
- Dossou, H.J.; Houéménou, G.; Tenté, B.; Mensah, G.A. Inventaire des rongeurs et de leurs ectoparasites en milieu urbain au Bénin: Transmission possible de zoonoses. Ann. Soc. Agron. 2015, 19, 545–574. [Google Scholar]
- Jobbins, S.E.; Alexander, K.A. Evidence of Leptospira sp. infection among a diversity of African wildlife species: Beyond the usual suspects. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 349–351. [Google Scholar] [CrossRef]
- Guernier, V.; Lagadec, E.; Cordonin, C.; Le Minter, G.; Gomard, Y.; Pagès, F.; Jaffar-Bandjee, M.C.; Michault, A.; Tortosa, P.; Dellagi, K. Human leptospirosis on Reunion Island, Indian Ocean: Are rodents the (only) ones to blame? PLoS Negl. Trop. Dis. 2016, 10, e0004733. [Google Scholar] [CrossRef] [PubMed]
- Roqueplo, C.; Marié, J.L.; André-Fontaine, G.; Kodjo, A.; Davoust, B. Serological survey of canine leptospirosis in three countries of tropical Africa: Sudan, Gabon and Ivory Coast. Comp. Immunol. Microbiol. Infect. Dis. 2015, 38, 57–61. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zone | Locality/Site | Environment | GPS | All | Rra | Rno | Mna | Cro | Other | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lat | Long | N | Pos | N | Pos | N | Pos | N | Pos | N | Pos | N | Pos | |||
| Cotonou | Cotonou/Abokicodji lagune | Urban | 6.363 | 2.442 | 16 | 1 | 12 | 0 | 3 | 0 | 1 | 1 | ||||
| Cotonou/Adogléta | Urban | 6.381 | 2.438 | 5 | 0 | 2 | 0 | 1 | 0 | 2 | 0 | |||||
| Cotonou/Agbato | Urban | 6.390 | 2.439 | 8 | 4 | 5 | 1 | 3 | 3 | |||||||
| Cotonou/Agontinkon | Urban | 6.374 | 2.404 | 5 | 2 | 4 | 2 | 1 | 0 | |||||||
| Cotonou/Ahouansori | Urban | 6.388 | 2.423 | 14 | 0 | 13 | 0 | 1 | 0 | |||||||
| Cotonou/Aïbatin | Urban | 6.358 | 2.363 | 5 | 0 | 5 | 0 | |||||||||
| Cotonou/Avotrou | Urban | 6.389 | 2.476 | 27 | 5 | 19 | 2 | 7 | 3 | 1 | 0 | |||||
| Cotonou/Ayimlofidé | Urban | 6.392 | 2.434 | 30 | 2 | 28 | 2 | 2 | 0 | |||||||
| Cotonou/Bokossi Tokpa | Urban | 6.365 | 2.438 | 9 | 0 | 2 | 0 | 5 | 0 | 2 | 0 | |||||
| Cotonou/Dandji | Urban | 6.373 | 2.477 | 2 | 2 | 1 | 1 | 1 | 1 | |||||||
| Cotonou/Dédokpo | Urban | 6.369 | 2.441 | 13 | 0 | 7 | 0 | 4 | 0 | 2 | 0 | |||||
| Cotonou/Djidjé | Urban | 6.384 | 2.434 | 7 | 0 | 3 | 0 | 2 | 0 | 2 | 0 | |||||
| Cotonou/Enagnon | Urban | 6.362 | 2.453 | 14 | 2 | 9 | 1 | 2 | 1 | 1 | 0 | 2 | 0 | |||
| Cotonou/Fidjrossé | Urban | 6.350 | 2.370 | 6 | 0 | 4 | 0 | 2 | 0 | |||||||
| Cotonou/Fifadji | Urban | 6.395 | 2.398 | 41 | 4 | 39 | 3 | 2 | 1 | |||||||
| Cotonou/Finagnon | Urban | 6.361 | 2.476 | 4 | 0 | 4 | 0 | |||||||||
| Cotonou/Marché Ganhi | Urban | 6.354 | 2.437 | 11 | 3 | 7 | 3 | 4 | 0 | |||||||
| Cotonou/Gankpodo | Urban | 6.393 | 2.456 | 15 | 1 | 12 | 0 | 1 | 1 | 2 | 0 | |||||
| Cotonou/Gbadji | Urban | 6.372 | 2.388 | 2 | 0 | 2 | 0 | |||||||||
| Cotonou/Gbénonkpo | Urban | 6.382 | 2.399 | 2 | 0 | 1 | 0 | 1 | 0 | |||||||
| Cotonou/Godomey | Urban | 6.413 | 2.312 | 13 | 0 | 12 | 0 | 1 | 0 | |||||||
| Cotonou/Haie Vive | Urban | 6.357 | 2.399 | 1 | 0 | 1 | 0 | |||||||||
| Cotonou/Houénoussou | Urban | 6.358 | 2.364 | 2 | 1 | 1 | 1 | 1 | 0 | |||||||
| Cotonou/Houéyiho | Urban | 6.367 | 2.387 | 1 | 0 | 1 | 0 | |||||||||
| Cotonou/Kowégbo | Urban | 6.387 | 2.469 | 12 | 0 | 11 | 0 | 1 | 0 | |||||||
| Cotonou/Kpankpan | Urban | 6.373 | 2.439 | 11 | 2 | 4 | 0 | 4 | 2 | 3 | 0 | |||||
| Cotonou/Ladji | Urban | 6.389 | 2.433 | 11 | 1 | 7 | 1 | 3 | 0 | 1 | 0 | |||||
| Cotonou/Maher | Urban | 6.392 | 2.434 | 3 | 0 | 3 * | 0 | |||||||||
| Cotonou/Marché Dantokpa | Urban | 6.374 | 2.430 | 15 | 1 | 12 | 1 | 2 | 1 | 1 | 0 | |||||
| Cotonou/Maro militaire | Urban | 6.363 | 2.421 | 3 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | |||||
| Cotonou/Minonchou | Urban | 6.391 | 2.457 | 14 | 2 | 11 | 2 | 1 | 0 | 1 | 0 | 1 | 0 | |||
| Cotonou/PAC (harbour) | Urban | 6.348 | 2.431 | 41 | 11 | 24 | 8 | 11 | 3 | 1 | 0 | 5 ** | 0 | |||
| Cotonou/Sèdami | Urban | 6.370 | 2.420 | 2 | 0 | 1 | 0 | 1 | 0 | |||||||
| Cotonou/Sodjatinmin | Urban | 6.368 | 2.456 | 1 | 0 | 1 | 0 | |||||||||
| Cotonou | Cotonou/Saint Jean | Urban | 6.363 | 2.418 | 4 | 0 | 4 | 0 | ||||||||
| Cotonou/Saint Jacques | Urban | 6.358 | 2.457 | 5 | 0 | 3 | 0 | 1 | 0 | 1 | 0 | |||||
| Cotonou/Suru Léré | Urban | 6.382 | 2.462 | 20 | 10 | 19 | 9 | 1 | 1 | |||||||
| Cotonou/Tchankpamé | Urban | 6.378 | 2.486 | 11 | 2 | 8 | 1 | 1 | 1 | 2 | 0 | |||||
| Togbin | Peri-urban | 6.357 | 2.302 | 42 | 0 | 6 | 0 | 35 | 0 | 1 | 0 | |||||
| Cotonou/Tokpa Hoho | Urban | 6.365 | 2.434 | 8 | 1 | 3 | 0 | 4 | 1 | 1 | 0 | |||||
| Cotonou/Vossa Kpodji | Urban | 6.397 | 2.400 | 27 | 2 | 22 | 2 | 3 | 0 | 1 | 0 | 1 | 0 | |||
| Cotonou/Wlacodji | Urban | 6.351 | 2.442 | 8 | 1 | 3 | 0 | 2 | 1 | 1 | 0 | 2 | 0 | |||
| Cotonou/Zogbohoué | Urban | 6.379 | 2.389 | 25 | 2 | 23 | 2 | 1 | 0 | 1 | 0 | |||||
| Total | 516 | 62 | 338 | 38 | 64 | 12 | 72 | 12 | 34 | 1 | 8 | 0 | ||||
| Ganvié | Ganvié | Lacustrial | 6.468 | 2.397 | 41 | 9 | 35 | 6 | 5 | 3 | 1 | 0 | ||||
| Total | 41 | 9 | 35 | 6 | 5 | 3 | 1 | 0 | ||||||||
| Porto Novo | Porto-Novo/Adjinan | Urban | 6.470 | 2.614 | 17 | 1 | 13 | 0 | 4 | 1 | ||||||
| Porto-Novo/Akonaboé | Urban | 6.514 | 2.605 | 13 | 4 | 1 | 0 | 2 | 0 | 10 | 4 | |||||
| Porto-Novo/Djegan-Daho | Urban | 6.487 | 2.651 | 24 | 7 | 4 | 1 | 1 | 1 | 9 | 2 | 10 | 3 | |||
| Porto-Novo/Dowa | Urban | 6.497 | 2.594 | 23 | 4 | 17 | 2 | 1 | 1 | 5 | 1 | |||||
| Porto-Novo/Gbékon | Urban | 6.469 | 2.635 | 29 | 7 | 17 | 3 | 7 | 2 | 5 | 2 | |||||
| Porto-Novo/Grand Marché | Urban | 6.475 | 2.630 | 39 | 10 | 12 | 8 | 27 | 2 | |||||||
| Porto-Novo/Hounsa | Urban | 6.511 | 2.634 | 11 | 1 | 2 | 1 | 1 | 0 | 2 | 0 | 6 | 0 | |||
| Porto-Novo/Marché Ouando 1 | Urban | 6.505 | 2.612 | 37 | 1 | 32 | 1 | 5 | 0 | |||||||
| Porto-Novo/Marché Ouando 2 | Urban | 6.508 | 2.611 | 31 | 5 | 29 | 5 | 2 | 0 | |||||||
| Porto-Novo/Zounkpa | Urban | 6.484 | 2.649 | 4 | 1 | 1 | 1 | 3 | 0 | |||||||
| Total | 228 | 41 | 127 | 21 | 4 | 1 | 30 | 10 | 67 | 9 | ||||||
| Ouidah | Savi/Minantinkpon | Peri-urban | 6.383 | 2.091 | 5 | 3 | 3 | 2 | 2 *** | 1 | ||||||
| Savi/Houéton | Rural | 6.428 | 2.104 | 20 | 0 | 10 | 0 | 8 | 0 | 2 | 0 | |||||
| Gakpé | Rural | 6.435 | 2.111 | 50 | 1 | 41 | 0 | 7 | 1 | 2 | 0 | |||||
| Pahou/Marché de Pahou | Urban | 6.384 | 2.208 | 32 | 3 | 14 | 0 | 6 | 3 | 2 | 0 | 10 | 0 | |||
| Pahou/Adjarra | Peri-urban | 6.409 | 2.200 | 26 | 1 | 19 | 0 | 6 | 1 | 1 | 0 | |||||
| Ouidah/Gbénan | Urban | 6.372 | 2.068 | 18 | 0 | 17 | 0 | 1 | 0 | |||||||
| Ouidah/Marché de Zobé | Urban | 6.359 | 2.087 | 17 | 1 | 9 | 0 | 3 | 0 | 5 | 1 | |||||
| Ouidah/Marché de Kpassé | Urban | 6.374 | 2.090 | 18 | 2 | 4 | 0 | 14 | 2 | |||||||
| Total | 186 | 11 | 110 | 0 | 6 | 3 | 34 | 4 | 34 | 3 | 2 | 1 | ||||
| Total | 971 | 123 | 610 | 65 | 79 | 19 | 136 | 26 | 136 | 13 | 10 | 1 | ||||
| Zone | Site | Cro | Mna | Rno | Rra |
|---|---|---|---|---|---|
| Ouidah | Pahou Adjarra | 1 int | |||
| Marché Zobé | 1 kir | ||||
| Marché Pahou | 1 bor + 2 int | ||||
| Minantinkpon | 1 bor | ||||
| Gakpé | 1 bor | ||||
| Marché Kpassé | 1 kir | ||||
| Cotonou | Abokicodji | 1 int | |||
| Agbato | 2 bor | ||||
| Agontikon | 2 bor | ||||
| Avotrou | 2 bor | 1 bor | |||
| Dandji | 1 bor | ||||
| Enagnon | 1 int | 1 int | |||
| Fifadji | 1 int | 1 bor | |||
| Ganhi | 2 int | ||||
| Gankpodo | 1 bor | ||||
| Houenoussou | 1 bor | ||||
| Kpankpan | 2 bor | ||||
| Marché Tokpa | 1 int | ||||
| PAC | 1 bor + 1 int | 6 int | |||
| Suru Léré | 1 bor | 6 bor + 1 int | |||
| Tchankpamé | 1 bor | ||||
| Tokpa Hoho | 1 int | ||||
| Vossa Kpodji | 1 bor + 1 int | ||||
| Wlacodji | 1 int | ||||
| Zogbohoué | 2 bor | ||||
| Ganvié | Ganvié | 2 int | |||
| Porto-Novo | Adjinan | 1 kir | |||
| Ouando 1 | 1 bor | ||||
| Ouando 2 | 5 bor | ||||
| Grand Marché | 1 kir + 1 bor | 8 bor | |||
| Djégan-Daho | 3 kir | 2 bor | 2 bor | ||
| Dowa | 1 kir | 1 bor | 2 bor | ||
| Akonaboé | 2 bor | ||||
| Gbékon | 2 bor | 2 bor | 2 bor |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houéménou, G.; Gauthier, P.; Etougbétché, J.; Badou, S.; Dossou, H.-J.; Agossou, D.; Picardeau, M.; Dobigny, G. Pathogenic Leptospira in Commensal Small Mammals from the Extensively Urbanized Coastal Benin. Urban Sci. 2019, 3, 99. https://0-doi-org.brum.beds.ac.uk/10.3390/urbansci3030099
Houéménou G, Gauthier P, Etougbétché J, Badou S, Dossou H-J, Agossou D, Picardeau M, Dobigny G. Pathogenic Leptospira in Commensal Small Mammals from the Extensively Urbanized Coastal Benin. Urban Science. 2019; 3(3):99. https://0-doi-org.brum.beds.ac.uk/10.3390/urbansci3030099
Chicago/Turabian StyleHouéménou, Gualbert, Philippe Gauthier, Jonas Etougbétché, Sylvestre Badou, Henri-Joël Dossou, David Agossou, Mathieu Picardeau, and Gauthier Dobigny. 2019. "Pathogenic Leptospira in Commensal Small Mammals from the Extensively Urbanized Coastal Benin" Urban Science 3, no. 3: 99. https://0-doi-org.brum.beds.ac.uk/10.3390/urbansci3030099