1. Introduction
Rice (
Oryza sativa L.) is an important food crop and source of income worldwide. In Africa, the rate of consumption of rice is increasing faster than most other staple food crops. For instance, between 2000 and 2010, the increase was estimated at 5.5% per year [
1]. This increased demand for rice has been accompanied by a rapid expansion in cultivated rice area of between 0.4–7.7% per year across sub-Saharan Africa (SSA) [
1]. Upland rice can be rotated with other crops, water and labor inputs (because it is not transplanted) are low compared to lowland rice, and above all, it often fetches higher returns on investment than most other crops [
2]. However, many rice-growing countries in Africa remain heavily reliant on imports to meet domestic rice demand [
1] and will thus need to increase the area under cultivation and more importantly, the production per unit area, to become more self-sufficient. Upland rice yield potential is not realized worldwide because production is often limited to less fertile and drought-prone lands [
3].
Nitrogen (N) management, in addition to water management, differs considerably across rice cropping systems [
3]. In Africa, low N application rates are predominant in upland rice production and rice mono-cropping is common. Nitrogen deficiencies are also common in upland rice because of uneven rainfall distribution during the growing season [
3,
4]. The low nitrogen use efficiency (NUE) levels (15–20 kg grain kg
−1 N applied) reported for rainfed upland rice [
5,
6] means that approximately 50–67 kg N is required for each 1 Mg of grain yield, which is costly for the average SSA rice farmer.
Studies on N fertilization, including under favorable growing conditions, are lacking for upland rice [
6]. In South Africa, some desktop feasibility studies reported upland rice production as viable under rainfed conditions, while others concluded that it was not viable [
7]. Previously, rice had also been grown successfully on a small scale in the KwaZulu-Natal Province [
8]. In Asia, Belder et al. [
5] reported that fertilization at 150 kg N ha
−1 improved water use efficiency (WUE) of irrigated rice from 4.0 to 7.5 kg mm
−1 (based on volume of water applied). Past N fertilization studies neglected water use when fertilizer rates were varied. Moreover, Belder et al. [
5] speculated that rice growth and yield were limited by water deficits at high N level, probably because N leads to increased transpiration [
9]. Kondo et al. [
4] observed that water uptake of potted rice under aerobic conditions increased with N fertilization, suggesting a modification in crop water demand by N. Thus, previously reported N recommendations may be too low to achieve high WUE levels.
Irrigating according to soil water depletion is a recommended practice to improve rainfall use efficiency and maximize yield [
10]. As rice has a shallow root system (<0.4 m depth) [
11], N management can be challenging, and as a result, it is important to optimize N applications to avoid nitrate (NO
3−) leaching and reduce production costs. Understanding the factors that enable high NUE and the relationship between crop water use/N uptake and growth is needed to close yield gaps in such cropping systems [
9]. It was therefore hypothesized that: (I) N fertilization increases seasonal water use of upland rice, and water management should thus be tailored to N availability; (II) the depth of water extraction is not affected by varying N rates when water is not limiting; and (III) the highest WUE and NUE can be achieved at the highest N rate. The overall aim of this study was to improve our understanding of upland rice response to N fertilizer rates under well-watered conditions to address knowledge gaps regarding optimal N application rates and seasonal water requirements.
3. Results
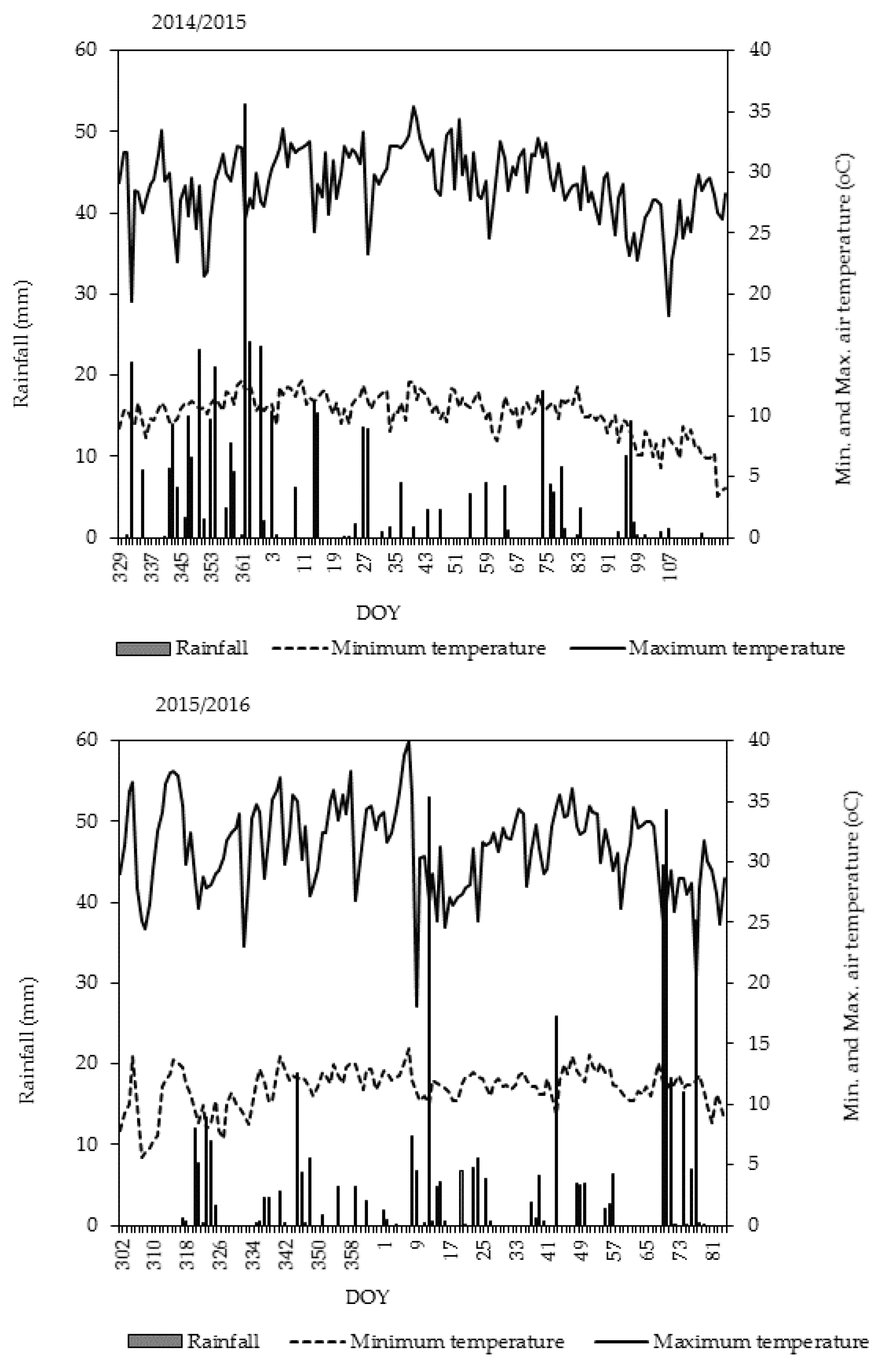
3.1. Seasonal Weather Conditions and Crop Water Demand during Growth
Figure 1 shows the daily weather data from sowing to harvest for selected parameters during the two seasons. Maximum and minimum temperatures averaged over the growing periods (29.0 °C and 15.3 °C in Y
1 and 31.2 °C and 13.7 °C in Y
2, respectively), were favorable for rice growth. Rainfall received during the period 52–112 DAE was 105 mm in the Y
1 and 175 mm in Y
2. Rainfall was more biased toward the early season in Y
1 and to the late season in Y
2 (
Figure 1). During day of the year (DOY) 69–77 in 2016 (late reproductive growth), heavy rainfall events coincided with the ripening stage (Y
2). In contrast, rainfall was more frequent during DOY 346–362 in 2014 (early vegetative growth of Y
1). The total amount received from sowing (DOY 324) to 20 DAE in Y
1 was 155 mm, compared to 48 mm for the same growth stage in Y
2. Although sowing was earlier (DOY 293) in Y
2 than in Y
1 (DOY 333), the cumulative amount of solar radiation during the season differed by only 77.5 MJ m
−2.
Irrigation amount was lower in Y
1 (130 mm) than in Y
2 (234 mm) due to more rainfall received in Y
1. Amount of irrigation water applied for a given N rate did not significantly (
p > 0.05, paired
t-test) differ between seasons. Although not consistent, irrigation amount generally tended to increase with increasing N rates (
Table 2). It was higher for 120 kg N ha
−1 (385 mm) than for 160 kg N ha
−1 (361 mm) and lowest for the zero-N treatment (336.5 mm).
Table 2 also shows that the zero-N treatment required a substantially lower irrigation amount, notably during vegetative growth in Y
1 and during reproductive growth in Y
2. Mean soil water content measured in the 0–0.4 m and 0–0.6 m layers at the end of each season for the zero-N and 40 kg N ha
−1 treatments was generally higher than for higher N rates, signifying that water extraction by plants was limited at lower N rates. These findings support the hypothesis that increasing N level increases crop water demand, even when water is not limiting growth.
One water meter was used per treatment and differences in irrigation amount between treatments could not be statistically separated.
A fairly constant soil water regime was maintained to avoid WS. However, it was important to note that θ was depleted from layers deeper than 0.4 m in plots receiving more than 80 kg N ha−1, notably during the reproductive growth stage. Soil water deficits for 160 kg N ha−1 were lower than for 120 kg N ha−1 over time across seasons. Results indicated that N levels above 120 kg N ha−1 reduced the amount of water extracted at maximum depth. Excessive N fertilization might have inhibited root growth and reduced water uptake despite the observed deep rooting and increased LAI at 160 kg N ha−1. These findings thus reject the hypothesis on the negligible effect of N on the depth of water extraction under well-watered conditions.
3.2. Crop Development, Leaf Area Index and Yield
Flowering for the 0 and 40 kg N ha−1 treatments was delayed (+6 days), and duration was shorter, compared to higher N rates, which could be because the low N fertilized plants took longer to achieve maximum tiller number. However, maturity was reached much earlier for these treatments than higher N rates during both years, resulting in a shorter grain filling period and lower final grain yields.
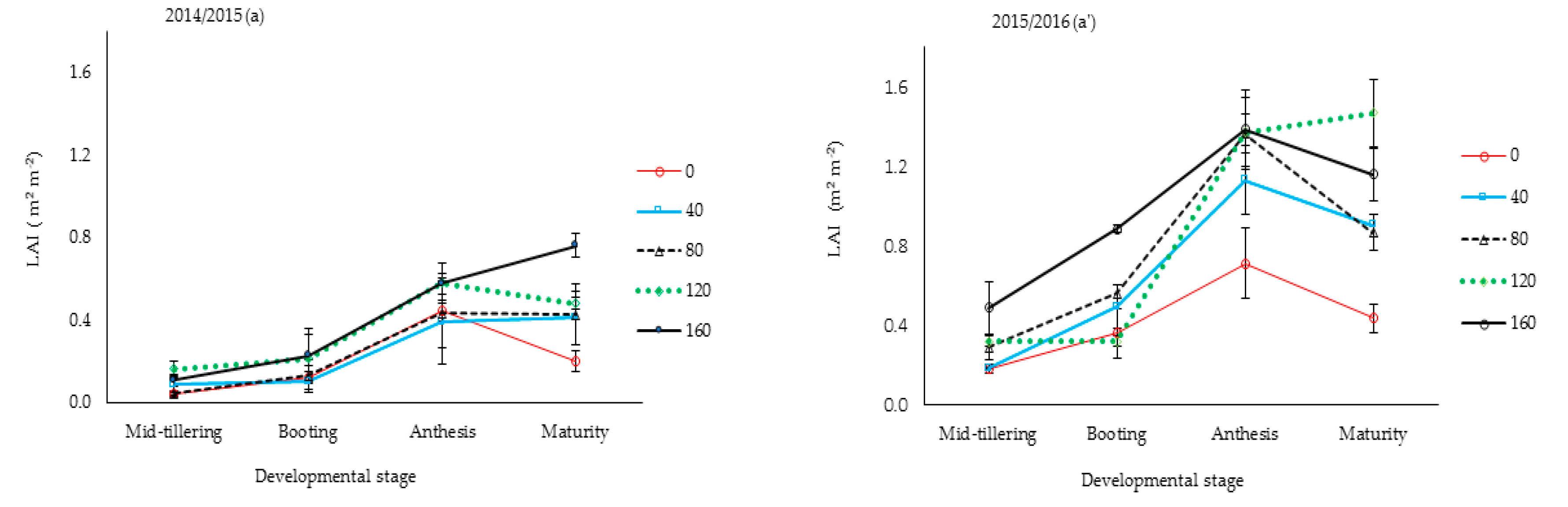
The interaction effect of season and N rates on growth (except for plant height) and TDM at maturity was not significant (
p > 0.05). Larger maximum LAI in Y
2 by 15–66%, compared to the same N treatment mean values than in Y
1 (
Figure 2), was attributed to inter-seasonal differences in daily temperatures.
Figure 2 also shows that LAI for the two highest N rates did not differ significantly from each other during the post-anthesis stage. Plants for the two highest N rates maintained greener canopies (stay green attribute) for longer periods than the other treatments.
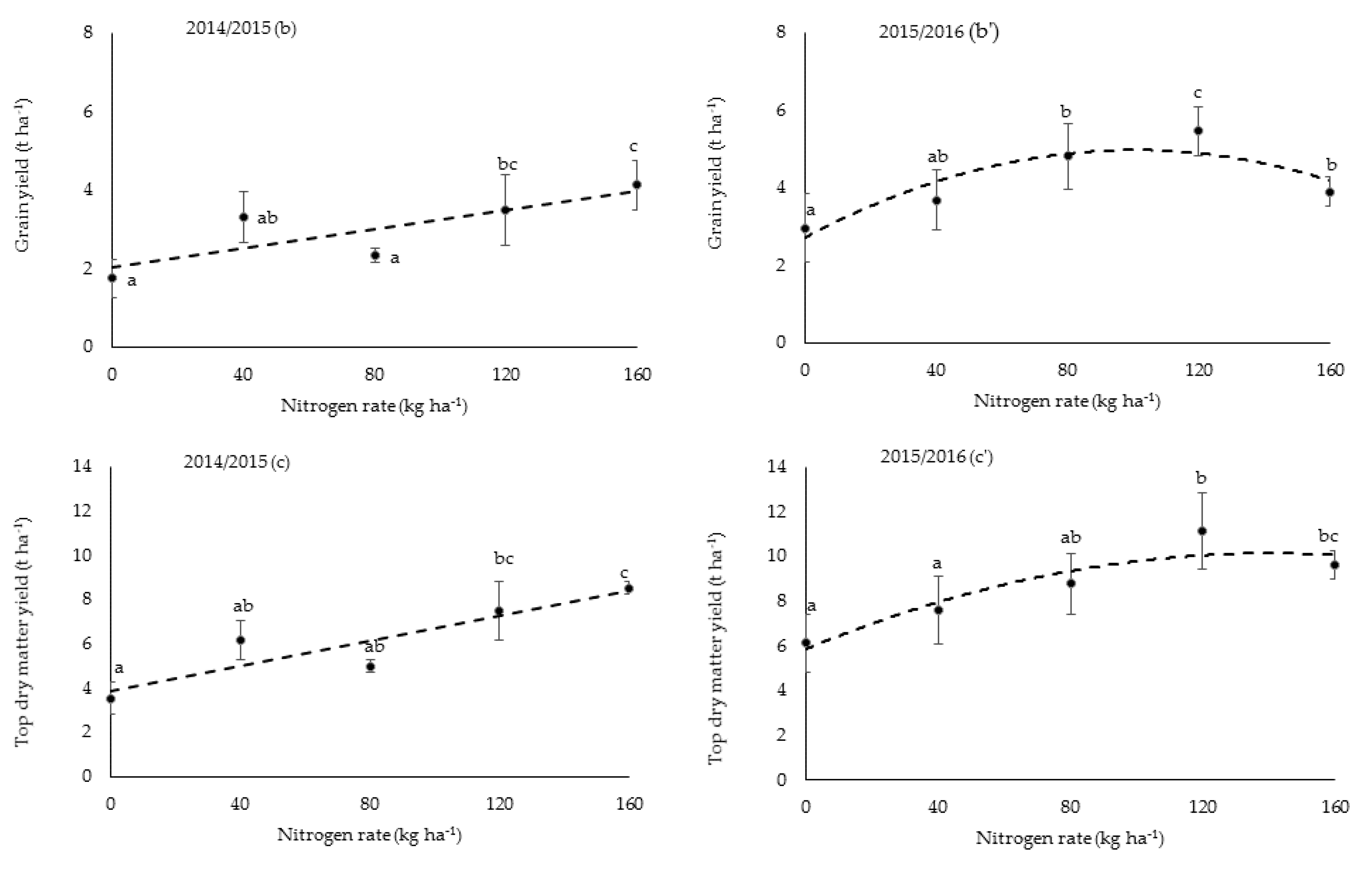
Grain yield (Y) increased significantly (p = 0.027) with increasing N rates (x), following a linear relationship () during Y1, and a quadratic relationship during Y2 with the maximum yield achieved at 120 kg N ha−1. High grain yield significantly correlated positively (p < 0.001) with the higher number of spikelets per panicle (59.3 ± 4.3 in Y1 and 92.1 ± 5.9 in Y2) and productive panicles m−2 (403.2 ± 82.5 in Y1 and 447.2 ± 73.7 in Y2) as a result of the larger LAI. Panicle length (17.0 ± 1.2 cm), 1000 grain mass (21.8 ± 0.6 g), and filled grains per panicle (7.0 ± 1.2%) were not different between seasons.
Correlation analysis revealed that grain yield variations due to N rates were closely linked to the number of spikelets (R2 = 0.91, p < 0.05) and panicles (R2 = 0.56, p < 0.05) per unit area. A reduction in grain yield at 160 kg N ha−1 in Y2 was linked to a significantly (p < 0.05) lower number of spikelets m−2 (45.63 × 103) than for the 120 kg N ha−1 treatment (64.30 × 103), lower leaf N concentration at maturity and more tillers, but a higher proportion of unproductive tillers.
3.3. Tiller Growth and Leaf N Concentrations
Mean tiller number was higher in Y
1 than in Y
2, except at final harvest for the zero-N treatment (
Table 3). The mean number of tillers counted during V growth, for instance at 65 DAE, was lowest (1.3 ± 0.1) for 0 kg N ha
−1, but not significantly (
p > 0.05) different from the other treatments. Tillering pattern over the season was very different between the seasons for the N-fertilized treatments. Tiller abortion, therefore, occurred only during Y
2, a season of normal tiller development, and was slightly higher at 0 kg N ha
−1 than for most N-fertilized treatments. During Y
2, maximum tiller number for 80–160 kg N ha
−1 was early (80 DAE) compared to 92 DAE for the lower N rates. The cause of tiller abortion was not mutual shading, because maximum canopy cover measured was low, at approximately 60% fractional interception.
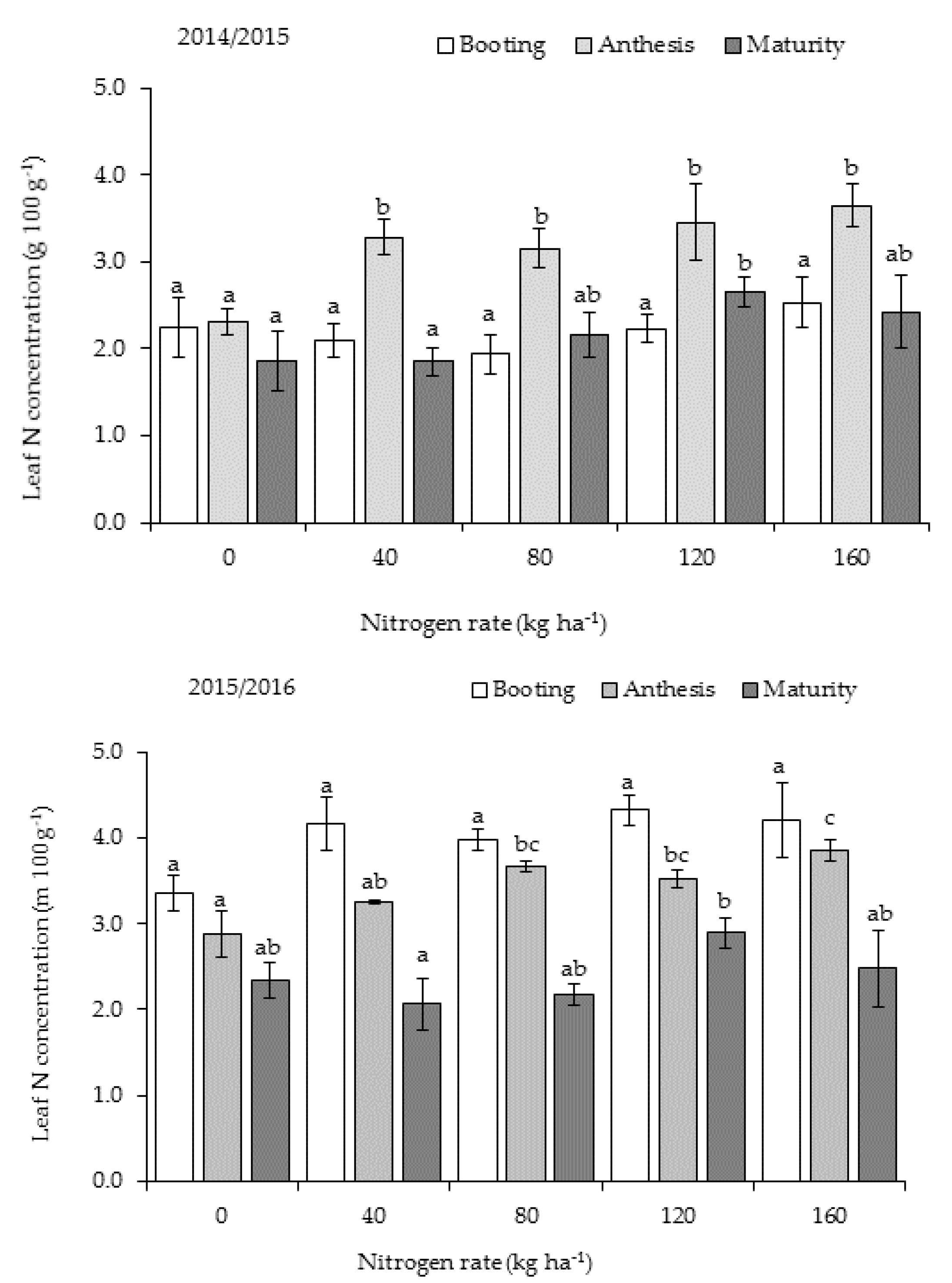
Leaf N concentration usually increased with increasing N application rate, independent of stage, although not always significant (
Figure 3). The highest N concentrations in Y
1 were measured during anthesis and in Y
2 during booting. Frequent rainfall events during the early vegetative growth in Y
1 (
Figure 1) may have caused anaerobic conditions that were detrimental to root growth and promoted denitrification, which could have resulted in reduced N uptake. In the lower leaves (without the flag leaf), N concentration was significantly higher (
p < 0.05) for 120 kg N ha
−1 (2.68 ± 0.16%) than other treatments, including 160 kg N ha
−1 (2.24 ± 0.08%). These findings suggest that the more N is applied, the more the crop takes up beyond what it needs for growth and grain production.
The decline in leaf N from booting to maturity was lower during Y1 than Y2, indicating that much N was retained in tillers (new tillers emerged post-flowering in Y1) and unavailable to the grains. The decline in tiller number post-flowering in Y2 at lower N rates was consistent, signifying a smaller maintenance burden at lower N rates, as there was less of a decline at higher N rate.
3.4. Water and N Use Efficiencies and Relationships and Estimated Crop N Uptake
Table 4 shows crop water use (ET), harvest index (HI), WUE, and NUE data as affected by N rates over the two growing seasons. The zero-N treatment consistently had the lowest ET. Although differences in total ET were negligible between N treatments, plants in the zero-N treatment had likely lower transpiration, but higher soil evaporation due to smaller LAI.
Grain quality did not change considerably with increasing N rate above 80 kg N ha−1. In addition to the yield response observed in Y2, the most efficient utilization of N fertilizer and water was thus achieved at a lower N rate than for maximum yield. It is thus important to achieve a balance between grain N concentration and yield when applying N fertilizers. Yield response was strongly affected by the recovery of N as indicated by a highly significant correlation (p < 0.001, R2 = 0.60) between apparent recovery and agronomic efficiency. Mean values of estimated AR indicate that the recovery of N was not affected by season and the interaction effect of N x season.
Grain N content per unit of water use increased with increasing N rate.
Table 4 shows that increasing N fertilizer rate from 40 to 160 kg N ha
−1 increased grain N uptake per unit crop water use from 8.7 to 10.6 kg N kg
−1 mm
−1 on average across years. Although N concentration in the stems was not analyzed, the highly significant (
p < 0.0001) stem DM yield differences measured at harvest (
Table 4) suggest that the total N retained in the stem parts for N fertilized treatments was high. The ratio of stem DM for N fertilized treatments to the zero-N treatment (1.8:1–2.8:1) suggests that stem N content for 120 kg N ha
−1 and 160 kg N ha
−1 was likely more than twice the level for 0 kg N ha
−1. It is thus likely that stems of plants receiving 120 kg N ha
−1 and 160 kg N ha
−1 contained more than twice the total amount of N of the zero-N treatment. Estimated total crop N uptake averaged 87.6 ± 29.5 kg N ha
−1 in Y
1 and 93.5 ± 23.3 kg N ha
−1 in Y
2. These results suggest that more N was locked up in the V parts. Hence, no further yield gains (though crop N uptake increased) would probably have been achieved above 160 kg N ha
−1.
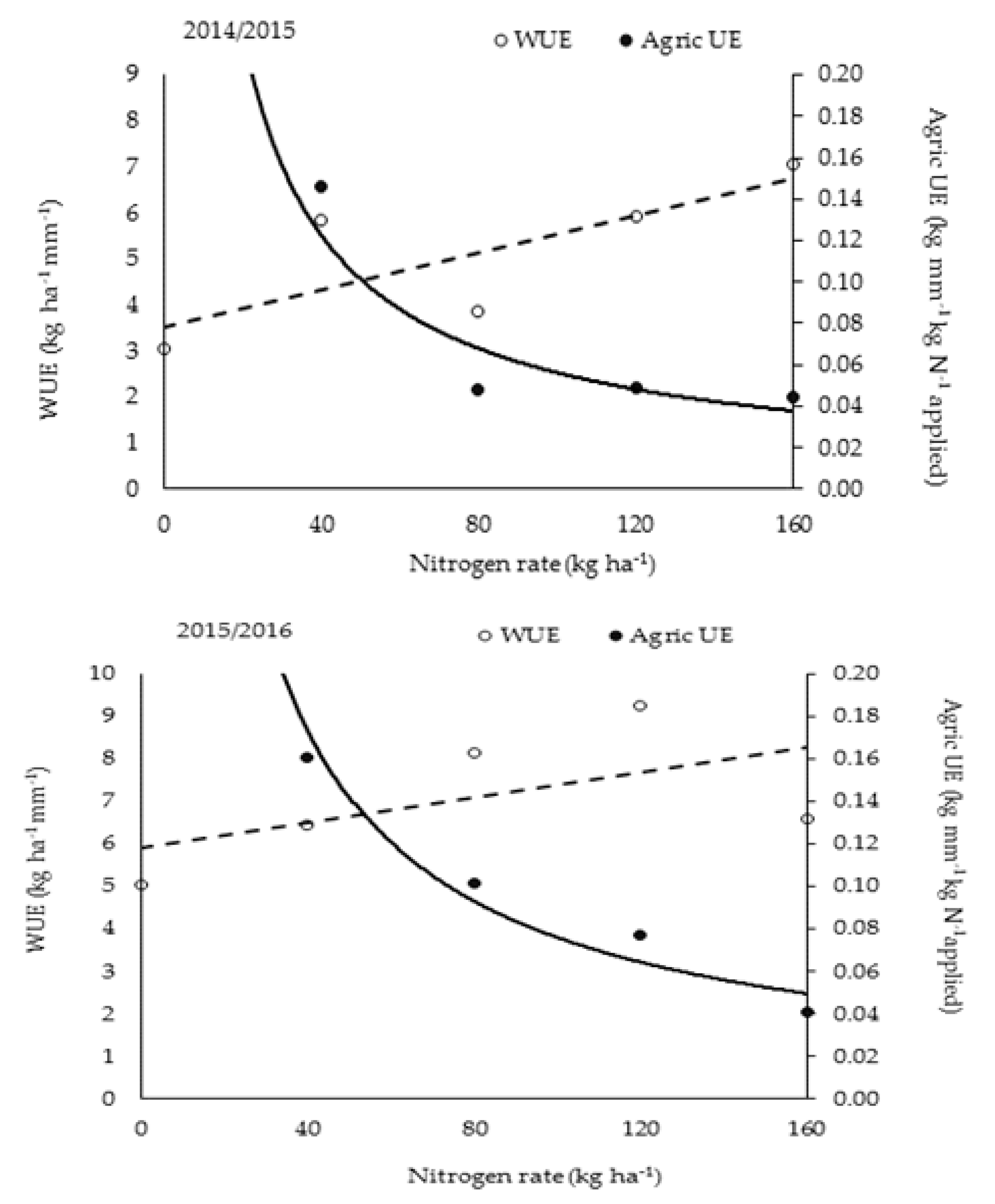
Agricultural UE decreased at a strongly (R
2 = 0.80 and 0.97) negative exponential rate with increase in N fertilizer rates in both years (
Figure 4). Mean values for 160 kg N ha
−1 treatment were the same in both years, despite a substantial reduction in grain yield for this rate in Y
2. Since grain quality did not change considerably with increasing N rate, except for 160 kg N ha
−1, the relationship indicates that to produce grain with 1.7% N (the grand average), lower N rates than for maximum yield will result in a more efficient utilization of N fertilizer and water. During both seasons, the intersection between the WUE and Agricultural UE functions occurred at a rate of ±55 kg N ha
−1 (
Figure 4), indicating that maximizing use efficiency for N and water simultaneously was achieved at this N rate.
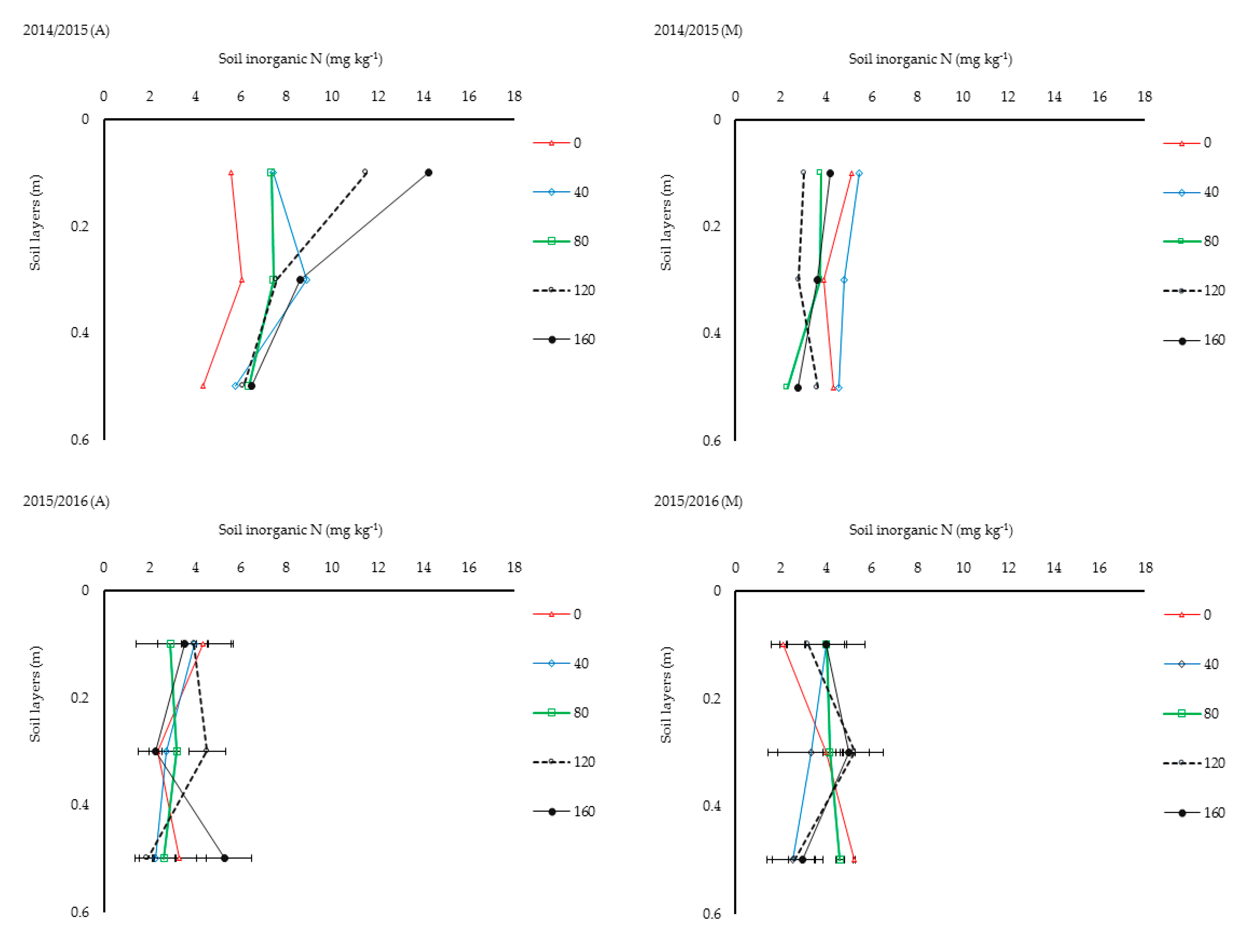
3.5. Soil Inorganic N Dynamics and Linkages with Soil Water Uptake
Soil inorganic N values in the top 0.6 m soil layer at harvest (2–8 mg kg
−1) across all treatments were generally lower than values measured at sowing (6.9–7.4 mg kg
−1). Differences in residual soil inorganic N levels were small in Y
2 compared to Y
1, most notably in the 0.4–0.6 m soil layer (
Figure 5), because late season rains (
Figure 1) probably diluted treatment effects. For the 0.4–0.6 m soil layer, plots without N fertilizer had higher mean soil inorganic N (4.32 mg kg
−1 in Y
1 and 5.23 mg kg
−1 in Y
2) than N fertilized treatments (3.31 ± 0.8 mg kg
−1 in Y
1 and 3.19 ± 0.8 mg kg
−1 in Y
2) at crop maturity. The high residual soil inorganic N for the 0 kg N ha
−1 treatment, approximately 16–47% more than the mean levels for the higher N rates in Y
1 and by 44–51% in Y
2, was most likely due to limited water uptake from deeper soil layers. This can probably be associated with poor root development and low root activity for this treatment. Furthermore, the lowest residual soil inorganic N measured for 120 kg N ha
−1, and the fact that the most soil water was depleted for this treatment, confirms that N fertilizer simultaneously increased water use and N uptake.
4. Discussion
Variation in rice yields between seasons for the same N rate was because canopy growth was greater during Y
2 than in Y
1. The inter-seasonal variations in LAI, growth, and yield shown in
Figure 2 could be explained by differences in rainfall patterns or distribution between years. The very wet conditions from frequent rainfall events in early Y
1, as shown in
Figure 1, likely affected N availability, which was reflected by low leaf N at booting in Y
1 compared to Y
2. Great variation in rice yields between seasons has been reported previously. Yoshida [
18] attributed inter-seasonal yield variation to differences in the available amount of solar radiation (R
s) during a season. There was no possibility of this effect in our study as cumulative R
s differed by only 77.5 MJ m
−2 between the seasons. The great inter-seasonal yield variation was a result of different rainfall patterns during the years.
The high grain yield in Y
2 compared to Y
1 was due to a generally higher number of spikelets per panicle and per unit area and productive panicles per square meter, as shown in
Table 3. The number of spikelets strongly depends on LAI at heading and the latter is positively related to crop N uptake at flowering [
18]. While yield dependence on spikelets m
−2 was significant in both years, the influence of leaf N was evident in Y
2. A reduction in grain yield at 160 kg N ha
−1 in Y
2 was linked to lower leaf N concentration at maturity and a lower number of spikelets than for the 120 kg N ha
−1 treatment. Furthermore, the diminishing increase in grain yields was caused by a higher proportion of unproductive tillers. In rice production, N is commonly applied during early vegetative stages to promote tillering and increase panicle number, followed by top-dressing around panicle initiation to increase spikelet number per panicle. Based on the findings of this study, it is important to promote reproductive growth to achieve high grain yield, because the increase in grain yield (which was often not significant between fertilized treatments) with increasing N rate was mainly attributed to more (but poorly filled) spikelets and more unproductive tillers.
Results on in-season leaf N concentrations suggest that the more N is applied, the more the crop takes up beyond it needs for growth and grain production, and the pattern for N-fertilized treatments can be altered by high soil water levels during tillering. It was expected that leaf N concentrations follow a normal pattern with time, which is initially high during early stages and decline gradually as the crop approached late developmental stages. In Y
1, the highest N concentrations were measured during anthesis, while in Y
2, the highest was at booting. The very wet soil conditions from frequent rainfall events for many days during the early vegetative growth stage (Y
1,
Figure 1) may have caused anaerobic conditions that were detrimental to root growth and promoted denitrification, which could have resulted in reduced N uptake. Nevertheless, findings in Y
1 showed that upland rice can still accumulate substantial N levels in the leaves during the reproductive growth stages, which were equal to levels in Y
2, even when N uptake is hampered during vegetative growth. Such ‘compensative uptake’ of N after flowering has not been documented for upland rice previously, but was observed in irrigated sugarcane (
Saccharum officinarum). Wood et al. [
19], for example, found late accumulation of leaf N just before maximum biomass production during the first season of a newly planted sugarcane crop.
The tillering patterns previously discussed (
Table 3) are strongly linked to N availability. In contrast to most crops, tillering in rice is widely spread over time. Tillers from a main or primary stem are at different chronological ages, and a substantial overlap between the vegetative and reproductive stages occurs [
11]. Uptake and utilization of N at a point in time can, therefore, differ considerably between a primary stem and secondary and tertiary tillers. This is similar to cotton (
Gossypium hirsutum L.), where the spread of boll production over time results in different N demands within a single cotton bush [
16]. Since the growth habit of rice is partly similar to that of cotton, competition for N between tillers in the vegetative growth stage as well as the grains can be expected, and the degree of competition depends on N availability. The decline in tiller number after flowering in Y
2 at lower N rates was consistent, signifying a smaller maintenance burden at lower N rates as there was less of a decline in higher N treatments. Differences in tillering between N-fertilized and zero-N treatments thus influenced the onset and duration of flowering, and to a lesser extent maturity. The present study findings on N influence on flowering contrast those of previous studies, for example, Belder et al. [
5] reported early maturity of aerobic rice in N-fertilized treatments under irrigation in a lowland. It is noteworthy that upland rice differs from lowland rice in several respects, among others, soil water availability.
The values of NUE and grain N in our study were slightly higher compared to the values (9–14 kg grain kg
−1 N for agronomic NUE, 1.13–1.28 g N 100 g
−1 for grain N, and 0.24–0.31 for HI) reported by Fageria et al. [
17] and Kaizzi et al. [
2]. One likely reason for the differences in NUE trends between studies is that the crops in the present study did not suffer from drought stress and the rice varieties were different.
The objective of N fertilization is to achieve high NUE and maximize profits. The pattern of Agricultural UE shown in
Figure 4, which was similar in both years, indicates that growth and resource use efficiencies declined with increasing N rate. The observed decline in Agric UE and in grain N per crop N uptake, means that N fertilizer should be optimized in irrigated rice production. The present study identified compensative uptake of N, increased rooting depth, and delayed senescence as specific physiological traits, which can be attributed to high NUE and sink capacity for this upland rice variety. However, it was noted that low panicle number due to few productive tillers in Y
1 occurred with the late accumulation of N in the leaves, which reduced grain yield and thus N utilization efficiency. Findings suggest that improving spikelet number through high planting density under well-watered conditions will compensate for low HI under high N rates and likely improve WUE.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}