Paraburkholderia phymatum STM815 σ54 Controls Utilization of Dicarboxylates, Motility, and T6SS-b Expression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Cultivation
2.2. Promoter Fusion Construction
2.3. RNA-Sequencing and Data Processing
2.4. Phenotypic Analysis
2.5. Statistical Analysis
3. Results
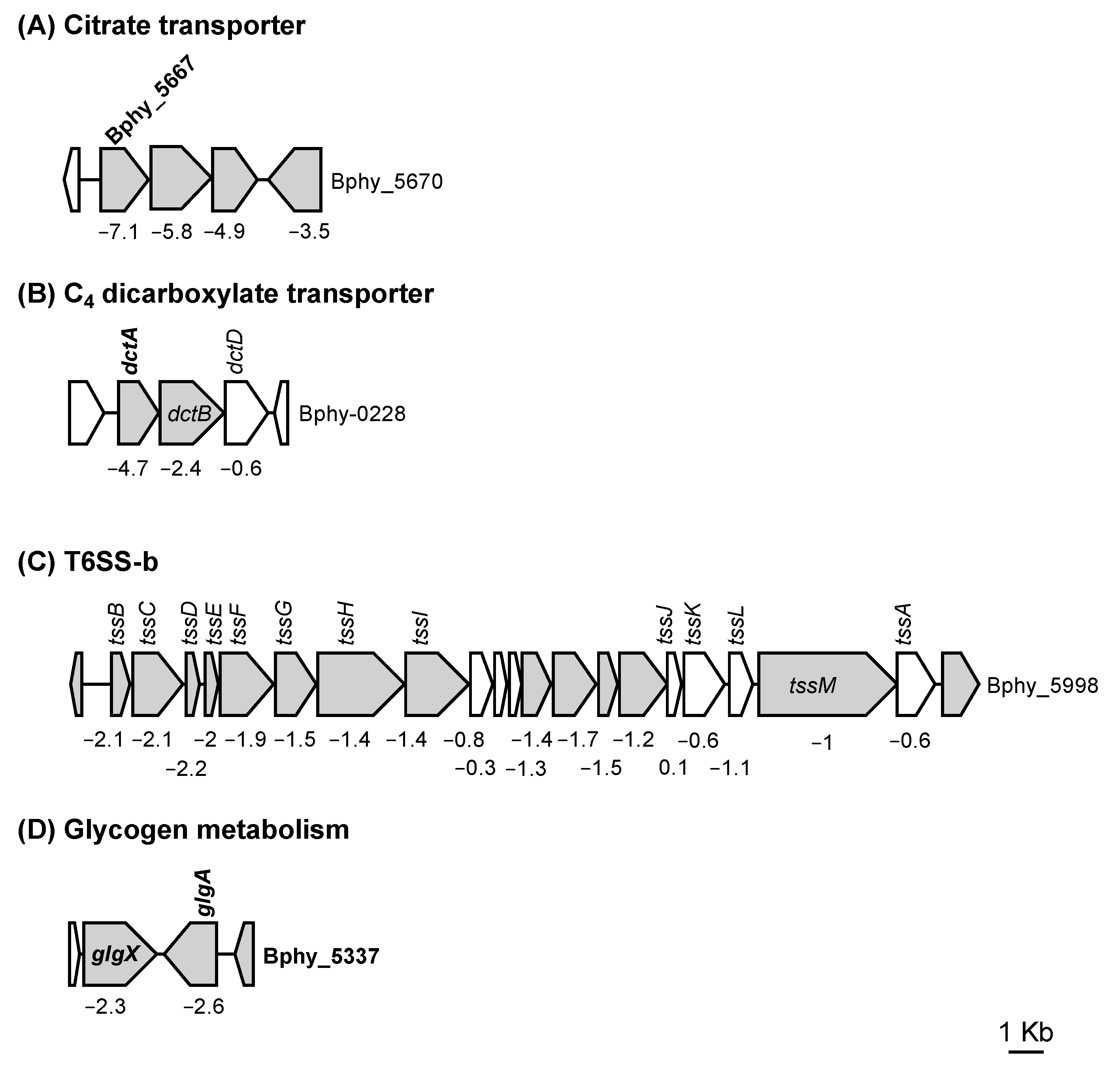
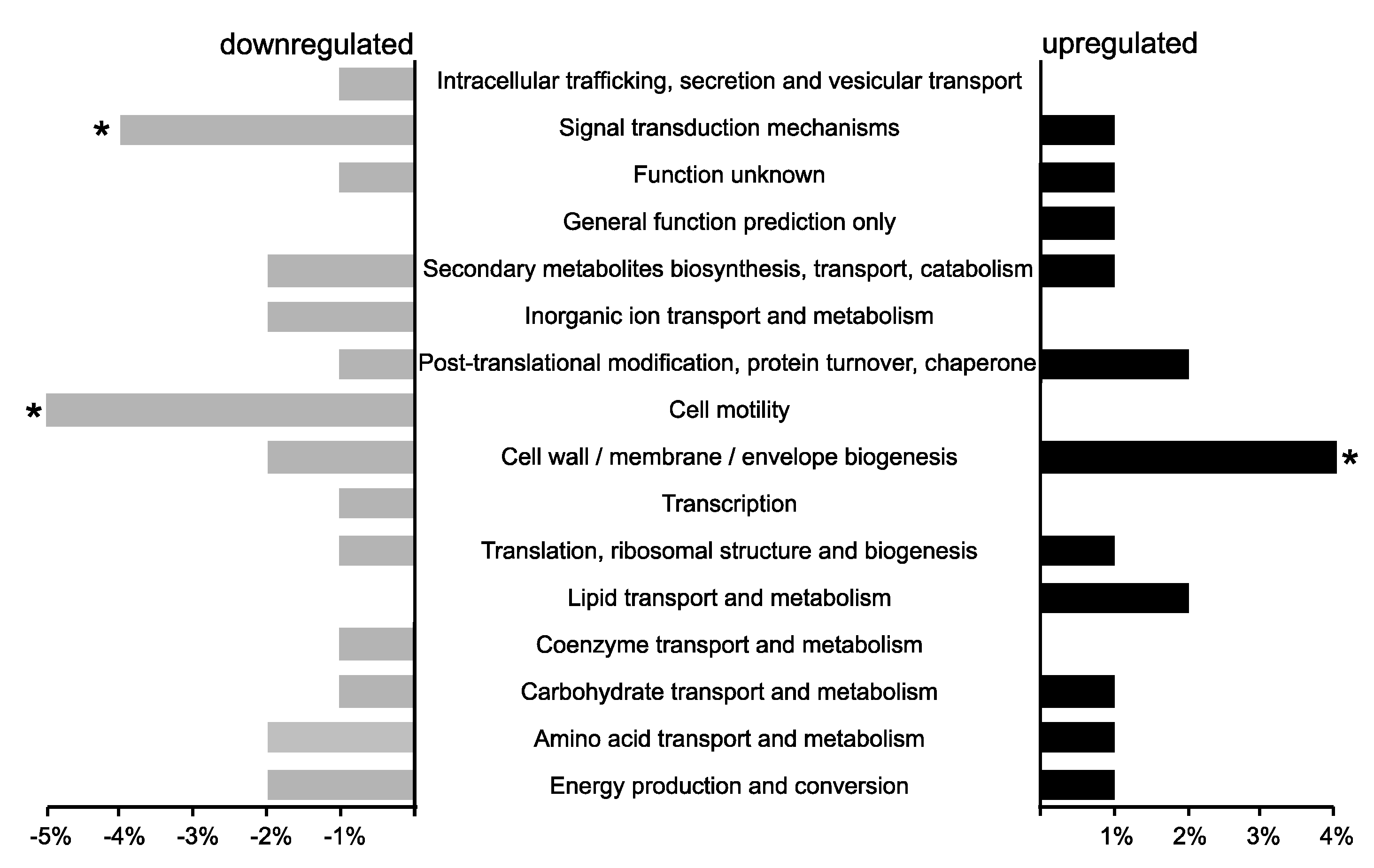
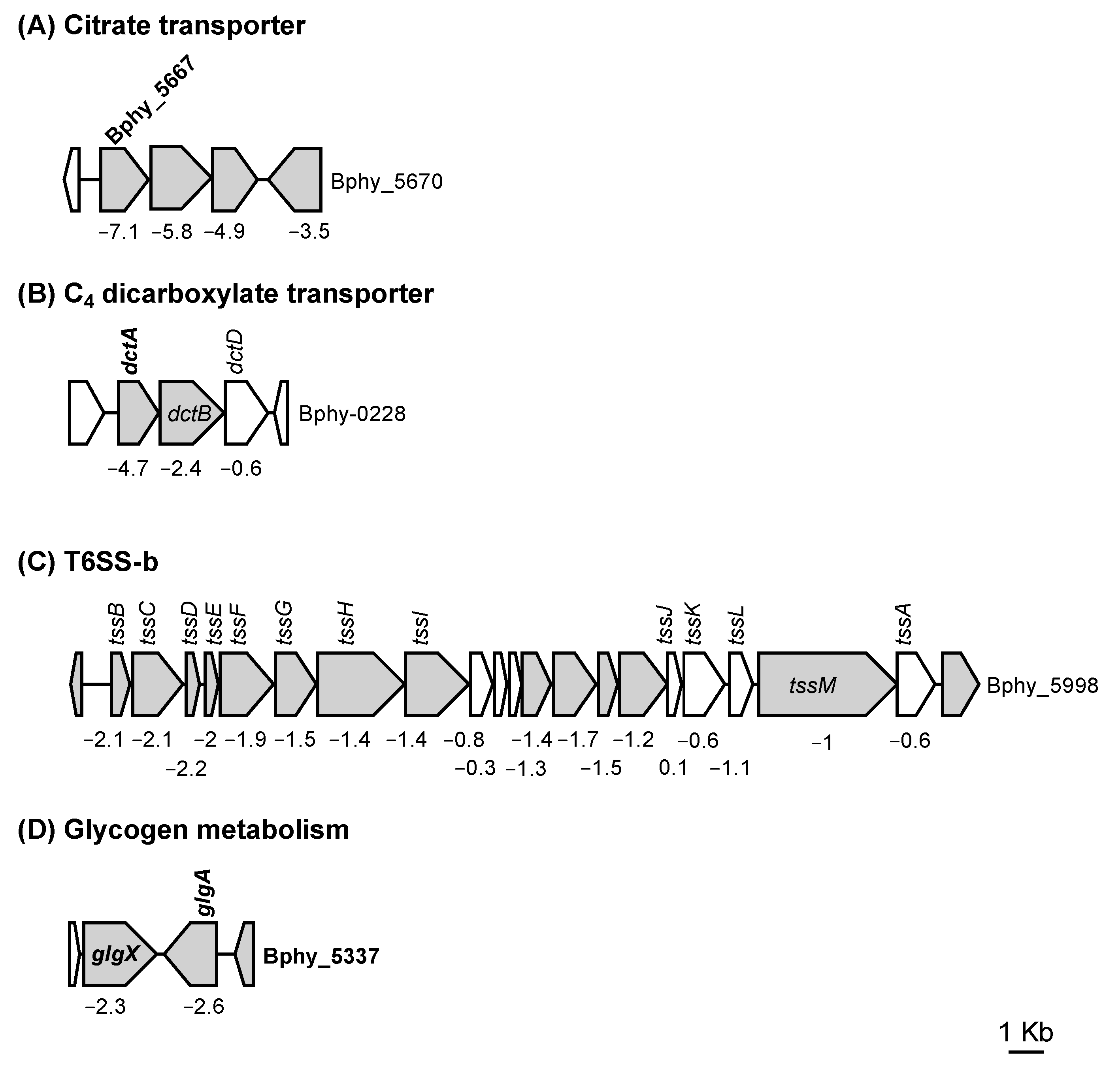
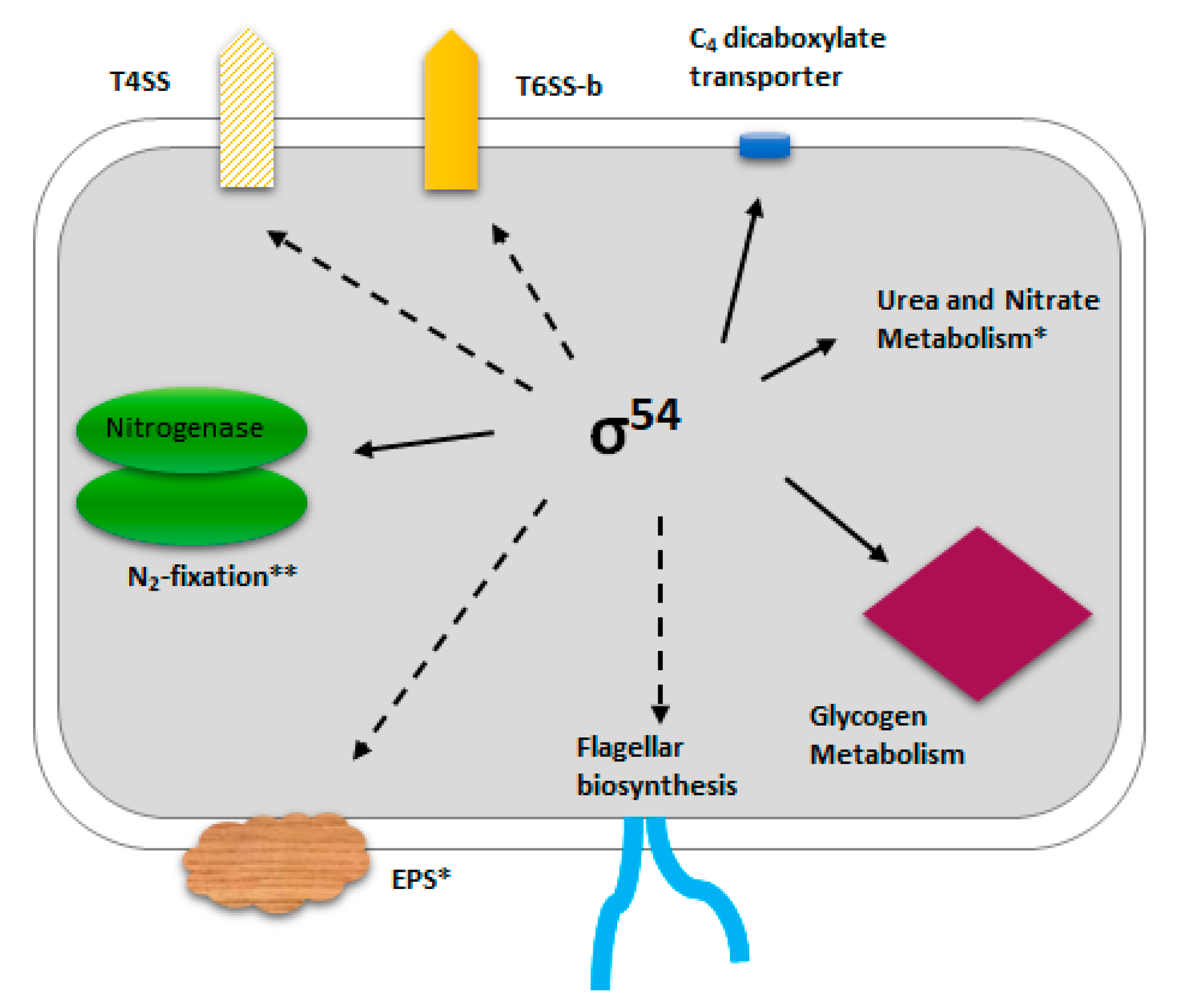
3.1. The P. phymatum STM815 σ54 Regulon in Free-Living Nitrogen-Limiting Conditions
3.2. Phenotypic Characterization of the σ54 Mutant
3.2.1. Role of σ54 for Assimilation of C4 Dicarboxylates
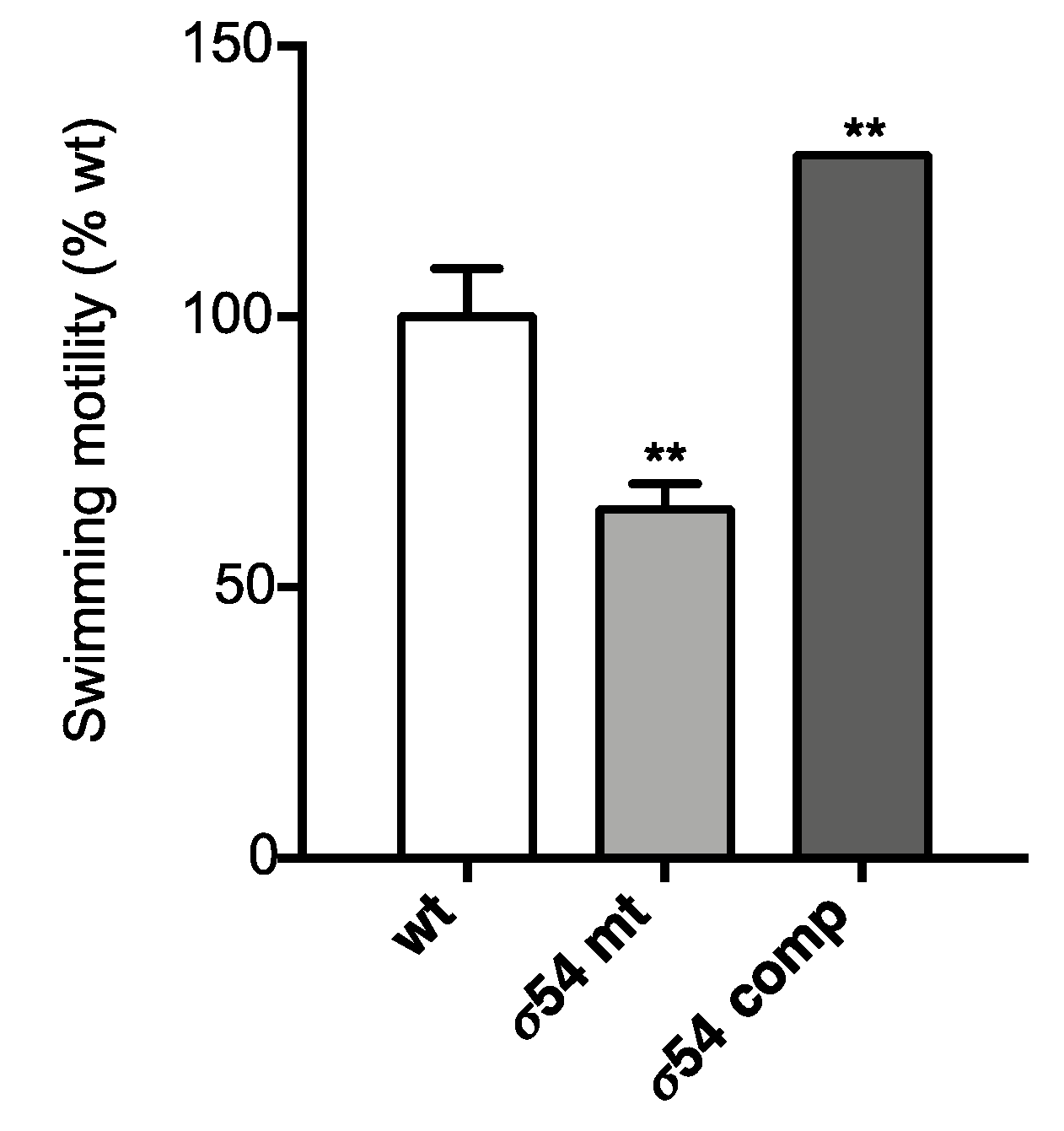
3.2.2. σ54 Positively Controls Swimming Motility
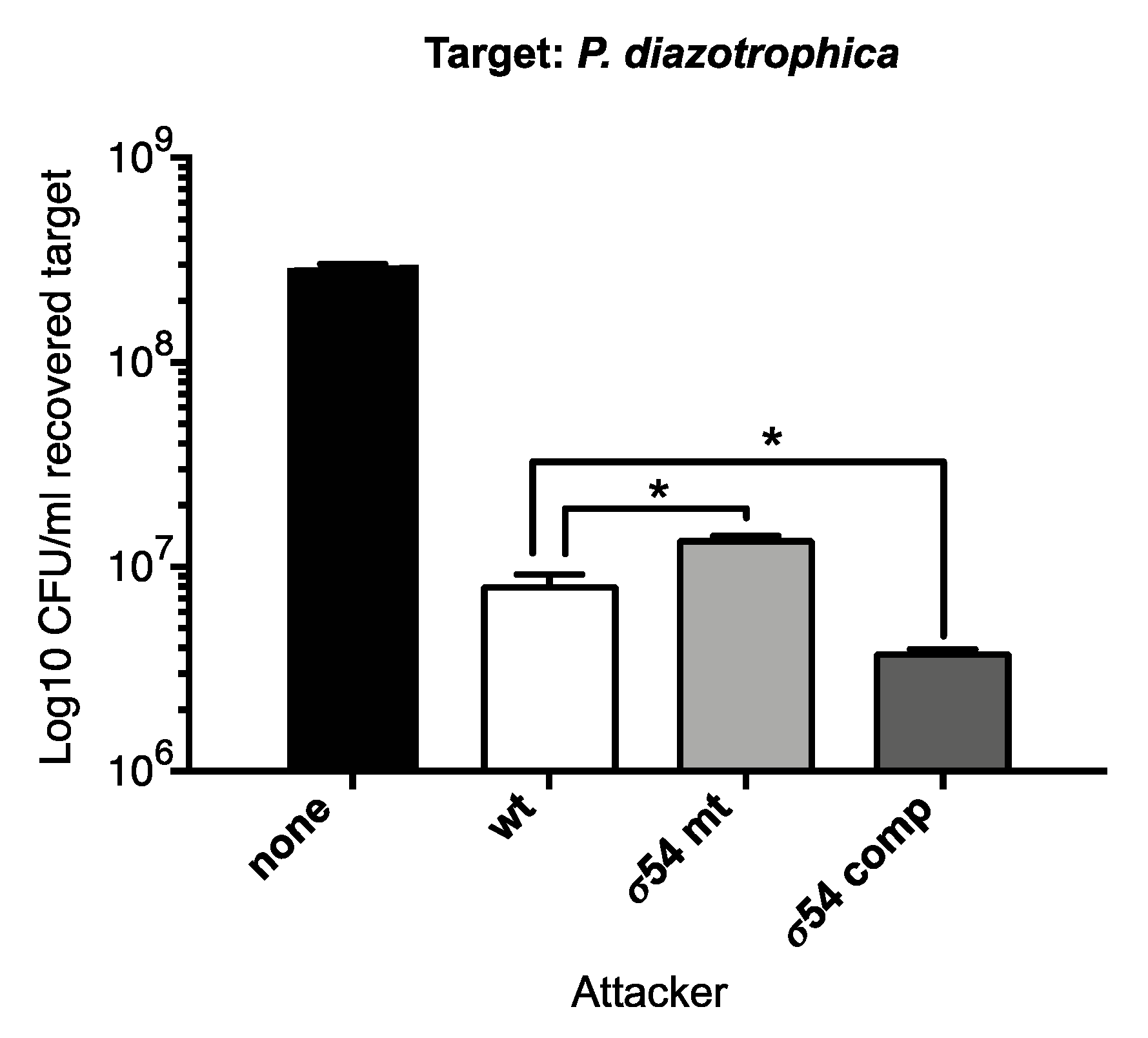
3.2.3. The Presence of σ54 Influences Interbacterial Competition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; Hättenschwiler, S.; Olander, L.P.; Allison, S. Nitrogen and Nature. Ambio 2002, 31, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Terpolilli, J.J.; Hood, G.A.; Poole, P.S. What determines the efficiency of N2-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol. 2012, 60, 325–389. [Google Scholar] [CrossRef]
- Yurgel, S.N.; Kahn, M.L. Dicarboxylate transport by rhizobia. FEMS Microbiol. Rev. 2004, 28, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C.; Munive, J.-A. Nodulation of legumes by members of the β-subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef]
- Chen, W.M.; Laevens, S.; Lee, T.M.; Coenye, T.; De Vos, P.; Mergeay, M.; Vandamme, P. Ralstonia taiwanensis sp. nov., isolated from root nodules of Mimosa species and sputum of a cystic fibrosis patient. Int. J. Syst. Evol. Microbiol. 2001, 51, 1729–1735. [Google Scholar] [CrossRef] [Green Version]
- Sawana, A.; Adeolu, M.; Gupta, R.S. Molecular signatures and phylogenomic analysis of the genus Burkholderia: Proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front. Genet. 2014, 5, 429. [Google Scholar] [CrossRef] [Green Version]
- Beukes, C.W.; Palmer, M.; Manyaka, P.; Chan, W.Y.; Avontuur, J.R.; Van Zyl, E.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; et al. Genome data provides high support for generic boundaries in Burkholderia sensu lato. Front. Microbiol. 2017, 8, 1154. [Google Scholar] [CrossRef]
- Chen, W.-M.; De Faria, S.M.; Straliotto, R.; Pitard, R.M.; Simões-Araújo, J.L.; Chou, J.-H.; Chou, Y.-J.; Barrios, E.; Prescott, A.R.; Elliott, G.N.; et al. Proof that Burkholderia strains form effective symbioses with legumes: A study of novel Mimosa-nodulating strains from South America. Appl. Environ. Microbiol. 2005, 71, 7461–7471. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wei, S.; Wang, F.; James, E.; Guo, X.; Zagar, C.; Xia, L.G.; Dong, X.; Wang, Y.P. Burkholderia and Cupriavidus spp. are the preferred symbionts of Mimosa spp. in Southern China. FEMS Microbiol. Ecol. 2012, 80, 417–426. [Google Scholar] [CrossRef]
- Mishra, R.P.; Tisseyre, P.; Melkonian, R.; Miché, L.; Klonowska, A.; Gonzalez, S.; Laguerre, G.; Chaintreuil, C.; Béna, G.; Moulin, L. Genetic diversity of Mimosa pudica rhizobial symbionts in soils of French Guiana: Investigating the origin and diversity of Burkholderia phymatum and other beta-rhizobia. FEMS Microbiol. Ecol. 2012, 79, 487–503. [Google Scholar] [CrossRef] [PubMed]
- Gehlot, H.S.; Tak, N.; Kaushik, M.; Mitra, S.; Chen, W.-M.; Poweleit, N.; Panwar, D.; Poonar, N.; Parihar, R.; Tak, A.; et al. An invasive Mimosa in India does not adopt the symbionts of its native relatives. Ann. Bot. 2013, 112, 179–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, B.; Dlodlo, O.; Chimphango, S.; Stirton, C.; Schrire, B.; Boatwright, J.S.; Honnay, O.; Smets, E.; Sprent, J.; James, E.K.; et al. Symbiotic diversity, specificity and distribution of rhizobia in native legumes of the Core Cape Subregion (South Africa). FEMS Microbiol. Ecol. 2015, 91, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, B.; Chimphango, S.B.M.; Stirton, C.; Rafudeen, M.S.; Honnay, O.; Smets, E.; Chen, W.-M.; Sprent, J.; James, E.; Muasya, A.M. Biogeographical patterns of legume-Nodulating Burkholderia spp.: From African fynbos to continental scales. Appl. Environ. Microbiol. 2016, 82, 5099–5115. [Google Scholar] [CrossRef] [Green Version]
- Gyaneshwar, P.; Hirsch, A.M.; Moulin, L.; Chen, W.-M.; Elliott, G.N.; Bontemps, C.; Estrada-de los Santos, P.; Gross, E.; Dos Reis, F.B.; Sprent, J.I.; et al. Legume-nodulating Betaproteobacteria: Diversity, host range, and future prospects. Mol. Plant Microbe Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Howieson, J.; De Meyer, S.E.; Vivas-Marfisi, A.; Ratnayake, S.; Ardley, J.K.; Yates, R. Novel Burkholderia bacteria isolated from Lebeckia ambigua—A perennial suffrutescent legume of the fynbos. Soil Biol. Biochem. 2013, 60, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Melkonian, R.; Moulin, L.; Béna, G.; Tisseyre, P.; Chaintreuil, C.; Heulin, K.; Rezkallah, N.; Klonowska, A.; Gonzalez, S.; Simon, M.F.; et al. The geographical patterns of symbiont diversity in the invasive legume Mimosa pudica can be explained by the competitiveness of its symbionts and by the host genotype: Competition for nodulation in α- and β-rhizobia. Environ. Microbiol. 2014, 16, 2099–2111. [Google Scholar] [CrossRef]
- Bontemps, C.; Rogel, M.A.; Wiechmann, A.; Mussabekova, A.; Moody, S.; Simon, M.F.; Moulin, L.; Elliott, G.N.; Lacercat-Didier, L.; DaSilva, C.; et al. Endemic Mimosa species from Mexico prefer alphaproteobacterial rhizobial symbionts. New Phytol. 2016, 209, 319–333. [Google Scholar] [CrossRef] [Green Version]
- Elliott, G.N.; Chou, J.-H.; Chen, W.-M.; Bloemberg, G.V.; Bontemps, C.; Martínez-Romero, E.; Velázquez, E.; Young, J.P.W.; Sprent, J.I.; James, E. Burkholderia spp. are the most competitive symbionts of Mimosa, particularly under N-limited conditions. Environ. Microbiol. 2009, 11, 762–778. [Google Scholar] [CrossRef]
- Lardi, M.; De Campos, S.B.; Purtschert, G.; Eberl, L.; Pessi, G. Competition experiments for legume infection identify Burkholderia phymatum as a highly competitive β-Rhizobium. Front. Microbiol. 2017, 8, 1527. [Google Scholar] [CrossRef] [Green Version]
- Elliott, G.N.; Chen, W.-M.; Chou, J.-H.; Wang, H.-C.; Sheu, S.-Y.; Perin, L.; Reis, V.M.; Moulin, L.; Simon, M.F.; Bontemps, C.; et al. Burkholderia phymatum is a highly effective nitrogen-fixing symbiont of Mimosa spp. and fixes nitrogen ex planta. New Phytol. 2007, 173, 168–180. [Google Scholar] [CrossRef] [PubMed]
- De Campos, S.B.; Lardi, M.; Gandolfi, A.; Eberl, L.; Pessi, G. Mutations in two Paraburkholderia phymatum Type VI secretion systems cause reduced fitness in interbacterial competition. Front. Microbiol. 2017, 8, 2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morett, E.; Buck, M. In vivo studies on the interaction of RNA polymerase-σ54 with the Klebsiella pneumoniae and Rhizobium meliloti nifH promoters: The role of NifA in the formation of an open promoter complex. J. Mol. Biol. 1989, 210, 65–77. [Google Scholar] [CrossRef]
- Francke, C.; Kormelink, T.G.; Hagemeijer, Y.; Overmars, L.; Sluijter, V.; Moezelaar, R.; Siezen, R.J. Comparative analyses imply that the enigmatic sigma factor 54 is a central controller of the bacterial exterior. BMC Genom. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lardi, M.; Liu, Y.; Purtschert, G.; De Campos, S.B.; Pessi, G. Transcriptome analysis of Paraburkholderia phymatum under nitrogen starvation and during symbiosis with Phaseolus vulgaris. Genes 2017, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Lardi, M.; Liu, Y.; Giudice, G.; Ahrens, C.H.; Zamboni, N.; Pessi, G. Metabolomics and transcriptomics identify multiple downstream targets of Paraburkholderia phymatum σ54 during symbiosis with Phaseolus vulgaris. Int. J. Mol. Sci. 2018, 19, 1049. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.-Y.; Chou, J.-H.; Bontemps, C.; Elliott, G.N.; Gross, E.; dos Reis Junior, F.B.; Melkonian, R.; Moulin, L.; James, E.K.; Sprent, J.I.; et al. Burkholderia diazotrophica sp. nov., isolated from root nodules of Mimosa spp. Int. J. Syst. Evol. Microbiol. 2013, 63, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.J.; Maaløe, O. DNA replication and the division cycle in Escherichia coli. J. Mol. Biol. 1967, 23, 99–112. [Google Scholar] [CrossRef]
- Miller, W.G.; Leveau, J.H.; Lindow, S.E. Improved gfp and inaZ broad-host-range promoter-probe vectors. Mol. Plant Microbe Interact. 2000, 13, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.-L.Y.; Beatson, S.A.; McEwan, A.G.; Schembri, M.A. Conjugative plasmid transfer and adhesion dynamics in an Escherichia coli biofilm. Appl. Environ. Microbiol. 2009, 75, 6783–6791. [Google Scholar] [CrossRef] [Green Version]
- Pessi, G.; Ahrens, C.H.; Rehrauer, H.; Lindemann, A.; Hauser, F.; Fischer, H.-M.; Hennecke, H. Genome-wide transcript analysis of Bradyrhizobium japonicum bacteroids in soybean root nodules. Mol. Plant Microbe Interact. 2007, 20, 1353–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulin, L.; Klonowska, A.; Caroline, B.; Booth, K.; Vriezen, J.A.; Melkonian, R.; James, E.; Young, J.P.W.; Béna, G.; Hauser, L.; et al. Complete Genome sequence of Burkholderia phymatum STM815T, a broad host range and efficient nitrogen-fixing symbiont of Mimosa species. Stand. Genom. Sci. 2014, 9, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, S.; Szklarczyk, D.; Trachana, K.; Roth, A.; Kuhn, M.; Muller, J.; Arnold, R.; Rattei, T.; Letunic, I.; Doerks, T.; et al. eggNOG v3.0: Orthologous groups covering 1133 organisms at 41 different taxonomic ranges. Nucleic Acids Res. 2012, 40, D284–D289. [Google Scholar] [CrossRef]
- Ledebur, H.C.; Gu, B.; Sojda, J.; Nixon, B.T. Rhizobium meliloti and Rhizobium leguminosarum dctD gene products bind to tandem sites in an activation sequence located upstream of sigma 54-dependent dctA promoters. J. Bacteriol. 1990, 172, 3888–3897. [Google Scholar] [CrossRef] [Green Version]
- Valentini, M.; Storelli, N.; Lapouge, K. Identification of C4-dicarboxylate transport systems in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2011, 193, 4307–4316. [Google Scholar] [CrossRef] [Green Version]
- Poole, P.; Allaway, D. Carbon and nitrogen metabolism in Rhizobium. Adv. Microb. Physiol. 2000, 43, 117–163. [Google Scholar] [CrossRef]
- Filloux, A. The rise of the Type VI secretion system. F1000Prime Rep. 2013, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.G.; Hu, B.; Christie, P.J. Biological and structural diversity of type IV secretion systems. Microbiol. Spectr. 2019, 7, 1–15. [Google Scholar] [CrossRef]
- Liu, Y.; Bellich, B.; Hug, S.; Eberl, L.; Cescutti, P.; Pessi, G. The exopolysaccharide cepacian plays a role in the establishment of the Paraburkholderia phymatum—Phaseolus vulgaris symbiosis. Front. Microbiol. 2020, 11, 1600. [Google Scholar] [CrossRef]
- Garcia, E.; Bancroft, S.; Rhee, S.G.; Kustu, S. The product of a newly identified gene, gInF, is required for synthesis of glutamine synthetase in Salmonella. Proc. Natl. Acad. Sci. USA 1977, 74, 1662–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Wu, G.; Liao, Y.; Zeng, Q.; Wang, H.; Liu, F. RpoN1 and RpoN2 play different regulatory roles in virulence traits, flagellar biosynthesis, and basal metabolism in Xanthomonas campestris. Mol. Plant Pathol. 2020, 21, 907–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldías, M.S.; Lamothe, J.; Wu, R.; Valvano, M.A. Burkholderia cenocepacia requires the RpoN sigma factor for biofilm formation and intracellular trafficking within macrophages. Infect. Immun. 2008, 76, 1059–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kullik, I.; Fritsche, S.; Knobel, H.; Sanjuan, J.; Hennecke, H.; Fischer, H.M. Bradyrhizobium japonicum has two differentially regulated, functional homologs of the σ54 gene (rpoN). J. Bacteriol. 1991, 173, 1125–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, B.; Mo, Z.-L.; Xiao, P.; Pan, H.-J.; Lan, X.; Li, G.-Y. Role of alternative sigma factor 54 (RpoN) from Vibrio anguillarum M3 in protease secretion, exopolysaccharide production, biofilm formation, and virulence. Appl. Microbiol. Biotechnol. 2012, 97, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Hayrapetyan, H.; Tempelaars, M.; Groot, M.N.; Abee, T. Bacillus cereus ATCC 14579 RpoN (sigma 54) is a pleiotropic regulator of growth, carbohydrate metabolism, motility, biofilm formation and toxin production. PLoS ONE 2015, 10, e0134872. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Liu, Y.; Chen, Y.; Yam, J.K.H.; Chew, S.C.; Chua, S.L.; Wang, K.; Givskov, M.; Yang, L. RpoN regulates virulence factors of Pseudomonas aeruginosa via modulating the PqsR quorum sensing regulator. Int. J. Mol. Sci. 2015, 16, 28311–28319. [Google Scholar] [CrossRef] [Green Version]
- Lardi, M.; Aguilar, C.; Pedrioli, A.; Omasits, U.; Suppiger, A.; Cárcamo-Oyarce, G.; Schmid, N.; Ahrens, C.H.; Eberl, L.; Pessi, G. σ54-dependent response to nitrogen limitation and virulence in Burkholderia cenocepacia strain H111. Appl. Environ. Microbiol. 2015, 81, 4077–4089. [Google Scholar] [CrossRef] [Green Version]
- Salazar, E.; Díaz-Mejía, J.J.; Moreno-Hagelsieb, G.; Martínez-Batallar, G.; Mora, Y.; Mora, J.; Encarnación, S. Characterization of the NifA-RpoN regulon in Rhizobium etli in free life and in symbiosis with Phaseolus vulgaris. Appl. Environ. Microbiol. 2010, 76, 4510–4520. [Google Scholar] [CrossRef] [Green Version]
- Hauser, F.; Pessi, G.; Friberg, M.; Weber, C.; Rusca, N.; Lindemann, A.; Fischer, H.-M.; Hennecke, H. Dissection of the Bradyrhizobium japonicum NifA+σ54 regulon, and identification of a ferredoxin gene (fdxN) for symbiotic nitrogen fixation. Mol. Genet. Genomics 2007, 278, 255–271. [Google Scholar] [CrossRef]
- Fazli, M.; Rybtke, M.; Steiner, E.; Weidel, E.; Berthelsen, J.; Groizeleau, J.; Bin, W.; Zhi, B.Z.; Yaming, Z.; Kaever, V.; et al. Regulation of Burkholderia cenocepacia biofilm formation by RpoN and the c-di-GMP effector BerB. Microbiologyopen 2017, 6, e00480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, S.J.; Golby, P.; Omrani, D.; Broad, S.A.; Harrington, V.L.; Guest, J.R.; Kelly, D.J.; Andrews, S.C. Inactivation and regulation of the aerobic C4-dicarboxylate transport (dctA) gene of Escherichia coli. J. Bacteriol. 1999, 181, 5624–5635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, C.J.; Poole, P.S. Roles of DctA and DctB in signal detection by the dicarboxylic acid transport system of Rhizobium leguminosarum. J. Bacteriol. 1998, 180, 2660–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finan, T.M.; Wood, J.M.; Jordan, D.C. Symbiotic properties of C4-dicarboxylic acid transport mutants of Rhizobium leguminosarum. J. Bacteriol. 1983, 154, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronson, C.W.; Astwood, P.M.; Downie, J.A. Molecular cloning and genetic organization of C4-dicarboxylate transport genes from Rhizobium leguminosarum. J. Bacteriol. 1984, 160, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelke, T.; Jagadish, M.N.; Pühler, A. Biochemical and genetical analysis of Rhizobium meliloti mutants defective in C4-dicarboxylate transport. Microbiology 1987, 133, 3019–3029. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Boller, T.; Wiemken, A. Trehalose becomes the most abundant non-structural carbohydrate during senescence of soybean nodules. J. Exp. Bot. 2001, 52, 943–947. [Google Scholar] [CrossRef]
- Streeter, J.G. Effect of trehalose on survival of Bradyrhizobium japonicum during desiccation. J. Appl. Microbiol. 2003, 95, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Streeter, J.G.; López-Gómez, M.L. Three enzymes for trehalose synthesis in Bradyrhizobium cultured bacteria and in bacteroids from soybean nodules. Appl. Environ. Microbiol. 2006, 72, 4250–4255. [Google Scholar] [CrossRef] [Green Version]
- Cytryn, E.J.; Sangurdekar, D.P.; Streeter, J.G.; Franck, W.L.; Chang, W.-S.; Stacey, G.; Emerich, D.W.; Joshi, T.; Xu, D.; Sadowsky, M.J. Transcriptional and physiological responses of Bradyrhizobium japonicum to desiccation-induced stress. J. Bacteriol. 2007, 189, 6751–6762. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, M.; Cytryn, E.J.; Sadowsky, M.J. Functional role of Bradyrhizobium japonicum trehalose biosynthesis and metabolism genes during physiological stress and nodulation. Appl. Environ. Microbiol. 2010, 76, 1071–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lardi, M.; Murset, V.; Fischer, H.-M.; Mesa, S.; Ahrens, C.H.; Zamboni, N.; Pessi, G. Metabolomic profiling of Bradyrhizobium diazoefficiens-induced root nodules reveals both host plant-specific and developmental signatures. Int. J. Mol. Sci. 2016, 17, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, R.; Wong, A.; Ramirez, M.; Barraza, A.; Orozco, M.D.C.; Cevallos, M.A.; Lara, M.; Hernández, G.; Iturriaga, G. Improvement of drought tolerance and grain yield in common bean by overexpressing trehalose-6-phosphate synthase in Rhizobia. Mol. Plant Microbe Interact. 2008, 21, 958–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, J.E.; Delorge, I.; Figueroa, C.M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J. 2014, 79, 544–567. [Google Scholar] [CrossRef]
- Delorge, I.; Figueroa, C.M.; Feil, R.; Lunn, J.E.; Van Dijck, P. Trehalose-6-phosphate synthase 1 is not the only active TPS in Arabidopsis thaliana. Biochem. J. 2015, 466, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Gonzalez-Uriarte, A.; Griffiths, C.A.; Hassani-Pak, K. The Role of trehalose 6-phosphate in crop yield and resilience. Plant Physiol. 2018, 177, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Hengge-Aronis, R.; Klein, W.; Lange, R.; Rimmele, M.; Boos, W. Trehalose synthesis genes are controlled by the putative sigma factor encoded by rpoS and are involved in stationary-phase thermotolerance in Escherichia coli. J. Bacteriol. 1991, 173, 7918–7924. [Google Scholar] [CrossRef] [Green Version]
- Flechard, M.; Fontenelle, C.; Blanco, C.; Goude, R.; Ermel, G.; Trautwetter, A. RpoE2 of Sinorhizobium meliloti is necessary for trehalose synthesis and growth in hyperosmotic media. Microbiology 2010, 156, 1708–1718. [Google Scholar] [CrossRef] [Green Version]
- Bernard, C.S.; Brunet, Y.R.; Gavioli, M.; Lloubès, R.; Cascales, E. Regulation of type VI secretion gene clusters by σ54 and cognate enhancer binding proteins. J. Bacteriol. 2011, 193, 2158–2167. [Google Scholar] [CrossRef] [Green Version]
- Sana, T.G.; Soscia, C.; Tonglet, C.M.; Garvis, S.; Bleves, S. Divergent control of two type VI secretion Systems by RpoN in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e76030. [Google Scholar] [CrossRef]
- Sheng, L.; Gu, D.; Wang, Q.; Liu, Q.; Zhang, Y. Quorum sensing and alternative sigma factor RpoN regulate type VI secretion system I (T6SSVA1) in fish pathogen Vibrio alginolyticus. Arch. Microbiol. 2012, 194, 379–390. [Google Scholar] [CrossRef]
- Dong, T.G.; Mekalanos, J.J. Characterization of the RpoN regulon reveals differential regulation of T6SS and new flagellar operons in Vibrio cholerae O37 strain V52. Nucleic Acids Res. 2012, 40, 7766–7775. [Google Scholar] [CrossRef] [Green Version]


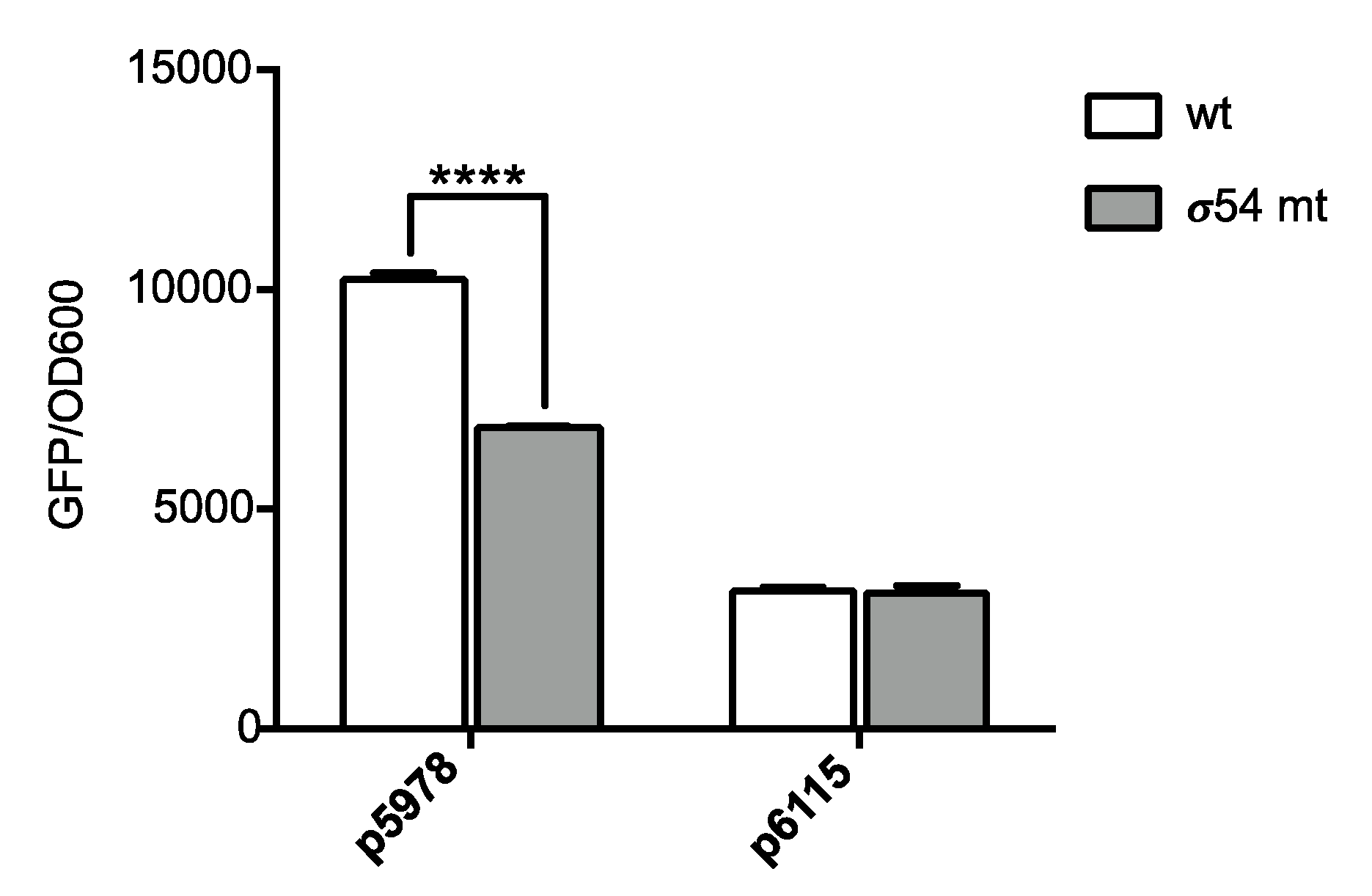



{kind=link}
{kind=link}
{kind=link}
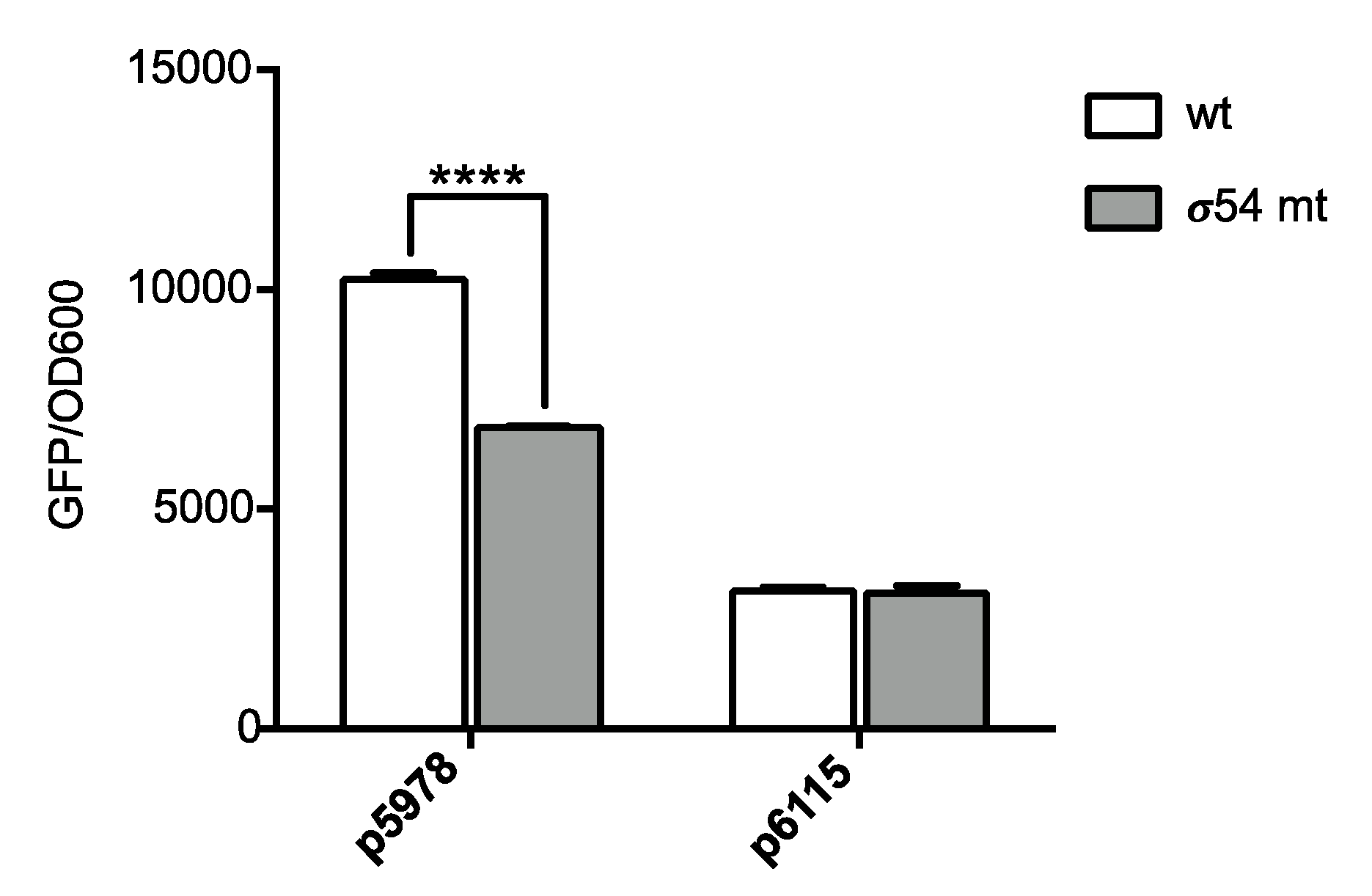{kind=link}
{kind=link}
{kind=link}
| Locus ID 1 | Description 1 | Gene Name | Log2FC (σ54 mt vs. wt) 2 |
|---|---|---|---|
| Amino Acid Transport and Metabolism | |||
| Bphy_0112 | extracellular ligand-binding receptor | −1.1 | |
| Bphy_0588 | extracellular ligand-binding receptor | −1.2 | |
| Bphy_0589 | inner-membrane translocator | −1.8 | |
| Bphy_0590 | inner-membrane translocator | −1.5 | |
| Bphy_0591 | ABC transporter-like protein | −1.4 | |
| Bphy_2572 | polar amino acid ABC transporter inner membrane subunit | −1.2 | |
| Bphy_2573 | extracellular solute-binding protein | −1.3 | |
| Bphy_3017 | extracellular ligand-binding receptor | −1.2 | |
| Bphy_3043 | extracellular ligand-binding receptor | −1.4 | |
| Bphy_3046 | inner-membrane translocator | −1.6 | |
| Bphy_3047 | extracellular ligand-binding receptor | −1.2 | |
| Bphy_3780 | cationic amino acid ABC transporter | −1.4 | |
| Bphy_5470 | 5-methyltetrahydropteroyltriglutamate-homocysteine methyltransferase | −1.9 | |
| Bphy_5613 | methionine gamma-lyase | −2.1 | |
| Carbohydrate Transport and Metabolism | |||
| Bphy_1237 | periplasmic binding protein/LacI transcriptional regulator | −1.2 | |
| Bphy_2145 | trehalose-6-phosphate synthase | otsA | −2.4 |
| Bphy_2615 | monosaccharide-transporting ATPase | −1.1 | |
| Bphy_5335 | glycogen debranching enzyme | glgX | −2.3 |
| Bphy_5336 | glycogen synthase | glgA | −2.6 |
| Bphy_5372 | extracellular solute-binding protein | −1.3 | |
| Cell Motility | |||
| Bphy_2938 | flagellar motor switch protein FliM | fliM | −3.2 |
| Bphy_2939 | flagellar motor switch protein FliN | fliN | −3.6 |
| Bphy_2956 | flagellar rod assembly protein/muramidase FlgJ | flgJ | −2.9 |
| Bphy_2962 | flagellar biosynthesis regulator FlhF | flhF | −4.1 |
| Bphy_5592 | methyl-accepting chemotaxis sensory transducer | −2.0 | |
| Cell Wall/Membrane/Envelope Biogenesis | |||
| Bphy_0254 | Porin | −1.1 | |
| Bphy_1082 | Porin | −1.6 | |
| Bphy_2167 | OmpC family outer membrane porin | −1.2 | |
| Bphy_2684 | Porin | −2.2 | |
| Bphy_3730 | glycosyl transferase family protein | −1.7 | |
| Bphy_3733 | exopolysaccharide tyrosine-protein kinase | −2.0 | |
| Bphy_5991 | aspartate racemase | −1.5 | |
| Coenzyme Transport and Metabolism | |||
| Bphy_3573 | putative aminotransferase | −1.4 | |
| Bphy_7651 | aminotransferase class-III | −1.3 | |
| Energy Production and Conversion | |||
| Bphy_0010 | aldehyde dehydrogenase | −2.3 | |
| Bphy_0225 | C4-dicarboxylate transporter | dctA | −4.7 |
| Bphy_1368 | isocitrate lyase | −1.4 | |
| Bphy_3971 | cytochrome o ubiquinol oxidase subunit IV | −1.4 | |
| Bphy_4774 | oxidoreductase alpha (molybdopterin) subunit | −1.1 | |
| Bphy_5667 | citrate carrier protein | −7.1 | |
| Bphy_5990 | sodium:dicarboxylate symporter | −1.7 | |
| Bphy_5992 | sodium:dicarboxylate symporter | −1.2 | |
| Function Unknown | |||
| Bphy_0467 | hypothetical protein | −1.1 | |
| Bphy_0675 | hypothetical protein | −1.4 | |
| Bphy_0848 | CsbD family protein | −1.3 | |
| Bphy_0980 | Ku protein | −1.4 | |
| Bphy_1016 | hypothetical protein | −1.3 | |
| Bphy_1217 | hypothetical protein | −1.0 | |
| Bphy_2791 | hypothetical protein | −1.2 | |
| Bphy_2880 | hypothetical protein | −1.8 | |
| Bphy_3671 | hypothetical protein | −1.7 | |
| Bphy_3731 | hypothetical protein | −1.7 | |
| Bphy_3807 | hypothetical protein | −2.5 | |
| Bphy_3844 | hypothetical protein | −3.6 | |
| Bphy_3917 | hypothetical protein | −1.4 | |
| Bphy_4035 | hypothetical protein | −1.5 | |
| Bphy_4046 | MbtH domain-containing protein | −2.3 | |
| Bphy_4787 | hypothetical protein | −3.1 | |
| Bphy_5272 | hypothetical protein | −1.6 | |
| Bphy_5337 | putative lipoprotein | −3.7 | |
| Bphy_5431 | hypothetical protein | −4.9 | |
| Bphy_5573 | hypothetical protein | −1.5 | |
| Bphy_5699 | hypothetical protein | −2.9 | |
| Bphy_5724 | hypothetical protein | −2.5 | |
| Bphy_5976 | hypothetical protein | −1.3 | |
| Bphy_5977 | hypothetical protein | −2.3 | |
| Bphy_5978 | type VI secretion protein | −2.1 | |
| Bphy_5979 | EvpB family type VI secretion protein | −2.1 | |
| Bphy_5980 | hypothetical protein | −2.2 | |
| Bphy_5981 | type VI secretion system lysozyme-related protein | −2.0 | |
| Bphy_5982 | type VI secretion protein | −1.9 | |
| Bphy_5983 | type VI secretion protein | −1.5 | |
| Bphy_5985 | type VI secretion system Vgr family protein | −1.4 | |
| Bphy_5996 | IcmF-like protein | −1.0 | |
| Bphy_5998 | hypothetical protein | −1.1 | |
| Bphy_7681 | hypothetical protein | −1.3 | |
| Bphy_7682 | hypothetical protein | −1.5 | |
| General Function Prediction Only | |||
| Bphy_5298 | PfpI family intracellular peptidase | −2.7 | |
| Bphy_5303 | transport-associated | −5.2 | |
| Bphy_5721 | amidohydrolase 3 | −2.1 | |
| Inorganic Ion Transport and Metabolism | |||
| Bphy_3175 | NMT1/THI5-like domain-containing protein | −1.8 | |
| Bphy_3959 | nitrate/sulfonate/bicarbonate ABC transporter periplasmic ligand-binding protein | −7.2 | |
| Bphy_3960 | ABC transporter related | −7.4 | |
| Bphy_3961 | binding-protein-dependent transport systems inner membrane component | −6.0 | |
| Bphy_4041 | periplasmic binding protein | −1.8 | |
| Intracellular Trafficking, Secretion and Vesicular Transport | |||
| Bphy_7526 | P-type DNA transfer ATPase VirB11 | −1.3 | |
| Bphy_7530 | VirB8 family protein | −1.5 | |
| Lipid transport and metabolism | |||
| Bphy_0466 | GDSL family lipase | −1.9 | |
| Post-translational Modification, Protein Turnover and Chaperones | |||
| Bphy_2992 | TPR repeat-containing protein | −1.3 | |
| Bphy_5984 | type VI secretion ATPase | −1.4 | |
| Secondary Metabolites Biosynthesis, Transport and Catabolism | |||
| Bphy_0174 | hypothetical protein | −1.1 | |
| Bphy_4036 | lysine/ornithine N-monooxygenase | −1.9 | |
| Bphy_4038 | amino acid adenylation domain-containing protein | −2.2 | |
| Bphy_5720 | isochorismatase hydrolase | −2.4 | |
| Signal Transduction Mechanisms | |||
| Bphy_0226 | integral membrane sensor signal transduction histidine kinase | −2.4 | |
| Bphy_3957 | histidine kinase | −1.8 | |
| Bphy_3958 | two component transcriptional regulator | −2.3 | |
| Bphy_3963 | heavy metal sensor signal transduction histidine kinase | −1.2 | |
| Bphy_4749 | PAS/PAC sensor hybrid histidine kinase | −1.2 | |
| Bphy_5314 | sigma-54 dependent transcriptional regulator | −1.6 | |
| Bphy_5338 | response regulator receiver protein | −3.5 | |
| Bphy_5668 | integral membrane sensor signal transduction histidine kinase | −5.8 | |
| Bphy_5669 | two component, sigma54 specific, Fis family transcriptional regulator | −4.9 | |
| Bphy_5670 | diguanylate phosphodiesterase | −3.5 | |
| Bphy_5974 | non-specific serine/threonine protein kinase | −2.6 | |
| Bphy_5975 | histidine kinase | −1.7 | |
| Bphy_5989 | extracellular solute-binding protein | −1.4 | |
| Bphy_6398 | CheB methylestease | −2.6 | |
| Transcription | |||
| Bphy_2946 | anti-sigma28 factor FlgM | flgM | −3.4 |
| Bphy_3962 | two component heavy metal response transcriptional regulator | −1.5 | |
| Bphy_4638 | transcriptional activator FlhC | flhC | −2.3 |
| Bphy_5304 | RNA polymerase factor sigma-54 | −2.9 | |
| Bphy_5333 | response regulator receiver protein | −3.7 | |
| Bphy_5662 | MarR family transcriptional regulator | −1.3 | |
| Bphy_7189 | Fis family GAF modulated sigma54 specific transcriptional regulator | −1.1 | |
| Translation, Ribosomal Structure and Biogenesis | |||
| Bphy_0314 | 50S ribosomal protein L25/general stress protein Ctc | −1.0 | |
| Not Assigned to a Functional Category | |||
| Bphy_1514 | putative lipoprotein | −3.3 | |
| Bphy_1768 | PRC-barrel domain-containing protein | −1.6 | |
| Bphy_5614 | phosphoesterase PA-phosphatase related | −3.3 | |
| Carbon Source | Utilization of Carbon | ||
|---|---|---|---|
| wt | σ54 mt | σ54 comp | |
| Fumaric acid | + | – | + |
| Malic acid | + | – | + |
| Succinic acid | + | – | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lardi, M.; Liu, Y.; Hug, S.; Bolzan de Campos, S.; Eberl, L.; Pessi, G. Paraburkholderia phymatum STM815 σ54 Controls Utilization of Dicarboxylates, Motility, and T6SS-b Expression. Nitrogen 2020, 1, 81-98. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen1020008
Lardi M, Liu Y, Hug S, Bolzan de Campos S, Eberl L, Pessi G. Paraburkholderia phymatum STM815 σ54 Controls Utilization of Dicarboxylates, Motility, and T6SS-b Expression. Nitrogen. 2020; 1(2):81-98. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen1020008
Chicago/Turabian StyleLardi, Martina, Yilei Liu, Sebastian Hug, Samanta Bolzan de Campos, Leo Eberl, and Gabriella Pessi. 2020. "Paraburkholderia phymatum STM815 σ54 Controls Utilization of Dicarboxylates, Motility, and T6SS-b Expression" Nitrogen 1, no. 2: 81-98. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen1020008