
Nitrogen Dynamics in Wetland Systems and Its Impact on Biodiversity

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Method
3. What Are Wetlands?
3.1. Classification of Wetlands
3.2. Functions of Wetlands
4. Constructed Wetlands and Nitrogen Cycling
4.1. Limitations
4.2. Types
4.2.1. Surface Flow System
4.2.2. Sub-Surface Flow System
4.2.3. Partially Saturated Vertical Flow Constructed Wetland: A Hybrid System
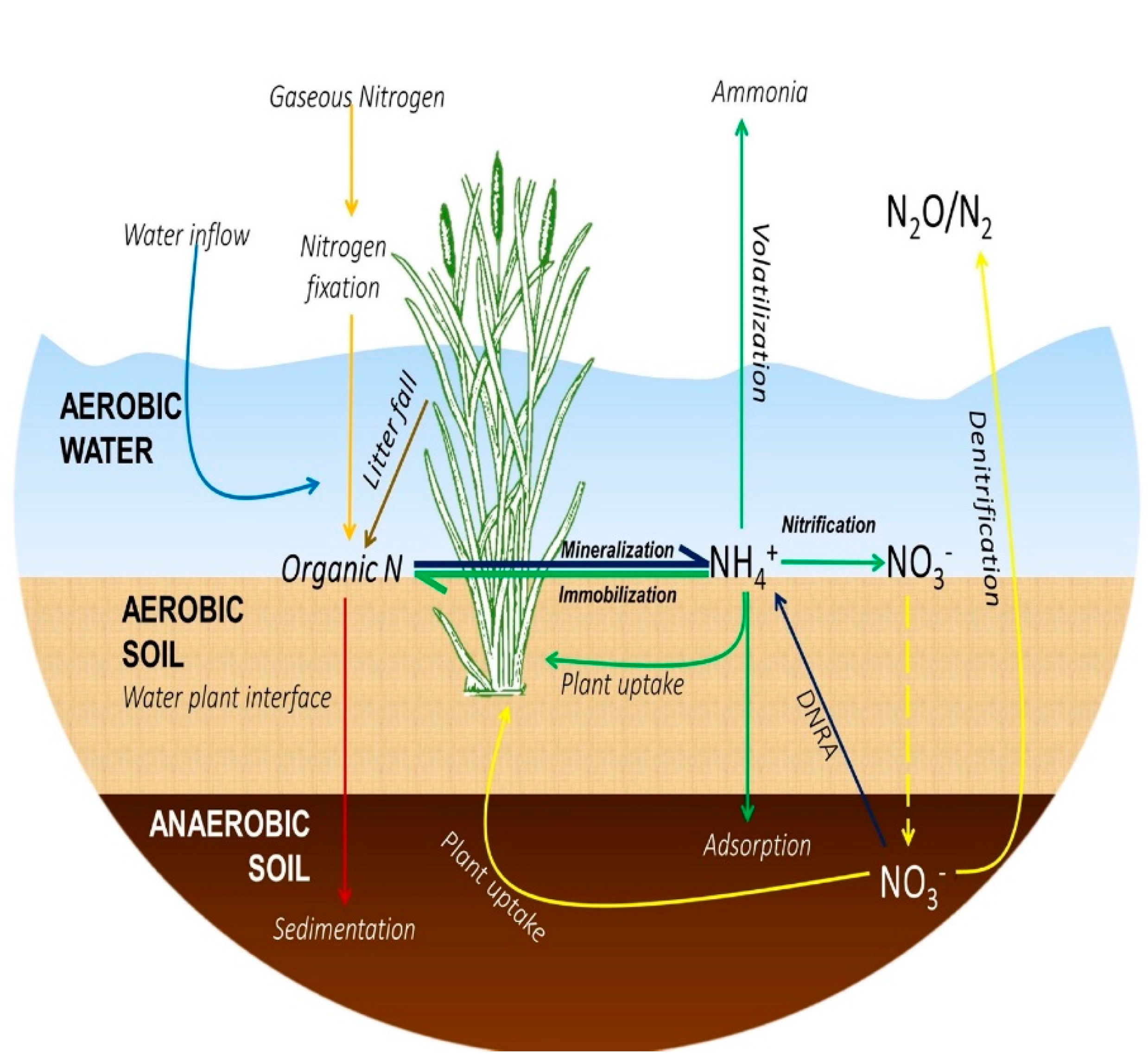
5. Nitrogen Dynamics in Wetland Systems
5.1. Mineralization
5.2. Nitrification
5.3. Denitrification
5.3.1. Measurement of Denitrification
5.3.2. Acetylene Inhibition Method
5.4. Volatilization
5.5. Sedimentation
5.6. Plant Uptake and Matrix Adsorption
6. Role of Faunal Communities in Wetlands
7. Impact of Vegetation on Nitrification and Denitrification
8. Oxygen Transport
9. Other Ecological Factors Influencing Nitrogen Removal Efficiency
9.1. Temperature
9.2. Hydraulic Residence Time
9.3. Vegetation Type
10. Wetland Nutrient Retention
11. Operation and Management of Wetlands for Efficient Nitrogen Attenuation
12. Role of Wetlands in Biodiversity Enhancement
13. Future Prospects
14. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- UNICEF Water: Sanitation and Hygiene Annual Report; UNICEF: New York, NY, USA, 2008.
- O’Geen, A.T.; Budd, R.; Gan, J.; Maynard, J.J.; Parikh, S.J.; Dahlgren, R.A. Mitigating nonpoint source pollution in agriculture with constructed and restored wetlands. Adv. Agron. 2010, 108, 1–76. [Google Scholar] [CrossRef]
- Howarth, R.W.; Billen, G.; Swaney, D.; Townsend, A.; Jaworski, N.; Lajtha, K.; Downing, J.A.; Elmgren, R.; Caraco, N.; Jordan, T.; et al. Regional nitrogen budgets and riverine inputs of N and P for the drainages to the North Atlantic Ocean: Natural and human influences. Biogeochemistry. 1996, 35, 75–139. [Google Scholar] [CrossRef]
- Hägg, H.E. Nitrogen Land-Sea Fluxes in The Baltic Sea Catchment—Empirical Relationships and Budgets. Ph.D. Thesis, University of Stockholm, Stockholm, Sweden, 2010. [Google Scholar]
- Encyclopedia of Soils in The Environment. In Nitrates; Powlson, D.S.; Addiscott, T.M.; Hillel, D. (Eds.) Elsevier: Amsterdam, The Netherlands, 2005; pp. 21–31. [Google Scholar] [CrossRef]
- Meisinger, J.J.; Palmer, R.E.; Timlin, D.J. Effects of tillage practices on drainage and nitrate leaching from winter wheat in the Northern Atlantic Coastal-Plain USA. Soil Tillage Res. 2015, 151, 18–27. [Google Scholar] [CrossRef]
- Richardson, C.J. Freshwater Wetlands: Transformers, Filters, or Sinks? In Freshwater Wetlands and Wildlife; Sharitz, R.R., Gibbons, J.W., Eds.; The Department of Energy (DOE) Office of Scientific and Technical Information: Oak Ridge, TN, USA, 1989; pp. 25–46. [Google Scholar]
- Mitsch, W.J. Global Wetlands: Old World and New; Elsevier Science: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Acreman, M.C. Impact Assessment of Wetlands: Focus on Hydrological and Hydrogeological Issues. Phase 2 Report. Environment Agency, Bristol (W6-091) and Centre for Ecology and Hydrology, Wallingford (C01996); Environment Agency: Bristol, UK, 2004.
- Cowardin, L.V.; Carter, F.C. Classification of Wetlands and Deep Water. Habitats of the United States; US Department of Interior, Fish and Wildlife Service: Washington, DC, USA, 1979.
- United States Environmental Protection Agency. 2006. Available online: http://www.epa.gov/wetlands (accessed on 28 April 2021).
- Sundaravadivel, M.; Vigneswaran, S. Constructed Wetlands for Wastewater treatment. Crit. Rev. Env. Sci. Technol. 2001, 31, 351–409. [Google Scholar] [CrossRef]
- National Wetland Policy of Nepal; Ministry of Forests and Soil Conservation, Singh Durbar: Kathmandu, Nepal, 2003.
- Thorsell, J.; Levy, R.F.; Sigaty, T. A Global Overview of Wetland and Marine Protected Areas on The World Heritage List. In Natural Heritage Programme; IUCN: Gland, Switzerland, 1997. [Google Scholar]
- Kadlec, R.H.; Knight, R.L. Treatment Wetlands; Lewis-CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- EPA (U.S. Environmental Protection Agency). Better Assessment Science Integrating Point and Nonpoint Sources (BASINS), User’s Manual, Version 3.0. EPA-823-H-01-001; Office of Water, U.S. Environmental Protection Agency: Washington, DC, USA, 2001.
- Richey, J.E.; Melak, J.M.; Aufdenkampe, A.K.; Ballester, V.M.; Hess, L.L. Outgassing from Amazonian rivers and wetlands as a large tropical source of atmospheric CO2. Nature 2002, 416, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Torrens, A.; Molle, P.; Boutin, C.; Salgot, M. Impact of design and operation variables on the performance of vertical-flow constructed wetlands and intermittent sand filters treating pond effluent. Water Res. 2009, 43, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Wallace, S.; Jiggins, J.; Crolla, A.; Kinsley, C.; Bachanc, A.; Verkuijl, S. Highrate Ammonium Removal in Aerated Engineered Wetlands. In Proceedings of the 10th International Conference on Wetland Systems for Water Pollution Control, Lisbon, Portugal, 23–29 September 2006. [Google Scholar]
- Tiedje, J.M. Ecology of Denitrification and Dissimilatory Nitrate Reduction to Ammonium. In Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John and Wiley & Sons, Inc.: New York, NY, USA, 1988; pp. 179–244. [Google Scholar]
- Woodward, K.B.; Fellows, C.S.; Conway, C.L.; Hunter, H.M. Nitrate removal, denitrification and nitrous oxide production in the riparian zone of an ephemeral stream. Soil Biol. Biochem. 2009, 41, 671–680. [Google Scholar] [CrossRef]
- Lin, Y.F.; Jing, S.R.; Lee, D.Y.; Wang, T.W. Effect of macrophytes and external carbon sources on nitrate removal from groundwater in constructed wetlands. Environ. Pollut. 2002, 119, 413–420. [Google Scholar] [CrossRef]
- Mayo, A.W.; Bigambo, T. Nitrogen transformation in horizontal subsurface flow constructed wetlands I: Model development. Phys. Chem. Earth Parts A B C 2005, 30, 658–667. [Google Scholar] [CrossRef]
- Groffman, P.M.; Altabet, M.A.; Böhlke, J.K.; Butterbach-Bahl, K.; David, M.B.; Firestone, M.K.; Giblin, A.E.; Kana, T.M.; Nielsen, L.P.; Voytek, M.A. Methods for Measuring Denitrification: Diverse Approaches to a Difficult Problem. Ecol. Appl. 2006, 16, 2091–2122. [Google Scholar] [CrossRef]
- Nelson, J.W.; Kadlec, J.A. A conceptual approach to relating habitat structure and macroinvertebrate production in freshwater wetlands. Trans. N. Amer. Wildl. Nat. Resourc. Conf. 1984, 49, 262–270. [Google Scholar]
- Swanson, G.A.; Meyer, M.I. Impact of fluctuating water levels on the feeding ecology of breeding blue-winged teal. J. Wildl. Manag. 1977, 41, 426–433. [Google Scholar] [CrossRef]
- Kramer, P.J.; Boyer, J.S. Water Relations of Plants and Soil; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Moreno, C.; Farahbakhshadzad, N.; Morrison, G.M. Ammonia removal from oil refinery effluent in vertical upflow macrophyte column systems. Water Air Soil Pollut. 2002, 135, 237–247. [Google Scholar] [CrossRef]
- Hume, N.P.; Fleming, M.S.; Horne, A.J. Denitrification potential and carbon quality of four aquatic plants in wetland microcosms. Soil Sci. Soc. Am. J. 2002, 66, 1706–1712. [Google Scholar] [CrossRef] [Green Version]
- Westlake, J.; Kvêt, J.; Szczepanski, A. The Production Ecology of Wetlands; Cambridge university press: Cambridge, UK, 1998. [Google Scholar]
- Hamilton, W.A. Biofilms: Microbial Interactions and Metabolic Activities. In Ecology of Microbial Communities; Fletcher, M., Gray, T.R.G., Jones, J.G., Eds.; Cambridge University Press: Cambridge, UK, 1987; pp. 361–385. [Google Scholar]
- Eriksson, P.G.; Andersson, J.L. Potential nitrification and cation exchange on litter of emergent, freshwater macrophytes. Freshwater Biol. 1999, 42, 479–486. [Google Scholar]
- Reddy, K.R.; D’Angelo, E.M.; DeBusk, T.A. Oxygen transport through aquatic macrophytes: The role in wastewater treatment. J. Environ. Qual. 1989, 19, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.B.; Liu, P.; Yang, Y.S.; Chen, W.R. Phytoremediation of urban wastewater by model wetlands with ornamental hydrophytes. J. Environ. Sci. 2007, 19, 902–990. [Google Scholar] [CrossRef]
- Zurita, F.; Anda, J.D.; Belmont, M.A. Performance of laboratory-scale wetlands planted with tropical ornamental plants to treat domestic wastewater. Water Qual. Res. 2006, 41, 410–417. [Google Scholar] [CrossRef]
- Green, C.; Tunstall, S.; Garner, J.; Ketteridge, A.M. Benefit Transfer: Rivers and Coasts. Paper Prepared for The CEGB Meeting on Benefit Transfer, H.M. Treasury. Publication No. 231; Flood Hazard Research Centre, Middlesex University: London, UK, 1994. [Google Scholar]
- Lahav, O.; Artzi, E.; Tarre, S.; Green, M. Ammonium removal using a novel unsaturated flow biological filter with passive aeration. Water Res. 2010, 35, 397–404. [Google Scholar] [CrossRef]
- Sun, G.; Austin, D. Completely autotrophic nitrogen-removal over nitrite in lab-scale constructed wetlands: Evidence from a mass balance study. Chemosphere 2007, 68, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yan, C.; Liu, J.; Guan, W.; Singh, R.P.; Cao, C.; Xiao, J. Feasibility Study of Vertical Flow Constructed Wetland for Tertiary Treatment of Nanosilver Wastewater and Temporal-Spatial Distribution of Pollutants and Microbial Community. J. Environ. Manag. 2019, 245, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wissing, F. Wasserreinigung mit Pflanzen; Verlag Eugen Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- Hammer, D.A. Designing constructed wetlands systems to treat agricultural nonpoint source pollution. Ecol. Eng. 1992, 1, 49–82. [Google Scholar] [CrossRef]
- Axler, R.; McCarthy, B.; Henneck, J. NERCC Individual Alternative Wastewater Treatment Systems: Pollutant Removal in 2003 and Long-Term Performance. Northeast Regional Correctional Center, St. Louis County, Duluth, Minnesota. NRRI Technical Report NRRI/TR-2004/28. Natural Resources Research Institute University of Minnesota-Duluth. 5013 Miller Trunk Highway; Natural Resources Research Institute University of Minnesota-Duluth: Duluth, MN, USA, 2004. [Google Scholar]
- Baker, C.J.; Maltby, E. Nitrate Removal by River Marginal Wetlands: Factors Affecting the Provision of a Suitable Denitrification Environment. In Hydrology and Hydrochemistry of British Wetlands; Hughes, J., Healthwaite, L., Eds.; Wiley: Chichester, UK, 1995; pp. 291–313. [Google Scholar]
- Bridgham, S.D.; Updegraff, K.; Pastor, J. Carbon, nitrogen, and phosphorus mineralization in northern wetlands. Ecology 1998, 79, 1545–1561. [Google Scholar] [CrossRef]
- Burt, T.P.; Matchett, L.S.; Haycock, N.E. Floodplain as Buffer Zones. In United Kingdom Floodplains; Westbury Publishing: Otley: West Yorlshire, UK, 1998. [Google Scholar]
- Busse, L.R.; Gunkel, G. Riparian alder fens-source or sink for nutrients and dissolved organic carbon?—2 Major sources and sinks. Limnologica 2002, 32, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Chauvelon, P. A wetland managed for agriculture as an interface between the Rhone river and the Vaccares lagoon (Camargue, France): Transfers of water and nutrients. Hydrobiologia 1998, 373, 181–191. [Google Scholar] [CrossRef]
- Cooper, P.F.; Findlater, B.C. Constructed Wetlands in Water Pollution Control; Pergamon Press: New York, NY, USA, 1990. [Google Scholar]
- Daniels, R.B.; Gilliam, J.W. Sediment and chemical load reduction by grass and riparian filters. Soil Sci. Soc. Amer. J. 1996, 60, 1231–1238. [Google Scholar] [CrossRef]
- Dorge, J. Modeling nitrogen transformations in freshwater wetlands. Estimating nitrogen retention and removal in natural wetlands in relation to their hydrology and nutrient loadings. Ecol. Model. 1994, 75–76, 409–420. [Google Scholar] [CrossRef]
- Fleischer, S.; Joelsson, A.; Stibe, L. The Potential Role of Ponds as Buffer Zones. In Buffer Zones: Their Processes and Potential in Water Protection; Haycock, N.E., Burt, T.P., Goulding, K.W.T., Pinay, G., Eds.; Quest Environmental: Harpenden, UK, 1997. [Google Scholar]
- Gersberg, R.M.; Elkins, B.V.; Goldman, C.R. Nitrogen removal in artificial wetlands. Water Res. 1983, 17, 1009–1014. [Google Scholar] [CrossRef]
- Green, M.B.; Upton, J. Reed Bed Treatment for Small Communities—UK Experience. Ch. 57 in Constructed Wetlands for Water Quality Improvement; Moshiri, G.A., Ed.; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Hansson, L.A.; Bronmark, C.; Nilsson, P.; Anders Abjornsson, K. Conflicting demands on wetland ecosystem services: Nutrient retention, biodiversity, or both? Freshw. Biol. 2005, 5, 705–714. [Google Scholar] [CrossRef]
- Hanson, G.C.; Groffman, P.M.; Gold, A.J. Denitrification in riparian wetlands receiving high and low groundwater nitrate inputs. J. Environ. Qual. 1994, 23, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Haycock, N.E.; Burt, T.P. Role of floodplain sediments in reducing the nitrate concentration of subsurface run-off: A case study in the Cotswolds, UK. Hydrol. Process. 1993, 7, 287–295. [Google Scholar] [CrossRef]
- Hey, D.L.; Kenimer, A.L.; Barrett, K.R. Water quality improvement by four experimental wetlands. Ecol. Eng. 1994, 3, 381–394. [Google Scholar] [CrossRef]
- Jansson, M.; Andersson, R.; Berggren, H.; Leonardson, L. Wetlands and lakes as nitrogen traps. Ambio 1994, 23, 320–325. [Google Scholar]
- Johnston, C.A.; Bubenzer, G.D.; Lee, G.B.; Madison, F.W.; McHenry, J.R. Nutrient trapping by sediment deposition in a seasonally flooded lakeside wetland. J. Environ. Qual. 1984, 13, 283–290. [Google Scholar] [CrossRef]
- Jordan, T.E.; Correll, D.L.; Weller, D.E. Nutrient interception by a riparian forest receiving inputs from adjacent cropland. J. Environ. Qual. 1993, 22, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Jordan, T.E.; Whigham, D.F.; Hofmockel, K.H.; Pittek, M.A. Nutrient and sediment removal by a restored wetland receiving agricultural runoff. J. Environ. Qual. 2003, 32, 1534–1547. [Google Scholar] [CrossRef]
- Kim, B.; Gautier, M.; Prost-Boucle, S.; Molle, P.; Michel, P.; Gourdon, R. Performance evaluation of partially saturated vertical-flow constructed wetland with trickling filter and chemical precipitation for domestic and winery wastewaters treatment. Ecol. Eng. 2014, 71, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Leonardson, L.; Bengtsson, L.; Davidsson, T.; Persson, T.; Emanuelsson, U. Nitrogen retention in artificially flooded meadows Nitrogen retention in artificially flooded meadows. Ambio 1994, 23, 332–341. [Google Scholar]
- Maltby, E.; Hogan, D.V.; Oliver, G.A. Wetland Soil Hydrological and Ecosystem Functioning. In Hydrology and Hydrochemistry of British Wetlands; Hughes, J.M.R., Heathwaite, A.L., Eds.; Wiley: Chicester, UK, 1995; pp. 325–362. [Google Scholar]
- Mander, U.; Lomus, K.; Kuusemets, V.; Ivask, M. The Potential Role of Wet Meadows and Grey Alder Forests as Buffer Zones. In Buffer Zones: Their Processes and Potential in Water Protection; Haycock, N.E., Burt, T.P., Goulding, K.W.T., Pinay, G., Eds.; Quest Environmental: Harpenden, UK, 1997. [Google Scholar]
- Nakase, C.; Zurita, F.; Nani, G.; Reyes, G.; Fernández-Lambert, G.; Cabrera-Hernández, A.; Sandoval, L. Nitrogen removal from domestic wastewater and the development of tropical ornamental plants in partially saturated mesocosm-scale constructed wetlands. Int. J. Environ. Res. Public Health 2019, 16, 4800. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.M.; Clausen, J.C.; Neafsey, J.A. Seasonal performance of a wetland constructed to process dairy milkhouse wastewater in Connecticut. Ecol. Eng. 2000, 14, 181–198. [Google Scholar] [CrossRef]
- Paturno, J.; Russell, J. Natural wetland polishing effluent discharging to Wooloweyah Lagoon. Water Sci. Technol. 1994, 29, 185–192. [Google Scholar] [CrossRef]
- Peterson, S.B.; Teal, J.M. The role of plants in ecologically engineered wastewater treatment systems. Ecol. Eng. 1996, 6, 137–148. [Google Scholar] [CrossRef]
- Phipps, R.G.; Crumpton, W.G. Factors affecting nitrogen loss in experimental wetlands with different hydrologic loads. Ecol. Eng. 1994, 3, 399–408. [Google Scholar] [CrossRef]
- Prior, H. Total Nitrogen and Total Phosphorous Cycling in Riparian Ecosystems. Ph.D. Thesis, University of Reading, Whiteknights, Reading, UK, 1998. [Google Scholar]
- Raisin, G.W.; Mitchell, D.S. The use of wetlands for the control of non-point source pollution. Water Sci. Technol. 1995, 32, 177–186. [Google Scholar] [CrossRef]
- Saeed, T.; Sun, G. Pollutant removals employing unsaturated and partially saturated vertical flow wetlands: A comparative study. Chem. Eng. J. 2017, 325, 332–341. [Google Scholar] [CrossRef]
- Sandoval, L.; Marín-Muñiz, J.L.; Zamora-Castro, S.A.; Sandoval-Salas, F.; Alvarado-Lassman, A. Evaluation of wastewater treatment by microcosms of vertical subsurface wetlands in partially saturated conditions planted with ornamental plants and filled with mineral and plastic substrates. Int. J. Environ. Res. Public Health 2019, 16, 167. [Google Scholar] [CrossRef] [Green Version]
- Schipper, L.A.; Vojvodic-Vukovic, M. Nitrate removal from groundwater and denitrification rates in a porous treatment wall amended with sawdust. Ecol. Eng. 2000, 14, 269–278. [Google Scholar] [CrossRef]
- Schwer, C.B.; Clausen, J.C. Vegetation filter treatment of diary milk house wastewater. J. Environ. Qual. 1989, 18, 446–451. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A. Nutrient dynamics in minerotrophic peat mires. Aquat. Bot. 1986, 25, 117–167. [Google Scholar] [CrossRef]
- Bang, W.H.; Jung, Y.; Park, J.W.; Lee, S.; Maeng, S.K. Effects of hydraulic loading rate and organic load on the performance of a pilot-scale hybrid VF-HF constructed wetland in treating secondary effluent. Chemosphere 2019, 218, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Ecology—Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; SchererLorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Reed, P.B. National List of Plant Species That Occur in Wetlands: National summary. U.S. Fish Wildl. Serv. Biol. Rep. 1988, 88, 244. [Google Scholar]
- Lugo, A.E.; Brinson, M.; Brown, S. Forested Wetlands. Ecosystems of the World; Elsevier Science: Amsterdam, The Netherlands, 1990; p. 15. [Google Scholar]
- Bacon, P.R. Use of wetlands for tourism in the Insular Caribbean. Ann. Tour. Res. 1987, 14, 104–117. [Google Scholar] [CrossRef]
- U.S. Fish & Wildlife Service. A Joint Venture Proposal for Designation as Wetlands of International Importance: The Cache River and Cypress Creek Wetlands Area of South Illinois. Report of the U.S. Fish & Wildlife Service and Illinois Department of Conservation; U.S. Fish and Wildlife Service: Washington, DC, USA, 1994.
- Bouchard, V.; Frey, S.D.; Gilbert, J.M.; Reed, S.E. Effects of macrophyte functional group richness on emergent freshwater wetland functions. Ecology 2007, 88, 2903–2914. [Google Scholar] [CrossRef]
- Weisner, S.E.; Thiere, G. Effects of vegetation state on biodiversity and nitrogen retention in created wetlands: A test of the biodiversity-ecosystem functioning hypothesis. Freshw. Biol. 2010, 55, 387–396. [Google Scholar] [CrossRef]
- Keddy, P. Putting the plants back into plant ecology: Six pragmatic models for understanding and conserving plant diversity. Ann. Bot. 2005, 96, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Ardon, M.; Montanari, S.; Morse, J.L.; Doyle, M.W.; Bernhardt, E.S. Phosphorus export from a restored wetland ecosystem in response to natural and experimental hydrologic fluctuations. J. Geophys. Res. Biogeosci. 2010, 115, G04031. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, M.; White, J.; Coghlan, C.; Reddy, K. Influence of hydropattern and vegetation type on phosphorus dynamics in flow-through wetland treatment systems. Ecol. Eng. 2011, 37, 1369–1378. [Google Scholar] [CrossRef]
- Walker, W.W.; Kadlec, R.H. Modeling phosphorus dynamics in Everglades wetlands and stormwater treatment areas. Crit. Rev. Environ. Sci. Technol. 2011, 41, 430–446. [Google Scholar] [CrossRef]
- Bastviken, S.K.; Weisner, S.E.; Thiere, G.; Svensson, J.M.; Ehde, P.M.; Tonderski, K.S. Effects of vegetation and hydraulic load on seasonal nitrate removal in treatment wetlands. Ecol. Eng. 2009, 35, 946–952. [Google Scholar] [CrossRef]
- Pollock, M.M.; Naiman, R.J.; Hanley, T.A. Plant species richness in riparian wetlands. A test of biodiversity theory. Ecol. 1998, 79, 94–105. [Google Scholar]
- Thiere, G.; Milenkovski, S.; Lindgren, P.E.; Sahlen, G.; Berglund, O.; Weisner, S.E.B. Wetland creation in agricultural landscapes: Biodiversity benefits on local and regional scales. Biol. Conserv. 2009, 142, 964–973. [Google Scholar] [CrossRef]
- Hey, D.L.; Cardamone, M.A.; Sather, J.H.; Mitsch, W.J. Restoration of Riverine Wetlands: The Des Plaines River Wetlands Demonstration Project; An Introduction to Ecotechnology; Mitsch, W.J., Jorgensen, S.E., Eds.; Ecological Engineering; John Wiley & Sons: New York, NY, USA, 1989; pp. 159–183. [Google Scholar]
- Ferguson, P.; Robinson, R.N.; Press, M.C.; Lee, J.A. Element concentrations in five Sphagnum species in relation to atmospheric pollution. Bryologist 1984, 13, 107–114. [Google Scholar]
- Wheeler, B.D.; Giller, K.E. Species richness of herbaceous fen vegetation in Broadland, Norfolk in relation to the quantity of above-ground plant material. J. Ecol. 1982, 70, 179–200. [Google Scholar] [CrossRef]
- Schuurkes, J.A.A.R.; Kok, C.J.; den Hartog, C. Ammonium ad nitrate uptake by aquatic plants from poorly buffered and acidified waters. Aquat. Bot. 1986, 24, 131–146. [Google Scholar]
- Blank, P.J.; Dively, G.P.; Gill, D.E.; Rewa, C.A. Bird community response to filter strips in Maryland. J. Wildl. Manag. 2011, 75, 116–125. [Google Scholar] [CrossRef]
- Morris, J.D. Effects of nitrogen loading on wetland ecosystems with particular reference to atmospheric deposition. Annu. Rev. Ecol. Syst. 1991, 22, 257–279. [Google Scholar] [CrossRef]
- Langley, J.A.; Megonigal, J.P. Ecosystem response to elevated CO2 levels limited by nitrogen-induced plant species shift. Nature 2010, 466, 96–99. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ref No. | Year | Author(s) | Wetland Name | Wetland Type | Nutrient Retention | Country |
|---|---|---|---|---|---|---|
| [42] | 2004 | Axler et al. | Unknown | Constructed | N, TN | Duloth, Minnesota |
| [43] | 1995 | Baker and Maltby | Kismeldon Meadows and Bradford Mill | Riparian | NO3, NO4 | UK |
| [44] | 1998 | Bridgham et al. | Unknown | Stream | N, P | Minnesota |
| [45] | 1998 | Burt et al. | R.Leach floodplain | Floodplain | NO3 | UK |
| [46] | 2002 | Busse and Gunkel | Unknown | Riparian alder fen | N, P | Berlin, Germany |
| [47] | 1998 | Chauvelon | Rhone river delta | Riverin delta | N, P | UK |
| [48] | 1990 | Cooper | Scotsman Valley, NZ | Riparian | NO3 | New Zealand |
| [49] | 1996 | Daniel and Gilliam | Cecil soil area | Riparian | N, P | USA |
| [50] | 1994 | Dorge | Rabis Baek | Peatland | NO3 | Denmark |
| [51] | 1993 | Fleischer et al. | Unknown | Ponds | N | Sweden |
| [52] | 1983 | Gersberg et al. | Unknown | Constructed | N, P | Santhe, CA |
| [53] | 1993 | Green and Upton | Unknown | Constructed | N, P | UK |
| [54] | 2005 | Hanson et al. | Unknown | Constructed | N, P | Southern Sweden |
| [55] | 1994 | Hanson et al. | nr Kingston | Riparian | NO3 | USA |
| [56] | 1993 | Haycock and Burt | R.Leach floodplain | Floodplain | NO3 | UK |
| [57] | 1994 | Hey et al. | Des Plains River Wetlands | Riparian | NO3, P | Chicago Illinois, USA |
| [58] | 1994 | Jansson et al. | River Raan catchment | Stream | N | Southern Sweden |
| [59] | 1984 | Johnston et al. | nr White Clay Lake | Marsh/swamp | N, P | USA |
| [60] | 1993 | Jordan et al. | Chester River Catchment | Floodplain | NO3, TP | USA |
| [61] | 2003 | Jordan et al. | Kent Island | Marsh/swamp | TN, TP | USA |
| [15] | 1996 | Kadlec and Knight | Unknown | Constructed | N, P | North America |
| [62] | 2014 | Kim et al. | Unknown | Partially saturated vertical flow wetland | NO3, TKN | France |
| [63] | 1994 | Leonardson et al. | Unknown | Constructed | N, NO3, NH4 | Sweden |
| [64] | 1995 | Maltby et al. | Floodplains in Devon | Floodplain | N | UK |
| [65] | 1997 | Mander et al. | Poriji River catchment | Riparian | N, P | Estonia |
| [66] | 2019 | Nakase et al. | Unknown | Partially saturated vertical flow wetland | NO3, TN | Mexico |
| [67] | 2000 | Newman et al. | Unknown | Constructed | N, TN, P | Connecticut |
| [68] | 1994 | Patruno and Russel | Yamba wetland | Marsh/swamp | N, P | Australia |
| [69] | 1996 | Peterson and Teal | Unknown | Constructed | N | New England |
| [70] | 1994 | Phipps and Grumpton | Unknown | Constructed | N, P | USA |
| [71] | 1998 | Prior | R.Lambourn floodplain | Floodplain | N, P | UK |
| [72] | 1995 | Raisin and Mitchell | Humphrey’s wetland | Marsh/swamp | TPN | Australia |
| [73] | 2017 | Saeed and Sun | Unknown | Partially saturated vertical flow wetland | TN | Bangladesh |
| [74] | 2019 | Sandoval et al. | Unknown | Partially saturated vertical flow wetland | NO3 | Mexico |
| [75] | 2000 | Schipper and Vojvodic’ Vukovic | Unknown | Constructed | NO3 | New Zealand |
| [76] | 1989 | Schwer and Clausen | nr Charlotte | Riparian | TP, TN | USA |
| [77] | 1986 | Verhoeven | Unknown | Peatmires/fens/bogs | N, P | US and Europe |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousaf, A.; Khalid, N.; Aqeel, M.; Noman, A.; Naeem, N.; Sarfraz, W.; Ejaz, U.; Qaiser, Z.; Khalid, A. Nitrogen Dynamics in Wetland Systems and Its Impact on Biodiversity. Nitrogen 2021, 2, 196-217. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen2020013
Yousaf A, Khalid N, Aqeel M, Noman A, Naeem N, Sarfraz W, Ejaz U, Qaiser Z, Khalid A. Nitrogen Dynamics in Wetland Systems and Its Impact on Biodiversity. Nitrogen. 2021; 2(2):196-217. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen2020013
Chicago/Turabian StyleYousaf, Anum, Noreen Khalid, Muhammad Aqeel, Ali Noman, Nayab Naeem, Wajiha Sarfraz, Ujala Ejaz, Zonaira Qaiser, and Arifa Khalid. 2021. "Nitrogen Dynamics in Wetland Systems and Its Impact on Biodiversity" Nitrogen 2, no. 2: 196-217. https://0-doi-org.brum.beds.ac.uk/10.3390/nitrogen2020013