Socio-Ecological Contingencies with Climate Changes over the Prehistory in the Mediterranean Iberia

 ,
,  ,
,
Abstract
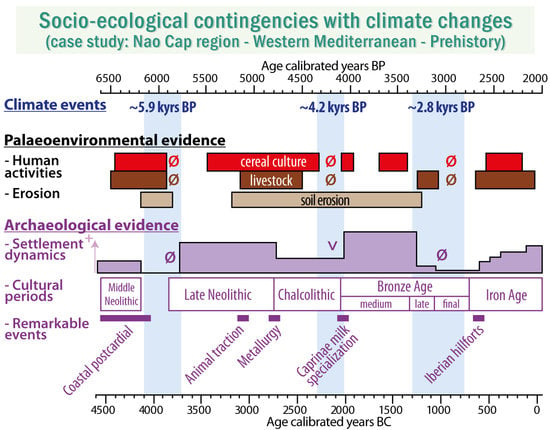
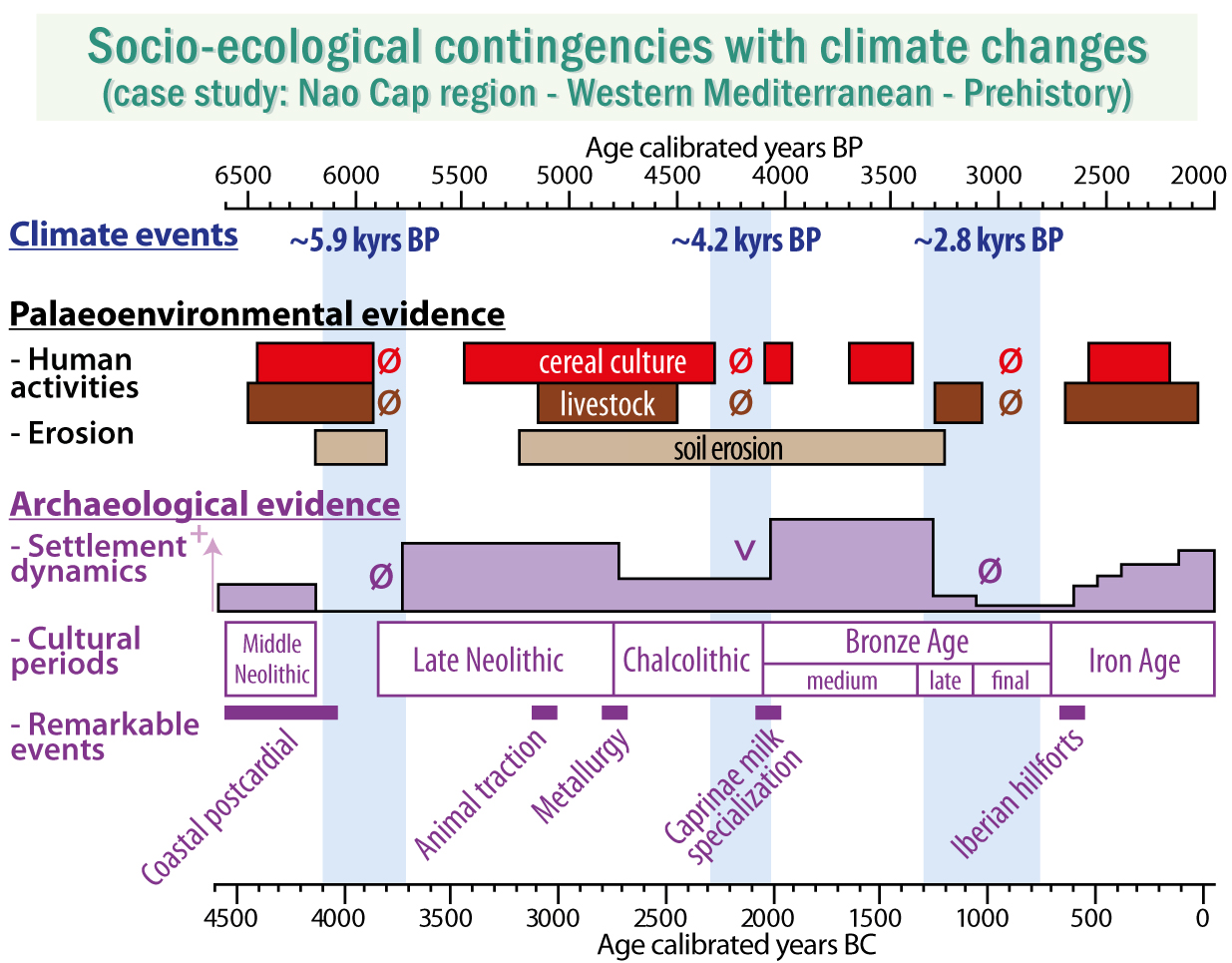
:
1. Introduction
- to reconstruct the morphogenetic evolution of the coastal plain of Pego-Oliva linked with the Mid-Holocene stabilization of the sea-level rise;
- to characterize vegetation successions, fire dynamics, and soil erosion at the interface between the coastal plain and abutting hills over the last 7000 years;
- to decipher the local impacts of the agropastoral activities from the Post-Cardial Neolithic (5000 cal. BCE) to the end of the Iron Age (2000 cal. BCE); and
- to investigate the influence of three Rapid Climate Changes (at ~5.9, ~4.2, and ~2.8 kyr BP events), on ancient socio-ecosystem functioning.
2. Study Site
3. Materials and Methods
4. Results
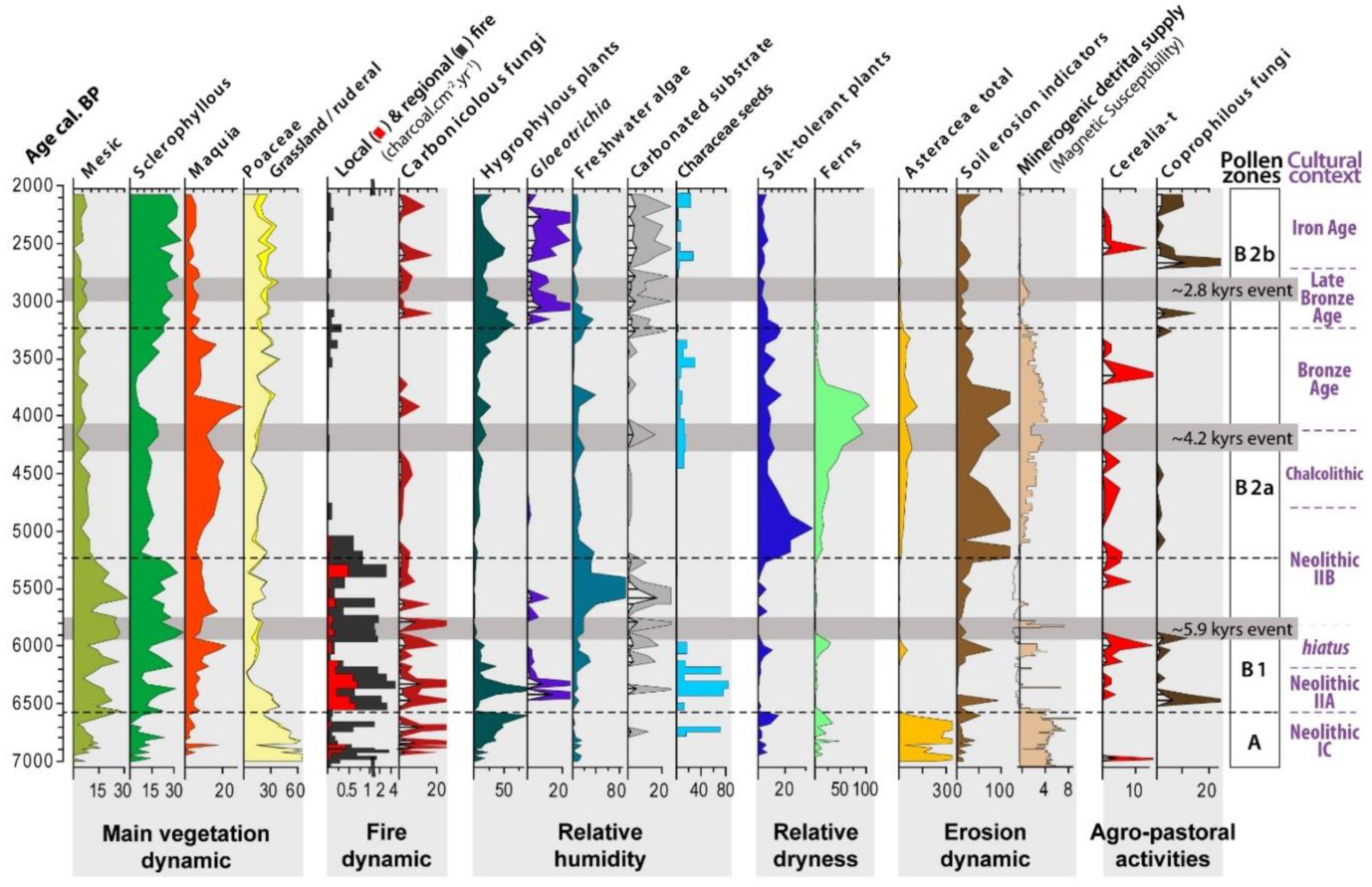
4.1. Sedimentology, Charcoal, and Chronology
4.2. Palynology
4.2.1. Zone A (366–438 cm, 7000–6600 cal. BP)
4.2.2. Sub-Zone B1 (253–362 cm, 6600–5300 cal. BP)
4.2.3. Sub-Zone B2a (154–249 cm, 5200–3200 cal. BP)
4.2.4. Sub-Zone B2b (48–150 cm, 3200–2000 cal. BP)
5. Interpretations and Discussion
5.1. Paludification of River Mouth with the Slowdown of the Sea-Level Rise (7420–6600 cal. BP)
5.2. Optimum of the Mid-Holocene Forest at the End of the Neolithic (6600–5300 cal. BP)
5.3. Mid-Holocene Forest Demise During the Chalcolithic and the Bronze Age (5300–3200 cal. BP)
5.4. Xerophilous Vegetation Turnover from Late Bronze to Iron Age (3200–2000 cal. BP)
6. Conclusions
- characterize the Mid-Holocene Climatic Optimum of the mesophilous oak forest that developed in the inner valley from the beginning of the sequence to 5300 cal. BP, confirmed by comparisons conducted with anthracological records from the archeological sites;
- underline the role of fire in the vegetation turnover at 5300 cal. BP (i.e., mixed oak forests replaced by sclerophyllous trees and matorral scrubs) and at 3200 cal. BP (i.e., the dominance of xerophytics);
- show the variability of karst spring outflow influencing the wetland ecology, with phases of fresh running water at 7000–6000 and 3200–2000 cal. BP, a pseudo-lacustrine freshwater phase (6000–5300 cal. BP), and a phase of desiccation (5300–3200 cal. BP) reaching a maximum at ~4200 cal. BP; and
- provide evidence of one short soil erosion event occurring at ca. 5900 cal. BP that likely relates to the lower level of vast coalescent dejection fans [33], and one long-lasting phase covering the period 5200–3800 cal. BP, that corresponds to the permanency of the agropastoral practices in the littoral hills.
- Agriculture and animal husbandry were common practices in this littoral area 7000 years ago, a fact confirmed by the archeological record. Indeed, the littoral of the Nao Cap is, to date, the oldest evidence of Neolithic settlement and culture diffusion inland in the Iberian Peninsula.
- The gaps of agropastoral indicators were very consistent to the local- and regional-scale archeological record (i.e., hiatus of occupations at the Mid- to Late Neolithic transition and Final Bronze Age, and a near cessation of the occupations at the end of the Chalcolithic), underlining the importance of considering the socio-ecological systems as a whole.
- Breakdowns in the socio-ecological trajectory matched three short-term climate events at ~5.9, ~4.2, and ~2.8 kyr BP, known to have affected the North Atlantic, and likely expressed as an accentuation of the aridity in the Iberian Peninsula. Water scarcity was likely a key cause of land abandonment; however, this interpretation was supported for the 4.2 and 2.8 kyr BP events only.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Erlandson, J.M. The Archaeology of Aquatic Adaptations: Paradigms for a New Millennium. J. Archaeol. Res. 2001, 9, 287–350. [Google Scholar] [CrossRef]
- Manen, C. Spatial, chronological and cultural dynamics of the neolithization in the Western Mediterranean. In The Neolithic transition in the Mediterranean; Manen, C., Perrin, T., Guilaine, J., Eds.; Errance/AEP: Paris, France, 2014; pp. 405–418. [Google Scholar]
- García-Puchol, O.; Diez Castillo, A.A.; Pardo-Gordó, S. Timing the Western Mediterranean Last Hunter-Gatherers and First Farmers. In Times of Neolithic Transition along the Western Mediterranean, Fundamental Issues in Archaeology; García-Puchol, O., Salazar-García, D.C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 69–99. [Google Scholar] [CrossRef]
- Zapata, L.; Peña-Chocarro, L.; Pérez-Jordá, G.; Stika, H.-P. Early Neolithic Agriculture in the Iberian Peninsula. J. World Prehist. 2004, 18, 283–325. [Google Scholar] [CrossRef]
- Antolín, F.; Jacomet, S.; Buxó, R. The hard knock life. Archaeobotanical data on farming practices during the Neolithic (5400–2300 cal BC) in the NE of the Iberian Peninsula. J. Archaeol. Sci. 2015, 61, 90–104. [Google Scholar] [CrossRef]
- Roberts, C.N.; Woodbridge, J.; Palmisano, A.; Bevan, A.; Fyfe, R.; Shennan, S. Mediterranean landscape change during the Holocene: Synthesis, comparison and regional trends in population, land cover and climate. Holocene 2019, 29, 923–937. [Google Scholar] [CrossRef]
- Van de Noort, R. Conceptualising climate change archaeology. Antiquity 2011, 85, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, J.; Molina, L. La Cova de Les Cendres; Serie Mayor 6; Fundación MARQ: Alicante, Spain, 2009; p. 236. [Google Scholar]
- Bernabeu Aubán, J.; Jiménez-Puerto, J.; Escriba Ruiz, P.; Pardo-Gordó, S. C14 y Poblamiento en las Comarcas Centro-Meridionales del País Valenciano (c. 7000–1500 cal BC). Recer. Mus. D’Alcoi 2018, 27, 35–48. [Google Scholar]
- García-Puchol, O.; Aura Tortosa, J.E. El Abric de la Falguera (Alcoi, Alacant): 8.000 Años de Ocupación Humana en la Cabecera del Río de Alcoi; Museu arqueologic municipal d’Alcoi: Alcoy, Spain, 2006; p. 301. [Google Scholar]
- Bernabeu Aubán, J.; Manen, C.; Pardo-Gordó, S. Spatial and Temporal Diversity During the Neolithic Spread in the Western Mediterranean: The First Pottery Productions. In Times of Neolithic Transition along the Western Mediterranean, Fundamental Issues in Archaeology; García-Puchol, O., Salazar-García, D.C., Eds.; Springer Publishing: Berlin/Heidelberg, Germany, 2017; pp. 373–397. [Google Scholar] [CrossRef]
- Balsera, V.; Aubán, J.B.; Costa-Caramé, M.; Díaz-del-Río, P.; Sanjuán, L.G.; Pardo, S. The Radiocarbon Chronology of Southern Spain’s Late Prehistory (5600–1000 cal BC): A Comparative Review. Oxf. J. Archaeol. 2015, 34, 139–156. [Google Scholar] [CrossRef]
- Jiménez-Puerto, J.; Bernabeu Aubán, J.; Orozco, T. Habitat evolution in lberian Eastern façade, from Neolithic to the Bronze Age. In Reactive Proactive, Architecture; University Politècnica València: Valencia, Spain, 2018; pp. 138–143. [Google Scholar]
- Carrión, J.S.; Navarro, C. Cryptogam spores and other non-pollen microfossils as sources of palaeoecological information: Case-studies from Spain. Ann. Bot. Fenn. 2002, 39, 1–14. [Google Scholar]
- Pérez-Jordà, G.; Peña-Chocarro, L. Agricultural production between the 6th and the 3rd millennium cal BC in the central part of the Valencia region (Spain). In Barely Surviving or More than Enough? The Environmental Archaeology of Subsistence, Specialisation and Surplus Food Production; Groot, M., Lentjes, D., Zeiler, J., Eds.; Sidestone Press: Leiden, The Netherlands, 2013; pp. 81–100. [Google Scholar]
- Bernabeu Aubán, J.; Michael Barton, C.; Pardo Gordó, S.; Bergin, S.M. Modeling initial Neolithic dispersal. The first agricultural groups in West Mediterranean. Ecol. Model. 2015, 307, 22–31. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A.; Burjachs, F.; Giardini, M.; Kouli, K.; Masi, A.; Picornell-Gelabert, L.; Revelles, J.; Sadori, L.; Servera-Vives, G.; et al. From influence to impact: The multifunctional land use in Mediterranean prehistory emerging from palynology of archaeological sites (8.0–2.8 ka BP). Holocene 2019, 29, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Badal, E.; Bernabeu Aubán, J.; Vernet, J.L. Vegetation changes and human action from the Neolithic to the Bronze Age (7000–4000 B.P.) in Alicante, Spain, based on charcoal analysis. Veg. Hist. Archaeobot. 1994, 3, 155–166. [Google Scholar] [CrossRef]
- Bernabeu Aubán, J.; Badal, E. A view of the vegetation and economic exploitation of the forest in the Late Neolithic sites of Les Jovades and Niuet (Alicante, Spain). Bull. Soc. Bot. Fr. 1992, 139, 697–714. [Google Scholar] [CrossRef]
- Bernabeu Aubán, J.; Orozco-Köhler, T.; Diez Castillo, A.A.; Gómez Puche, M.; Molina Hernández, F.J. Mas d’Is (Penàguila, Alicante): Aldeas y recintos monumentales del Neolítico Inicial en el valle del Serpis. Trab. Prehist. 2003, 60, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S.; Fernández, S.; González-Sampériz, P.; Gil-Romera, G.; Badal, E.; Carrión-Marco, Y.; López-Merino, L.; López-Sáez, J.A.; Fierro, E.; Burjachs, F. Expected trends and surprises in the Lateglacial and Holocene vegetation history of the Iberian Peninsula and Balearic Islands. Rev. Palaeobot. Palynol. 2010, 162, 458–475. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S. Paleoflora y Paleovegetación de la Península Ibérica e Islas Baleares; Universidad de Murcia: Murcia, Spain, 2012; p. 994. [Google Scholar]
- Planchais, N.; Parra, I. Analyses polliniques de sédiments lagunaires et côtiers en Languedoc, en Roussillon et dans la province de Castellón (Espagne); Bioclimatologie. Bull. Soc. Bot. Fr. 1984, 131, 97–105. [Google Scholar] [CrossRef]
- Marco-Barba, J.; Burjachs, F.; Reed, J.M.; Santisteban, C.; Usera, J.M.; Alberola, C.; Expósito, I.; Guillem, J.; Patchett, F.; Vicente, E.; et al. Mid-Holocene and historical palaeoecology of the Albufera de València coastal lagoon. Limnetica 2019, 38, 353–389. [Google Scholar] [CrossRef]
- Dupré, M.; Fumanal, M.P.; Sanjaume, E.; Santisteban, C.; Usera, J.; Viñals, M.J. Quaternary evolution of the Pego coastal lagoon (Southern Valencia, Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 68, 291–299. [Google Scholar] [CrossRef]
- Viñals, M.J.; Belluomini, G.; Fumanal, M.P.; Dupré, M.; Usera, J.; Mestres, J.; Manfra, L. Rasgos paleoambientales holocenos en la Bahía de Xàbia (Alicante). In Estudios Sobre Cuatern; Fumanal, M.P., Bernabeu, J., Eds.; Universitat de Valencia: Valencia, Spain, 1993; pp. 107–114. [Google Scholar]
- Court-Picon, M.; Vella, C.; Chabal, L.; Bruneton, H. Paléo-environnements littoraux depuis 8000 ans sur la bordure occidentale du Golfe du Lion. Le lido de l’Etang de Thau (carottage SETIF, Sète, Hérault). Quaternaire 2010, 21, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Devillers, B.; Bony, G.; Degeai, J.-P.; Gascò, J.; Lachenal, T.; Bruneton, H.; Yung, F.; Oueslati, H.; Thierry, A. Holocene coastal environmental changes and human occupation of the lower Hérault River, southern France. Quat. Sci. Rev. 2019, 222, 105912. [Google Scholar] [CrossRef]
- Melis, R.T.; Di Rita, F.; French, C.; Marriner, N.; Montis, F.; Serreli, G.; Sulas, F.; Vacchi, M. 8000 years of coastal changes on a western Mediterranean island: A multiproxy approach from the Posada plain of Sardinia. Mar. Geol. 2018, 403, 93–108. [Google Scholar] [CrossRef]
- Poher, Y.; Ponel, P.; Médail, F.; Andrieu-Ponel, V.; Guiter, F. Holocene environmental history of a small Mediterranean island in response to sea-level changes, climate and human impact. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 465, 247–263. [Google Scholar] [CrossRef]
- Revelles, J.; Ghilardi, M.; Rossi, V.; Currás, A.; López-Bultó, O.; Brkojewitsch, G.; Vacchi, M. Coastal landscape evolution of Corsica island (W. Mediterranean): Palaeoenvironments, vegetation history and human impacts since the early Neolithic period. Quat. Sci. Rev. 2019, 225, 105993. [Google Scholar] [CrossRef]
- Ejarque, A.; Julià, R.; Reed, J.M.; Mesquita-Joanes, F.; Marco-Barba, J.; Riera, S. Coastal Evolution in a Mediterranean Microtidal Zone: Mid to Late Holocene Natural Dynamics and Human Management of the Castelló Lagoon, NE Spain. PLoS ONE 2016, 11, e0155446. [Google Scholar] [CrossRef] [Green Version]
- Brisset, E.; Burjachs, F.; Ballesteros Navarro, B.J.; de Pablo, J.F.L. Socio-ecological adaptation to Early-Holocene sea-level rise in the western Mediterranean. Glob. Planet. Chang. 2018, 169, 156–167. [Google Scholar] [CrossRef]
- Planchais, N. Palynologie lagunaire: L’exemple du Languedoc-Roussillon. Ann. Géogr. 1984, 93, 268–275. [Google Scholar]
- Fumanal, M.P.; Mateu, G.; Rey, J.; Somoza, L.; Viñals, M.J. Las unidades morfosedimentarias cuaternarias del litoral del Cap de la Nau (Valencia-Alicante) y su correlación con la plataforma continental. In Estudios Sobre Cuatern; Fumanal, M.P., Bernabeu, J., Eds.; Universitat de Valencia: Valencia, Spain, 1993; pp. 53–64. [Google Scholar]
- Anthony, E.J.; Marriner, N.; Morhange, C. Human influence and the changing geomorphology of Mediterranean deltas and coasts over the last 6000years: From progradation to destruction phase? Earth-Sci. Rev. 2014, 139, 336–361. [Google Scholar] [CrossRef]
- Fumanal, M.P.; Viñals, M.J.; Ferrer, C.; Aura, E.; Bernabeu, J.; Casabó, J.; Gisbert, J.; Sentí, M.A. Litoral y poblamiento en el litoral valenciano durante el Cuaternario reciente: Cap de Cullera-Puntal de Moraira. In Estudios Sobre Cuatern; Fumanal, M.P., Bernabeu, J., Eds.; Universitat de Valencia: Valencia, Spain, 1993; pp. 249–259. [Google Scholar]
- Viñals, M.J.; Fumanal, M.P. Quaternary development and evolution of the sedimentary environments in the Central Mediterranean Spanish coast. Quat. Int. 1995, 29, 119–128. [Google Scholar] [CrossRef]
- Carmona, P.; Ruiz, J.M. Historical morphogenesis of the Turia River coastal flood plain in the Mediterranean littoral of Spain. CATENA 2011, 86, 139–149. [Google Scholar] [CrossRef]
- Carmona, P.; Ruiz-Pérez, J.-M.; Blázquez, A.-M.; López-Belzunce, M.; Riera, S.; Orengo, H. Environmental evolution and mid–late Holocene climate events in the Valencia lagoon (Mediterranean coast of Spain). Holocene 2016, 26, 1750–1765. [Google Scholar] [CrossRef]
- Rey, J.; Fumanal, P.M. The Valencian coast (Western Mediterranean): Neotectonics and geomorphology. Quat. Sci. Rev. 1996, 15, 789–802. [Google Scholar] [CrossRef]
- Ballesteros Navarro, B.; Domínguez Sánchez, J.A.; Díaz-Losada, E.; García Menéndez, O. Zonas húmedas mediterráneas y acuíferos asociados. Condicionantes hidrogeológicos del Marjal de Pego-Oliva (Alicante-Valencia). Bol. Geo. Min. 2009, 120, 459–478. [Google Scholar]
- Cirujano, S.; Medina, L.; Peris, J.B.; Stübing, G. Estudio de la Flora y Vegetacion de las Marjales de Pego-Oliva y la Safor Orientado a su Gestion, Valencia, Spain. 1995, p. 284. Available online: http://www.humedalesibericos.com/sesiones/humedales1/web/publicaciones/sp_35.pdf (accessed on 3 July 2020).
- Rivas-Martínez, S.; Penas, Á.; Díaz González, T.E.; Cantó, P.; del Río, S.; Costa, J.C.; Herrero, L.; Molero, J. Biogeographic Units of the Iberian Peninsula and Baelaric Islands to District Level. A Concise Synopsis. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Plant and Vegetation; Springer Publishing: Berlin/Heidelberg, Germany, 2017; Volume 1, pp. 131–188. [Google Scholar] [CrossRef]
- Soriano, P.; Costa, M. The Coastal Levantine Area. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Plant and Vegetation; Springer Publishing: Berlin/Heidelberg, Germany, 2017; Volume 1, pp. 589–625. [Google Scholar]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Ramsey, C.B.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef] [Green Version]
- Blaauw, M. Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat. Geochronol. 2010, 5, 512–518. [Google Scholar] [CrossRef]
- Nowaczyk, N.R. Logging of Magnetic Susceptibility. In Tracking Environmental Change Using Lake Sediments: Basin Analysis, Coring, and Chronological Techniques, Developments in Paleoenvironmental Research; Last, W.M., Smol, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 155–170. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a fire proxy. In Tracking Environmental Change Using Lake Sediments: Terrestrial, Algal, and Siliceous Indicators; Last, W.M., Smol, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 75–97. [Google Scholar] [CrossRef]
- Goeury, C.; De Beaulieu, J.L. À propos de la concentration du pollen à l’aide de la liqueur de Thoulet dans les sédiments minéraux. Pollen Spores 1979, XXI, 239–251. [Google Scholar]
- Girard, M. Nouvelles techniques de préparation en palynologie appliqués à trois sédiments du Quaternaire final de l’Abri Cornille (Istres -Bouches du Rhône). Quaternaire 1969, 6, 275–278. [Google Scholar] [CrossRef]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de Botanique Historique et Palynologie Editions: Marseille, France, 1992; p. 543. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis; John Wiley & Sons: Chichester, UK, 1989; p. 328. [Google Scholar]
- Van Geel, B.; Buurman, J.; Brinkkemper, O.; Schelvis, J.; Aptroot, A.; van Reenen, G.; Hakbijl, T. Environmental reconstruction of a Roman Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J. Archaeol. Sci. 2003, 30, 873–883. [Google Scholar] [CrossRef]
- Revelles, J.; Burjachs, F.; van Geel, B. Pollen and non-pollen palynomorphs from the Early Neolithic settlement of La Draga (Girona, Spain). Rev. Palaeobot. Palynol. 2016, 225, 1–20. [Google Scholar] [CrossRef]
- Revelles, J.; van Geel, B. Human impact and ecological changes in lakeshore environments. The contribution of non-pollen palynomorphs in Lake Banyoles (NE Iberia). Rev. Palaeobot. Palynol. 2016, 232, 81–97. [Google Scholar] [CrossRef]
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia, Tilia-Graph and TGView; Illinois State Museum: Springfield, IL, USA, 1991; Available online: https://www.tiliait.com/ (accessed on 3 July 2020).
- Gelorini, V.; Verbeken, A.; van Geel, B.; Cocquyt, C.; Verschuren, D. Modern non-pollen palynomorphs from East African lake sediments. Rev. Palaeobot. Palynol. 2011, 164, 143–173. [Google Scholar] [CrossRef]
- Torres, T.; Ortiz, J.E.; Martín-Sánchez, D.; Arribas, I.; Moreno, L.; Ballesteros, B.; Blázquez, A.; Domínguez, J.A.; Estrella, T.R. The long Pleistocene record from the Pego-Oliva marshland (Alicante-Valencia, Spain). Geol. Soc. 2014, 388, 429–452. [Google Scholar] [CrossRef]
- Vacchi, M.; Ghilardi, M.; Melis, R.T.; Spada, G.; Giaime, M.; Marriner, N.; Lorscheid, T.; Morhange, C.; Burjachs, F.; Rovere, A. New relative sea-level insights into the isostatic history of the Western Mediterranean. Quat. Sci. Rev. 2018, 201, 396–408. [Google Scholar] [CrossRef]
- Carrión, J.S.; Fierro, E.; Ros, M.; Munuera, M.; Fernández, S.; Ochando, J.; Amorós, G.; Navarro, F.; Rodríguez-Estrella, T.; Manzano, S.; et al. Ancient Forests in European drylands: Holocene palaeoecological record of Mazarrón, south-eastern Spain. Proc. Geol. Asso. 2018, 129, 512–525. [Google Scholar] [CrossRef]
- Court-Picon, M.; Buttler, A.; de Beaulieu, J.-L. Modern pollen/vegetation/land-use relationships in mountain environments: An example from the Champsaur valley (French Alps). Veg. Hist. Archaeobot. 2006, 15, 151–168. [Google Scholar] [CrossRef]
- Deza-Araujo, M.; Morales-Molino, C.; Tinner, W.; Henne, P.D.; Heitz, C.; Pezzatti, G.B.; Hafner, A.; Conedera, M. A critical assessment of human-impact indices based on anthropogenic pollen indicators. Quat. Sci. Rev. 2020, 236, 106291. [Google Scholar] [CrossRef]
- Bernabeu Aubán, J.; Martí, B. The first agricultural groups in the Iberian Peninsula. In La Transition Néolithique en Méditerranée; Errance Publishing: Paris, France, 2014; pp. 419–438. [Google Scholar]
- Bernabeu Aubán, J.; Molina, L.L.; Esquembre-Bebiá, M.A.; Ortega, J.R.; Boronat, J. La cerámica impresa mediterránea en el origen del Neolítico de la península ibérica. In De Méditerranée et D’ailleurs. Mélanges Offerts à J. Guilaine; Les Archives d’Ecologie Préhistorique: Toulouse, France, 2009; pp. 83–95. [Google Scholar]
- Pantaléon-Cano, J.; Yll, E.-I.; Pérez-Obiol, R.; Roure, J.M. Palynological evidence for vegetational history in semi-arid areas of the western Mediterranean (Almería, Spain). Holocene 2003, 13, 109–119. [Google Scholar] [CrossRef]
- Carrión, J.S.; Fernández, S.; Jiménez-Moreno, G.; Fauquette, S.; Gil-Romera, G.; González-Sampériz, P.; Finlayson, C. The historical origins of aridity and vegetation degradation in southeastern Spain. J. Arid Environ. 2010, 74, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S.; Van Geel, B. Fine-resolution Upper Weichselian and Holocene palynological record from Navarrés (Valencia, Spain) and a discussion about factors of Mediterranean forest succession. Rev. Palaeobot. Palynol. 1999, 106, 209–236. [Google Scholar] [CrossRef]
- Carrión-Marco, Y. La Vegetación Mediterránea y Atlántica de la Península Ibérica: Nuevas Secuencias Antracológicas; Servicio de Investigación Prehistórica: Valencia, Spain, 2005; p. 314. [Google Scholar]
- López Sáez, J.A.; Van Geel, B.; Sánchez, M.M. Aplicación de los microfósiles no polínicos en Palinología Arqueológica. In Congresso de Arqueologia Peninsular; ADECAP: Vila Real, Portugal, 2000; pp. 11–20. [Google Scholar]
- Esquembre-Bebiá, M.A.; Soler, J.D.D.B.; Maestre, F.J.J.; Hernández, F.J.M.; Navas, A.L.; de Pablo, J.F.L.; Valle, R.M.; Eres, M.P.I.; García, C.F.; Pastor, R.R.; et al. El yacimiento neolítico del Barranquet de Oliva (Valencia). In IV Congreso del Neolítico Peninsular; Museo Arqueológico Alicante: Alicante, Spain, 2008; pp. 183–190. [Google Scholar]
- Bernabeu Aubán, J.; Molina, L.; Diez, A.; Orozco, T. Inequalities and Power. Three Millennia of Prehistory in Mediterranean Spain (5600—2000 cal BC). In Social Inequality in Iberian Late Prehistory; BAR International Series; Archaeopress: Oxford, UK, 2006; pp. 97–116. [Google Scholar]
- Miquel Calatayud, P.; Guitart Perarnau, I.; Martinez Valle, R.; Mata Parreño, C.; Pascual Benito, J.L. L’ocupació Prehistòrica de la Cova de Bolumini (Beniarbeig-Benimeli-Marina Alta); Actes del III Congres de Història de la Marina Alta; Institut d’Estudis Comarcals de la Marina Alta: Pedreguer, Spain, 1992; pp. 31–48. [Google Scholar]
- Alonso López, A.M. Las dinámicas de poblamiento ibérico y romano en la Vall d’Albaida, L’Alcoià y El Comtat (ss. IV a.C.-II d.C.): Un estudio comparativo del patrón de asentamiento. Recer. Mus. D’Alcoi 2018, 27, 63–78. [Google Scholar]
- García Atiénzar, G.; Barciela González, V. La Prehistòria a la Marina Alta (Alacant): De Les Primeres Poblacions Humanes al Final de L’Edat del Bronze. In VI Jornades d’Estudis Carmel Giner Bolufer de Pego i Les Valls Actes; De Pego, A., Ed.; Institut d’Estudis Comarcals de la Marina Alta: Pego, Spain, 2018; pp. 103–114. [Google Scholar]
- Grau Mira, I. El Poblamiento de Época Ibérica en la Region Centro-Meridional del País Valenciano. Ph.D. Thesis, Universidad de Alicante, Alicante, Spain, 2000; p. 524. [Google Scholar]
- Grau Mira, I. Les Valls de Pego en el Marc del Paisatge Ibèric de la Marina Alta (Segles vii-ii a.n.e.). In VI Jornades d’Estudis Carmel Giner Bolufer de Pego I Les Valls Actes; De Pego, A., Ed.; Institut d’Estudis Comarcal de la Marina Alta: Pego, Spain, 2018; pp. 111–128. [Google Scholar]
- Bond, G.; Kromer, B.; Beer, J.; Muscheler, R.; Evans, M.N.; Showers, W.; Hoffmann, S.; Lotti-Bond, R.; Hajdas, I.; Bonani, G. Persistent Solar Influence on North Atlantic Climate During the Holocene. Science 2001, 294, 2130–2136. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, J.; Climent, S. Sobre la pesca en la edad del bronce. ARSE 1985, 20, 481–485. [Google Scholar]
- Bernabeu Aubán, J.; García Puchol, O.; Barton, C.M.; McClure, S.; Pardo Gordó, S. Radiocarbon dates, climatic events, and social dynamics during the Early Neolithic in Mediterranean Iberia. Quat. Int. 2016, 403, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Schröder, T.; van’t Hoff, J.; López-Sáez, J.A.; Viehberg, F.; Melles, M.; Reicherter, K. Holocene climatic and environmental evolution on the southwestern Iberian Peninsula: A high-resolution multi-proxy study from Lake Medina (Cádiz, SW Spain). Quat. Sci. Rev. 2018, 198, 208–225. [Google Scholar] [CrossRef]
- Scott, L. Environmental implications and origin of microscopic Pseudoschizaea Thiergart and Frantz ex R. Potonié emend. in sediments. J. Biogeogr. 1992, 19, 349–354. [Google Scholar] [CrossRef]
- Connor, S.E.; Vannière, B.; Colombaroli, D.; Anderson, R.S.; Carrión, J.S.; Ejarque, A.; Romera, G.G.; González-Sampériz, P.; Hoefer, D.; Morales-Molino, C.; et al. Humans take control of fire-driven diversity changes in Mediterranean Iberia’s vegetation during the mid–late Holocene. Holocene 2019, 29, 886–901. [Google Scholar] [CrossRef]
- Rodriguez-Lloveras, X.; Machado, M.J.; Sanchez-Moya, Y.; Calle, M.; Medialdea, A.; Sopeña, A.; Benito, G. Impacts of sediment connectivity on Holocene alluvial records across a Mediterranean basin (Guadalentín River, SE-Spain). CATENA 2020, 187, 104321. [Google Scholar] [CrossRef]
- Badal, E. Estudio antracológico de la secuencia holocena de la Cova de les Cendres. In La Cova de Les Cendres: (Moraira-Teulada, Alicante); Museo Arqueológico de Alicante-MARQ: Alicante, Spain, 2009; pp. 125–134. [Google Scholar]
- Fyfe, R.M.; Woodbridge, J.; Palmisano, A.; Bevan, A.; Shennan, S.; Burjachs, F.; Legarra Herrero, B.; García Puchol, O.; Carrión, J.-S.; Revelles, J.; et al. Prehistoric palaeodemographics and regional land cover change in eastern Iberia. Holocene 2019, 29, 799–815. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu Auban, J.; Moreno Martín, A.; Barton, C.M. Complex systems, social networks and the evolution of social complexity. In The Prehistory of Iberia: Debating Early Social Stratification and the State; Berrocal, M., García Sanjuán, L., Gilman, A., Eds.; Routledge: New York, NY, USA, 2012; pp. 23–37. [Google Scholar]
- Knipper, C.; Rihuete-Herrada, C.; Voltas, J.; Held, P.; Lull, V.; Micó, R.; Risch, R.; Alt, K.W. Reconstructing Bronze Age diets and farming strategies at the early Bronze Age sites of La Bastida and Gatas (southeast Iberia) using stable isotope analysis. PLoS ONE 2020, 15, e0229398. [Google Scholar] [CrossRef]
- García-Puchol, O.; Bernabeu Aubán, J.; Carrión, M.; Molina Balaguer, Y.; Perez Jorda, G.; Gómez Puche, M. A funerary perspective on Bell Beaker period in the Western Mediterranean. Reading the social context of individual burials at La Vital (Gandía, Valencia). Trab. Prehist. 2013, 70, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu Aubán, J.; Pascual, J.L.; Orozco, T.; Badal, E.; Fumanal, M.P.; García Puchol, O. Niuet (L’Alqueria d’Asnar). Poblado del III Milenio aC; Museu d’Alcoi: Alcoy, Spain, 1994; pp. 9–74. [Google Scholar]
- Murillo-Barroso, M.; Martinón-Torres, M.; Massieu, M.D.C.; Socas, D.M.; González, F.M. Early metallurgy in SE Iberia. The workshop of Las Pilas (Mojácar, Almería, Spain). Archaeol. Anthropol. Sci. 2017, 9, 1539–1569. [Google Scholar] [CrossRef]
- Pérez Ripoll, M. La explotación ganadera durante el IIIr milenio aC en la Península Ibérica. In II Congres del Neolític a la Península Ibèrica. SAGUNTUM; Universitat de València: Valencia, Spain, 1999; Volume 2, pp. 95–103. [Google Scholar]
- Ruan, J.; Kherbouche, F.; Genty, D.; Blamart, D.; Cheng, H.; Dewilde, F.; Hachi, S.; Edwards, R.L.; Régnier, E.; Michelot, J.-L. Evidence of a prolonged drought ca. 4200 yr BP correlated with prehistoric settlement abandonment from the Gueldaman GLD1 Cave, Northern Algeria. Clim. Past 2016, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bini, M.; Zanchetta, G.; Perşoiu, A.; Cartier, R.; Català, A.; Cacho, I.; Dean, J.R.; Rita, F.D.; Drysdale, R.N.; Finnè, M.; et al. The 4.2 ka BP Event in the Mediterranean region. Clim. Past 2019, 15, 555–577. [Google Scholar] [CrossRef] [Green Version]
- Cartier, R.; Brisset, E.; Guiter, F.; Sylvestre, F.; Tachikawa, K.; Anthony, E.J.; Paillès, C.; Bruneton, H.; Bard, E.; Miramont, C. Multiproxy analyses of Lake Allos reveal synchronicity and divergence in geosystem dynamics during the Lateglacial/Holocene in the Alps. Quat. Sci. Rev. 2018, 186, 60–77. [Google Scholar] [CrossRef]
- Cartier, R.; Sylvestre, F.; Paillès, C.; Sonzogni, C.; Couapel, M.; Alexandre, A.; Mazur, J.-C.; Brisset, E.; Miramont, C.; Guiter, F. Diatom-oxygen isotope record from high-altitude Lake Petit (2200 m a.s.l.) in the Mediterranean Alps: Shedding light on a climatic pulse at 4.2 ka. Clim. Past 2019, 15, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Brisset, E.; Miramont, C.; Anthony, E.J.; Bruneton, H.; Rosique, T.; Sivan, O. Sediment budget quantification of a sub-Alpine river catchment since the end of the last glaciation. CATENA 2014, 114, 169–179. [Google Scholar] [CrossRef]
- Brisset, E.; Miramont, C.; Guiter, F.; Anthony, E.J.; Tachikawa, K.; Poulenard, J.; Arnaud, F.; Delhon, C.; Meunier, J.-D.; Bard, E.; et al. Non-reversible geosystem destabilisation at 4200 cal. BP: Sedimentological, geochemical and botanical markers of soil erosion recorded in a Mediterranean alpine lake. Holocene 2013, 23, 1863–1874. [Google Scholar] [CrossRef]
- Lull, V.; Micó, R.; Rihuete, C.; Risch, R. Límites históricos y limitaciones del conocimiento arqueológico: La transición entre los grupos arqueológicos de Los Millares y El Argar. In Arqueología, Sociedad, Territorio y Paisaje, Homenaje a D. Fernández Posse; Bueno Ramírez, P., Ed.; Consejo Superior de Investigaciones Científicas, CSIC, Instituto de Historia: Madrid, Spain, 2010; pp. 75–94. Available online: https://www.torrossa.com/digital/fcov/2010/2471828_FCOV.pdf#page=73 (accessed on 3 July 2020).
- Carozza, L.; Berger, J.F.; Burens-Carozza, A.; Marcigny, C. Society and environment in Southern France from the 3rd millennium BC to the beginning of the 2nd millennium BC: 2200 BC a tipping point? In Proceedings of the 7th Archaeological Conference of Central Germany, Halle, Germany, 23–26 October 2014; pp. 335–362. Available online: https://halshs.archives-ouvertes.fr/halshs-01245488/ (accessed on 3 July 2020).
- Carrión, J.S.; Andrade, A.; Bennett, K.D.; Navarro, C.; Munuera, M. Crossing forest thresholds: Inertia and collapse in a Holocene sequence from south-central Spain. Holocene 2001, 11, 635–653. [Google Scholar] [CrossRef]
- Burjachs, F.; Jones, S.E.; Giralt, S.; de Pablo, J.F.L. Lateglacial to Early Holocene recursive aridity events in the SE Mediterranean Iberian Peninsula: The Salines playa lake case study. Quat. Int. 2016, 403, 187–200. [Google Scholar] [CrossRef]
- Pérez Jordà, G.; Alonso Martínez, N.; Iborra Eres, M.P. Agricultura y ganadería protohistóricas en la Península Ibérica: Modelos de gestión. In Arqueología de la Tierra: Paisajes Rurales de la Protohistoria Peninsular: VI Cursos de Verano Internacionales de la Universidad de Extremadura (Castuera, 5–8 de Julio de 2005); Universidad de Extremadura, Servicio de Publicaciones: Cáceres, Spain, 2007; pp. 327–372. [Google Scholar]
- Pérez Jordà, G. La Agricultura en el País Valenciano Entre el VI y el I Milenio a.C. Ph.D. Thesis, Universitat de València, Valencia, Spain, 2013; p. 374. [Google Scholar]
- Gisbert Santonja, J.A. El Tossalet de les Mondes (Pego); Univ. de València, Saguntum: Valencia, Spain, 1993; pp. 207–231. [Google Scholar]
- Aranegui Gascó, C. El Sucronenesis Sinus en Época Ibérica. Saguntum Extra; Univ. de València, Saguntum: Valencia, Spain, 2015; p. 201. [Google Scholar]
- Barton, C.M. Complexity, social complexity, and modeling. J. Archaeol. Method Theory 2014, 21, 306–324. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core Name | Core Depth (cm) | Material | Laboratory Code | Age 14C BP | Min Age cal. BP | Max Age cal. BP |
|---|---|---|---|---|---|---|
| P17-9 | 37.5 | charcoals | Poz-109495 | 1980 ± 30 | 1876 | 1992 |
| P17-9 | 107.5 | charcoals | Poz-109496 | 2410 ± 30 | 2351 | 2684 |
| P17-9 | 154.5 | charcoals | Poz-94011 | 3065 ± 35 | 3180 | 3363 |
| P17-9 | 187.5 | charcoals, peat | Poz-95362 | 3365 ± 35 | 3485 | 3694 |
| P17-9 | 227.5 | charcoals | Poz-109439 | 4070 ± 35 | 4438 | 4804 |
| P17-9 | 251.5 | charcoals | Poz-94012 | 4675 ± 35 | 5316 | 5572 |
| P17-9 | 327.5 | charcoals | Poz-95364 | 5240 ± 40 | 5918 | 6177 |
| P17-9 | 357.5 | charcoals | Poz-95020 | 5810 ± 40 | 6498 | 6718 |
| P17-9 | 397.5 | charcoals | Poz-95364 | 7240 ± 40 | 7979 | 8161 |
| P17-9 | 423.5 | charcoals | Poz-95365 | 6040 ± 40 | 6786 | 6993 |
| P17-8 | 550.0 | charcoals | Beta-460429 | 6420 ± 30 | 7280 | 7421 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brisset, E.; Revelles, J.; Expósito, I.; Bernabeu Aubán, J.; Burjachs, F. Socio-Ecological Contingencies with Climate Changes over the Prehistory in the Mediterranean Iberia. Quaternary 2020, 3, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030019
Brisset E, Revelles J, Expósito I, Bernabeu Aubán J, Burjachs F. Socio-Ecological Contingencies with Climate Changes over the Prehistory in the Mediterranean Iberia. Quaternary. 2020; 3(3):19. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030019
Chicago/Turabian StyleBrisset, Elodie, Jordi Revelles, Isabel Expósito, Joan Bernabeu Aubán, and Francesc Burjachs. 2020. "Socio-Ecological Contingencies with Climate Changes over the Prehistory in the Mediterranean Iberia" Quaternary 3, no. 3: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030019