
Dating of the Lower Pleistocene Vertebrate Site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, Magnetostratigraphy, and Cosmogenic Radionuclides

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
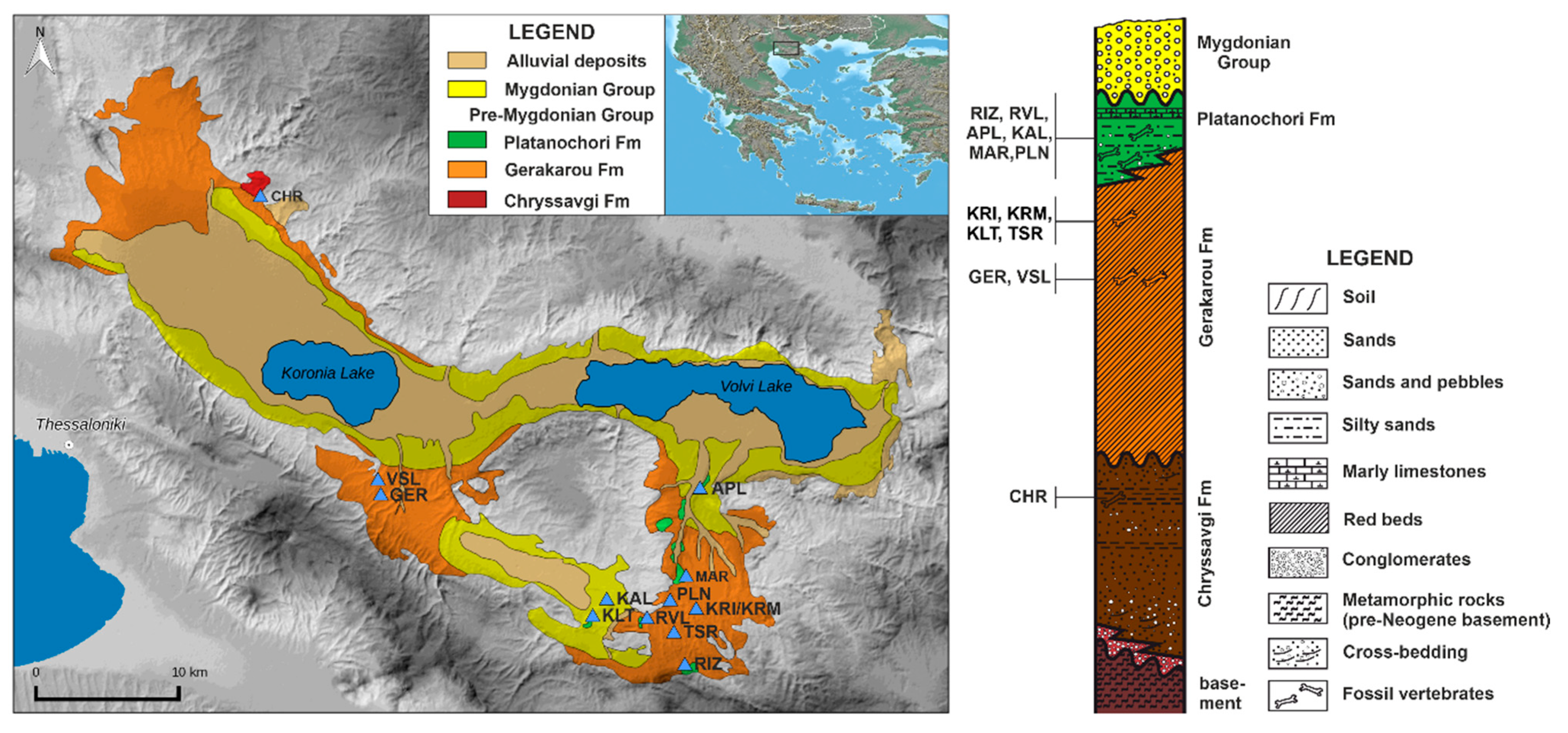
2. Geological Setting and Fossiliferous Locality
3. Methodology
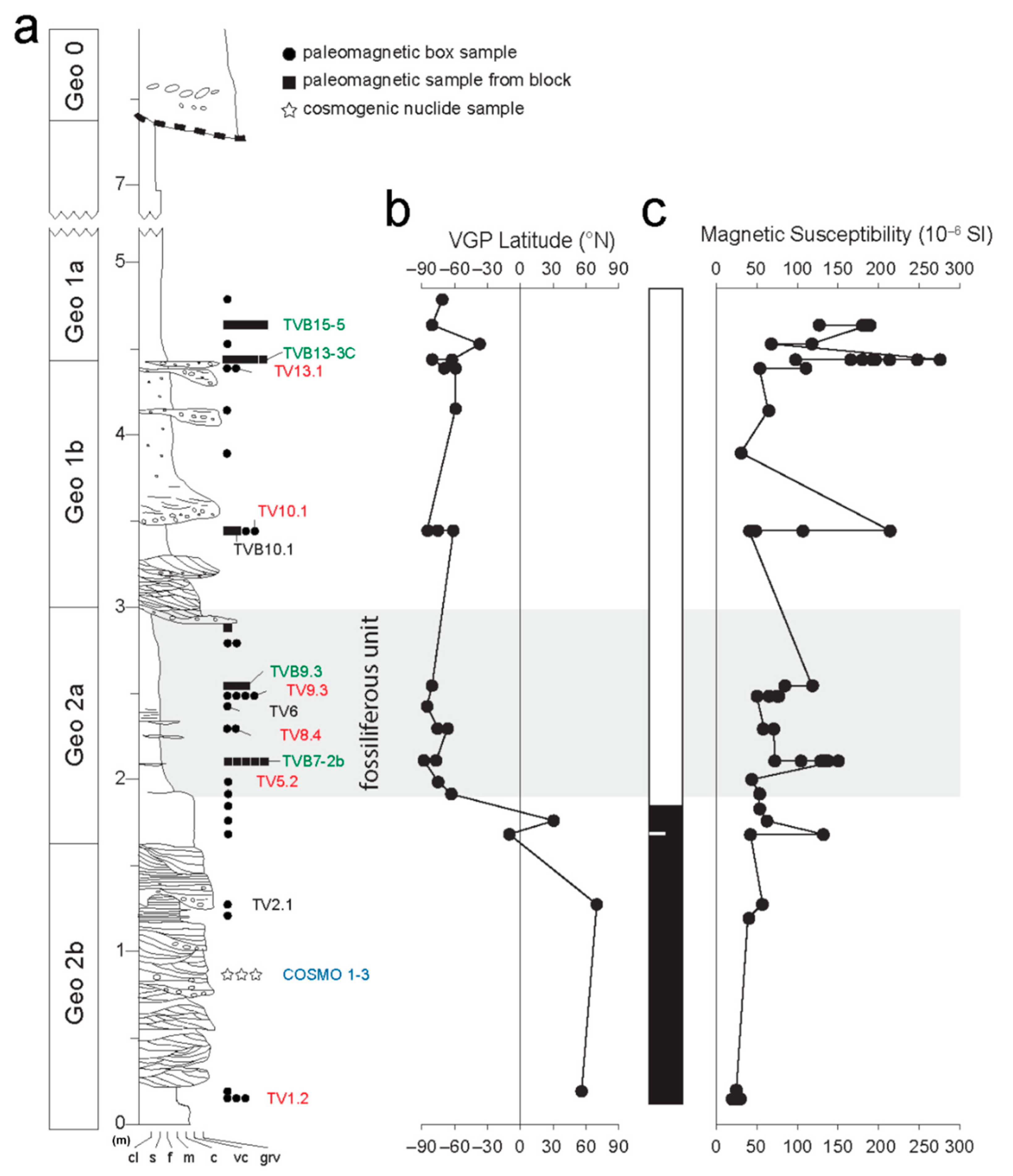
4. Results
4.1. Cosmogenic Radionuclides
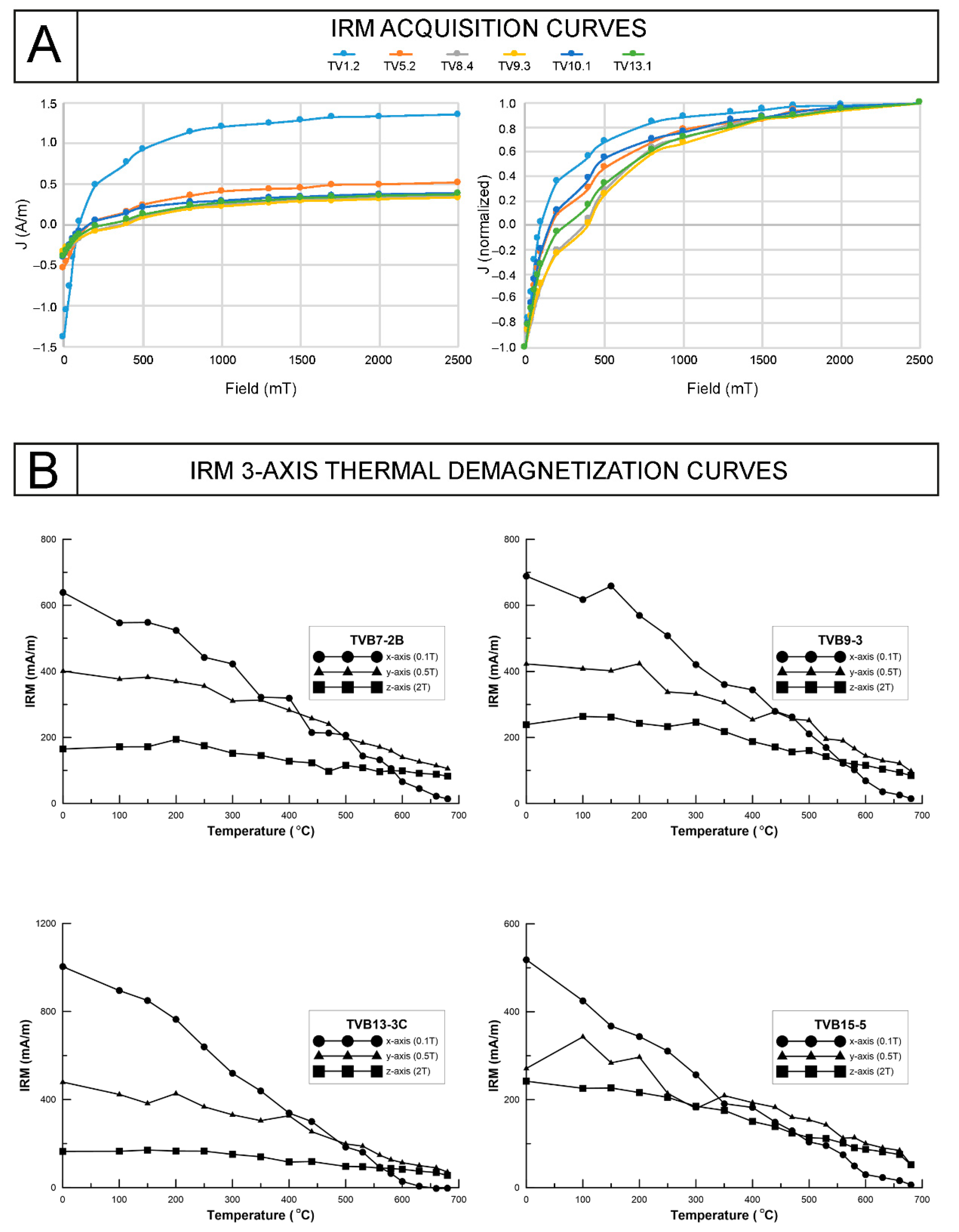
4.2. Magnetic Properties
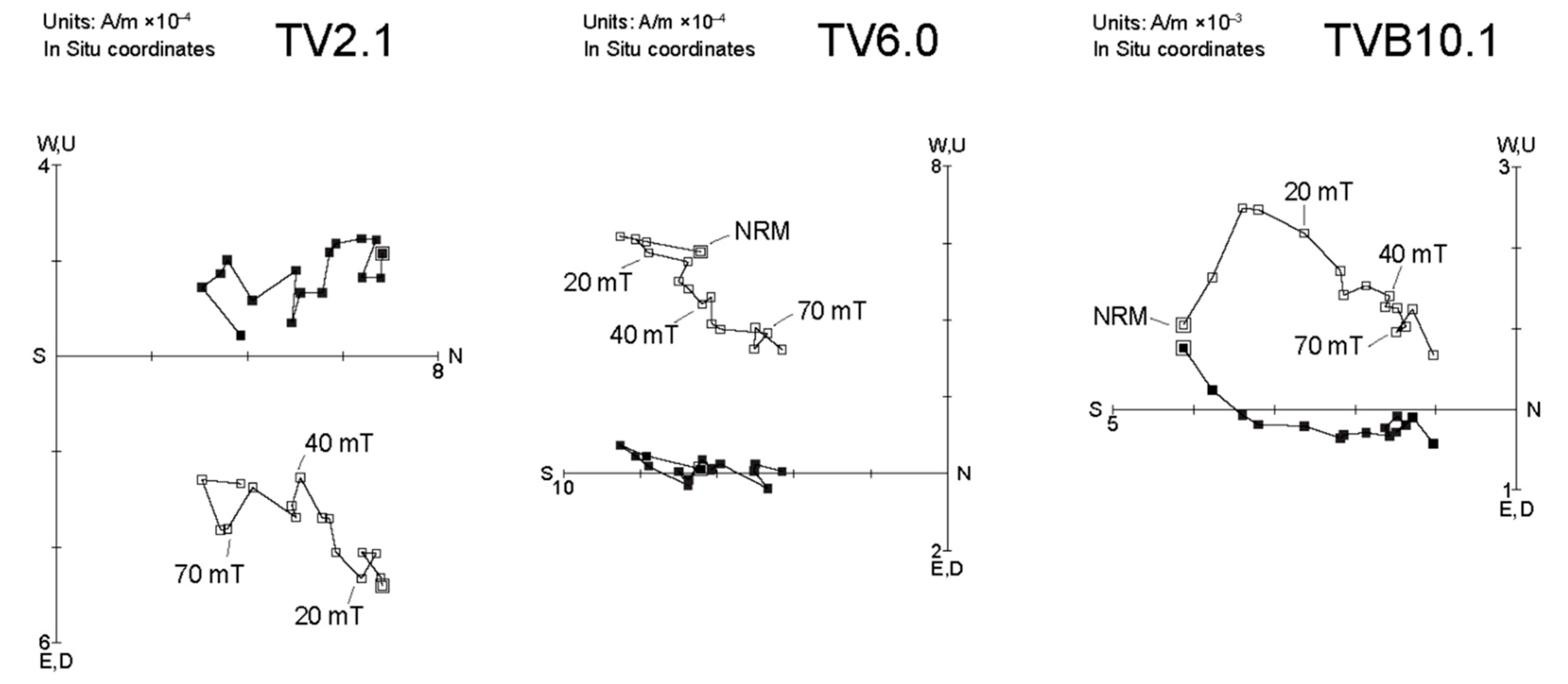
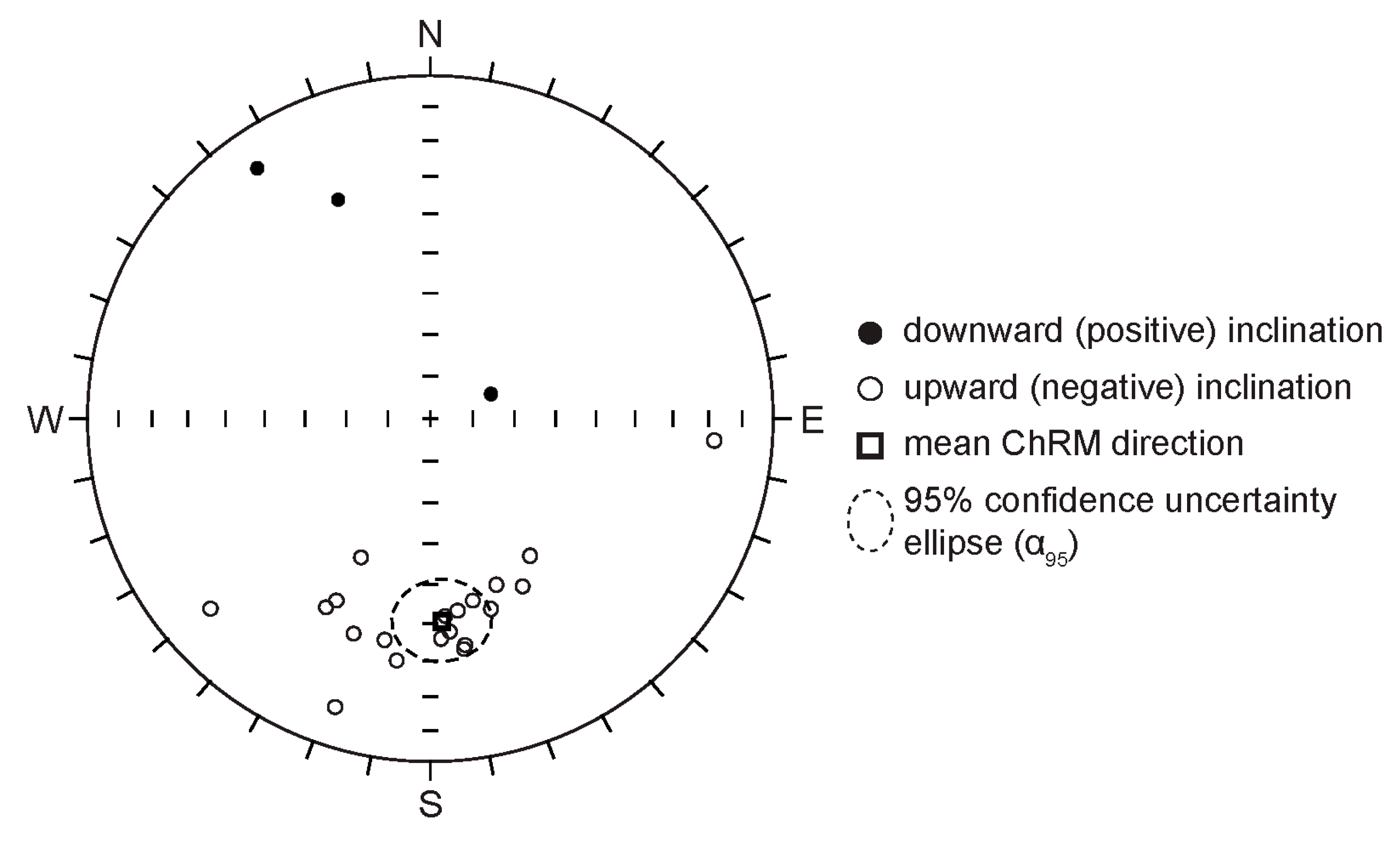
4.3. Magnetostratigraphy
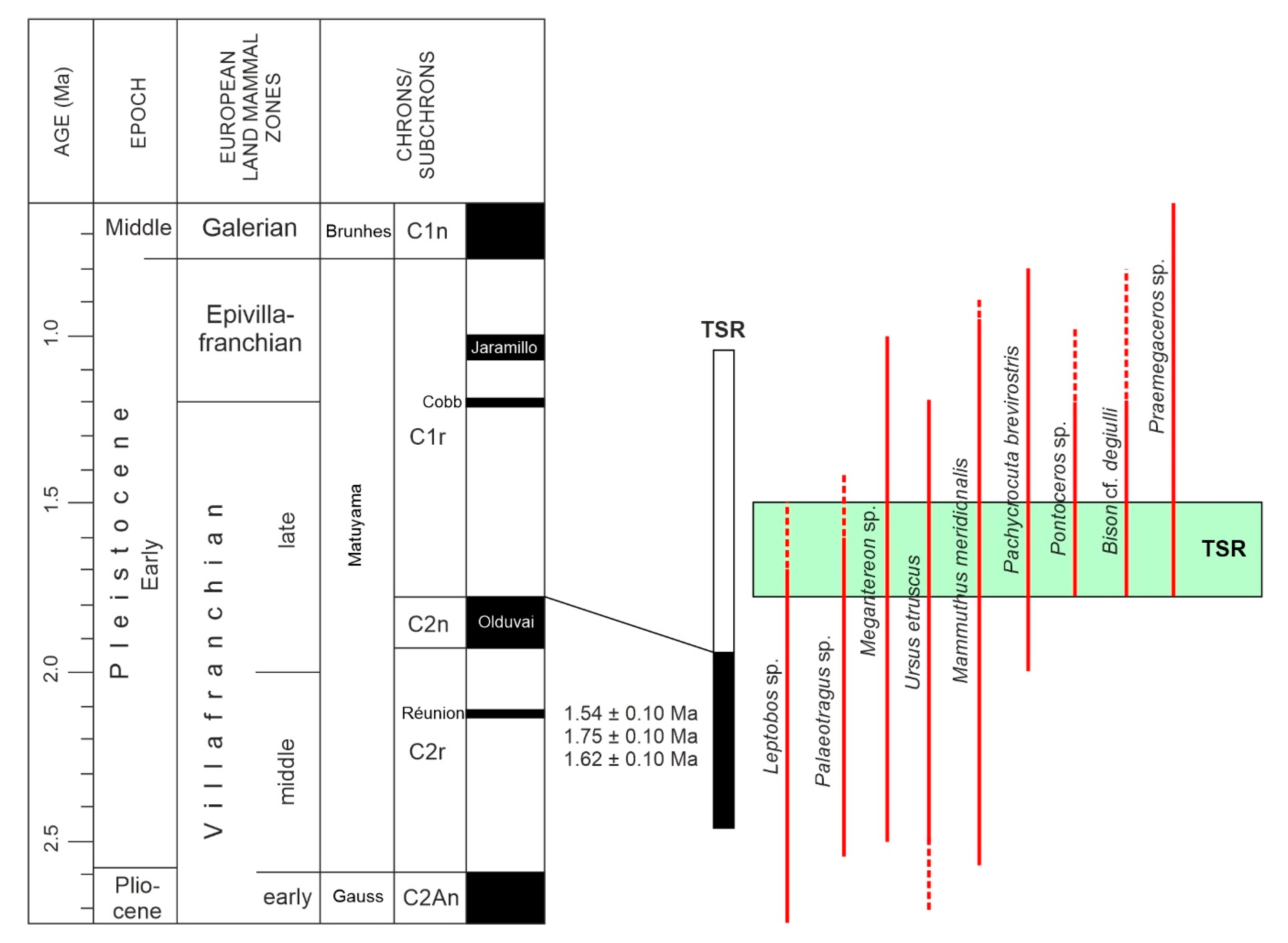
4.4. Biochronology
5. Discussion
5.1. Age Estimation: Age Constraints
5.2. Placement of TSR within the Greek and European Context
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rook, L.; Martínez-Navarro, B. Villafranchian: The long story of a Plio-Pleistocene European large mammal biochronologic unit. Quat. Int. 2010, 219, 134–144. [Google Scholar] [CrossRef]
- Martínez-Navarro, B. Early Pleistocene faunas of Eurasia and hominin dispersals. In Out of Africa I. The first hominin colonization of Eurasia; Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., Leakey, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 207–224. [Google Scholar]
- Napoleone, G.; Albianelli, A.; Azzaroli, A.; Bertini, A.; Magi, M.; Mazzini, I. Calibration of the upper Valdarno basin to the Plio-Pleistocene for correlating the Apennine continental sequences. Il Quaternario 2003, 16, 131–166. [Google Scholar]
- Kahlke, R.D. Late Early Pleistocene European large mammals and the concept of an Epivillafranchian biochron. In Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations. Vol. II. Proceedings of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar); Kahlke, R.D., Maul, L.C., Mazza, P., Eds.; Courier Forschungsinstitut Senckenberg: Frankfurt a. M., Germany, 2007; Volume 259, pp. 265–278. [Google Scholar]
- Palombo, M.R. Large mammals faunal dynamics in Southwestern Europe during the late Early Pleistocene: Implications for the biochronological assessment and correlation of mammalian faunas. Alpine Mediterr. Quat. 2016, 29, 143–168. [Google Scholar]
- Bellucci, L.; Sardella, R.; Rook, L. Large mammal biochronology framework in Europe at Jaramillo: The Epivillafranchian as a formal biochron. Quat. Int. 2015, 389, 84–89. [Google Scholar] [CrossRef]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Leroy, S.A.G.; Arpe, K.; Mikolajewicz, U. Vegetation context and climatic limits of the Early Pleistocene hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1448–1463. [Google Scholar] [CrossRef] [Green Version]
- Palombo, M.R. Deconstructing mammal dispersals and faunal dynamics in SW Europe during the Quaternary. Quat. Sci. Rev. 2014, 96, 50–71. [Google Scholar] [CrossRef]
- Ferring, R.; Oms, O.; Agustí, J.; Berna, F.; Nioradze, M.; Shelia, T.; Tappen, M.; Vekua, A.; Zhvania, D.; Lordkipanidze, D. Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. Proc. Natl. Acad. Sci. USA 2011, 108, 10432–10436. [Google Scholar] [CrossRef] [Green Version]
- Lordkipanidze, D.; Jashashvili, T.; Vekua, A.; de Leon, M.S.P.; Zollikofer, C.P.E.; Rightmire, G.P.; Pontzer, H.; Ferring, R.; Oms, O.; Tappen, M.; et al. Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 2007, 449, 305–310. [Google Scholar] [CrossRef]
- Palombo, M.R. Faunal dynamics in SW Europe during the late Early Pleistocene: Palaeobiogeographical insights and biochronological issues. In European Early Pleistocene Biogeography and Ecology Based on the Mammal Record: Case Studies and Preliminary Syntheses; Kostopoulos, D.S., Konidaris, G., Tesakov, A., van den Hoek Ostende, L.W., Rook, L., Eds.; Comptes Rendus Palevol: Amsterdam, The Netherlands, 2018; Volume 17, pp. 247–261. [Google Scholar]
- Muttoni, G.; Scardia, G.; Kent, D.V. Early hominins in Europe: The Galerian migration hypothesis. Quat. Sci. Rev. 2018, 180, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Palombo, M.R. Biochronology of terrestrial mammals and Quaternary subdivisions: A case study of large mammals from the Italian peninsula. Il Quaternario 2009, 22, 291–306. [Google Scholar]
- Palombo, M.R. Discrete dispersal bioevents of large mammals in Southern Europe in the post-Olduvai Early Pleistocene: A critical overview. Quat. Int. 2017, 431, 3–19. [Google Scholar] [CrossRef]
- Sakellariou-Mane, E.; Psilovikos, A.; Koufos, G. Contribution to the study of Villafranchian in Northern Chalkidiki (Macedonia, Greece). Sci. Annals, Fac. Phys. Mathem., Univ. Thessaloniki 1979, 19, 279–293. [Google Scholar]
- Zamanis, A.; Faugères, L.; Bonis, L.d.; Fountoulis, D.; Simeakis, C.; Panayotis, I.; Dimitrakopoulos, R.; Mercier, J.L. Découverte d’une faune de Mammifères du Quaternaire ancien dans les formations du lac Langhada (Macédoine centrale, Grèce). Implications néotectoniques. C. R. Acad. Sci. Paris 1980, 291, 813–816. [Google Scholar]
- Koufos, G.D.; Melentis, J.K. New data from the Villafranchian mammal locality of Gerakarou (Macedonia-Greece). Prak. Akad. Ath. 1983, 58, 185–191. [Google Scholar]
- Koufos, G.D.; Syrides, G.E.; Kostopoulos, D.S.; Koliadimou, K.K. Preliminary results about the stratigraphy and the palaeoenvironment of Mygdonia Basin, Macedonia, Greece. Geobios 1995, M.S. 18, 243–249. [Google Scholar] [CrossRef]
- Tsoukala, E.; Chatzopoulou, K. A new Early Pleistocene (latest Villafranchian) site with mammals in Kalamotó (Mygdonia Basin, Macedonia, Greece) – Preliminary report. Mitt. Komm. Quartärforsch. Österr. Akad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Konidaris, G.E.; Tourloukis, V.; Kostopoulos, D.S.; Thompson, N.; Giusti, D.; Michailidis, D.; Koufos, G.D.; Harvati, K. Two new vertebrate localities from the Early Pleistocene of Mygdonia Basin (Macedonia, Greece): Preliminary results. C. R. Palevol 2015, 14, 353–362. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S. The Plio-Pleistocene large mammal record of Greece: Implications for early human dispersals into Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 269–280. [Google Scholar]
- Kostopoulos, D.S.; Vassiliadou, K.; Koufos, G.D. The beginning of Pleistocene in the Balkan area according to the mammal record; palaeozoogeographical approach. Ann. Géol. Pays Hellén. 2002, 39, 253–278. [Google Scholar]
- Spassov, N. Southeastern Europe as a route for the earliest dispersal of Homo toward Europe: Ecological conditions and the timing of the first human occupation of Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 281–290. [Google Scholar]
- Tourloukis, V.; Harvati, K. The Palaeolithic record of Greece: A synthesis of the evidence and a research agenda for the future. Quat. Int. 2018, 66, 48–65. [Google Scholar] [CrossRef]
- Harvati, K.; Panagopoulou, E.; Runnels, C. The paleoanthropology of Greece. Evol. Anthropol. 2009, 18, 131–143. [Google Scholar] [CrossRef]
- Harvati, K.; Röding, C.; Bosman, A.M.; Karakostis, F.A.; Grün, R.; Stringer, C.; Karkanas, P.; Thompson, N.C.; Koutoulidis, V.; Moulopoulos, L.A.; et al. Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature 2019, 571, 500–504. [Google Scholar] [CrossRef]
- Athanassiou, A. Pleistocene vertebrates from the Kyparíssia lignite mine, Megalopolis Basin, S. Greece: Rodentia, Carnivora, Proboscidea, Perissodactyla, Ruminantia. Quat. Int. 2018, 497, 198–221. [Google Scholar] [CrossRef]
- Athanassiou, A.; Michailidis, D.; Vlachos, E.; Tourloukis, V.; Thompson, N.; Harvati, K. Pleistocene vertebrates from the Kyparíssia lignite mine, Megalopolis Basin, S. Greece: Testudines, Aves, Suiformes. Quat. Int. 2018, 497, 178–197. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Harvati, K. The skeleton of a straight-tusked elephant (Palaeoloxodon antiquus) and other large mammals from the Middle Pleistocene butchering locality Marathousa 1 (Megalopolis Basin, Greece): Preliminary results. Quat. Int. 2018, 497, 65–84. [Google Scholar] [CrossRef]
- Benammi, M.; Aidona, E.; Merceron, G.; Koufos, G.D.; Kostopoulos, D.S. Magnetostratigraphy and chronology of the Lower Pleistocene primate bearing Dafnero fossil site, N. Greece. Quaternary 2020, 3, 22. [Google Scholar] [CrossRef]
- Psilovikos, A.A. Palaeogeographic development of the basin and lake of Mygdonia (Langada-Volvi area), Greece. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1977. [Google Scholar]
- Giusti, D.; Konidaris, G.E.; Tourloukis, V.; Marini, M.; Maron, M.; Zerboni, A.; Thompson, N.; Koufos, G.D.; Kostopoulos, D.S.; Harvati, K. Recursive anisotropy: A spatial taphonomic study of the Early Pleistocene vertebrate assemblage of Tsiotra Vryssi, Mygdonia Basin, Greece. Boreas 2019, 48, 713–730. [Google Scholar]
- Konidaris, G.E.; Kostopoulos, D.S.; Koufos, G.D.; Tourloukis, V.; Harvati, K. Tsiotra Vryssi: A new vertebrate locality from the Early Pleistocene of Mygdonia Basin (Macedonia, Greece). In Proceedings of the XIV Annual Meeting of the European Association of Vertebrate Palaeontologists, Haarlem, The Netherlands, 6–10 July 2016; p. 37. [Google Scholar]
- Kostopoulos, D.S.; Maniakas, I.; Tsoukala, E. Early bison remains from Mygdonia Basin (Northern Greece). Geodiversitas 2018, 40, 283–319. [Google Scholar] [CrossRef] [Green Version]
- Koufos, G.D.; Konidaris, G.E.; Harvati, K. Revisiting Ursus etruscus (Carnivora, Mammalia) from the Early Pleistocene of Greece with description of new material. Quat. Int. 2018, 497, 222–239. [Google Scholar]
- Granger, D.E.; Kirchner, J.W.; Finkel, R.C. Quaternary downcutting rate of the New River, Virginia, measured from differential decay of cosmogenic 26Al and 10Be in cave-deposited alluvium. Geology 1997, 25, 107–110. [Google Scholar] [CrossRef]
- Granger, D.E.; Muzikar, P.F. Dating sediment burial with in situ-produced cosmogenic nuclides: Theory, techniques, and limitations. Earth Planet. Sci. Lett. 2001, 188, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Granger, D.E. Cosmogenic nuclide burial dating in archaeology and paleoanthropology. In Treatise on Geochemistry, 2nd ed.; Holland, H.D., Turekian, K.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 81–97. [Google Scholar]
- Schaller, M.; Ehlers, T.A.; Stor, T.; Torrent, J.; Lobato, L.; Christl, M.; Vockenhuber, C. Timing of European fluvial terrace formation and incision rates constrained by cosmogenic nuclide dating. Earth Planet. Sci. Lett. 2016, 451, 221–231. [Google Scholar] [CrossRef]
- Nishiizumi, K. Preparation of 26Al AMS standards. Nucl. Instrum. Methods Phys. Res. B 2004, 223–224, 388–392. [Google Scholar] [CrossRef]
- Nishiizumi, K.; Imamura, M.; Caffee, M.W.; Southon, J.R.; Finkel, R.C.; McAninch, J. Absolute calibration of 10Be AMS standards. Nucl. Instrum. Methods Phys. Res. B 2007, 258, 403–413. [Google Scholar] [CrossRef]
- Norris, T.L.; Gancarz, A.J.; Rokop, D.J.; Thomas, K.W. Half-life of 26Al. J. Geophys. Res. Solid Earth 1983, 88, B331–B333. [Google Scholar] [CrossRef]
- Chmeleff, J.; von Blanckenburg, F.; Kossert, K.; Jakob, D. Determination of the 10Be half-life by multicollector ICP-MS and liquid scintillation counting. Nucl. Instrum. Methods Phys. Res. B 2010, 268, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Korschinek, G.; Bergmaier, A.; Faestermann, T.; Gerstmann, U.C.; Knie, K.; Rugel, G.; Wallner, A.; Dillmann, I.; Dollinger, G.; von Gostomski, C.L.; et al. A new value for the half-life of 10Be by Heavy-Ion Elastic Recoil Detection and liquid scintillation counting. Nucl. Instrum. Methods Phys. Res. B 2010, 268, 187–191. [Google Scholar] [CrossRef]
- Braucher, R.; Merchel, S.; Borgomano, J.; Bourlès, D.L. Production of cosmogenic radionuclides at great depth: A multi element approach. Earth Planet. Sci. Lett. 2011, 309, 1–9. [Google Scholar] [CrossRef]
- Borchers, B.; Marrero, S.; Balco, G.; Caffee, M.; Goehring, B.; Lifton, N.; Nishiizumi, K.; Phillips, F.; Schaefer, J.; Stone, J. Geological calibration of spallation production rates in the CRONUS-Earth project. Quat. Geochronol. 2016, 31, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Marrero, S.M.; Phillips, F.M.; Borchers, B.; Lifton, N.; Aumer, R.; Balco, G. Cosmogenic nuclide systematics and the CRONUScalc program. Quat. Geochronol. 2016, 31, 160–187. [Google Scholar] [CrossRef] [Green Version]
- Lifton, N.; Sato, T.; Dunai, T.J. Scaling in situ cosmogenic nuclide production rates using analytical approximations to atmospheric cosmic-ray fluxes. Earth Planet. Sci. Lett. 2014, 386, 149–160. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T. Paleontological data analysis; Blackwell Publishing: Oxford, UK, 2006; p. 351. [Google Scholar]
- Arribas, A.; Garrido, G.; Viseras, C.; Soria, J.M.; Pla, S.; Solano, J.G.; Garcés, M.; Beamud, E.; Carrión, J.S. A mammalian lost world in Southwest Europe during the Late Pliocene. PLoS ONE 2009, 4, e7127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlke, R.D. Die Unterpleistozäne Komplexfundstelle Untermaßfeld-Zusammenfassung des Kenntnisstandes sowie synthetische Betrachtungen zu Genesemodell, Paläoökologie und Stratigraphie. In Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen), Teil 3; Kahlke, R.D., Ed.; Monographien des Römisch-Germanischen Zentralmuseums Mainz, Dr. Rudolf Habelt: Bonn, Germany, 2001; pp. 931–1030. [Google Scholar]
- Kahlke, R.D. (Ed.) The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany). Part 4; Monographien des Römisch-Germanischen Zentralmuseums Mainz: Mainz, Germany, 2020; Volume 40. [Google Scholar]
- Koufos, G.D. The Villafranchian mammalian faunas and biochronology of Greece. Boll. Soc. Paleontol. Ital. 2001, 40, 217–223. [Google Scholar]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe. Rev. de Paléobiologie 2003, 22, 197–229. [Google Scholar]
- Mazza, P.P.A.; Bertini, A.; Magi, M. The Late Pliocene site of Poggio Rosso (Central Italy): Taphonomy and paleoenvironment. Palaios 2004, 19, 227–248. [Google Scholar] [CrossRef]
- Pavia, M.; Zunino, M.; Coltorti, M.; Angelone, C.; Arzarello, M.; Bagnus, C.; Bellucci, L.; Colombero, S.; Marcolini, F.; Peretto, C.; et al. Stratigraphical and palaeontological data from the Early Pleistocene Pirro 10 site of Pirro Nord (Puglia, south eastern Italy). Quat. Int. 2012, 267, 40–55. [Google Scholar] [CrossRef]
- Huguet, R.; Saladié, P.; Cáceres, I.; Díez, C.; Rosell, J.; Bennàsar, M.; Blasco, R.; Esteban-Nadal, M.; Gabucio, M.J.; Rodríguez-Hidalgo, A.; et al. Successful subsistence strategies of the first humans in south-western Europe. Quat. Int. 2013, 295, 168–182. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Minwer-Barakat, R.; Alba, D.M.; Garcés, M.; Gómez, M.; Aurell-Garrido, J.; Ros-Montoya, S.; Moyà-Solà, S.; Berástegui, X. The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 2010, 29, 3972–3982. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Alba, D.M.; Aurell-Garrido, J. Villafranchian large mammals from the Iberian Peninsula: Paleobiogeography, paleoecology and dispersal events. J. Iber. Geol. 2014, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Medin, T.; Martínez-Navarro, B.; Rivals, F.; Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Figueirido, B.; Rook, L.; Palmqvist, P. Late Villafranchian Ursus etruscus and other large carnivorans from the Orce sites (Guadix-Baza basin, Andalusia, southern Spain): Taxonomy, biochronology, paleobiology, and ecogeographical context. Quat. Int. 2017, 431, 20–41. [Google Scholar] [CrossRef]
- Vislobokova, I.A.; Agadjanyan, A.K. New data on age of the Pleistocene fauna from the Trlica locality (Montenegro, Central Balkans) and its correlation with other faunas of Europe. Stratigr. Geol. Correl. 2016, 24, 188–202. [Google Scholar] [CrossRef]
- Michel, V.; Shen, C.C.; Woodhead, J.; Hu, H.M.; Wu, C.C.; Moullé, P.É.; Khatib, S.; Cauche, D.; Moncel, M.H.; Valensi, P.; et al. New dating evidence of the early presence of hominins in Southern Europe. Sci. Rep. 2017, 7, 10074. [Google Scholar] [CrossRef]
- Cherin, M.; D’Allestro, V.; Masini, F. New bovid remains from the Early Pleistocene of Umbria (Italy) and a reappraisal of Leptobos merlai. J. Mamm. Evol. 2019, 26, 201–224. [Google Scholar] [CrossRef]
- Kostopoulos, D.S.; Aidona, E.; Benammi, M.; Gkeme, A.; Grasset, L.; Guy, F.; Koufos, G.D.; Kynigopoulou, Z.; Le Maitre, A.; Novello, A.; et al. The Lower Pleistocene primate-bearing fossil site of Dafnero (W. Macedonia, Greece): New data from classic and innovative approaches. In Proceedings of the 15th International Congress of the Geological Society of Greece, Athens, Greece, 22–24 May 2019. [Google Scholar]
- Lopatin, A.V.; Vislobokova, I.A.; Lavrov, A.V.; Startsev, D.B.; Gimranov, D.O.; Zelenkov, N.V.; Maschenko, E.N.; Sotnikova, M.V.; Tarasenko, K.K.; Titov, V.V. The Taurida cave, a new locality of Early Pleistocene vertebrates in Crimea. Dokl. Biol. Sci. 2019, 485, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Lowrie, W. Identification of ferromagnetic minerals in a rock by coercivity and unblocking temperature properties. Geophys. Res. Lett. 1990, 17, 159–162. [Google Scholar] [CrossRef]
- McFadden, P.L.; McElhinny, M.W. Classification of the reversal test in palaeomagnetism. Geophys. J. Int. 1990, 103, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Sotnikova, M.; Rook, L. Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the Late Miocene to Early Pleistocene. Quat. Int. 2010, 212, 86–97. [Google Scholar] [CrossRef]
- Cherin, M.; Bertè, D.F.; Rook, L.; Sardella, R. Re-defining Canis etruscus (Canidae, Mammalia): A new look into the evolutionary history of early Pleistocene dogs resulting from the outstanding fossil record from Pantalla (Italy). J. Mamm. Evol. 2014, 21, 95–110. [Google Scholar] [CrossRef]
- Koufos, G.D. The Pleistocene carnivores of the Mygdonia basin (Macedonia, Greece). Ann. de Paléontol. 1992, 78, 205–257. [Google Scholar]
- Spassov, N. Biochronology and zoogeographic affinities of the Villafranchian faunas of Bulgaria and South Europe. Hist. Nat. Bulgarica 2000, 12, 89–128. [Google Scholar]
- Masini, F.; Sala, B. Large- and small-mammal distribution patterns and chronostratigraphic boundaries from the Late Pliocene to the Middle Pleistocene of the Italian peninsula. Quat. Int. 2007, 160, 43–56. [Google Scholar] [CrossRef]
- Vislobokova, I.A.; Agadjanian, A.K. New data on large mammals of the Pleistocene Trlica fauna, Montenegro, the Central Balkans. Paleontol. J. 2015, 49, 651–667. [Google Scholar] [CrossRef]
- Vislobokova, I.A.; Titov, V.V.; Lavrov, A.V.; Gimranov, D.O.; Startsev, D.B.; Tasasenko, K.K. Early Pleistocene spiral-horned antelopes (Artiodactyla, Bovidae) from the Taurida Cave (Crimea, Russia). Paleontol. J. 2020, 54, 81–90. [Google Scholar] [CrossRef]
- Abbazzi, L. La fauna de cérvidos de Barranco León y Fuente Nueva-3. In Ocupaciones Humanas en el Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza; Toro, I., Martínez-Navarro, B., Agustí, J., Eds.; Consejería de Cultura, Junta de Andalucía, Arqueología Monografías: Sevilla, Spain, 2010; pp. 273–290. [Google Scholar]
- Athanassiou, A. New giraffid (Artiodactyla) material from the Lower Pleistocene locality of Sésklo (SE Thessaly, Greece): Evidence for an extension of the genus Palaeotragus into the Pleistocene. Zitteliana B 2014, 32, 71–89. [Google Scholar]
- Masini, F.; Palombo, M.R.; Rozzi, R. A reappraisal of the Early to Middle Pleistocene Italian Bovidae. Quat. Int. 2013, 288, 45–62. [Google Scholar] [CrossRef]
- Sorbelli, L.; Madurell-Malapeira, J.; Prat-Vericat, M.; Azzarà, B.; Romi, P.; Gentili, S.; Cherin, M. Primitive Bison from the late Villafranchian site of Pietrafitta (Italy). In Proceedings of the Invertebrate and Vertebrate Paleontology day (IVPday), Pisa, Italy, 17–20 November 2020; pp. 112–116. [Google Scholar]
- Sardella, R.; Bellucci, L.; Bona, F.; Cherin, M.; Iurino, D.A.; Rook, L. Before and after the earliest Homo dispersal in Europe: Evidence from the early Pleistocene sites of the Italian Peninsula. In European Early Pleistocene Biogeography and Ecology Based on the Mammal Record: Case Studies and Preliminary Syntheses; Kostopoulos, D.S., Konidaris, G., Tesakov, A., van den Hoek Ostende, L.W., Rook, L., Eds.; Comptes Rendus Palevol: Amsterdam, The Netherlands, 2018; Volume 17, pp. 287–295. [Google Scholar]
- Cohen, K.M.; Gibbard, P.L. Global chronostratigraphical correlation table for the last 2.7 million years, version 2019 QI-500. Quat. Int. 2019, 500, 20–31. [Google Scholar] [CrossRef]
- Guérin, C.; Faure, M.; Argant, A.; Argant, J.; Crégut-Bonnoure, É.; Debard, É.; Delson, E.; Eisenmann, V.; Hugueney, M.; Limondin-Lozouet, N.; et al. Le gisement pliocène supérieur de Saint-Vallier (Drôme, France): Synthèse biostratigraphique et paléoécologique. Geobios 2004, 37, S349–S360. [Google Scholar]
- Delson, E.; Faure, M.; Guerin, C.; Aprile, L.; Argant, J.; Blackwell, B.A.B.; Debard, E.; Harcourt-Smith, W.; Martin-Suarez, E.; Monguillon, A.; et al. Franco-American renewed research at the Late Villafranchian locality of Senèze (Haute-Loire, France). Cour. Forsch.-Inst. Senckenberg 2006, 256, 275–290. [Google Scholar]
- Radulescu, C.; Samson, P.M. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Romania. Boll. Soc. Paleontol. Ital. 2001, 40, 285–291. [Google Scholar]
- Terhune, C.E.; Curran, S.; Croitor, R.; Drăgușin, V.; Gaudin, T.; Petculescu, A.; Robinson, C.; Robu, M.; Werdelin, L. Early Pleistocene fauna of the Olteţ River Valley of Romania: Biochronological and biogeographic implications. Quat. Int. 2020, 553, 14–33. [Google Scholar] [CrossRef]
- Bellucci, L.; Sardella, R. The last Antilopini bovids from the Early Pleistocene of Italy. Quat. Int. 2015, 357, 245–252. [Google Scholar] [CrossRef]
- Martínez-Navarro, B.; Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Medin, T.; Hortolà, P.; Palmqvist, P. The Epivillafranchian and the arrival of pigs into Europe. Quat. Int. 2015, 389, 131–138. [Google Scholar] [CrossRef]
- Steensma, K.J. Plio-/Pleistozäne Großsäugetiere (Mammalia) aus dem Beckem von Kastoria/Grevena, südlich von Neapolis - NW-Griechenland. Ph.D. Thesis, Technische Universität Clausthal, Clausthal, Germany, 1988. [Google Scholar]
- Palombo, M.R. To what extent could functional diversity be a useful tool in inferring ecosystem responses to past climate changes? Quat. Int. 2016, 413, 15–31. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus | Species |
|---|---|---|---|
| Proboscidea | Elephantidae | Mammuthus | meridionalis |
| Carnivora | Canidae | Canis | sp. |
| Ursidae | Ursus | etruscus | |
| Hyaenidae | Pachycrocuta | brevirostris | |
| Felidae | Megantereon | sp. | |
| Perissodactyla | Equidae | Equus | sp. (medium-sized) |
| Equus | sp. (large-sized) | ||
| Rhinocerotidae | Stephanorhinus | sp. | |
| Artiodactyla | Giraffidae | Palaeotragus | sp. |
| Cervidae | Cervus | sp. | |
| Praemegaceros | sp. | ||
| Bovidae | Pontoceros | sp. | |
| Leptobos | sp. | ||
| Bison | cf. degiulli |
| Sample ID | Lab ID | Grain Size (μm) | Qtz Dissolved (g) | m(27Al) (ppm) | 26Al/27Al | 1σ Error (%) | 26Al Conc. 104 atoms/g(qtz) | m(9Be) (mg) | 10Be/9Be | 1σ Error (%) | 10Be Conc. 104 atoms/g(qtz) | 26Al/10Be | Simple Burial Age (Myr) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COSMO 1 | GL11 | 250–500 | 46.76 | 121 | 1.39 × 10−13 | 4.46 | 37.07 ± 1.91 | 0.3397 | 2.16 × 10−13 | 3.60 | 10.51 ± 0.42 | 3.53 ± 0.23 | 1.54 ± 0.10 |
| COSMO 2 | GL7 | 250–500 | 76.52 | 81 | 1.80 × 10−13 | 5.23 | 32.48 ± 1.70 | 0.2868 | 4.10 × 10−13 | 3.34 | 10.23 ± 0.34 | 3.18 ± 0.20 | 1.75 ± 0.10 |
| COSMO 3 | GL8 | 250–500 | 55.97 | 65 | 2.20 × 10−13 | 5.44 | 31.88 ± 1.74 | 0.2874 | 2.76 × 10−13 | 3.51 | 9.40 ± 0.33 | 3.39 ± 0.22 | 1.62 ± 0.10 |
| Mean Directions In Situ | Palaeopole | ||||||
|---|---|---|---|---|---|---|---|
| N | k | α95 | Dec. | Inc. | Lat. | Long. | A95 |
| 23 | 9.0 | 10.7° | 176.8 °E | −40.5° | 72.4 °N | 213.2 °E | 7.8° |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konidaris, G.E.; Kostopoulos, D.S.; Maron, M.; Schaller, M.; Ehlers, T.A.; Aidona, E.; Marini, M.; Tourloukis, V.; Muttoni, G.; Koufos, G.D.; et al. Dating of the Lower Pleistocene Vertebrate Site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, Magnetostratigraphy, and Cosmogenic Radionuclides. Quaternary 2021, 4, 1. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4010001
Konidaris GE, Kostopoulos DS, Maron M, Schaller M, Ehlers TA, Aidona E, Marini M, Tourloukis V, Muttoni G, Koufos GD, et al. Dating of the Lower Pleistocene Vertebrate Site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, Magnetostratigraphy, and Cosmogenic Radionuclides. Quaternary. 2021; 4(1):1. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4010001
Chicago/Turabian StyleKonidaris, George E., Dimitris S. Kostopoulos, Matteo Maron, Mirjam Schaller, Todd A. Ehlers, Elina Aidona, Mattia Marini, Vangelis Tourloukis, Giovanni Muttoni, George D. Koufos, and et al. 2021. "Dating of the Lower Pleistocene Vertebrate Site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, Magnetostratigraphy, and Cosmogenic Radionuclides" Quaternary 4, no. 1: 1. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4010001