
Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications


1
Departamento de Arqueología, Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires, Avda. del Valle 5737, Olavarría, Buenos Aires B7400JWI, Argentina
2
Departamento de Paleobiología, Museo Nacional de Ciencias Naturales (CSIC), José Gutiérrez Abascal, 2, 28006 Madrid, Spain
*
Author to whom correspondence should be addressed.
Quaternary 2021, 4(2), 15; https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020015
Submission received: 8 February 2021
/
Revised: 22 March 2021
/
Accepted: 20 April 2021
/
Published: 11 May 2021
(This article belongs to the Special Issue Advances in Quaternary Studies: The Contribution of Mammalian Fossil Record II)
Abstract
:The Pampean Region contains sedimentary sequences with abundant mammal fossil records, which constitute the chronological outline of the Plio–Pleistocene of South America. These classic localities have been used for more than a century to correlate with other South American regions. Throughout this time, a series of misinterpretations have appeared. To understand the stratigraphic significance of these localities and the geochronological situation of each unit referring to the Pleistocene, a critical historical study of the antecedents was carried out, evaluating the state of each unit. The biostratigraphic studies of the Pampean Region’s mammalian faunas improved the understanding of biogeographic changes taking into account the environmental fluctuations of the Pleistocene.
1. Introduction
In 1833, Charles Darwin explored the Pampas of southern Argentina. His annotations, as well as those of D’Orbigny [1], were developed within the framework of the ideas of his time that proclaimed a scheme of the Earth’s past in which the rest of the regions of the world were incorporated [2]. Darwin visited a large part of the Pampas landscape. He was surprised by the vast extension and abundance of fossil remains, which were the most significant aspects that he mentioned when summarizing the Argentine. These notes were the basis that Ameghino [3,4] used to develop a stratigraphic scheme.
The surface sediments that cover most of the Pampas Region were originally called “terrains pampéenes” and “argile pampéenes” [1], “pampean formation” [5], and “pampa formation” [6]. Ameghino [3] proposed a subdivision of the “Pampean formation” into three units: Lower Pampean, Upper Pampean, and Lacustrine Pampean. These deposits are characterized by presenting a lithological homogeneity, which added to the observed discontinuities and led to the search for other criteria to order them. One of these criteria for separating the deposits into different stratigraphic units was the content of fossil mammals. A significant case was the classic localities between Mar del Plata and Miramar, which present laterally continuous exposures and allowed subdivision based on their paleontological content [4]. Another fact to highlight is that the first stratigraphic analyses of the Pampean deposits were favored by large excavations due to the construction of the ports of Buenos Aires and La Plata, which provided new sections for the analysis [7]. This unit scheme is based on the content of fossil mammals derived from the South America Land Mammals Ages (SALMA) [8], a stratigraphic arrangement widely accepted among paleontologists. We use the evolutionary degree of the faunal assemblages in the stratigraphic ordering of the Pampean deposits.
Recently, a new contribution with different locations and more precise records, a biostratigraphic scheme, was designed in Pleistocene Pampean deposits [7,9,10,11,12,13,14,15,16,17]. Through this scheme, appropriate relationships of superposition of first and last appearance of mammalian record can be established, and thus a relative chronology can be established.
In this paper, we summarize the main biotic and abiotic events that occurred in Pampean Region in the Pleistocene: glaciations; diastrophism, marine regressions, and transgressions; marked replacements of distribution and quality of the vast southern plains; a geographical connection between the Americas; and the Great American Biotic Interchange (GABI) and the Quaternary Megafauna Extinction (QME) modified the landscape of the region, and strongly influenced its present status.
2. Characteristics of the Pampean Region
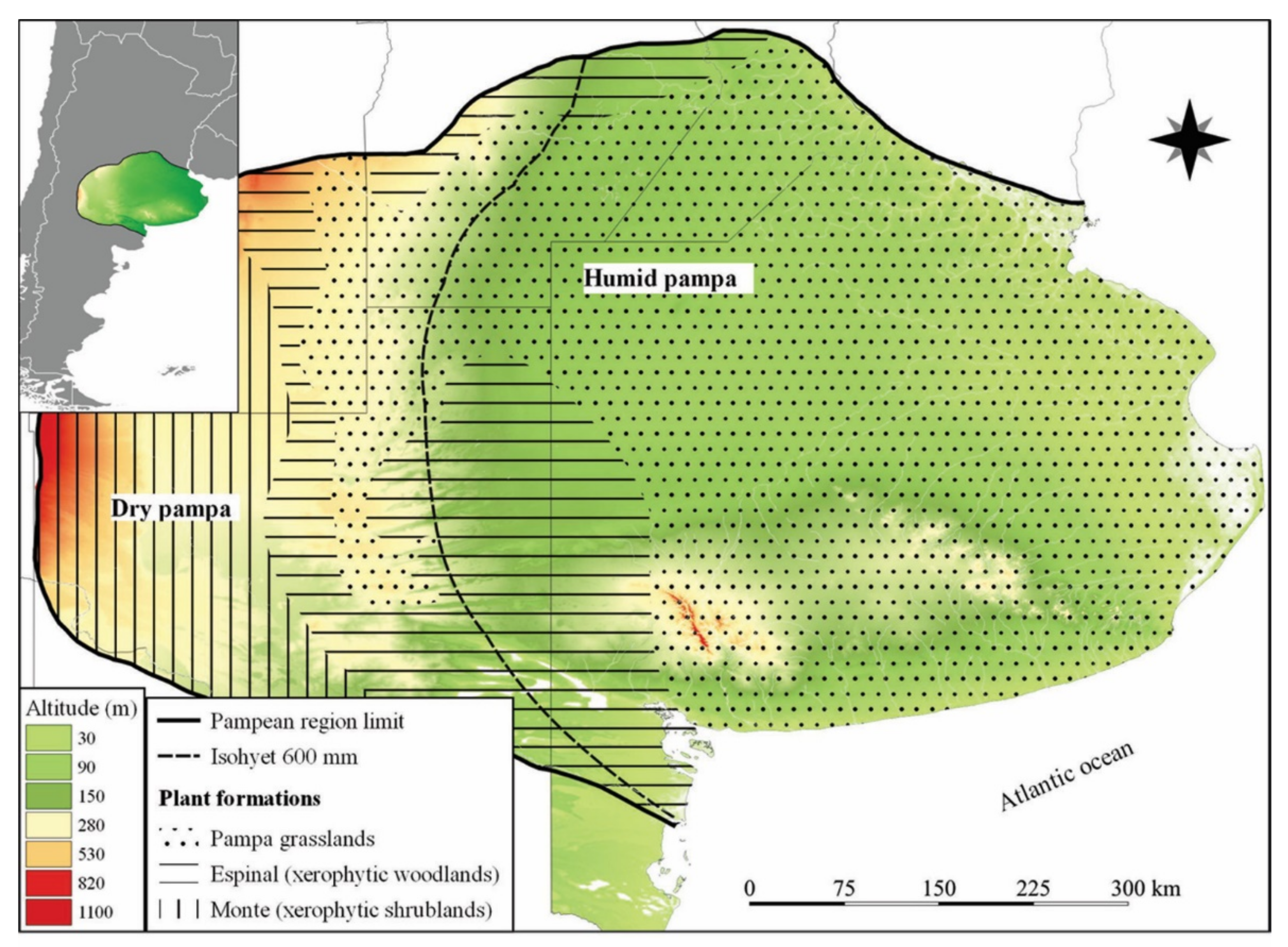
The Pampean Region (Figure 1) is located approximately between latitude 30–39° S and longitude 57–66° W and occupies a large area (~ 600,000 km2). The prevailing climate in this region is temperate to subtropical and humid. The annual average temperature can oscillate between 13 and 18 °C, and the annual precipitation records are between 600 and 1200 mm. The climate of the region is controlled by the Atlantic Anticyclone, whose effect decreases in influence from the northeast to the southwest [18]. In the Pampean Region, two subregions are recognized: the Humid Pampa in the northeast and the Dry Pampa in the southwest [19,20]. This division of the region enables the differentiation of three different ecoregions: Pampa (same extent as the Humid Pampa), Espinal, and Low Monte (the Dry Pampa includes both Espinal and Low Monte) [19]. From a biogeographic approach, the study area is within the Pampean zoogeographic domain of the Guayano–Brazilian subregion [21] and the phytogeographic Pampean province of the Chaqueño domain [22]. Recent reviews of the biogeography of South America include the Region Pampeana in the province called Pampa [23]. The vegetation is predominantly herbaceous steppe or pseudo-steppe, although communities of grass prairies and psammophytic and halophytic steppes are registered [24].
The region presents a mainly flat landscape with some slight undulations. A deep mantle of loess and loessic silts is interrupted by the relatively low hills called a system of Ventana and Tandil [25]. The Pleistocene sediments of the Pampean Region were deposited mainly under arid to semi-arid climatic conditions, alternating with indicator levels of more humid conditions [26]. The paleoclimate record indicates that this fluctuation results from the interaction between the air masses of the Pacific and Atlantic anticyclones [27]. At present, Pacific Anticyclone loses its humidity when crossing the Andes Mountains and arrives cold and dry in the Pampean Region. For its part, the winds of the Atlantic Anticyclone are frequently warm and humid.
3. Geology
The Pampean Region includes a large sedimentary basin whose strata date back from the Cretaceous to the Holocene [28]. The younger strata are loess deposits dating back to the Late Cenozoic. For this reason, some authors have included these sediments in a single lithostratigraphic unit, called the Pampeana Formation [26,29,30]. For their part, Riggi et al. [31] and Tonni et al. [32] recognized the Early–Middle Ensenada Formation and the Middle–Late Pleistocene Buenos Aires Formation [33]. Most of these deposits correspond to aquatic facies and a few correspond to primary eolian facies [34]. Moreover, Kraglievich [35] proposed other units based on other correlated deposits in the coastal cliffs between Mar del Plata and Miramar. The Ensenada and Buenos Aires Formations are assignable to the Ensenada and Bonaerian–Lujanian Stage, respectively.
Several hypotheses have been put forward on the genesis of the Pampean sediments. González Bonorino [36] proposed that the Pampean sediments come mainly from the weathering, transport, and erosion of tertiary formations (tertiary basalts of the Patagonian region and crystalline basalt of the Pampean mountain ranges) and Pleistocene volcanism in the Andes Mountains. More recently, it was proposed that there are several loess contribution zones (in addition to the classic ones) that are located to the north and west of the Pampean Region and involve different stages [37]. Rabassa et al. [38] attribute the Pampean loess deposit to the wind activity generated by the climatic conditions derived from the glacial advance, which influences the Pacific’s marine anticyclonic centers, shifting their axis to the north. The development of paleosol is widespread in the Pampas sediments. Overlapping paleosols, resulting from the succession of pedogenetic processes, are found in Pliocene and Pleistocene sediments at different regional locations.
Fidalgo et al. [29,30,39] reviewed the Late Quaternary nomenclature and stratigraphy of the Pampean Region. With new geomorphological data, plus analysis of texture and composition of Late Pleistocene and Holocene deposits, they defined two stratigraphic units: the Luján Formation, which includes the Guerrero and Río Salado members; and the La Postrera Formation. The first is of fluvial-lake origin, and the second is eolian (Figure 2).
The La Postrera Formation that covers the entire region includes loess and very fine stratified sands [41]. The sedimentological analysis of eolian material [42] suggests that volcanism in the Andean region was the predominant source of the contribution, specifically, as a contribution of pyroclastic deposits that are made by erosive fluvial and wind processes. Iriondo [43] described a Pampean Wind System based on geomorphological evidence, which would partly explain the origin of these sediments in the central region of Argentina. He proposed that the sediments were redistributed from the Colorado River system. This hypothesis partially fits with the interpretation proposed by Zárate and Blasi [41,42]. For these authors, the sandy loess that covers the southeastern Pampean region would have its origin in the alluvial plains of Colorado, and Negro and the southwesterly winds are predominant in the area today [18,44]. On the other, geomorphological data, such as the orientation of lunettes and dunes, indicate westerly winds in the past [45]. Gardenal [46] proposed four main groups of aeolian sediments in the Pampean region’s western part. However, the calibration of these depositional episodes is complex. According to Zárate and Blasi [42], the first and second episodes correspond to the late Pleistocene, the third to the Middle Holocene, and the last to historical times.
Likewise, the Luján Formation is made up of highly developed alluvial plain deposits together with a wide fluvial system of the province of Buenos Aires [29]. The lower levels of this formation correspond essentially to alluvial deposits, such as the La Chumbiada and Guerrero members. In contrast, the upper levels are mainly lacustrine, such as the Río Salado member. In two localities (Paso Otero and Empalme Querandies), the Guerrero member has two facies (one predominantly brown, the other green), among which an ecological succession is evidenced, as its faunas indicate environmental deterioration [47]. The lower brown stratum (members of La Chumbiada, sensu Dillon and Rabassa [48]) is characterized by a fauna of mammals with a high frequency of browsers. On the other hand, the upper green levels (Guerrero member) show an abundance of large herbivores and a decrease in browsing forms [47].
Two pedostratigraphic units are also recognized. The first one, called Puesto Callejón Viejo Soil occurs in the superior part of the Guerrero Member. The other, known as Puesto Berrondo Soil, developed in the Río Salado Member [39]. The latter soil indicates a decrease in humidity at a regional scale with the fluvio-lacustrine environments’ desiccation corresponding to the Rio Salado Member [49].
In the southern coastal zone of the Pampean Region, there are two marine transgressions assigned to the last interglacial and the Middle Holocene [50]. The Pascua Formation is made up of Late Pleistocene marine deposits [29] and occurs in different locations along the coast [51,52,53]. They are also correlated with the Chuy Formation recognized in Uruguay [54] and the transgression event described for the Late Pleistocene of Brazil [55].
4. Paleoclimatic History and Mammal Diversity
The Late Neogene in South America was marked by active diastrophism and a high frequency of sea-level changes [56]. Both events are related and connected to the various causes of cyclical and climate changes. The sea level’s successive descents are connected to the progressive events of diastrophism called the Diaguita Phase. Thus, the most apparent event of this diastrophic phase was the establishment of the Panamanian Land Bridge that interrupted communication between the Atlantic and Pacific oceans. This event radically changed both oceans’ circulation, facilitating the emergence of the Arctic ice cap [57]. The cold Circum–Antarctic Current of the Southern Hemisphere was established, and the ice sheet formation in Antarctica began [58].
The main differences between the climatic–environmental conditions of the Tertiary and the Quaternary are linked to the amplitude and frequency of environmental changes. In the Tertiary, geological and paleontological records show long intervals of uniform environmental conditions, sporadically interrupted by geologically short but highly disruptive change intervals. On the other hand, the climatic changes of the Quaternary had greater amplitude and frequency. Both types of changes have different effects on biota. Environmental changes during the Tertiary significantly altered the taxonomic and ecological composition of mammalian communities, operating through cladogenetic processes, extinction, and immigration from other areas. The changes that occurred during the Quaternary are responsible for chorological variations, expanding or contracting the distribution areas of species, communities, and biomes [59,60,61]. The successive retreat and advance of the glaciers produced, in non-engulfed areas such as the Pampeana Region, an alternation of arid (savanna, steppes) and humid (subtropical forests) biomes. This particularity favored high extinction rates as a consequence of the reduction of favorable areas [62]. During the coldest and driest periods, there was a retraction of the subtropical areas, which facilitated the expansion and interconnection of open biomes [63,64].
Since the Pampean Region can currently be considered an ecotone between two important biogeographic areas (Brazilian and Patagonian), the fauna associations included mixtures of taxa from both areas at different times during the Pleistocene. In cold and dry climatic conditions, a contraction of the areas occupied by tropical and subtropical fauna was observed, which resulted in the interconnection of the open environment fauna. On the other hand, when open areas retreated during interglacial periods, the forest expanded. The arid and semi-arid conditions allowed the expansion of the Patagonian and Central mammals (Lestodelphis, Microcavia, and Zaedyus) into areas occupied by the subtropical mammals (i.e., echimyids, procyonids, and tayassuids), which presently inhabit warmer and more humid environments [9,65].
Paleoecological studies by Fariña [66] and Croft [67] suggest that savanna and grassland habitats expanded under dry climates in the Pampean Region. In those times, the steppes were well developed, favored by cold and arid or semi-arid conditions. Some typical mammals of the current Patagonian and Central subregions (i.e., Microcavia australis and Pediolagus salinicola) are also recorded during the Late Pleistocene in the Pampean Region [9]. Tonni and Cione [10] pointed out that there are also some hot and humid climatic periods, of short duration, interspersed between more extended periods with cooler and more arid conditions.
The range of δO18 calculated for the Equidae of the Pampean Region corresponds to a variation of 4.2 °C in the mean annual temperatures throughout the Pleistocene [68]. As a reference, the current temperature ranges between 13.4 °C in Mar del Plata (38 °S), 16.4 °C in Buenos Aires (34 °S), and 19.2 °C in Santa Fé (32 °S).
According to Sánchez et al. [68], carbon isotope analyses in Equidae from the Pampas Region suggest a change in the pasture: Early Pleistocene horses consumed predominantly C3 plants, and then consumed a C3–C4 mixture in the Middle–Late Pleistocene; towards the Late Pleistocene, they returned to a strict diet of C3 plants.
Cione and Tonni [10] point out that the mammal communities of the Marplatan stage suggest warm and humid conditions. In the Early Ensenada stage, recorded mammals suggest the beginning of a cold, semi-arid period [9]. Mammal records from the Late Ensenadana stage indicate the return of warm and humid conditions [10].
The record of Echimyidae, Dasyproctidae, and Noctilionidae [69] in the Early Bonaerian stage indicates the beginning of a new warm event. The presence of Ctenomys kraglievichi and Tayassuidae suggests a strong warm pulse around 0.5 Ma [15,70]. Evidence from other continents also suggests a warm pulse in the Middle Pleistocene, between 0.7 and 0.3 Ma [71].
The interglacial conditions of MIS5e (130 ka) can be inferred from records of mammals of the Lujanian stage [72]. It also happens that during the Last Glacial Maximum (18 ka), mammalian indicators of arid and cold conditions predominated [73]. The pollen record indicates that Late Pleistocene environments were arid, associated with more continental environments and lower sea levels [70]. Isotopes found in terrestrial gastropods indicate a change to a more humid environment in the Pleistocene–Holocene transition [74].
5. The Land Mammals Ages and Biostratigraphy
Ameghino [75,76] proposed a stratigraphic scale that was a reference standard for continental Cenozoic South America. It is a sequence of stages grouped in higher-order units [75,76]. This Ameghino proposal, based on biostratigraphic criteria, is still partly valid today. He proposed a subdivision of the “Pampean formation” into three units: Lower Pampean, Upper Pampean, and Lacustrine Pampean. Ameghino’s stratigraphic conception does not differ from that which was developing in Europe at the same time. Ameghino [75] introduced terms such as “formation” and “floor”, which are currently used to refer to units of the lithostratigraphic and chronostratigraphic classification. However, for Ameghino, a “formation” was a unit of time that was inferred based on its fossil content, where lithology was not a necessary element for its name, and even less the only one. Later, when Frenguelli [77,78] proposed his stratigraphic scheme, he used Ameghino’s names but with another connotation.
The Pampian stages of Frenguelli are the Chapalmalense, Ensenadense, and Bonaerense. The post-Pampa stages are the Lujanense, Platense, Querandinense, Cordobense, and Aimarense. The three stages of Pampian are assigned to the Pleistocene, and the five stages after Pampian represent the Holocene (Figure 2).
Kraglievich [35] established another stratigraphic scheme for the southeast of the Pampean Region, where he recognized several formations based on some of the classic names of Ameghino, which were redefined; for example, the “Chapalmalense horizon” or “Chapalmalense stage” became the “Chapadmalal formation”. Although Kraglievich described the lithological characters somewhat, the discrimination between the units rested fundamentally on the paleontological content.
Until the 1940s, there was considerable confusion because the same names were often used to distinguish rock, time, and faunal units. Simpson’s [79] study brought order to this vast body of information. Later, Wood et al. [80] proposed a new scheme based on several Land Mammals Ages (LMA) for North American records. LMA was a valuable scheme to order and establish a relative age of the Cenozoic terrestrial deposits on this continent. Woodburne [81] used the term North American Terrestrial Mammal Ages (NALMAs) as a schema of formal biochronological units, which was later taken as the basis for the proposed schema of Archibald et al. [82], among others. Pascual et al. [8,83] proposed a new synthesis, based on the criteria of Ameghino (evolutionary grade of the faunas), that closely corresponds with the LMA concept proposed by Savage [84]. The SALMA arrangement comprises unique taxon associations that are inferred to have existed during a restricted time interval (Table 1). These Ages were established using knowledge of the faunas’ evolutionary grade, in the same way that Ameghino did, by noting the times of first and last taxa appearances in the fossil record and other related changes in faunal assemblages over time [59]. The LMA are stratigraphic units that, for the moment, have not been formally recognized by the stratigraphic code of nomenclature of Argentina. However, as a classifying scheme, they have been very useful in evolutionary and biogeographic mammal studies [84,85]. Pascual et al. [59], among others, have underlined its usefulness in establishing intra- and intercontinental correlations of Cenozoic time.
6. Mammal Biozone Scheme for the Pampas Region
In recents years, four types of regional stage were distinguished, namely (1) alternative regional stages; (2) concurrent regional stages; (3) biogeographical regional stages; and (4) regional land stages [40]. All regional stages are essentially incomparable with the global stages, established with Global Boundary Stratotype Section and Point (GSSP).
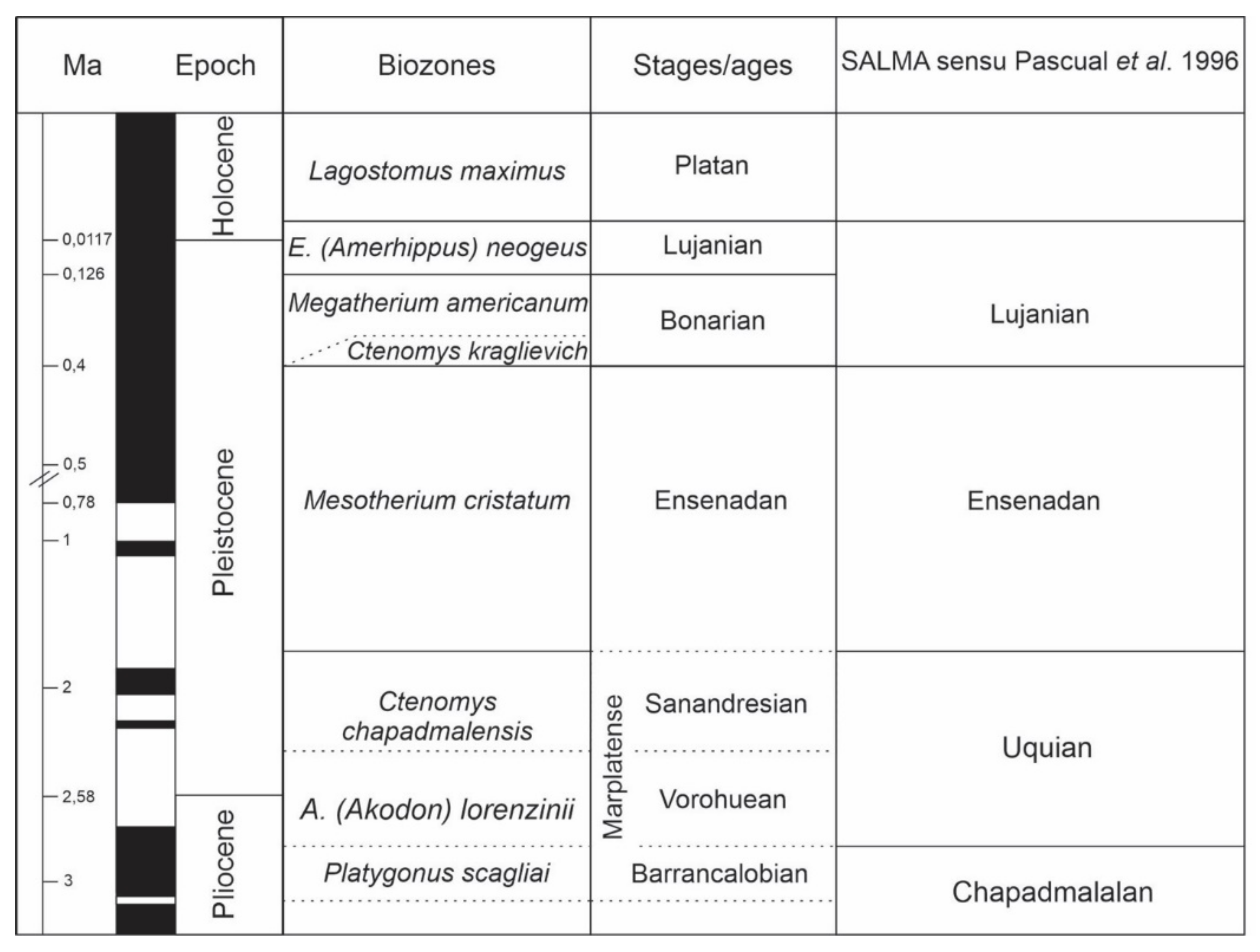
In South America, the regional land stages (type 4) are the South American Land Mammal Ages, defined by Pascual et al. [8] and are alternatives to the chronostratigraphic units developed from the terrestrial record. These land mammals ages are used widely. Some can be confidently linked to global chronostratigraphic divisions whereas the correlation of some others is not so clear. These alternative regional stages should be abandoned and, where possible, replaced by biozones or lithostratigraphic units since they make no contribution in the development of chronostratigraphy. For example, Cione and Tonni [86] has demonstrated the efficacy of the Neogene mammal zones in Europe in contrast to the European Land Mammal Ages. In the last decades, Cione and Tonni [10] recommended the substitution of the LMA by a chronostratigraphic/geochronological classification based on biostratigraphy, according to the stratigraphic code of Argentina. In successive presentation, these authors identified five biozones (Association Zones, or Interval Zones) for the Pleistocene [87,88]. These biozones are those of Ctenomys chapadmalensis, Mesotherium cristatum, Megatherium americanum and Equus neogeus. They constitute the biostratigraphic base that sustains the Marplatan, Ensenadense, Bonaerense, and Lujanense Stages/Ages. Considering that the “International Commission on Stratigraphy” [89] established the Plio-Pleistocene boundary in 2.58 Ma, the Sanandresense Sub-Stage of the Marplatan represents in the Pampean Region the beginning of the Quaternary (Table 1).
This new scheme, the sequence of names proposed by Ameghino is used with few modifications for the Quaternary of Pampean Region. Consequently, the use of a term must be associated with its definition in the classification system; otherwise, it will generate confusion and incorrect interpretations. In this proposal, the biostratigraphic units are based on characters, such as the spatial location of the taxa, which are verified in the field; however, there is still a certain level of abstraction in their recognition (Figure 3).
6.1. Ctenomys chapalmalensis Biozone
This biozone coincides with the Upper Marplatan (Early Pleistocene), particularly with the San Andrés Formation. Ctenomys chapalmalensis is an exclusive taxon from the San Andrés Formation, and it represents the oldest record of Ctenomys in this region. The deposit of this formation and its fauna content was correlated with Patagonia’s glacial advances to MIS82 to 78 [90]. This biozone is characterized by the last record of Eumysops, whose species were widespread during the Pliocene and Early Pleistocene. [91].
6.2. Mesotherium cristatum Biozone
This biozone coincides with the Ensenadan (Lower to Middle Pleistocene), particularly with the Ensenada Formation in the Pampean Region [32] and with Miramar Formation [35], in the southeast. Numerous taxa of biostratigraphic importance are recorded in the Ensenadan, some exclusive to this stage (Panochthus intermedius, Glyptodon munizi, Neosclerocalyptus pseudornatus, Neosclerocalyptus ornatus, Megatherium gallardoi, Scelidotherium bravardi, Antifer ensenadensis, Catagonus metropolitanus, Theriodictis platensis, Arctotherium angustidens, and Mesotherium cristatum).
As the type profiles in which the fossils used by Ameghino [74] as the basis for his Ensenadan (Puerto de La Plata, Ensenada) were found are not currently available, Tonni et al. [32] suggest as a new type locality a quarry located in Hernández (La Plata County), whose sequence includes characteristic fossils of the Mesotherium cristatum biozone [88].
6.3. Megatherium americanum Biozone
This biozone coincides with the Bonaerian (Middle Pleistocene). Its lower limit coincides with the lower part of the Buenos Aires Formation in the northeast of the Pampean Region [32,33] and probably with Arroyo Seco Formation [35] in the southeast. According to the fauna evidence, it begins with a warm event, with a predominance of pedogenesis and typical Brazilian mammals’ registry. Considering these particularities and the stratigraphic situation, Verzi et al. [15] suggest that the Bonaerian begins in the interglacial period corresponding to MIS11, around 0.4 Ma [90,92]. This interglacial event was the warmest and the longest in the last 0.5 Ma, generating a significant rise in sea level, observed in the beach sediments of the southern province of Buenos Aires. [52,88].
Numerous taxa with biostratigraphic importance are recorded for the first time on this stage (Panochthus tuberculatus, Megatherium americanum, Glyptodon elongatus, Glyptodon reticulatus, Lestodon armatus, Mylodon darwini, Scelidotherium leptocephalum, Glossotherium robustum, Eutatus seguini, Macrauchenia patachonica, Toxodon platensis, Artotherium bonariense, Pecari tajacu, and Lama gracilis).
6.4. Equus neogeus Biozone
This biozone coincides with the Lujanian (Upper Pleistocene, [12,94]. However, Cione and Tonni [40] suggest that this biozone does not agree with the SALMA Lujanian proposed by Pascual et al. [8], who also include the Bonarian stage.
The guide fossil Equus neogeus is recorded from the base of the unit, which is correlated with MIS5e (ca.130 ka BP) [13,87]. This interglacial period is probably represented by the coastal sea levels assigned to the Pascua Formation [29]. Recent dating suggests that a part of the deposits referred to as the Pascua Formation and the “Belgranense” can be correlated with part of the MIS3 [95,96].
Some exclusive taxa are registered (Doedicurus clavicaudatus, Neosclerocalyptus paskoensis, and Equus neogeus). In contrast, others are registered for the first time in the region (Panochthus greslebini, Microcavia australis, Dolichotis patagonum, Eligmodontia typus, Galea musteloides, and Lycalopex gymnocercus).
7. Great American Biotic Interchange
Biogeographic studies have shown that South America was an insular continent, isolated from the rest of the continents, for a period of time that spans much of the Cenozoic [97,98]. Under these particular conditions, South American land mammals evolved in their world until the Isthmus of Panama. By the Late Miocene, the descendants of these autochthonous lineages (condylarths, notoungulates, lithopterns, marsupials and edentates) and immigrants from Africa arriving in the Late Eocene or Early Oligocene (platyrrhine primates and caviomorph rodents), had achieved a balance in their communities. A peculiarity of these faunas is the coexistence of carnivorous and omnivorous marsupials with placental ungulates and edentates [78,98].
The GABI was the most important event from a biogeographical perspective in the Late Cenozoic [99,100]. The earliest appearances of migrating terrestrial animals demonstrate intercontinental dispersals’ timings and help to understand when the Isthmus of Panama formed [101]. Recent works on the paleobiology of the mammals from the Argentine Pampas have refined our comprehension of its final formation [102]. Around 2.8 Ma ago, the tectonic activities of the Pacific margin connected the two Americas. A habitat corridor was opened, which helped the dispersal of plants and animals between the Americas, thus producing the GABI [103]. This land corridor functioned selectively [104]. The partial replacement of native South American mammals by North American immigrants is due to three different causes. First, the arrival of new ungulates and carnivores to South America. The native ungulates suffered pressure from the holarctic placental predators. For its part, there was an imbalance in the diversity of mammalian species on each continent because they are areas of different sizes. Third, global climate changes caused changes in the Isthmus of Panama’s environments that lead to an imbalance between the number of South American and North American migrants [103]. It has been suggested that there were three main types of habitat corridors: tropical mesic forest, mesic savanna, and xeric scrub savanna [104]. In the humid interglacial phase, jungle environments dominated the tropics, and the main movement of fauna was from the Amazon to Central America. During the more arid glacial phase, the directional pattern was reversed [103]. Various groups of mammals moved from one continent to another, although the result seems more favorable for North American immigrants. There was a significant change in the structure of the communities in South America [105,106].
8. Quaternary Megafauna Extinction
The most outstanding feature of the Pleistocene fauna of South America is the extinction of its rich megafauna [20,109]. The extinction event of species and genera of mammals (and in some cases of entire families) is frequently recorded since the Middle Miocene in the Pampean Region. These events are related to the environmental climate changes that occurred in that period time. However, these events can be considered normal, such as background extinction, compared to the Late Pleistocene’s extinction. The latter was a particularly extraordinary event where all mega-mammal species (more than 1000 kg) and most large mammals (more than 44 kg) disappeared [110,111]. This QME, whose victims were large mammals, mainly North Americans, South Americans, and Australians, has been attributed to different factors [112]. Various types of theories have been proposed for this extinction. On the one hand, some propose hypotheses related to climatic and ecological changes [113], while another group blames human activity for the disappearance of megafauna [114]. Evidence for meteorites has also been proposed [115]. The most recent approaches to QME suggest a synergy between increased human impacts and rapid climate change [116,117]. The idea of factor synergy in QME originates from the observation that extinction was most intense and rapid in central North America, where the arrival of the first Clovis hunters coincides with climate change at the end of the Pleistocene. It is also well documented that climate change itself caused at least some extinctions. This is true for species that run into impervious barriers while trying to stay in their habitat when climatic zones change. The question is to what extent the combination of rapid climate change and increasing human pressures amplifies extinction compared to the extinctions that result from each cause.
Martin [114] suggested that the extinction of large North American mammals is closely related to human populations’ rapid dispersal. This hypothesis, called “Overkill”, is based on the synchronism of extinction with this arrival. In contrast, the South American registry does not support this hypothesis. This window of coexistence would represent a less extended period than that of North America [118,119]. Archaeological sites with megafauna are not abundant and there is no evidence of mass slaughter sites in the Pampean Region [120]. In the Pampean registry, it is seen that of the total of 16 extinct genera, only five show evidence of human exploitation [121]. The hypothesis about the human pressure that contributed to the extinction is now stronger than in the past. The appearance of works on chronology, simulations, paleoclimatology, paleontology, and archeology support this debate. However, it is an oversimplification to say that an abrupt hunt in a short period time caused extinction [122,123]. The various evidence show that blitzkrieg in the strict sense can be rejected in South America.
Paleoclimate records do not suggest a unique late Pleistocene transition. Oxygen isotope records indicate that the recent glacial retreat was neither faster nor greater in magnitude than other Pleistocene changes [124]. Climate change has a great influence on the diversity of plants and animals. The question is whether the climatic changes of the late Pleistocene were sufficient to have triggered an atypical ecological response. Three ecological theories were postulated to explain the Pleistocene extinction: (1) the key species model [125], (2) coevolutionary disequilibrium, and (3) the mosaic-nutrient model. Cione et al. [110] have proposed an alternative model called “broken Zig-Zag”. This hypothesis proposes an alternation of the megafauna’s relative frequency as a consequence of the replacement between the habitats of savannas and forests of the glacial and interglacial intervals. During the last interglacial period, humans entered South America. They broke the zig-zag, hunting large mammals and putting pressure on their populations that were in decline due to changes in environmental conditions.
9. Final Remark
Two factors affecting the diversity of mammals during the Pleistocene in this region. In the first place, the glaciations and the climatic and sea-level changes associated with them. Second, the arrival of immigrants from North America [58,59,88].
Since the Pliocene, these latitudes’ climatic conditions changed from hot and humid to colder, drier, accentuating seasonality. The alternation in the advance and retreat of glaciers caused a coming and going of arid (savanna and steppes) and humid (tropical and subtropical forests) biomes. The diversity of mammals shows more variations than in previous times, highlighting the heterogeneity of the habitat. This cyclical pattern allowed the provincialism that characterizes the mammalian faunas of the Pampean Region during the Pleistocene. The evidence suggests that, biogeographically, the Pampean Region is an ecotonal area very sensitive to paleoclimatic changes. Therefore, although some taxa have a wide distribution in South America, their records do not necessarily reflect synchronicity. Consequently, is not easy to establish a precise correlation between distant locations without isotopic or paleomagnetic data.
The Pleistocene from the Pampean Region includes the Sanandresian, Ensenadan, Bonarian, and Lujanian stages, each one of which is the carrier of a specific mammalian fauna that characterizes and defines the following corresponding biozone: Ctenomys chapalmalensis, Mesotherium cristatum, Megatherium americanum, and Equus neogeus.
Author Contributions
J.L.P. and M.T.A. conceptualized the research. J.L.P., M.T.A. and J.B. gathered the data. J.L.P. wrote the first version of the paper, and M.T.A. and J.B. contributed to the final version. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Project ANPCYT PICT 2015-1512 and PICT 2019-03480 to J.L.P.; DGICYT CGL2016-79334-P, PIAR-2019 and COOP-B20287 from Spain to M.T.A.; and a Grant of the National University of Central Argentina (UNICEN) and CONICET to INCUAPA.
Acknowledgments
We wish to express our thanks to Maria Rita Palombo for inviting us to collaborate in this volume. We thank three reviewers and Ricardo Bonini for helpful comment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- d’Orbigny, A. Voyage dans l’Amérique Méridionale (le Brésil, la République Orientale del’Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou), exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833. Tome Troisième, 4° Partie: Paléontologie. Paris: P. Bertrand; Strasbourg: V. Levrault. 1842, p. 340. Available online: https://www.biodiversitylibrary.org/item/180098#page/7/mode/1up (accessed on 1 May 2021).
- Zárate, M.A.; Folguera, A.; Folguera, A. On the formations of the Pampas in the footsteps of Darwin: South of the Salado. Rev. Asoc. Geológica Argent. 2009, 64, 124–136. [Google Scholar]
- Ameghino, F. La Antigüedad del Hombre en el Plata 2; G. Masson: Paris, France; Igon Hermanos: Buenos Aires, Argentina, 1881; p. 640. [Google Scholar]
- Ameghino, F. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapalmalán. An. Mus. Nac. Buenos Aires 1908, 3, 343–428. [Google Scholar]
- Darwin, C.R. Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage of H.M.S. Beagle Round the World, under the Command of Capt. Fitz Roy, 2nd ed.; R.N. John Murray: London, UK, 1845; p. 520. [Google Scholar]
- Bravard, A. Geología de las Pampas; Registro Estadístico del Estado de Buenos Aires: Buenos Aires, Argentina, 1857; pp. 1–22. [Google Scholar]
- Soibelzon, E.; Soibelzon, L.H.; Gasparini, G.M.; Tonni, E.P. El Pleistoceno de la provincia de Buenos Aires y sus mamíferos. In Mioceno al Pleistoceno del centro y norte de Argentina; Nasif, N., Esteban, G., Chiesa, J., Zurita, A., Georgieff, S., Eds.; Opera Lilloana: Tucumán, Argentina, 2019; pp. 606–637. [Google Scholar]
- Pascual, R.; Ortega Hinojosa, E.J.; Gondar, D.; Tonni, E.P. Las edades del Cenozoico mamalífero de Argentina con especial atención a aquellas del territorio bonaerense. An. Com. Investig. Científica 1965, 6, 165–193. [Google Scholar]
- Tonni, E.P.; Alberdi, M.T.; Prado, J.L.; Bargo, M.S.; Cione, A.L. Changes of mammal assemblages in the pampean region (Argentina) and their relation with the Plio-Pleistocene boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1992, 95, 179–194. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Chronostratigraphy and ‘land mammal-ages’: The Uquian problem. J. Paleontol. 1995, 69, 135–159. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Inchasi, a Chapadmalalan (Pliocene) locality in Bolivia. Comments on the Pliocene-Pleistocene continental scale of southern South America. J. South. Am. Earth Sci. 1996, 9, 221–236. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Biostratigraphy and chronological scale of upper-most Cenozoic in the Pampean area, Argentina. In Quaternary Vertebrate Palaeontology in South America. Quaternary in South America and Antarctic Peninsula; Tonni, E.P., Cione, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; Volume 12, pp. 23–52. [Google Scholar]
- Pardiñas, U.F.J.; Gelfo, J.; San Cristóbal, J.; Cione, A.L.; Tonni, E.P. Una asociación de organismos marinos y continentales en el Pleistoceno superior en el sur de la provincia de Buenos Aires, Argentina. XIII Congr. Geológico Argent. III Congr. Explor. Hidrocarb. 1996, 5, 95–112. [Google Scholar]
- Verzi, D.H.; Lezcano, M. Status sistemático y antigüedad de Megactenomys kraglievichi Rusconi, 1930 (Rodentia, Octodontidae). Rev. Mus. La Plata 1996, 9, 239–246. [Google Scholar]
- Verzi, D.H.; Deschamps, C.M.; Tonni, E.P. Biostratigraphic and paleoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae). Palaeogeogr. Palaeoclimatol. Palaeocology 2004, 212, 315–329. [Google Scholar] [CrossRef]
- Deschamps, C.M.; Tomassini, R.L. Late Cenozoic vertebrates from the southern Pampean Region: Systematic and bio-chronostratigraphic update. Publicación Electrónica Asoc. Paleontológica Argent. 2016, 16, 202–225. [Google Scholar] [CrossRef] [Green Version]
- Soibelzon, E.; Prevosti, F.J.; Bidegain, J.C.; Rico, Y.; Tonni, E.P.; Verzi, D. Correlation of cenozoic sequences of southeast Buenos Aires Province. Biostratigraphy and magnetostratigraphy. Quat. Int. 2009, 210, 51–56. [Google Scholar] [CrossRef]
- Burgos, J.J. El clima de la provincia de Buenos Aires en relación con la vegetación y el suelo. Flora de la provincia de Buenos Aires, parte 1. In Colección Científica; INTA: Buenos Aires, Argentina, 1969; pp. 33–100. [Google Scholar]
- Mancini, M.V.; Paez, M.M.; Prieto, A.R.; Stutz, S.; Tonello, M.; Vilanova, I. Mid-Holocene climatic variability reconstruction from pollen records (32°–52°S, Argentina). Quat. Int. 2005, 132, 47–59. [Google Scholar] [CrossRef]
- Prado, J.L.; Martinez-Maza, C.; Alberdi, M.T. Megafauna extinction in South America: A new chronology for the Argentine Pampas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 425, 41–49. [Google Scholar] [CrossRef]
- Ringuelet, R.A. Rasgos fundamentales de la zoogeografía de la Argentina. Phycis 1961, 22, 151–170. [Google Scholar]
- Cabrera, A.L. Fitogeografía de la República Argentina. Boletín Soc. Argent. Botánica 1971, 14, 1–42. [Google Scholar]
- Morrone, J.J. Biogeografía de América Latina y el Caribe. Man. Tesis Soc. Entomológica Aragonesa 2001, 3, 1–144. [Google Scholar]
- Cabrera, A.L. Regiones fitogeográficas argentinas. In Enciclopedia Argentina de Agricultura y Jardinería; T II. Editorial; ACME SACI: Buenos Aires, Argentina, 1976; p. 85. [Google Scholar]
- Zárate, M.A.; Tripaldi, A. The aeolian system of central Argentina. Aeolian Res. 2012, 3, 401–417. [Google Scholar] [CrossRef]
- Tonni, E.P.; Fidalgo, F. Consideraciones sobre los cambios climáticos durante el Pleistoceno tardio-reciente en la provincia de Buenos Aires. Aspectos ecológicos y zoogeográficos relacionados. Ameghiniana 1978, 15, 235–253. [Google Scholar]
- Iriondo, M.; Garcia, M.O. Climatic variations in the Argentine Plains during the last 18,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 101, 201–220. [Google Scholar] [CrossRef]
- Irigoyen, M.R. Geología del Subsuelo y Plataforma Continental. In Proceedings of the Geología del Subsuelo y Plataforma Continental, Bahía Blanca, Argentina, 21–27 September 1975; pp. 139–169. [Google Scholar]
- Fidalgo, F.; de Francesco, O.; Colado, U. Geología superficial en las Hojas Castelli, J.M. Cobo y Monasterio (provincia de Buenos Aires). Relat. Congr. Geológico Argent. 1973, 4, 27–39. [Google Scholar]
- Fidalgo, F.; De Francesco, F.; Pacual, R. Geología superficial de la llanura bonaerense. Relat. Geol. Prov. Buenos Aires Sexto Congr. Geológico Argent. 1975, 103–138. [Google Scholar]
- Riggi, J.; Hidalgo, F.; Martínez, O.; Porro, N. Geología de los ‘‘Sedimentos Pampeanos’’ en el Partido de La Plata. Rev. Asoc. Geológica Argent. 1986, 41, 316–333. [Google Scholar]
- Tonni, E.P.; Nabel, P.; Cione, A.L.; Etchichury, M.; Tófalo, R.; Scillato Yané, G.; San Cristóbal, J.; Carlini, A.; Vargas, D. The Ensenada and Buenos Aires formations (Pleistocene) in a quarry near La Plata, Argentina. J. S. Am. Earth Sci. 1999, 12, 273–291. [Google Scholar] [CrossRef]
- Navel, P.; Cione, A.; Tonni, P. Environmental changes in the Pampean area of Argentina at the Matuyama–Brunhes (C1r–C1n) Chrons boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 403–412. [Google Scholar]
- Zárate, M. El Cenozoico Tardío continental de la provincia de Buenos Aires. In 16 Congreso Geológico Argentino (La Plata); Relatorio: La Plata, Argentina, 2005; pp. 139–158. [Google Scholar]
- Kraglievich, J.L. El perfil geológico de Chapadmalal y Miramar, Provincia de Buenos Aires. Rev. Mus. Munic. Cienc. Nat. Tradic. 1952, 1, 1–8. [Google Scholar]
- González Bonorino, F. Mineralogía de las fracciones arcilla y limo del Pampeano en el área de la ciudad de Buenos Aires y su significado estratigráfico y sedimentológico. Rev. Asoc. Geológica Argent. 1965, 20, 67–148. [Google Scholar]
- Zárate, M.A. Loess of southern South America. Quat. Sci. Rev. 2003, 22, 1987–2006. [Google Scholar] [CrossRef]
- Rabassa, J.; Coronato, A.M.; Salemme, M.C. Chronology of the Late Cenozoic Patagonia glaciations and their correlation with biostratigraphic units of the pampean region (Argentina). J. S. Am. Earth Sci. 2005, 20, 81–103. [Google Scholar] [CrossRef]
- Fidalgo, F.; Riggi, J.C.; Gentile, R.; Correa, H.; Porro, N. Los ‘sedimentos post-pampeanos’ continentales en el ámbito sur bonaerense. Rev. Asoc. Geológica Argent. 1991, 46, 239–256. [Google Scholar]
- Cione, A.L.; Tonni, E.P. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. Geología y Recursos Minerales de la Provincia de Buenos Aires. 16 Congreso Geológico Argentino. Relatorio 2005, 11, 183–200. [Google Scholar]
- Zárate, M.; Blasi, A. Late Pleistocene–Holocene Eolian deposits of the Southern Buenos Aires province, Argentina: A preliminary model. Quat. Int. 1993, 17, 15–20. [Google Scholar] [CrossRef]
- Zárate, M.; Blasi, A. Late Pleistocene and Holocene loess deposits of the southereastern Buenos Aires province, Argentina. Geojournal 1991, 24, 211–220. [Google Scholar] [CrossRef]
- Iriondo, M. Map of the South American plains. Its present state. Quat. S. Am. Antarct. Penins. 1988, 6, 297–308. [Google Scholar]
- García, N.O. South American climatology. Quat. Int. 1994, 21, 7–28. [Google Scholar] [CrossRef]
- Salazar Lea Plaza, J.C. Inventario Integrado de los Recursos Naturales de la provincia de La Pampa. Clima, Geomorfología, Suelo y Vegetación. INTA, provincia de La Pampa-UNL Pampa. Reg. Fisiográficas 1980, 98, 1–444. [Google Scholar]
- Gardenal, M. Geomorfología del partido de Saliquelló, provincia de Buenos Aires. CIC Unpubl. Rep. 1986, 60, 1. [Google Scholar]
- Prado, J.L.; Menegaz, A.; Tonni, E.P.; Salemme, M. Los mamíferos de la fauna local de Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y biostratigráficos. Ameghiniana 1987, 24, 217–233. [Google Scholar]
- Dillon, A.; Rabassa, J. Miembro La Chumbiada, Formación Luján (Pleistoceno, Provincia de BuenosAires), una nueva unidad estratigráfica del valle del río Salado. I J. Geol. Bonaer. 1985, 27, 1. [Google Scholar]
- Zárate, M.; Kemp, R.A.; Espinosa, M.; Ferrero, L. Pedosedimentary and palaeoenvironmental significance of a Holocene alluvial sequence in the southern Pampas, Argentina. Holocene 2000, 10, 481–488. [Google Scholar] [CrossRef]
- Isla, F.; Rutter, N.; Schnack, E.; Zárate, M. La transgresión Belgranense en Buenos Aires. Una revisión a cien años de su definición. Asoc. Geológica Argent. Serie D Publicación Espec. 2000, 4, 3–14. [Google Scholar]
- Aguirre, M.L.; Whatley, R.C. Late Quaternary marginal marine deposits and palaeoenvironments from northeastern Buenos Aires province, Argentina: A review. Quat. Sci. Rev. 1995, 14, 223–254. [Google Scholar] [CrossRef]
- Colado, U.; Figini, A.; Fidalgo, R.; Fucks, E. Los depósitos marinos del Cenozoico superior aflorantes en la zona comprendida entre Punta Indio y el Río Samborombón, provincia de Buenos Aires. Actas IV Jorn. Geol. Bonaer. 1995, 1, 151–158. [Google Scholar]
- Cione, A.L.; Tonni, E.P.; San Cristóbal, J.; Hernández, P.; Benítez, A.; Bordignon, F.; Perí, J. Putative meteoritic craters in Rıo Cuarto, Central Argentina interpreted as eolian landforms. Earth, Moon, and Planets. Int. J. Solar Syst. Sci. 2002, 91, 9–24. [Google Scholar]
- Martínez, S.; Ubilla, M.; Verde, M.; Perea, D.; Rojas, A.; Guérèquiz, R.; Pinñeiro, G. Paleoecology and Geochronology of Uruguayan Coastal Marine Pleistocene deposits. Quat. Res. 2001, 55, 246–254. [Google Scholar] [CrossRef]
- Suguio, K.; Martin, L. Quaternary marine formations of the State of Sao Paulo and Southern Rio de Janeiro. Int. Symp. Coast. Evol. Quat. 1978, 1, 1–55. [Google Scholar]
- Haq, B.U.; Hardenbol, J.; Vail, P.R. Chronology of fluctuation sea levels since the Triasic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, N.J. New Data on the Evolution of Pliocene Climatic Variability. In Paleoclimate and Evolution with Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 1995; pp. 242–248. [Google Scholar]
- Rabassa, J. Cuaternario de la cordillera patagónica y Tierra del Fuego. In Anales del Instituto de Geología y Recursos Minerales; Caminos, R., Ed.; Geología Argentina: Buenos Aires, Argentina, 1999; pp. 710–714. [Google Scholar]
- Pascual, R.; Ortiz Jaureguizar, E. Evolving climates and mammal faunas in Cenozoic South America. In The Platyrrhine Fossil Record; Fleagle, J.G., Rosenberg, A.L., Eds.; Academic Press: London, UK, 1990; pp. 23–60. [Google Scholar]
- Pascual, R.; Ortiz Jaureguizar, E.; Prado, J.L. Land mammals: Paradigm for Cenozoic South American Geobiotic evolution. Münchner Geowissenchaftliche Abh. A 1996, 30, 265–320. [Google Scholar]
- Ortiz Jaureguizar, E.; Cladera, G. Paleoenvironmental evolution of southern South America during the Cenozoic. J. Arid Environ. 2006, 66, 489–532. [Google Scholar] [CrossRef]
- Marshall, L.G.; Cifelli, R. Analysis of changing diversity patterns in Cenozoic Land mammal age faunas, South America. Palaeovertebrata 1990, 19, 169–210. [Google Scholar]
- Strömberg, C.A.; Dunn, R.E.; Madden, R.H.; Kohn, M.J.; Carlini, A.A. Decoupling the spread of grasslands from the evolution of grazer-type herbivores in South America. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Pascual, R. Late Tertiary mammals of southern South America as indicators of climatic deterioration. Quat. S. Am. Antarct. Penins. 1984, 2, 1–30. [Google Scholar]
- Tonni, E.P.; Fidalgo, F. Geología y paleontología de los sedimentos del Pleistoceno en el área de Punta Hermengo (Miramar, Prov. de Buenos Aires, Rep. Argentina): Aspectos paleoclimáticos. Ameghiniana 1982, 19, 79–108. [Google Scholar]
- Fariña, R.A. Limb bone strength and habits in large glyptodonts. Lethaia 1995, 28, 189–196. [Google Scholar] [CrossRef]
- Croft, D.A. Cenozoic environmental change in South America as indicated by mammalian body size distributions (cenograms). Diversity Distrib. 2001, 7, 271–287. [Google Scholar] [CrossRef]
- Sánchez, B.; Prado, J.L.; Alberdi, M.T. Ancient feeding, ecology, and extinction of Pleistocene horses from the Pampean Region, Argentina. Ameghiniana 2006, 43, 427–436. [Google Scholar]
- Vucetich, M.G.; Verzi, D.H. First record of Dasyproctidae (Rodentia) in the Pleistocene of Argentina: Paleoclimatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 178, 67–73. [Google Scholar] [CrossRef]
- Quattrocchio, M.E.; Borromei, A.M.; Deschampsc, C.M.; Grilla, S.C.; Zavala, C.A. Landscape evolution and climate changes in the Late Pleistocene–Holocene, southern Pampa (Argentina): Evidence from palynology, mammals and sedi- mentology. Quat. Int. 2008, 181, 123–138. [Google Scholar] [CrossRef]
- Vrba, E.S. Environment and evolution: Alternative causes of the temporal distribution of evolutionary events. S. Afr. J. Sci. 1985, 81, 229–236. [Google Scholar]
- Pardiñas, U.F.J.; Cione, A.L.; San Cristóbal, J.; Verzi, D.H.; Tonni, E.P. A new interglacial continental vertebrate assemblage in central-eastern Argentina. Curr. Res. Pleistocene 2004, 21, 111–112. [Google Scholar]
- Tonni, E.P.; Huarte, R.A.; Carbonari, J.E.; Figini, A.J. New radiocarbon chronology for the Guerrero Member of the Luján Formation (Buenos Aires, Argentina): Palaeoclimatic significance. Quat. Int. 2003, 109–110, 45–48. [Google Scholar]
- Bonadonna, F.P.; Leone, G.; Zanchetta, G. Composición isotópica de los fósiles de gasterópodos continentales de la provincia de Buenos Aires. Indicaciones paleoclimáticas. In Evolución Biológica y Climática de la Región Pampeana Durante los Últimos Cinco Millones de Años; Alberdi, M.T., Leone, G., Tonni, E.P., Eds.; Monografías del Museo Nacional de Madrid: Madrid, Spain, 1995; Volume 12, pp. 77–104. [Google Scholar]
- Ameghino, F. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas Acad. Nac. Cienc. 1889, VI, 1027. [Google Scholar]
- Ameghino, F. Le Diprothomo platensis: Un precurseur de l’homme du Pliocene inférieur de Buenos Aires. An. Mus. Nac. Hist. Nat. Buenos Aires 1909, 19, 107–209. [Google Scholar] [CrossRef]
- Frenguelli, J. Rasgos generales de la morfología y la geología de la Provincia de Buenos Aires. Publ. Lab. Ens. Mater. Investig. Tecnológicas 1950, 33, 1–72. [Google Scholar]
- Frenguelli, J. Argentina de Estudios geográficos GAEA. Neozoico 1957, 2–3, 218. [Google Scholar]
- Simpson, G.G. Review of the mammal-bearing tertiary of South America. Proc. Am. Philos. Soc. 1940, 83, 649–710. [Google Scholar]
- Wood, H.E., 2nd; Chaney, R.W.; Clark, J.; Colbert, E.H.; Jepsen, G.L.; Reeside, J.B., Jr. Committee. Nomenclature and correlation of the North American continental Tertiary. Bull. Geol. Soc. Am. 1941, 52, 1–48. [Google Scholar]
- Woodburne, M.O. Definition and characterization in mammalian chronostratigraphy. J. Paleontol. 1977, 51, 220–234. [Google Scholar]
- Archibald, J.D.; Clemens, W.A.; Gingerich, P.D.; Krause, D.K.; Lindsay, E.H.; Rose, K.D. First North American land-mammal ages of the Cenozoic Era. In Cenozoic Mammals of North America; Woodburne, M.O., Ed.; University of California Press: Berkeley, CA, USA, 1987; pp. 24–76. [Google Scholar]
- Pascual, R.; Ortega Hinojosa, E.J.; Gondar, D.; Tonni, E.P. Las edades del Cenozoico mamalífero de la provincia de Buenos Aires. In Paleontología Bonaerense; Borrello, A.V., Ed.; Comisión de Investigaciones Científicas de la Provincia de Buenos Aires: Buenos Aires, Argentina, 1966; pp. 3–12. [Google Scholar]
- Savage, D.E. Cenozoic geochronology of the fossil mammals of the Western Hemisphere. Rev. Mus. Argent. Cienc. Nat. 1962, 8, 53–67. [Google Scholar]
- Simpson, G.G. The evolution of marsupials in South America. Annais Acad. Bras. Ciências 1971, 43, 103–118. [Google Scholar]
- Cione, A.L.; Gasparini, G.M.; Soibelzon, E.; Soibelzon, L.H.; Tonni, E.P. The Great American Biotic Interchange. A South. American Perspective; Springer Brief Monographies in Earth System Sciences. South America and the Southern Hemisphere; Springer International Publishing AG: Cham, Switzerland, 2015; p. 97. [Google Scholar]
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.-X. The ICS International Chronostratigraphic Chart. Episodes 2013, 36, 199–204. [Google Scholar]
- Tonni, E.P. Los mamíferos del Cuaternario de la región pampeana de Buenos Aires, Argentina. In Quaternario do Rio Grande do Sul: Integrando Conhecimentos; Ribeiro, A.M., Girardi Bauermann, S., Saldanha Scherer, C., Eds.; Monografías da Sociedades Brasileira de Paleontologia: Porto Alegre, Brazil, 2009; pp. 207–216. [Google Scholar]
- Olivares, A.I.; Verzi, D.H.; Vucetich, M.G. Definición del género Eumysops Ameghino, 1888 (Rodentia, Echimyidae) y revisión de las especies del Plioce- no temprano de Argentina central. Ameghiniana 2012, 49, 198–216. [Google Scholar] [CrossRef]
- Merino, M.; Lutz, A.; Verzi, D.; Tonni, E.P. The fishing bat Noctilio (Mam- malia, Chiroptera) in the middle Pleistocene of central Argentina. Acta Chiropterologica 2007, 9, 401–407. [Google Scholar] [CrossRef]
- Deschamps, C.M. Late Cenozoic mammal bio-chronostratigraphy in south western Buenos Aires Province, Argentina. Ameghiniana 2005, 42, 733–750. [Google Scholar]
- Cione, A.L.; Tonni, E.P. Correlation of Pliocene to Holocene southern South American and European vertebrate-bearing units. Neogene and Quaternary Continental Stratigraphy and Mammal Evolution. Boll. Soc. Paleontol. Ital. 2001, 40, 167–173. [Google Scholar]
- Tonni, E.P.; Carbonari, J.E.; Huarte, R. Marine sediments attributed to marine isotope stage 3 in the southeastern Buenos Aires province, Argentina. Curr. Res. Pleistocene 2010, 27, 154–156. [Google Scholar]
- Gasparini, G.M.; Tonni, E.P. La fauna y los ambientes en el Cuaternario de la región Pampeana. Historia Evolutiva y Paleobiogeografía de los Vertebrados de América del Sur. XXX Jornadas Argentinas de Paleontología de Vertebrados, Volumen Especial. Contrib. Mus. Argent. Cienc. Nat. Bernardino Rivadavia 2016, 6, 395–402. [Google Scholar]
- Simpson, G.G. History of the fauna of Latin America. Am. Sci. 1950, 38, 261–389. [Google Scholar]
- Patterson, B.; Pascual, R. The Fossil Mammal Fauna of South America. In Evolution, Mammals and Southern Continents; Keast, A., Erk, F.C., Glass, B., Eds.; University of New York Press: Albany, NY, USA, 1972; pp. 247–309. [Google Scholar]
- Simpson, G.G. The Curious History of South American Mammals. In Splendid Isolation; Yale University Press: New Haven, CT, USA; London, UK, 1980; pp. 1–266. [Google Scholar]
- Webb, S.D. Late Cenozoic mammal dispersals between the Americas. In The Great American Biotic Interchange; Stehli, F.G., Webb, S.D., Eds.; Plenum Press: New York, NY, USA; London, UK, 1985; pp. 357–386. [Google Scholar]
- O’Dea, A.; Lessios, H.A.; Coates, A.G.; Eytan, R.I.; Restrepo-Moreno, S.A.; Cione, A.L.; Collins, L.S.; de Queiroz, A.; Farris, D.W.; Norris, R.D.; et al. Formation of the Isthmus of Panama. Sci. Adv. 2016, 2, e1600883. [Google Scholar] [CrossRef] [Green Version]
- Domingo, L.; Tomassini, R.L.; Montalvo, C.I.; Sanz-Pérez, D.; Alberdi, M.T. The Great American Biotic Interchange revisited: A new perspective from the stable isotope record of Argentine Pampas fossil mammals. Sci. Rep. 2020, 10, 1–10. [Google Scholar]
- Webb, S.D. Ecogeography and the Great American Interchange. Paleobiology 1991, 17, 266–280. [Google Scholar] [CrossRef]
- Webb, S.D. A history of savanna vertebrates in the New World. Part II: South America and the great interchange. Ann. Rev. Ecol. Syst. 1978, 9, 393–426. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Leone, G.; Tonni, E.P. Evolución Biológica y Climática de la Región Pampeana Durante los Últimos cinco Millones de años; Monografías del Museo Nacional de Madrid: Madrid, Spain, 1995; Volume 12, pp. 1–423. [Google Scholar]
- Prado, J.L.; Alberdi, M.T. Quaternary mammalian faunas of the Pampean Region. Quat. Int. 2010, 212, 176–186. [Google Scholar] [CrossRef]
- Reguero, M.A.; Candela, A.M.; Alonso, R.N. Biochronology and biostratigraphy of the Uquía Formation (Pliocene–early Pleistocene, NW Argentina) and its significance in the Great American Biotic Interchange. J. S. Am. Earth Sci. 2007, 23, 1–16. [Google Scholar] [CrossRef]
- Woodburne, M.O.; Cione, A.L.; Tonni, E.P. Central American provincialism and the Great American Biotic Interchange. In Advances in Late Tertiary Vertebrate Paleontology in Mexico and The Great American Biotic Interchange; Carranza-Castañeda, O., Lindsay, E.H., Eds.; Publicación Especial del Instituto de Geología y Centro de Geociencias de la Universidad Nacional Autónoma de México: Mexcio City, Mexico, 2006; Volume 4, pp. 73–101. [Google Scholar]
- Nieto, M.; Hortal, J.; Martínez-Maza, C.; Morales, J.; Ortiz-Jaureguizar, E.; Peláez-Campomanes, P.; Pickford, M.; Prado, J.L.; Rodríguez, J.; Senut, B.; et al. Historical determinants of mammal diversity in Africa: Evolution of mammalian body mass distribution in Africa and South America during Neogene and Quaternary times. In African Biodiversity. Molecules, Organisms, Ecosystems; Huber, B.A., Sinclair, B.J., Lampe, K.H., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 287–295. [Google Scholar]
- Cione, A.L.; Tonni, E.P.; Soibelzon, L.H. Did humans cause large mam- mal late Pleistocene-Holocene extinction in South America in a context of shrinking open areas. In American megafaunal extinctions at the end of the Pleistocene; Vertebrate Paleobiology and Paleontology Series; Haynes, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 125–144. [Google Scholar]
- Barnosky, A.D.; Lindsey, E.L.; Villavicencio, N.A.; Bostelmann, E.; Hadly, E.A.; Wanket, J.; Marshall, C.R. Variable impact of late-Quaternary megafaunal extinction in causing ecological state shifts in North and South America. Proc. Natl. Acad. Sci. USA 2016, 113, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnosky, A.D.; Lindsey, E.L. Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quat. Int. 2010, 217, 10–29. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P.; Soibelzon, L. The broken zig-zag: Late Cenozoic large mammal and turtle extinction in South America. Rev. Mus. Argent. Cienc. Nat. Bernardino Rivadavia 2003, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.S. Prehistorc Overkill: The Global Model. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 354–403. [Google Scholar]
- Firestone, R.B.; West, A.; Kennett, J.P.; Becker, L.; Bunch, T.E.; Revay, Z.S.; Schultz, P.H.; Belgya, T.; Kennett, D.J.; Erlandson, J.M.; et al. Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and the Younger Dryas cooling. Proc. Natl. Acad. Sci. USA 2007, 104, 16016–16021. [Google Scholar] [CrossRef] [Green Version]
- Barnosky, A.D.; Bell, C.J.; Emslie, S.D.; Goodwin, H.T.; Mead, J.I.; Repenning, C.A.; Scott, E.; Shabel, A.B. Exceptional record of mid-Pleistocene vertebrates helps differentiate climatic from anthropogenic ecosystem perturbations. Proc. Natl. Acad. Sci. USA 2004, 101, 9297–9302. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio, N.A.; Lindsey, E.L.; Martin, F.M.; Borrero, L.A.; Moreno, P.I.; Marshall, C.R.; Barnosky, A.D. Combination of humans, climate, and vegetation change triggered Late Quaternary megafauna extinction in the Última Esperanza region, southern Patagonia, Chile. Ecography 2016, 39, 125–140. [Google Scholar] [CrossRef]
- Steele, J.; Politis, G. AMS 14C dating of early human occupation of southern South America. J. Archaeol. Sci. 2009, 36, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Borrero, L.A. The elusive evidence: The archeological record of the South American extinct megafauna. In American Megafaunal Extinctions at the End of the Pleistocene; Haynes, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 145–168. [Google Scholar]
- Politis, G.G.; Prado, J.L.; Beukens, R.P. The human impact in Pleistocene–Holocene extinctions in South America. The Pampean case. In Ancient Peoples and Landscapes; Johnson, E., Ed.; Museum of Texas Tech University: Lubbock, TX, USA, 1995; pp. 187–205. [Google Scholar]
- Gutiérrez, M.; Martínez, G. Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean Region (Argentina). Quat. Int. 2008, 191, 53–68. [Google Scholar] [CrossRef]
- Prates, L.; Politis, G.; Steele, J. Radiocarbon chronology of the early human occupation of Argentina. Quat. Int. 2013, 301, 104–122. [Google Scholar] [CrossRef]
- Prates, L.; Politis, G.G.; Perez, S.I. Rapid radiation of humans in South America after the last glacial maximum: A radiocarbon-based study. PLoS ONE 2020, 15, e0236023. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.D. Megafauna biomass tradeoff as a driver of Quaternary and future extinctions. Proc. Natl. Acad. Sci. USA 2008, 105, 11543–11548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen-Smith, R.N. Pleistocene extinctions: The pivotal role of megaherbivores. Paleobiology 1987, 13, 351–362. [Google Scholar] [CrossRef]
- Graham, R.W.; Lundelius, E.L. Coevolutionary disequilibrium and Pleistocene extinctions. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 223–249. [Google Scholar]
- Guthrie, R.D. Mosaics, allelochemics and nutrients. An ecological theory of late Pleistocene Megafaunal Extinction. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 259–298. [Google Scholar]
Figure 1.
Location of the Pampean Region in Argentina.


Figure 3.
Four taxa from Argentinian Pampa that define the biozones. 1, Skull in lateral view of Equus neogeus; 2, Skull in lateral view of Mesotherium cristatum; 3, Femur of Megatherium americium; 4(a,b), Skull in ventral view and mandible in lateral view of Ctenomys chapalmalensis.
Figure 3.
Four taxa from Argentinian Pampa that define the biozones. 1, Skull in lateral view of Equus neogeus; 2, Skull in lateral view of Mesotherium cristatum; 3, Femur of Megatherium americium; 4(a,b), Skull in ventral view and mandible in lateral view of Ctenomys chapalmalensis.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The main stratigraphic schemes proposed for the Pampean Region.
| Doering (1882) | Ameghino (1908) | Kraglievich (1952) | Frenguelli (1957) | Fidalgo et al. (1973) | Riggi et al. (1986) | |
|---|---|---|---|---|---|---|
| Fm Querandina | Post pampeano | Platense | Fm de Loberia | Serie postpampeano | Fm La Postrera/Fm Luján | |
| Fm Tehuelche | Lujanense | Fm Buenos Aires | ||||
| Bonaerense | Fm de Arroyo Seco | Bonaerense | ||||
| Fm Pampeana | Ensenadense | Fm de Miramar | ||||
| Fm de San Andrés | Ensenadense | |||||
| Fm Pampeano | Fm Ensenada | |||||
| Fm de Vorohué | Fm Pampiano | |||||
| Fm de Barranca de los Lobos | ||||||
| Fm Araucano | Chapadmalense | Fm de Chapadmalal | Chapadmalense | |||
| Monte-hermosense | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prado, J.L.; Alberdi, M.T.; Bellinzoni, J. Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications. Quaternary 2021, 4, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020015
AMA Style
Prado JL, Alberdi MT, Bellinzoni J. Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications. Quaternary. 2021; 4(2):15. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020015
Chicago/Turabian StylePrado, José Luis, María Teresa Alberdi, and Jonathan Bellinzoni. 2021. "Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications" Quaternary 4, no. 2: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020015