
Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau


1
School of Natural Sciences, Private Bag 55, University of Tasmania, Hobart, TAS 7001, Australia
2
Department of Primary Industries, Parks, Water and Environment, Hobart, TAS 7001, Australia
*
Author to whom correspondence should be addressed.
Fire 2021, 4(3), 32; https://0-doi-org.brum.beds.ac.uk/10.3390/fire4030032
Submission received: 4 May 2021
/
Revised: 13 June 2021
/
Accepted: 19 June 2021
/
Published: 24 June 2021
(This article belongs to the Special Issue Bushfire in Tasmania)
Abstract
:Australian montane sclerophyll shrubland vegetation is widely considered to be resilient to infrequent severe fire, but this may not be the case in Tasmania. Here, we report on the vegetative and seedling regeneration response of a Tasmanian non-coniferous woody montane shrubland following a severe fire, which burned much of the Great Pine Tier in the Central Plateau Conservation Area during the 2018–2019 fire season when a historically anomalously large area was burned in central Tasmania. Our field survey of a representative area burned by severe crown fire revealed that more than 99% of the shrubland plants were top-killed, with only 5% of the burnt plants resprouting one year following the fire. Such a low resprouting rate means the resilience of the shrubland depends on seedling regeneration from aerial and soil seedbanks or colonization from plants outside the burned area. Woody species’ seedling densities were variable but generally low (25 m). The low number of resprouters, and reliance on seedlings for recovery, suggest the shrubland may not be as resilient to fire as mainland Australian montane shrubland, particularly given a warming climate and likely increase in fire frequency.
1. Introduction
Shrubs, defined here as woody plants less than 3 m high, have often been considered as grassland and woodland invaders, contributing to woody thickening. Shrub invasion can be indicative of declining rangeland productivity because they are unpalatable to livestock or low in feed value [1,2,3]. However, shrublands provide important ecosystem functions and services, such as wildlife habitat and food resource, sizeable carbon sinks, prevention of soil erosion, and hydrological regulation [2,4,5,6]. These two opposing points of view of shrublands affect how natural and anthropogenic wildfires are viewed. If shrubs are viewed as nuisance invaders, then planned and unplanned fires can be used as an important management tool in their control [7,8,9,10]. Alternatively, if shrublands are an important component of the landscape then fire management is critical. Both cases demand understanding of the resilience (capacity to return to its pre-disturbance state [11]) of shrublands to fire. Climate change complicates fire management in shrublands. Shrubs can be one of the most flammable components within a montane landscape, and under a warming climate shrub encroachment into grasslands may increase [12,13]. Further, severe wildfires are expected to become more frequent with a rapidly warming climate [14,15,16].
Life-history traits affect the dynamics of plant populations and vegetation communities following disturbances, such as fire [17,18,19,20]. In the case of plant populations top-killed by fire, regeneration can occur in three ways: resprouting from dormant buds (apical, epicormic, or basal); by seed germination; or colonization from elsewhere [21]. Trade-offs exist between resprouting and investment in seedling regeneration [22,23], and these trade-offs are also controlled by fire severity and fire intensity. Fire intensity is defined as the amount of energy released during a fire per unit time [24], whereas fire severity is defined as the biological impact of combustion, affecting ecosystem structure and function [24]. Fire severity is often, but not always, strongly correlated with fire intensity. Obligate post-fire seeder species are typically killed by all but the lowest severity fires and regenerate from aerial or soil seedbanks, whereas post-fire resprouters typically rapidly recover vegetatively from all but the most severe fires [21]. Following severe fire, obligate seeder species will require time for seed to germinate, grow and mature and produce the next batch of seed. If fire recurs before this cycle is completed, these species face a significant risk of local extirpation; this risk is exacerbated if climate change also reduces growth rates [25].
Understanding responses of shrub species to fire is key to managing vegetation dynamics on the Central Plateau of Tasmania, which features an extensive montane-alpine shrubland/grassland mosaic. Increases in fire severity and frequency due to a warming climate pose risks to ecosystems where historically fires may have been rare [12,14,26]. The Central Plateau is such a system, with increasing risk of severe fire [15] threatening the montane-alpine woodland, shrubland and grassland communities that characterize the region [27]. Historical records suggest that ’fire-stick’ farming was practiced by indigenous people and European settlers adopted these practices to maintain grassy vegetation for forage for domestic stock [28,29,30]. There is anecdotal evidence that the taller woody shrubs have increased in density following the cessation of this practice (Harold Riley, personal communication). Furthermore, following European settlement, the area was heavily grazed by cattle and sheep [31] until around 1989, when grazing was banned within the Central Plateau Conservation Area (CPCA) [32]. It is possible that burning by graziers suppressed shrubs and maintained grassiness, and with the cessation of livestock grazing and associated burning, shrubs have now encroached. Alternatively, overgrazing and frequent burning by graziers created bare soil and favored establishment of unpalatable shrubs, requiring graziers to burn more, creating a positive feedback between burning and shrubs [33,34,35]. If the Central Plateau shrubland is in fact invaded grassland, management burning may be warranted to control the shrubs and allow the grassland to re-establish, in conjunction with the continued exclusion of livestock grazing.
The fire season of 2018–2019 was particularly intense in Tasmania burning over 210,000 ha [36]. In February 2019, a wildfire severely burned areas of the eastern CPCA, destroying almost all above ground biomass. This fire provided an excellent opportunity to better understand the resilience to fire of the CPCA shrubland in the absence of livestock grazing. With the exception of deciduous heath and coniferous heath communities, many Tasmanian montane species are thought to be able to regenerate following infrequent fires [37], but there is little empirical data on the response to fire of Tasmanian non-coniferous montane shrub communities. Our field research focused on the resprouting response of woody shrubs within the CPCA to fire. In particular, we characterized the fire severity using both satellite imagery and ground-based methods. We then investigated how many species resprouted after fire, and whether resprouting rates were affected by fire severity and/or shrub size. We also measured post-fire seedling recruitment, to determine whether fire severity influenced the density of overall seedling recruitment, one-year post-fire.
2. Materials and Methods
2.1. Study Region
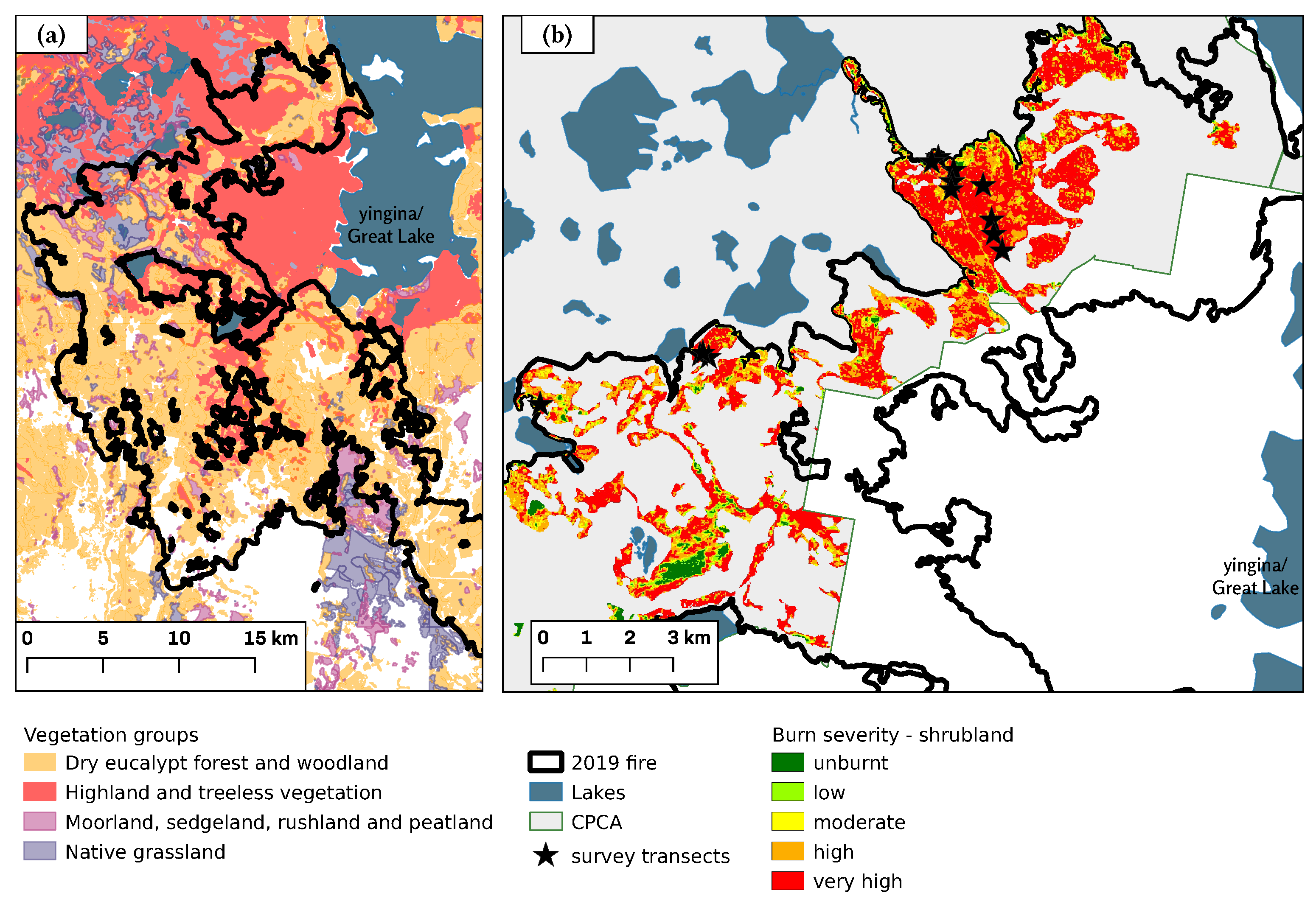
Our study area was the montane shrublands within the CPCA, near Lake Augusta, to the west of yingina/Great Lake, on Tasmania’s Central Plateau at an elevation of approximately 1100 m (Figure 1 and Figure 2). The CPCA covers an area of 94,510 ha and is part of the Tasmanian Wilderness World Heritage Area (TWWHA). In contrast to many other alpine and subalpine areas globally that are dominated by grasses and herbs [38], approximately 55% of the CPCA is classed as highland and treeless vegetation, of which 71% (36,600 ha) is scleromorphic alpine heathland and less than 1% is grassland (Figure 2) [39].
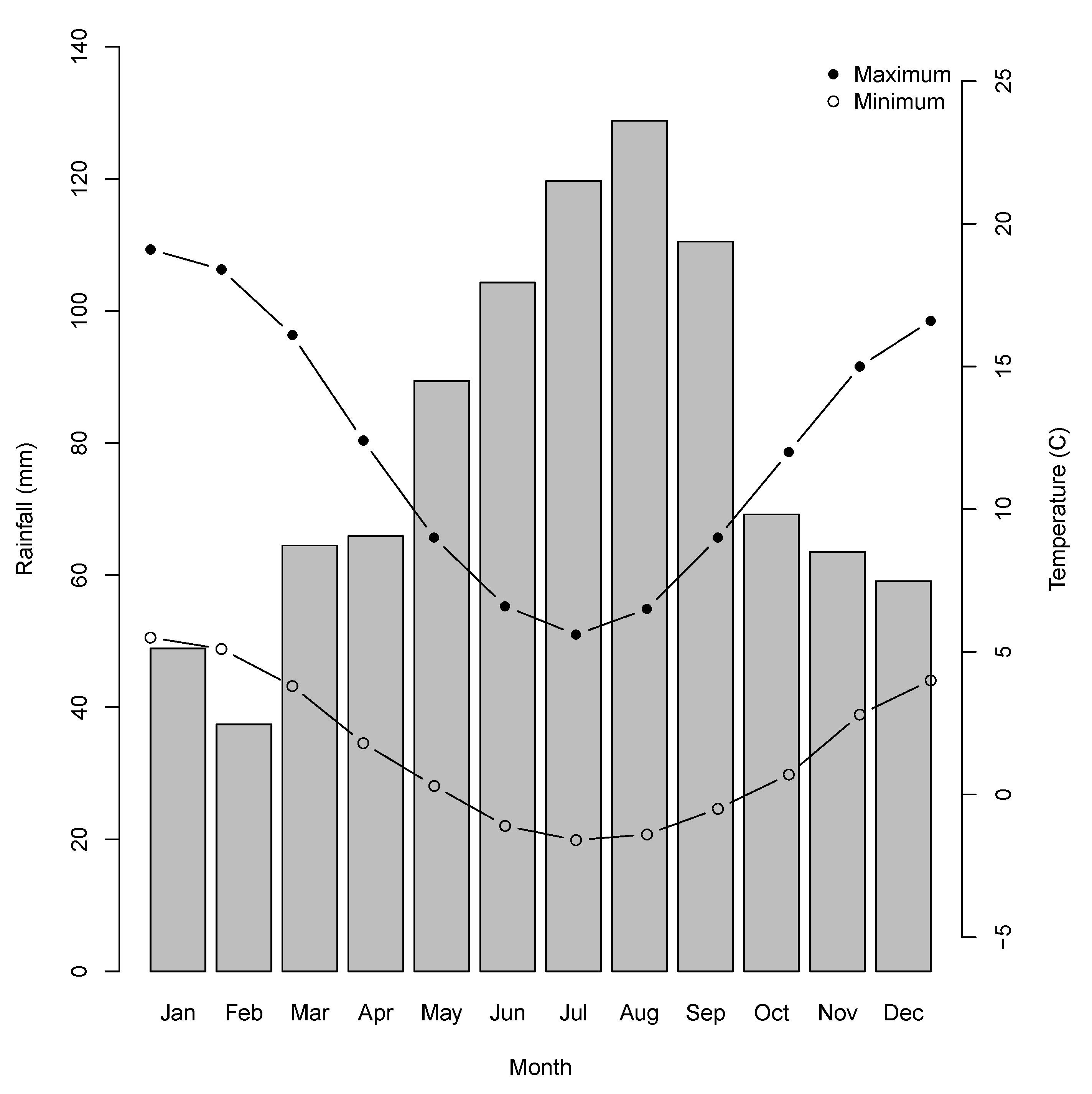
The Central Plateau was extensively glaciated during the Last Glacial Maximum, which has left a legacy of myriad small lakes and tarns. The underlying geology is predominantly Jurassic dolerite [32,40]. The climate of the Central Plateau region is perhumid cold to humid cold, with heavy rainfall in the Western Plateau region becoming lighter towards the east [41]. Snow falls frequently during the winter months but rarely lies on the ground for extended periods [41,42]. At Liawenee, the nearest weather station to Lake Augusta, mean monthly maximum temperatures range from 5.6 °C in July to 19.1 °C in January, and the mean minimum monthly temperature ranges from −1.6 °C in July to 5.5 °C in January (Figure 3), but minimum temperatures below zero can occur during any month of the year [43]. The mean annual precipitation is 923 mm, with the wettest months occurring during winter (Figure 3). In the 2018 calendar year precipitation was 998 mm, but January 2019 (the month in which the fire started) was very dry, with only 4 mm rain, compared with an average for January of 50 mm. The twelve-month period following the 2019 fire was slightly drier than average, with a total 771 mm precipitation [43].
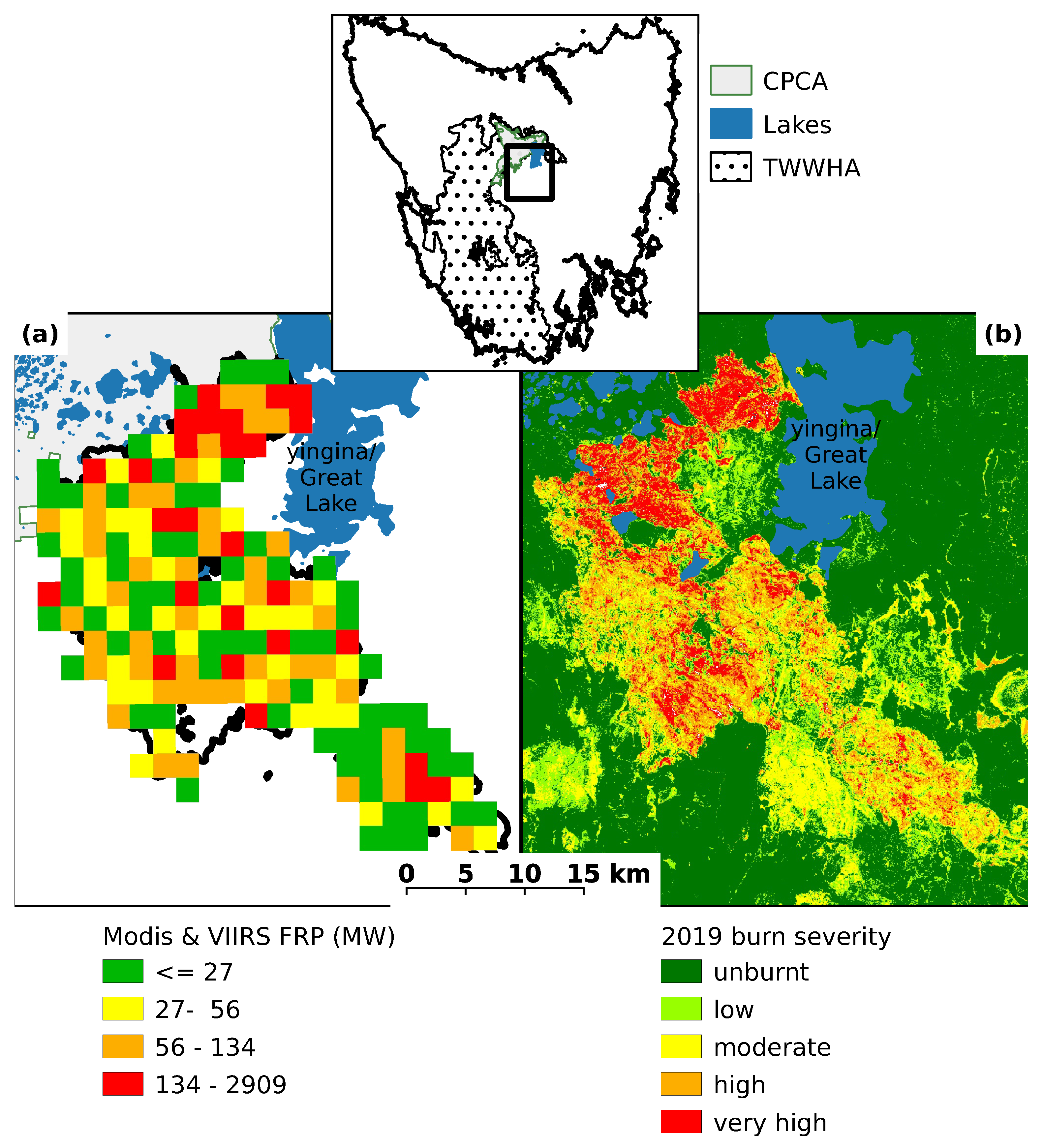
Although the CPCA is likely to have experienced many small fires from burning grasslands for stock grazing, there are no historical records of large fires within the region until 1960/61, when some 57,500 ha was burnt, purportedly by highland graziers to enhance forage quality [29]. Little is known about the ecological effects of the 1961 fire on the montane shrubland. The 2018–2019 summer in Tasmania was the second warmest on record with many areas of the state experiencing very little rain from late December to early February resulting in extreme dryness, including in areas that would normally be too moist to burn [36]. On 15 January 2019, dry lightning ignited several small fires which combined to form the Great Pine Tier fire. Over a period of three weeks, the fire burnt 51,224 ha, of which 10,094 ha was within the TWWHA. The CPCA shrubland burnt between 29 January and 3 February 2019 [36] (Figure 1), and MODIS hotspot data for this period were downloaded from the MODIS website. Values for fire radiative power (FRP), a proxy for fire intensity, were calculated from the MODIS and VIIRS active fire products [44,45]. FRP for the Great Pine Tier fire varied between 0 and 2909 MW, and averaged 142 MW (Figure 1a), which is substantially less than the average 362 MW reported for fire in sedge heathland in south-west Tasmania in 2013 [46]. The CPCA shrubland was severely impacted by the fire, with approximately 84% of the shrubland area being burnt by a high (26%) or very high (58%) severity fire (as measured by the differenced Normalized Burn Ratio (dNBR)). By comparison, approximately 59% of the total area burnt at a high severity (38%) or very high severity (21%) (Figure 1b and Table 1).
2.2. Field Survey
The field survey took place during February and March 2020, 12 months after the 2019 Pine Tier fire. Eleven traverses were made of the burnt shrubland with measurements made within variable length belt plots established at approximately 50-m intervals along each traverse (Figure 2b). Each traverse was located in easily accessible areas where the presence of burnt stems indicated that pre-fire the vegetation consisted of woody shrubs rather than woodland or grassland. Each plot was at a right angle to the main traverse, with a width of 1, 2, or 4 m and ranging in length between 1.2 and 39.7 m in order to capture between 10 and 20 woody shrubs per plot, depending on the local shrub density. All plots were placed at least 50 m from the Lake Augusta Road, and the starting point for each plot was determined by haphazardly dropping a stick. In total, we established 55 plots in the burnt area and 5 plots in unburnt shrubland just outside the fire boundary. The pre-fire similarity of vegetation in burnt and unburnt plots was determined by inspection of high-resolution aerial imagery captured before the 2019 fire, on-ground assessment of vegetation present post-fire and local environment (topography, soil, hydrology), as well as on the basis of the authors’ familiarity with the study area. Such side-by-side comparisons of adjoining burned and unburned area are a pragmatic method to contextualize the effects of fire [47]. GPS coordinates of the start of each plot were recorded using the Handy GPS Application, Version 34.4 (www.binaryearth.net, accessed on 18 October 2019), operating on an android smart phone.
Initially, all woody plants were recorded, but, for the later plots, only those plants with at least one stem with a basal diameter ≥ 2 cm were recorded. Within each plot, all woody shrubs were identified to species level where possible. Where a group of stems were growing close together, they were considered to be part of the same plant if they appeared to be connected underground. For each plant, we recorded whether it was burnt or not, condition (alive, dead, or resprouting), number of stems, and width and length of the lignotuber, from which we calculated shrub basal area. As a proxy for fire severity, we measured the diameter (to the nearest mm) of the smallest burnt twig. One measurement was recorded for each plant within the plot. Larger minimum burnt twig measurements indicate a more severe fire [48,49] and have been well correlated with survival of a fire-sensitive Tasmanian conifer [50]. Minimum burnt twig measurement was recorded as ‘0’ for all plants that had an unburnt canopy, including the occasional unburnt plant within the burnt plots. For those plants resprouting, the number of resprouting shoots, and height of tallest resprout was recorded. For plants within the unburnt plots, the presence or absence of a canopy seedbank was noted.
Three 50 × 50 cm quadrats were established at the beginning, center, and end of each belt plot. We counted the number of seedlings present in each quadrat. We were not able to identify species due to the very small size of the seedlings. Within each quadrat, the total number of scats and type of animal (macropod, wombat, rabbit, deer) were also recorded. Plots were characterized by visually estimating the percentage of rock, forbs, grasses, and bare soil for a 1 m quadrat located at the center of the belt plot. Due to the very open and flat terrain of the study site, we did not record slope or aspect.
Travel restrictions due to Covid-19 were imposed towards the end of the planned field work, so we measured fewer unburnt plots than planned. In addition, we observed resprouting Olearia myrsinoides shrubs but were unable to survey populations of this species or include them in the results.
2.3. Data Analysis
We compared our percentage estimate of environmental variables in burned and unburned quadrats using t-tests, after confirming that the data were normally distributed and variances equal. In the burnt plots, most stems were largely consumed by the fire. Therefore, we used the shrub basal area (calculated as length x width occupied by the shrub stems at ground level, thereby incorporating below ground lignotuber or rhizome) and number of stems per plant as indicators of plant size, which would be loosely correlated with shrub age.
Orites revoluta was the only species in our survey plots that resprouted in sufficient numbers for analysis. The resprouting response (whether dead or resprouting) of burnt O. revoluta shrubs was analyzed using a binomial generalized linear model (glm) with resprouting as a binary response variable and fire severity (represented by burnt twig diameter at the plant level) and shrub basal area as explanatory variables.
We investigated the relationship between seedling density and fire severity by comparing Poisson, zero-inflated Poisson and negative binomial glms using average minimum burnt twig diameter per plot as a continuous predictor variable. This plot level fire severity proxy, calculated by averaging burnt twig measurement for all burnt plants within the plot, was used as a fire severity proxy because seedling data were recorded at the plot level. We also tested whether there was a humped response to fire severity by comparing linear models with those that also included a quadratic (twig diameter squared) term. Vuong tests and AIC were used to select the best model [51,52].
All data exploration and statistical analysis was undertaken using R statistical software (version 3.6.3) [53].
3. Results
3.1. Site Condition
The percentage of bare ground, rock, and vegetation cover varied considerably between the plots, although there was no significant difference between the mean cover of the burnt plots compared to unburnt according to t-tests. For the burnt plots, bare ground ranged from 0% to 80%, with a mean of 28%, and rock cover ranged from 0% to 90% with a mean of 19%. Forbs and grass cover in the burnt plots ranged from 0% to 80% and 70%, respectively, with means of 15% and 9%. Scats (macropod, rabbit, wombat, and deer) were present in most plots surveyed (n = 49; 82%). Most scats were native macropods, with feral fallow deer and introduced rabbit scats recorded in only 9 plots and 1 plot, respectively. Only macropod scats were present in the unburnt plots (Table A1).
3.2. Plant Response
Of the 1036 woody shrubs surveyed in total, 92% (952) had been burnt, with more than 99% (946) of the burnt plants top-killed (Table 2 and Figure 4c,d). By contrast, most plants (94%) in the unburnt plots were alive (Table 2). Within the burnt plots, the level of resprouting was low, with only 5% (43) of the burnt shrubs observed to be resprouting from basal buds.
We observed resprouting in three species (Orites revoluta, Leptospermum rupestre and Bauera rubioides) (Figure A1) in our study plots (in addition to Olearia myrsinoides, observed outside our survey plots). Of the 90 plants from the three species observed to resprout, 48% (43) were resprouting, and 93% (40) of the resprouters were O. revoluta (Table 2). The other resprouters were 2 L. rupestre shrubs (representing 17% of burnt L. rupestre) and the sole observed B. rubioides shrub (Figure A1).
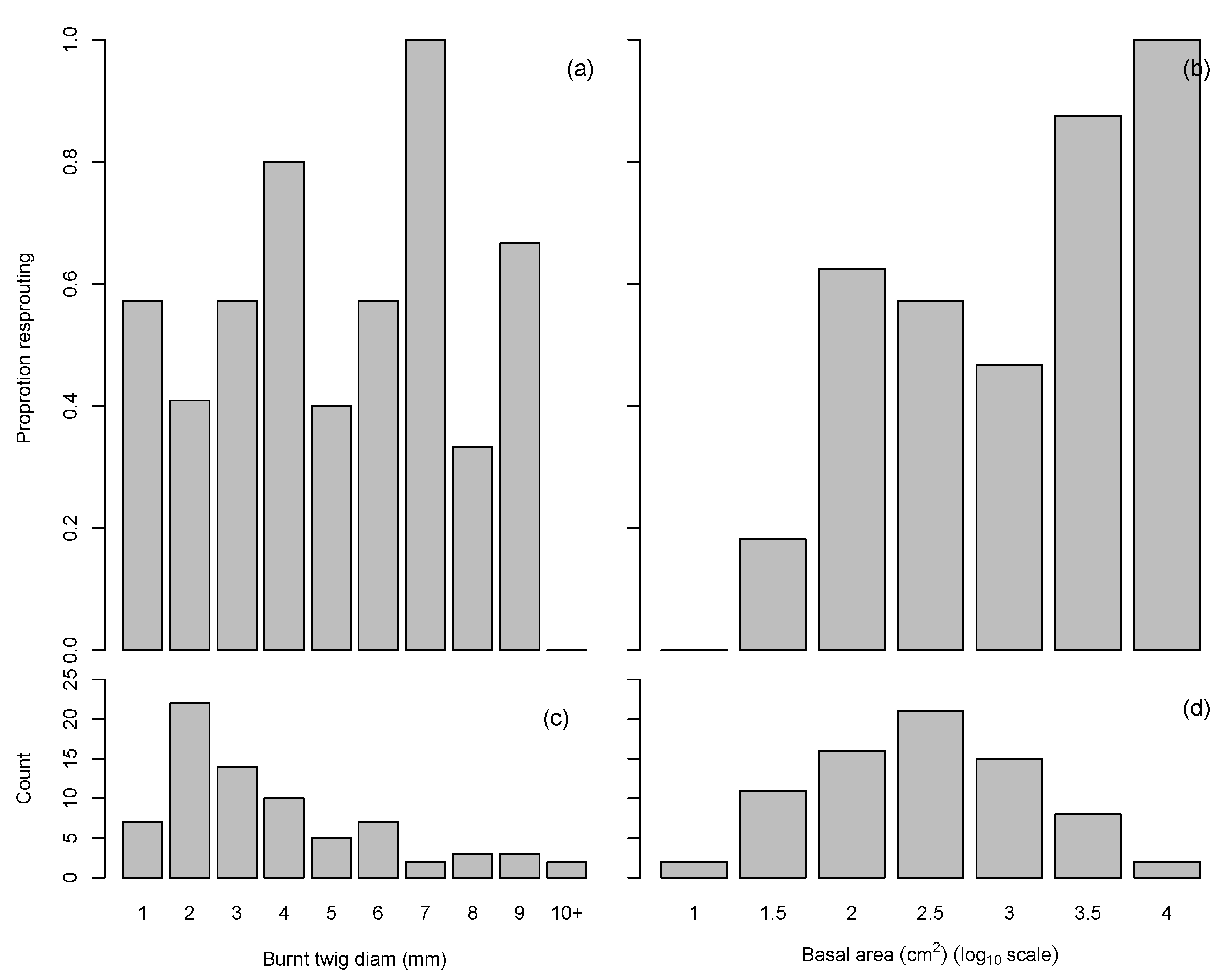
Of the 75 O. revoluta plants that were burnt, 53% (40) were resprouting (Table 2). There was no significant relationship between fire severity and resprouting of O. revoluta (Figure 5), although this may be due to low sample size at the higher fire severities. There was a significant positive relationship (p < 0.01) between the proportion of O. revoluta plants resprouting and shrub basal area (Figure 5b,d). Resprouting was prolific in the surviving O. revoluta plants (Figure A1a). The median number of resprout shoots per resprouting plant was 21, with three plants having at least 100 resprout shoots. The height of the tallest resprout shoot ranged from 3 cm to 90 cm, with a median of 14 cm.
3.3. Seedling Recruitment
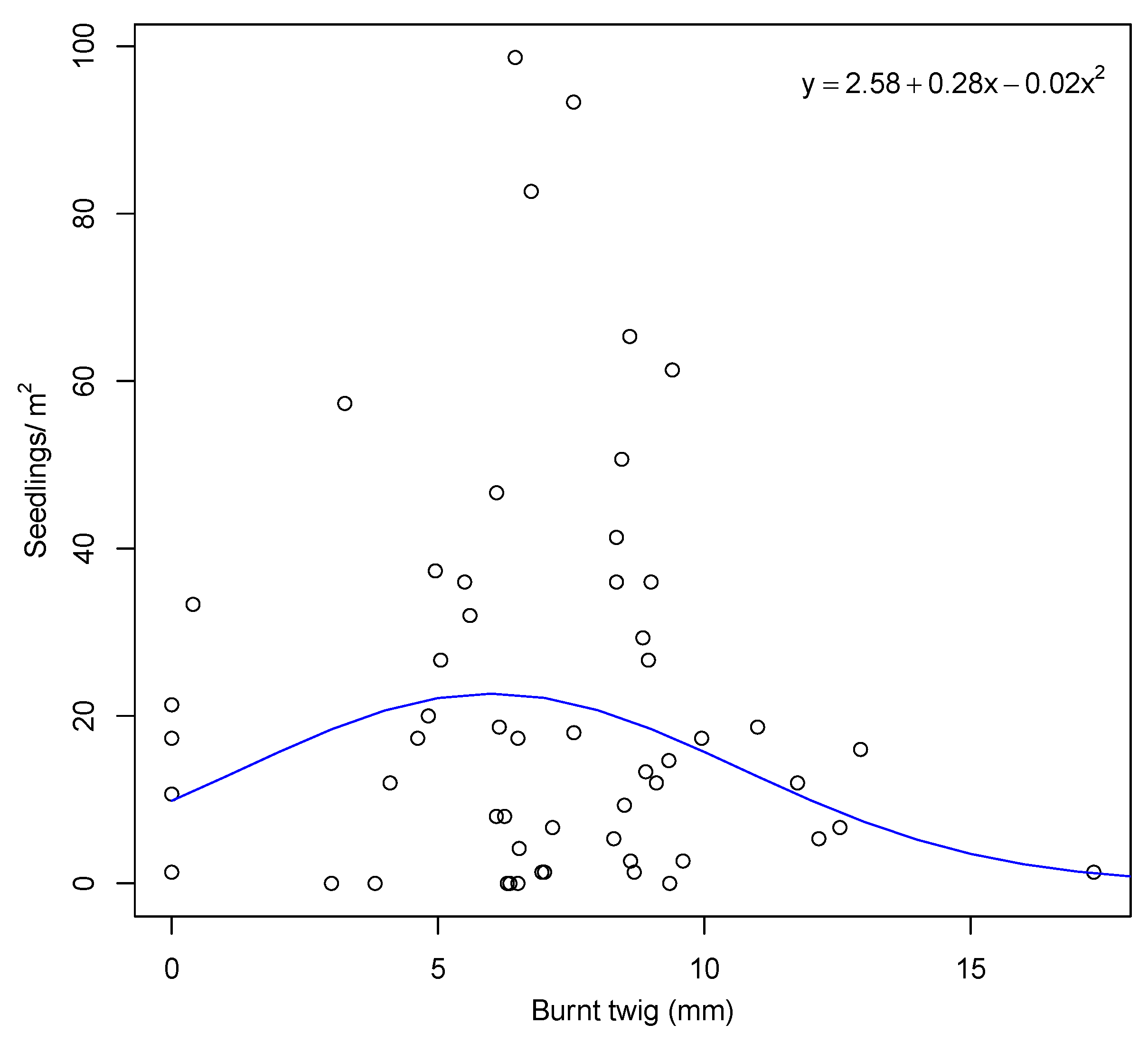
Seedlings were present in all unburnt plots, and most plants in the unburnt plots had a canopy seedbank at the time of the survey. Seedlings were absent in nine of the burnt plots (15%), and most of these were in plots burnt at lower than average fire severity. Across all plots, seedling density was very variable, ranging from zero up to a maximum density of 242 m, with a mean density of 25 m. The best model between seedling density and fire severity was the negative binomial quadratic (AIC = 465), followed by zero-inflated poisson quadratic (AIC = 1255) and poisson quadratic regression (AIC = 1680). This shows a humped relationship, with higher seedling density occurring under intermediate fire severity conditions (Figure 6).
4. Discussion
Our study contributes to understanding the fire ecology of Tasmanian montane shrublands which are rarely burned, and provides a record of some of the impacts of the anomalous 2018–2019 fire season that impacted several regions within the Tasmanian Wilderness World Heritage Area including the Central Plateau Conservation Area (CPCA). We found montane shrubland in central Tasmania is largely comprised of post-fire obligate seeder species and the shrubland was severely burnt by the 2019 Great Pine Tier fire. It is unknown if the fire regimes for this shrubland community are restricted to such stand replacing crown fire or also include mixed, or low severity fires. Despite our sampling design that sought to be representative of burned shrublands, we acknowledge that our survey plots were slightly biased against the low and moderate fire severities, but high or very high severity fire predominated in both our survey plots (87%) and the CPCA shrubland (84%). High consumption of shrub biomass by surface fires is common, with a study of 38 fires in temperate Australia finding a median 80% consumption [47]. Overall post-fire resprouting in our survey was very low, with only 5% of all burnt plants, and only three species, observed to resprout. Such a low resprouting rate is similar to the 7.5% found by Reference [54] following a severe fire in shrubby and heathy coastal vegetation on Clarke Island, off the north east coast of Tasmania, but less than the 16% observed by Reference [46] in heath sedgeland in south-west Tasmania. However, in both the Nicholson and French studies, a higher proportion of recorded species had at least one plant resprouting (approximately 60% and 80%, respectively, compared with only 38% in the current study). The 38% of species resprouting in the current study was also substantially less than the 85% observed in subalpine shrubland in Kosciuszko National Park, New South Wales, following the 2003 fires [55]. The observed resprouter rate is also lower than the 53% resprouting response for woody shrubs that has been recorded in the New England Tableland Bioregion in central eastern Australia [56] and up to 80% in southwest Western Australia [57].
The dominant resprouter species, O. revoluta, was a vigorous resprouter, with individual plants sometimes producing more than 100 basal shoots. There was also a higher proportion of O. revoluta individuals resprouting than observed by Reference [54] for any species on Clarke Island. This highlights the variability among species in the vigour and consistency of the resprouting response, which can vary with factors, such as plant size, and the type and severity of stress [58,59]. Interestingly, O. revoluta did not display the apparent humped relationship between resprouting rates and fire severity reported by Reference [54], but we encountered too few individuals that experienced high severity fire to be confident that resprouting rates were maintained at these more extreme severities. It is possible that more species could have resprouted with fire that is less severe than occurred in 2019. For instance, this could be why we found no instances of resprouting in Ozothamnus hookeri, which is reported to be an early successional resprouter that produces large quantities of wind-dispersed seed [60]. It has also been suggested that pre-fire drought combined with a high severity fire may significantly reduce resprouting success [61]. The rainfall records for Liawenee do not show an unusually prolonged dry period immediately prior to the 2019 fire, so short-term drought stress is unlikely to explain the lack of resprouting. However, carbohydrate reserves of resprouters may have declined over a longer-term dry trend reported for this region, which is known to be affecting the population health of some woody plants [62,63].
Given the weak resprouting response following the 2019 fire, recovery of the pre-fire shrubland will depend on regeneration from seed, which, in this region, can be precarious. The main germination period for seedlings is autumn, but the harsh winter environment on the Central Plateau, and particularly frost heave events, result in substantial seedling losses [32]. Therefore, woody shrubs tend to establish in bare ground in the less exposed sites, such as under existing vegetation cover [32,64,65]. Additionally, the size of the bare ground gaps can influence regeneration, with smaller gaps favoring grasses and larger gaps favoring woody shrub seedlings [66]. The other main threat to seedling survival is herbivory [64,67,68,69]. Fire can result in increased macropod densities [70]. In addition to native marsupials, other important herbivores are the introduced rabbits [30,31,64] and feral fallow deer [71]. We found scant evidence of rabbit or deer presence, but macropod scats were present in most survey plots, suggesting that, in our survey area, the dominant herbivory pressure immediately post-fire came from native herbivores.
The low percentage of species that we found resprouting supports the findings of Reference [72] that Tasmanian montane flora recovers very slowly, and appears less resilient to infrequent severe fire than mainland Australian montane flora. The low rate of resprouting, together with the patchy seedling regeneration observed, indicates that recovery of this shrubland will likely take a lot longer than the 8 years postulated by Reference [73] for recovery of the alpine Bogong High Plains in Victoria, Australia. The marked reduction we observed in live shrub cover is likely to persist for many years, given slow growth rates in this area: twelve months after the fire, median resprout height was 14 cm, and, at this rate, it would take at least eight years for the few surviving shrubs to attain the height of those in unburnt areas. We have observed high grass cover in the burnt areas, consistent with findings of Reference [72], that, in burnt alpine coniferous heath, there was an increase in grass and herb cover, which persisted for about 30 years, thereby increasing the risk of subsequent fire.
With only one resprouting species (O. revoluta) currently dominating the shrub regeneration in our study area, it will be interesting to see whether the fire causes long term reductions in the woody shrub diversity of the shrubland. Identification of shrub seedlings will be needed to develop a detailed picture of how the floristics of this shrubland will change following the fire. A longer-term study focusing on regeneration of species over time would improve our understanding of the impact of the 2019 fire on the vegetation dynamics of the shrubland, allowing better informed management decisions to be made.
The dominance of obligate seeders within the shrubland may mean this community has reduced resilience in response to anthropogenic climate change and associated increased frequency of droughts and fires [74]. Under warmer, drier climate conditions plants may grow more slowly and produce fewer seeds, and therefore require longer periods between fires to recover biomass and re-establish a seedbank. Additionally, unfavorable weather conditions, such as drought and/or frosts, following a severe fire may reduce post-fire recruitment. At the same time, a drier, warmer climate is likely to lead to increased fire frequency. The combination of shorter fire intervals and slower growth rates caused by a warming, drying climate has been termed interval squeeze by Reference [25] and poses a particular risk for obligate seeder systems [74], such as the CPCA shrubland. This could lead to a shift from shrubland to grassland [75,76,77], that combined with a corresponding increase in fire frequency, may become an irreversible state change.
Woody encroachment by shrubs and small trees has been reported in many areas of the world. The major underlying cause appears to be increased grazing pressure from domestic animals reducing the cover and continuity of flammable grasses, which reduces fire frequency and severity, enhancing survival of woody plants, which are generally less palatable than grasses [34,78,79,80]. It is possible that this process has led to an expansion and/or thickening of shrub cover during the decades when sheep-grazing occurred on the Central Plateau. However, this study is unable to advance this question of whether the current shrubland distribution is a legacy of past fire and grazing since European settlement, including the disruption of Aboriginal fire management regimes. Further research needs to focus on historical land cover changes using both palaeoecological and historical techniques, which have been applied in other landscape settings in Tasmania (e.g., References [81,82]), as well as contrasting post-fire recovery of montane shrublands that are still subjected to livestock grazing with those where livestock grazing does not occur.
In conclusion, the 2019 Great Pine Tier fire that swept through the Central Plateau Conservation Area in February 2019 was a high severity crown fire that killed almost all above ground vegetation in the shrubland. Only three out of eight species identified as being present prior to the fire resprouted in the 12 months following the fire, and only one, Orites revoluta, exhibited a strong resprouting response. In the absence of additional resprouting, regeneration of this shrubland will rely on seedling recruitment or colonization and low fire frequency to enable seedlings to mature. It is unclear if the pre-fire shrubland resulted from the combined effects of frequent fire and intense stock grazing on natural grasslands. If this can be shown to be the case, then more frequent fires may be beneficial in reducing shrub cover and restoring grassland. Conversely, fires that are too frequent could endanger populations of the predominantly obligate seeder shrub species found in the area, requiring minimizing fire frequency in order to assure long term persistence of these species. Understanding past trends in shrub cover and fire activity will assist managers in implementing ecologically appropriate fire regimes.
Author Contributions
Conceptualization, D.M.J.S.B. and S.W.J.L.; methodology, D.M.J.S.B., J.A.F., L.D.P., and S.W.J.L.; formal analysis, J.A.F.; writing—original draft preparation, J.A.F. and L.D.P.; writing—review and editing, J.A.F., L.D.P., D.M.J.S.B., and S.W.J.L.; supervision, D.M.J.S.B.; funding acquisition, D.M.J.S.B. and S.W.J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a quick response grant from the Bushfire & Natural Hazards CRC, and the Professor Newton Barber Honours Scholarship in Plant Science from the University of Tasmania awarded to J.A.F.
Data Availability Statement
The data presented in this study are openly available in FigShare at 10.6084/m9.figshare.14524371.
Acknowledgments
We thank Mary McVarish and Ross Corkrey for assistance in the field, and Ross Corkrey for additional statistical advice.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Scat presence and number within survey plots.
| Animal | Number of Plots | Number of Scats | ||
|---|---|---|---|---|
| Burnt | Unburnt | Burnt | Unburnt | |
| Macropod | 39 | 5 | 217 | 105 |
| Wombat | 3 | 0 | 16 | 0 |
| Deer | 9 | 0 | 18 | 0 |
| Rabbit | 1 | 0 | 21 | 0 |
| Total | 272 | 105 | ||
Figure A1.
Resprouting shrubs. (a) Orites revoluta from large basal lignotuber; (b) Bauera rubioides and (c) Leptospermum rupestre. Photographs by J. Foulkes.
Figure A1.
Resprouting shrubs. (a) Orites revoluta from large basal lignotuber; (b) Bauera rubioides and (c) Leptospermum rupestre. Photographs by J. Foulkes.

References
- McArthur, E.D.; Kitchen, S.G. Shrubland ecosystems: Importance, distinguishing characteristics, and dynamics. In Proceedings: Shrubland Dynamics–Fire and Water, Lubbock, TX, USA, 10–12 August 2004; Sosebee, R.E., Wester, D.B., Britton, C.M., McArthur, E.D., Kitchen, S.G., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2007; RMRS-P-47; pp. 3–10. [Google Scholar]
- McKell, C.M. Shrubs—A neglected resource of arid lands. Science 1975, 187, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Van Auken, O.W. Causes and consequences of woody plant encroachment into western North American grasslands. J. Environ. Manag. 2009, 90, 2931–2942. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Soliveres, S. Are shrubs really a sign of declining ecosystem function? Disentangling the myths and truths of woody encroachment in Australia. Aust. J. Bot. 2014, 62, 594–608. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Kröel-Dulay, G.; Ransijn, J.; Schmidt, I.K.; Beier, C.; De Angelis, P.; De Dato, G.; Dukes, J.S.; Emmett, B.; Estiarte, M.; Garadnai, J.; et al. Increased sensitivity to climate change in disturbed ecosystems. Nat. Commun. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, K.C. Shrub recruitment response to intensity and season of fire in a semi-arid woodland. J. Appl. Ecol. 1991, 28, 60–70. [Google Scholar] [CrossRef]
- Hodgkinson, K.C.; Harrington, G.N. The case for prescribed burning to control shrubs in eastern semi-arid woodlands. Aust. Rangel. J. 1985, 7, 64–74. [Google Scholar] [CrossRef]
- Joubert, D.F.; Smit, G.N.; Hoffman, M.T. The role of fire in preventing transitions from a grass dominated state to a bush thickened state in arid savannas. J. Arid Environ. 2012, 87, 1–7. [Google Scholar] [CrossRef]
- Lohmann, D.; Tietjen, B.; Blaum, N.; Joubert, D.F.; Jeltsch, F. Prescribed fire as a tool for managing shrub encroachment in semi-arid savanna rangelands. J. Arid Environ. 2014, 107, 49–56. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Camac, J.S.; Williams, R.J.; Wahren, C.H.; Hoffmann, A.A.; Vesk, P.A. Climatic warming strengthens a positive feedback between alpine shrubs and fire. Glob. Chang. Biol. 2017, 23, 3249–3258. [Google Scholar] [CrossRef] [Green Version]
- Holz, A.; Wood, S.W.; Veblen, T.T.; Bowman, D.M.J.S. Effects of high-severity fire drove the population collapse of the subalpine Tasmanian endemic conifer Athrotaxis cupressoides. Glob. Chang. Biol. 2015, 21, 445–458. [Google Scholar] [CrossRef]
- Flannigan, M.; Cantin, A.S.; De Groot, W.J.; Wotton, M.; Newbery, A.; Gowman, L.M. Global wildland fire season severity in the 21st century. For. Ecol. Manag. 2013, 294, 54–61. [Google Scholar] [CrossRef]
- Fox-Hughes, P.; Harris, R.; Lee, G.; Grose, M.; Bindoff, N. Future fire danger climatology for Tasmania, Australia, using a dynamically downscaled regional climate model. Int. J. Wildland Fire 2014, 23, 309–321. [Google Scholar] [CrossRef]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J. Resprouting as a key functional trait in woody plants—challenges to developing new organizing principles. New Phytol. 2010, 188, 651–654. [Google Scholar] [CrossRef]
- Noble, I.R.; Slatyer, R.O. The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 1980, 43, 5–21. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef] [Green Version]
- Prior, L.D.; Bowman, D.M.J.S. Classification of post-fire responses of woody plants to include pyrophobic communities. Fire 2020, 3, 15. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.M.J.S.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Westerling, A.L.; Turner, M.G.; Smithwick, E.A.H.; Romme, W.H.; Ryan, M.G. Continued warming could transform Greater Yellowstone fire regimes by mid-21st century. Proc. Natl. Acad. Sci. USA 2011, 108, 13165–13170. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Bliss, A.; Bowman, C.J.W.; Prior, L.D. Fire caused demographic attrition of the Tasmanian palaeoendemic conifer Athrotaxis cupressoides. Austral Ecol. 2019, 44, 1322–1339. [Google Scholar] [CrossRef]
- Jackson, W.D. Vegetation of the Central Plateau. In The Lake Country; Banks, M.R., Ed.; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 61–86. [Google Scholar]
- Johnson, K.; Marsden-Smedley, J.B. Fire history of the northern part of the Tasmanian Wilderness World Heritage Area and its associated regions. Pap. Proc. R. Soc. Tasman. 2002, 136, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, R.R. Land use on the Central Plateau with Special Reference to the Grazing Industry. In The Lake Country; Banks, M.R., Ed.; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 161–176. [Google Scholar]
- Cullen, P. Land Degradation on the Central Plateau, Tasmania: The Legacy of 170 Years of Exploitation; Earth Science Section, Parks and Wildlife Service, Dept. of Environment and Land Management: Hobart, Australia, 1995. [Google Scholar]
- Misiak, W.M. Limits to Plant Regeneration in Alpine Vegetation on Tasmania’s Central Plateau. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2007. [Google Scholar]
- Costin, A.B.; Wimbush, D.J.; Kerr, D.; Gay, L.W. Studies in Catchment Hydrology in the Australian Alps. I. Trends in Soils and Vegetation; Technical Paper no.13; CSIRO: Canberra, Australia, 1959. [CrossRef]
- Williams, R.J.; Ashton, D.H. Effects of disturbance and grazing by cattle on the dynamics of heathland and grassland communities on the Bogong High Plains, Victoria. Aust. J. Bot. 1987, 35, 413–431. [Google Scholar] [CrossRef]
- Williamson, G.J.; Murphy, B.P.; Bowman, D.M.J.S. Cattle grazing does not reduce fire severity in eucalypt forests and woodlands of the Australian Alps. Austral Ecol. 2014, 39, 462–468. [Google Scholar] [CrossRef]
- AFAC. A Review of the Management of the Tasmanian Fires of December 2018 to March 2019; Technical Report; Australasian Fire and Emergency Service Authorities Council Limited: East Melbourne, VIC, Australia, 2019. [Google Scholar]
- Kirkpatrick, J.B.; Dickinson, K.J.M. The impact of fire on Tasmanian alpine vegetation and soils. Aust. J. Bot. 1984, 32, 613–629. [Google Scholar] [CrossRef]
- Balmer, J.; Whinam, J.; Kelman, J.; Kirkpatrick, J.B.; Lazarus, E. A Review of the Floristic Values of the Tasmanian Wilderness World Heritage Area. Nature Conservation Report 2004/3; Department of Primary Industries Water and Environment: Tasmania, Australia, 2004.
- Tasmanian Vegetation Mapping Program (TVMP). TASVEG Version 4.0, Tasmanian Vegetation Map 1:25,000; Department of Primary Industries, Water and Environment: Hobart, Australia, 2020.
- Banks, M.R. (Ed.) General Geology. In The Lake Country; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 25–33. [Google Scholar]
- Gentilli, J. Australian Climate Patterns; Thomas Nelson (Australia) Limited: Melbourne, Australia, 1972. [Google Scholar]
- Kirkpatrick, J.B. Treeless plant communities of the Tasmanian high country. Proc. Ecol. Soc. Aust. 1983, 12, 61–77. [Google Scholar]
- Bureau of Meteorology. Climate Data Online. Climate Services. Bureau of Meteorology. Available online: http://www.bom.gov.au/ (accessed on 17 September 2020).
- Giglio, L.; Descloitres, J.; Justice, C.O.; Kaufman, Y.J. An enhanced contextual fire detection algorithm for MODIS. Remote Sens. Environ. 2003, 87, 273–282. [Google Scholar] [CrossRef]
- Williamson, G.J.; Price, O.F.; Henderson, S.B.; Bowman, D.M.J.S. Satellite-based comparison of fire intensity and smoke plumes from prescribed fires and wildfires in south-eastern Australia. Int. J. Wildland Fire 2013, 22, 121–129. [Google Scholar] [CrossRef]
- French, B.J.; Prior, L.D.; Williamson, G.J.; Bowman, D.M.J.S. Cause and effects of a megafire in sedge-heathland in the Tasmanian temperate wilderness. Aust. J. Bot. 2016, 64, 513–525. [Google Scholar] [CrossRef]
- Murphy, B.P.; Prior, L.D.; Cochrane, M.A.; Williamson, G.J.; Bowman, D.M.J.S. Biomass consumption by surface fires across Earth’s most fire prone continent. Glob. Chang. Biol. 2018, 25, 254–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.M.; Oechel, W.C. A simple method for estimating fire intensity after a burn in California chaparral. Acta Oecol./Oecol. Plant 1989, 10, 57–68. [Google Scholar]
- Whight, S.; Bradstock, R. Indices of fire characteristics in sandstone heath near Sydney, Australia. Int. J. Wildland Fire 1999, 9, 145–153. [Google Scholar] [CrossRef]
- Bliss, A.; Prior, L.D.; Bowman, D.M.J.S. Lack of reliable post-fire recovery mechanisms makes the iconic Tasmanian conifer Athrotaxis Cupressoides susceptible to population decline. Aust. J. Bot. 2021, 69, 162–173. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Ishiguro, M.; Kitagawa, G. Akaike Information Criterion Statistics; D. Reidel: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Vuong, Q.H. Likelihood ratio tests for model selection and non-nested hypotheses. Econom. J. Econom. Soc. 1989, 57, 307–333. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Nicholson, A.; Prior, L.D.; Perry, G.L.W.; Bowman, D.M.J.S. High post-fire mortality of resprouting woody plants in Tasmanian Mediterranean-type vegetation. Int. J. Wildland Fire 2017, 26, 532–537. [Google Scholar] [CrossRef]
- Walsh, N.G.; McDougall, K.L. Progress in the recovery of the flora of treeless subalpine vegetation in Kosciuszko National Park after the 2003 fires. Cunninghamia 2004, 8, 439–452. [Google Scholar]
- Clarke, P.J.; Knox, K.J.E.; Campbell, M.L.; Copeland, L.M. Post-fire recovery of woody plants in the New England Tableland Bioregion. Cunninghamia 2009, 11, 221–239. [Google Scholar]
- Bell, D.T. Ecological response syndromes in the flora of southwestern Western Australia: Fire resprouters versus reseeders. Bot. Rev. 2001, 67, 417–440. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Sprouting ability across diverse disturbances and vegetation types worldwide. J. Ecol. 2004, 92, 310–320. [Google Scholar] [CrossRef]
- Vesk, P.A.; Warton, D.I.; Westoby, M. Sprouting by semi-arid plants: Testing a dichotomy and predictive traits. Oikos 2004, 107, 72–89. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Bridle, K.L.; Wild, A.S. Succession after fire in alpine vegetation on Mount Wellington, Tasmania. Aust. J. Bot. 2002, 50, 145–154. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; Wonkka, C.L.; Taylor, C.A., Jr.; Kreuter, U.P. Extreme prescribed fire during drought reduces survival and density of woody resprouters. J. Appl. Ecol. 2016, 53, 1585–1596. [Google Scholar] [CrossRef] [Green Version]
- Calder, J.A.; Kirkpatrick, J.B. Climate change and other factors influencing the decline of the Tasmanian cider gum (Eucalyptus gunnii). Aust. J. Bot. 2008, 56, 684–692. [Google Scholar] [CrossRef]
- Piper, F.I.; Paula, S. The role of nonstructural carbohydrates storage in forest resilience under climate change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bridle, K.L.; Kirkpatrick, J.B.; Cullen, P.; Shepherd, R.R. Recovery in alpine heath and grassland following burning and grazing, Eastern Central Plateau, Tasmania, Australia. Arctic Antarct. Alp. Res. 2001, 33, 348–356. [Google Scholar] [CrossRef]
- McPhail, D.C.; Kirkpatrick, J.B. Patch mosaic cyclic succession associated with the growth and senescence of an alpine shrub. Aust. J. Bot. 2016, 64, 111–119. [Google Scholar] [CrossRef]
- Williams, R.J. Gap dynamics in subalpine heathland and grassland vegetation in south-eastern Australia. J. Ecol. 1992, 80, 343–352. [Google Scholar] [CrossRef]
- Bridle, K.L.; Kirkpatrick, J.B. Comparative effects of stock and wild vertebrate herbivore grazing on treeless subalpine vegetation, Eastern Central Plateau, Tasmania. Aust. J. Bot. 1999, 47, 817–834. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Bridle, K.L. Grazing and the absence of fire promote the dominance of an unpalatable shrub in a patch mosaic cyclic successional system. Aust. J. Bot. 2016, 64, 45–50. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Marsden-Smedley, J.B.; Folco, M.B.D.; Leonard, S.W.J. Influence of grazing and vegetation type on post-fire floristic and lifeform composition in Tasmania, Australia. Plant Ecol. 2016, 217, 57–69. [Google Scholar] [CrossRef]
- Styger, J.K.; Kirkpatrick, J.B.; Marsden-Smedley, J.O.N.; Leonard, S.W.J. Fire incidence, but not fire size, affects macropod densities. Austral Ecol. 2011, 36, 679–686. [Google Scholar] [CrossRef]
- Locke, S. The Distribution and Abundance of Fallow Deer in the Central Plateau Conservation Area and Adjacent Areas in Tasmania: A Baseline Monitoring Program; Nature Conservation Report 07/02; Department of Primary Industries and Water: Hobart, Australia, 2007.
- Kirkpatrick, J.B.; Bridle, K.L.; Dickinson, K.J.M. Decades-scale vegetation change in burned and unburned alpine coniferous heath. Aust. J. Bot. 2010, 58, 453–462. [Google Scholar] [CrossRef]
- Williams, R.J.; Wahren, C.H.; Shannon, J.M.; Papst, W.A.; Heinze, D.A.; Camac, J.S. Fire regimes and biodiversity in Victoria’s alpine ecosystems. Proc. R. Soc. Vic. 2012, 124, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- Batllori, E.; Ackerly, D.D.; Moritz, M.A. A minimal model of fire-vegetation feedbacks and disturbance stochasticity generates alternative stable states in grassland–shrubland–woodland systems. Environ. Res. Lett. 2015, 10, 034018. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Briggs, J.M.; Blair, J.M. Fire dynamics distinguish grasslands, shrublands and woodlands as alternative attractors in the Central Great Plains of North America. J. Ecol. 2014, 102, 1374–1385. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, Z.; Nippert, J.B.; Ocheltree, T.W. Abrupt transition of mesic grassland to shrubland: Evidence for thresholds, alternative attractors, and regime shifts. Ecology 2014, 95, 2633–2645. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, A.T. Grazing systems, ecosystem responses, and global change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Noble, J.C.; Hik, D.S.; Sinclair, A.R.E. Landscape ecology of the burrowing bettong: Fire and marsupial biocontrol of shrubs in semi-arid Australia. Rangel. J. 2007, 29, 107–119. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub invasions of North American semiarid grasslands. Annu. Rev. Ecol. Syst. 2000, 31, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Wood, S.W.; Neyland, D.; Sanders, G.J.; Prior, L.D. Contracting Tasmanian montane grasslands within a forest matrix is consistent with cessation of Aboriginal fire management. Austral Ecol. 2013, 38, 627–638. [Google Scholar] [CrossRef]
- Romanin, L.M.; Hopf, F.; Haberle, S.G.; Bowman, D.M.J.S. Fire regime and vegetation change in the transition from Aboriginal to European land management in a Tasmanian eucalypt savanna. Aust. J. Bot. 2016, 64, 427–440. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Location of the study area. Map of Tasmania inset showing location of the 2019 fire (black box respresents area shown in (a,b)) in relation to the Tasmanian Wilderness World Heritage Area (TWWHA) and Central Plateau Conservation Area (CPCA). (a) Variation in fire intensity (MODIS and VIIRS Fire Radiative Power (FRP) in MW). To illustrate trend in fire intensity, FRP values have been rasterized with pixel width approximately 2 km. (b) Burn severity (based on differenced Normalized Burn Ratio) of the 2019 fire for pre- and post-fire Sentinel 2 imagery. Image dates: Pre-fire 31 December 2018 and 13 January 2019; post-fire 1 March 2019. dNBR burn severity values: unburnt ≤ 0.09999; low > 0.09999 ≤ 0.19999; moderate > 0.19999 ≤ 0.39999; high > 0.59999, ≤ 0.99999; very high > 0.99999. Note that shrubland areas generally burned at very high severities due to canopy combustion.
Figure 1.
Location of the study area. Map of Tasmania inset showing location of the 2019 fire (black box respresents area shown in (a,b)) in relation to the Tasmanian Wilderness World Heritage Area (TWWHA) and Central Plateau Conservation Area (CPCA). (a) Variation in fire intensity (MODIS and VIIRS Fire Radiative Power (FRP) in MW). To illustrate trend in fire intensity, FRP values have been rasterized with pixel width approximately 2 km. (b) Burn severity (based on differenced Normalized Burn Ratio) of the 2019 fire for pre- and post-fire Sentinel 2 imagery. Image dates: Pre-fire 31 December 2018 and 13 January 2019; post-fire 1 March 2019. dNBR burn severity values: unburnt ≤ 0.09999; low > 0.09999 ≤ 0.19999; moderate > 0.19999 ≤ 0.39999; high > 0.59999, ≤ 0.99999; very high > 0.99999. Note that shrubland areas generally burned at very high severities due to canopy combustion.

Figure 2.
(a) Broad vegetation groups within the vicinity of the 2019 Great Pine Tier fire (black outline) (from Reference [39]). Uncolored areas indicate other vegetation groups. (b) Burn severity of the montane treeless shrubland (a subset of the Highland and treeless vegetation group depicted in (a)) and the location of the survey transects. Burn severity is based on differenced Normalized Burn Ratio of the 2019 fire for pre- and post-fire Sentinel 2 imagery. Image dates: Pre-fire 31 December 2018 and 13 January 2019; post-fire 1 March 2019. dNBR burn severity values: unburnt ≤ 0.09999; low > 0.09999 ≤ 0.19999; moderate > 0.19999 ≤ 0.39999; high > 0.59999, ≤ 0.99999; very high > 0.99999.
Figure 2.
(a) Broad vegetation groups within the vicinity of the 2019 Great Pine Tier fire (black outline) (from Reference [39]). Uncolored areas indicate other vegetation groups. (b) Burn severity of the montane treeless shrubland (a subset of the Highland and treeless vegetation group depicted in (a)) and the location of the survey transects. Burn severity is based on differenced Normalized Burn Ratio of the 2019 fire for pre- and post-fire Sentinel 2 imagery. Image dates: Pre-fire 31 December 2018 and 13 January 2019; post-fire 1 March 2019. dNBR burn severity values: unburnt ≤ 0.09999; low > 0.09999 ≤ 0.19999; moderate > 0.19999 ≤ 0.39999; high > 0.59999, ≤ 0.99999; very high > 0.99999.

Figure 3.
Climate data for Liawenee for the period 2001 to 2019. Bars show mean monthly rainfall (mm). Solid circles are mean monthly maximum temperatures (C), and open circles mean monthly minimum temperatures.
Figure 3.
Climate data for Liawenee for the period 2001 to 2019. Bars show mean monthly rainfall (mm). Solid circles are mean monthly maximum temperatures (C), and open circles mean monthly minimum temperatures.

Figure 4.
Photographs of Central Plateau Conservation Area shrubland. (a,b) Unburnt shrubland in the vicinity of Lake Augusta. (c,d) The shrublands 12 months following the 2019 fire, illustrating the severity of the fire. Photographs: (a,b) R. Corkrey; (c,d) J. Foulkes.
Figure 4.
Photographs of Central Plateau Conservation Area shrubland. (a,b) Unburnt shrubland in the vicinity of Lake Augusta. (c,d) The shrublands 12 months following the 2019 fire, illustrating the severity of the fire. Photographs: (a,b) R. Corkrey; (c,d) J. Foulkes.

Figure 5.
Proportion of Orites revoluta resprouting in relation to (a) burnt twig diameter and (b) shrub basal area. The total number of O. revoluta plants is also shown in relation to (c) burnt twig diameter and (d) shrub basal area. Note that Shrub basal area is shown on a log scale.
Figure 5.
Proportion of Orites revoluta resprouting in relation to (a) burnt twig diameter and (b) shrub basal area. The total number of O. revoluta plants is also shown in relation to (c) burnt twig diameter and (d) shrub basal area. Note that Shrub basal area is shown on a log scale.

Figure 6.
Seedling density as a function of fire severity (measured by mean burnt twig diameter per plot). Note that one outlier was removed for illustration purposes, but was included in the analysis.
Figure 6.
Seedling density as a function of fire severity (measured by mean burnt twig diameter per plot). Note that one outlier was removed for illustration purposes, but was included in the analysis.

Table 1.
Proportion of shrubland within the fireground of the Central Plateau Conservation Area (CPCA) burnt at differing fire severities (as measured by differenced Normalised Burn Ratio), and the proportion of plots we surveyed within each burn severity category. Note that the percentages shown for the CPCA are similar to that for the total fire area.
Table 1.
Proportion of shrubland within the fireground of the Central Plateau Conservation Area (CPCA) burnt at differing fire severities (as measured by differenced Normalised Burn Ratio), and the proportion of plots we surveyed within each burn severity category. Note that the percentages shown for the CPCA are similar to that for the total fire area.
| Fire Severity | CPCA Fire Area | Survey Plots | ||
|---|---|---|---|---|
| % Total Area | % Shrubland | % Total Area | % Shrubland | |
| Unburnt | 5.2 | 4.4 | 5 | 8.3 |
| Low | 4.7 | 3.2 | 1 | 1.7 |
| Medium | 14.3 | 8.2 | 2 | 3.3 |
| High | 29.2 | 26.1 | 8 | 13.3 |
| Very high | 46.6 | 58.0 | 44 | 73.3 |
Table 2.
Number of shrubs in the unburnt and burnt plots, condition of shrubs in the burnt plots and mean basal area of shrubs in the unburnt and burnt plots. Note that only 5 of the unburnt shrubs were dead, and none was resprouting, so their condition is not presented here.
Table 2.
Number of shrubs in the unburnt and burnt plots, condition of shrubs in the burnt plots and mean basal area of shrubs in the unburnt and burnt plots. Note that only 5 of the unburnt shrubs were dead, and none was resprouting, so their condition is not presented here.
| Species | Unburnt | Burnt | Condition of Shrubs in the Burnt Plots | Shrub Basal Area (cm) ± s.d. | |||
|---|---|---|---|---|---|---|---|
| Alive | Dead | Resp. | Unburnt | Burnt | |||
| Acrothamnus montanus (R.Br.) Quinn | 0 | 1 | 1 | 0 | 0 | n.a. | 250 |
| Bauera rubioides Andrews | 0 | 1 | 0 | 0 | 1 | n.a | 135 |
| Leptospermum rupestre Hook.f. | 0 | 15 | 1 | 12 | 2 | n.a | 169 ± 256 |
| Orites acicularis (R.Br.) Roem. & Schult. | 14 | 40 | 0 | 40 | 0 | 303 ± 394 | 225 ± 216 |
| Orites revoluta R.Br. | 9 | 77 | 2 | 35 | 40 | 223 ± 294 | 528 ± 1130 |
| Ozothamnus hookeri Sond. | 30 | 13 | 0 | 13 | 0 | 141 ± 223 | 108 ± 101 |
| Richea acerosa (Lindl.) F.Muell. | 24 | 54 | 9 | 45 | 0 | 224 ± 306 | 178 ± 228 |
| Richea sprengeloides (R.Br.) F.Muell. | 0 | 43 | 0 | 43 | 0 | n.a | 65 ± 72 |
| Unknown | 0 | 715 | 0 | 715 | 0 | n.a | 72 ± 97 |
| Total | 77 | 959 | 8 | 901 | 43 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Foulkes, J.A.; Prior, L.D.; Leonard, S.W.J.; Bowman, D.M.J.S. Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau. Fire 2021, 4, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/fire4030032
AMA Style
Foulkes JA, Prior LD, Leonard SWJ, Bowman DMJS. Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau. Fire. 2021; 4(3):32. https://0-doi-org.brum.beds.ac.uk/10.3390/fire4030032
Chicago/Turabian StyleFoulkes, Judy A., Lynda D. Prior, Steven W. J. Leonard, and David M. J. S. Bowman. 2021. "Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau" Fire 4, no. 3: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/fire4030032