NG2 Glia: Novel Roles beyond Re-/Myelination


1
Department of Neuroscience Rita Levi Montalcini, University of Turin, 10126 Turin, Italy
2
Neuroscience Institute of the Cavalieri Ottolenghi Foundation (NICO), Regione Gonzole 10, 10043 Orbassano (Turin), Italy
*
Author to whom correspondence should be addressed.
Neuroglia 2018, 1(1), 151-175; https://0-doi-org.brum.beds.ac.uk/10.3390/neuroglia1010011
Submission received: 10 June 2018
/
Revised: 24 June 2018
/
Accepted: 29 June 2018
/
Published: 4 July 2018
Abstract
:Neuron-glia antigen 2-expressing glial cells (NG2 glia) serve as oligodendrocyte progenitors during development and adulthood. However, recent studies have shown that these cells represent not only a transitional stage along the oligodendroglial lineage, but also constitute a specific cell type endowed with typical properties and functions. Namely, NG2 glia (or subsets of NG2 glia) establish physical and functional interactions with neurons and other central nervous system (CNS) cell types, that allow them to constantly monitor the surrounding neuropil. In addition to operating as sensors, NG2 glia have features that are expected for active modulators of neuronal activity, including the expression and release of a battery of neuromodulatory and neuroprotective factors. Consistently, cell ablation strategies targeting NG2 glia demonstrate that, beyond their role in myelination, these cells contribute to CNS homeostasis and development. In this review, we summarize and discuss the advancements achieved over recent years toward the understanding of such functions, and propose novel approaches for further investigations aimed at elucidating the multifaceted roles of NG2 glia.
1. Introduction
During central nervous system (CNS) ontogenesis, myelinating oligodendrocytes (OLs) originate from parenchymal precursors expressing the neuron-glia antigen 2 (NG2) chondroitin sulfate proteoglycan, and therefore, commonly referred to as oligodendrocyte precursor cells or NG2-expressing glia (NG2 glia) [1]. These cells persist in the adult CNS parenchyma, where they comprise about 5% of all CNS cells [2], and can serve as a rapidly responding reservoir for new OLs in case of demyelination [1,3,4]. In intact adult nervous tissue, NG2 glia can also be engaged in proliferation and maturation to sustain a certain degree of oligodendrogenesis [5,6] and myelin plasticity [7,8,9,10,11]. Notably, recent in vivo longitudinal two-photon imaging studies showed that, in the murine cortex, OL density continues to increase until two years of age [10], and more than half of the OLs present in middle aged mice are produced during adult life (i.e., after fourmonths of age [9]). The generation of such adult-born OLs is required to maintain proper axonal functions [12], and is involved in the production of new myelin segments [9] and changes of the circuit properties subserving experience-dependent plasticity (i.e., motor skills learning [7,8]). However, in both adult human and rodent CNS, the density, distribution and proliferative rate of NG2 glia seem to be independent of the presence/density of myelinated fibres. NG2 glia distribute homogenously throughout grey and white matter parenchyma. Further, although gray matter cells have a longer cell cycle length than their white matter counterparts [13], NG2 glia proliferative fractions do not differ in highly myelinated vs. non-/poorly myelinated brain regions (i.e., the granular and the molecular layers of the cerebellum, respectively; our unpublished observations). In intact conditions, most NG2 glia appear as “quiescent” cells (i.e., neither progressing along the lineage, nor re-entering the cell cycle), and overall, the fraction of NG2 glia actively engaged in oligodendrogenesis is minimal [14,15]. These observations suggest that in basal conditions such an abundant pool of resident NG2 glia may exert additional roles beyond their functions in oligodendrogenesis and myelin production.
Although the molecular aspects of NG2 glia response to injury are far from being completely understood, data accumulated so far indicate that, phenotypically, NG2 glia provide a stereotypic reaction (i.e., increased NG2 expression, retraction of cell processes, cell body swelling, cell proliferation and migration toward the lesion site) to almost all kind of injury, independently of the extent of myelin loss [14,16,17,18,19]. Further, upon experimental demyelination in the subcortical white matter, transient populations of NG2 glia coming from the subventricular zone (SVZ) of the lateral ventricles immediately amplify and invade the lesion site, but ultimately seem not to contribute to myelin repair, and are lost [20,21,22]. This indicates that the reactivity and amplification of NG2 glia (or at least of some NG2 glia subsets) per se does not simply reflect a regenerative event. The post-injury emergence of transient NG2 glia populations is reminiscent of the developmental scenario, where three waves of NG2 glia are sequentially produced in the forebrain, and the first embryonically-generated cells are entirely replaced during the first week of life [23]. The generation of such cell populations that do not contribute to myelination suggests again that NG2 glia/glial subsets exert additional/alternative functions in specific developmental/post-injury phases.
In line with this view, a subpopulation of NG2 glia has been observed to serve as multipotent progenitors, producing protoplasmic astrocytes as well OLs in the embryonic ventral forebrain or following some kinds of CNS injury [19,24,25,26,27]. It has also been proposed that adult NG2 glia may activate spontaneous neurogenic events within restricted brain areas [28]. In view of achieving neuronal replacement in the adult CNS, the idea that NG2 glia can be the source for new neurons is particularly attractive because of their abundance and ubiquitous distribution. However, this has been proven true in vivo only upon specific cell reprogramming approaches, and clear evidence that adult NG2 glia spontaneously contribute to parenchymal neurogenesis or astrogliogenesis is lacking [4,28,29,30].
Based on these and other findings, four years ago we proposed that, beyond sustaining re-/myelination, NG2 glia may participate in the nervous tissue homeostasis, neuromodulation, developmental and post-injury events [28]. Such a view has recently been substantiated by a growing body of evidence, including descriptive observations and results of functional in vivo/in vitro studies. Here, we summarize and discuss the advancements obtained in the field in the last years and propose novel approaches for further investigations aimed at understanding the multifaceted roles of NG2 glia.
2. NG2 Glia Distribution, Self-Maintenance and Anatomical Relationships with Central Nervous System Cells
Recent studies showed that NG2 glia distribution and density are tightly and homeostatically regulated in adult CNS tissue. In vivo two-photon imaging analyses showed that NG2 glia processes constantly survey their local environment with highly motile filopodia and growth cone-like structures that retract upon contact with other NG2 cells [31,32]. As a result, NG2 glia are arranged in a grid-like manner, where cells are equally spaced and, similar to astrocytes, occupy tridimensional non-overlapping domains. In the mammalian and zebrafish CNS, NG2 glia division and short distance migration are the mechanisms by which these cells maintain their uniform density in space and time: once an individual NG2-expressing cell is “lost” because of maturation (i.e., loss of progenitor features including expression of cell surface signal molecules) or experimental ablation, a neighboring NG2 cell enters the cell cycle and migrate to fill the unoccupied space (Figure 1A) [31,32,33,34]. Notably, similar to stem cells, NG2 glia can divide asymmetrically in vivo and give rise to a mixed progeny, either keeping a progenitor phenotype or proceeding to differentiation (Figure 1B) [5,6,35]. This mechanism may allow the generation of OLs, while preventing the progressive exhaustion of NG2 glia during adulthood. NG2 glia maintenance is also regulated by specific synapse-mediated signals received from γ-aminobutyric acid (GABA)-ergic neurons (see also below), as demonstrated by the progressive depletion of the NG2 glia pool lacking the GABA-A receptor subunit γ2 [36]. The discovery of such efficient—and somehow redundant—regulation mechanisms by which a constant population of NG2 glia is homeostatically maintained in time and space, even in non-/poorly-myelinated CNS regions, challenges the assumption that NG2 glia exclusively serve as progenitors for new OLs, and suggests functions related to the surveillance and modulation of the activity of the surrounding neuropil, as formerly assessed for astrocytes and microglia [37,38,39,40].
In line with this view, confocal and electron microscopy studies showed that NG2 glia establish physical contacts with functionally relevant neuronal domains, including dendrites [41,42], somata [14,41,43,44], nodes of Ranvier [45,46] and synapses [47,48] (Figure 1C). About 30–50% of the nodes are contacted by both NG2 glia and astrocyte processes in the optic nerve, corpus callosum, and spinal cord of young adult rodents. Ultrastructural analyses revealed that NG2 glia processes extend fine, finger-like projections that contact the nodal membrane at discrete points, while astrocytes had broader processes that surround the entire nodes [46] (Figure 1C). Similarly, NG2 glia and astrocytes often contact the same axon terminals, with NG2 glia thin processes interdigitating between the pre- and post-synaptic elements and large astrocytic processes ensheating the entire synapses [41,47] (Figure 1C). These observations suggest different roles of the contacts established by the two glial cell types at nodes and synaptic sites.
What makes NG2 glia unique among the glial cells is their connections with neurons through unidirectional neuron-to-NG2 glia synapses. Most, if not all NG2 glia in the grey and white matter receive functional glutamatergic and/or GABAergic synaptic contacts, whose machinery and ultrastructure is remarkably similar to that of conventional neuron-to-neuron synapses. Such organization includes the tight alignment of neuron and NG2 glia cell membranes, the presence of an active zone with accumulation of synaptic vesicles on the neuronal side, and an electron dense postsynaptic density on the NG2 cell side. Notably, neuron-to-NG2 glia synapses are established in parallel with neuronal synaptogenesis [14,49], and are lost during NG2 glia progression in maturation [48,49]; this is in line with a specific role of this form of communication in undifferentiated NG2 glia. Remarkably, while glutamatergic neuron-to-NG2 glia synapses usually derive from long-range axons impinging almost exclusively on NG2 glia processes [50,51,52], GABAergic synapses derive from local interneurons [49,53] with a specific distribution of the synaptic contacts from fast spiking (located at NG2 glia somata and proximal parts of the processes) and non-fast spiking interneurons (located at the distal parts of NG2 glia processes) [49]. Although both glutamatergic and GABAergic neuron-to-NG2 glia synaptic inputs induce depolarization (since the measured chloride reversal potential in these cells is around −30/−40 mV), such well-defined arrangement suggests distinct roles for the different types of synapses impinging onto NG2 glia. In line with this idea, while the glutamatergic inputs onto NG2 glia increase in frequency and amplitude during CNS maturation [54], at least in the cerebral cortex, GABAergic neuron-to-NG2 glia synaptic transmission is a phenomenon restricted to developmental stages [49,55].
Intimate physical interactions also occur between NG2 glia and astrocytes [56,57], microglia [58], and myelinating OLs. Notably, while NG2 glia are never coupled via gap-junctions [59,60], some of them express connexin 32 [61] and partly couple to mature OLs [62]. Further, electron microscopy analysis showed that NG2 glia processes often contact the paranodal loops of myelin [46]. These data suggest a privileged communication between NG2 glia and other elements in the oligodendroglial lineage.
In the embryonic mouse and human brain, NG2 glia are closely associated with the developing blood vessels, by being either positioned at the sprouting tip or tethered along the abluminal surface of the endothelium via the basal lamina [63,64]. Some populations of NG2 glia maintain a perivascular distribution also in the mouse neonatal brain and adult white matter [65,66,67,68]. In such perivascular niche, NG2 glia establish direct interactions through their processes or somata with both endothelial cells and pericytes. Beside these perivascular NG2 glia, a large fraction of parenchymal NG2 cells, whose somata are located away from blood vessels, extend some processes to physically interact with pericytes and microvessels [41,46] (Figure 1C). Such a distribution suggests some form of crosstalk among NG2 glia, endothelial cells and pericytes (see below).
3. NG2 Glia as Sensors of Neuronal Activity
NG2 glia contacts/contiguity with functionally relevant neuronal domains put them in a strategic position to monitor the activity of neuronal circuitries, integrate distinct inputs, and possibly respond to changes of the firing patterns of the surrounding neurons [69]. Indeed, a number of studies have shown that NG2 glia dynamically react to alterations in neurotransmission [70]. Pioneer experiments have shown that suppression of neural activity due to intraocular injection of tetrodotoxin (TTX) remarkably reduces NG2 glia proliferation in the optic nerve [71]. Consistently, deprivation of sensory experience (i.e., whisker removal at developmental stages) negatively affects NG2 glia proliferation and survival, and alters their distribution in the somatosensory cortex [6,52]. In contrast, electrical stimulation of the corticospinal tract promotes NG2 cell proliferation and differentiation in the adult rat [72]. Similarly, neuronal activity induced by optogenetic stimulation in Thy1-channelrhodopsin-2 transgenic mice, elicits a mitogenic response in NG2 glia and increases their differentiation [73]. Again in line with an activity-dependent regulation of the number of NG2 glia and OLs, increased rates of NG2 glia proliferation have been observed in response to wheel running and environmental enrichment [74]. However, in later studies, voluntary physical exercise has instead been reported as being accompanied by a reduction of the proliferation rate, premature differentiation, and changes in NG2 glia division modality in the adult mouse brain [5,75,76]. Indeed, a recent study showed that the pattern of neuronal activity, rather than just the presence or absence of activity, determines the activity-dependent behavior of NG2 glia in vivo. By implanting an electrode array into the corpus callosum of adult mice, Nagy and colleagues [77] showed that NG2 glia respond differently when callosal axons are stimulated at 5, 25 or 300 hertz (Hz). Namely, stimulation at 5 Hz promoted NG2 glia maturation, while stimulation at 25 or 300 Hz stimulated NG2 glia proliferation. These findings are particularly noteworthy, because the rate and timing of neuronal firing are the main carriers of information about the features of a task or stimulus, and also because they imply that NG2 glia are somehow capable of discriminating between different patterns of neuronal firing (see below).
How do NG2 glia sense neuronal activity? Indeed, another feature that makes NG2 glia unique among glial cells is the expression of a large repertoire of typically “neuronal” proteins, including ion channels and neurotransmitter receptors. These comprise Ca2+-permeable and impermeable α-amino-3-hydroxy-5-methyl-4-isoxazolepropionicacid (AMPA) and N-methyl-d-aspartate (NMDA) ionotropic glutamate receptors; group I (mGluR1 and mGluR5), group II (mGluR3) and group III (mGluR4) metabotropic glutamate receptors; ionotropic and metabotropic GABA receptors; Ca2+-permeable and impermeable nicotinic acetylcholine receptors (nAChRs); muscarinic acetylcholine receptors (mAChRs); ionotropic glycine receptors (GlyRs); adrenergic, dopamine, serotonin and purinergic receptors [78]. The expression of these receptors is not universal in NG2 cell populations (e.g., only 60% of the callosal NG2 cells are able to respond to NMDA [79]), indicating a certain degree of heterogeneity within NG2 glia as regards their potential to sense and respond to changes in neurotransmission. Despite the fact that some of these neurotransmitter receptors (i.e., AMPA and ionotropic GABA receptors) have been shown to mediate neuron-to-NG2 glia synaptic communication, activation of most of them is thought to occur through extrasynaptic mechanisms [48], and is associated with large, widespread intracellular Ca2+ elevations [78].
Beside such activity-dependent “paracrine” interactions with neurons, studies show that NG2 cells are particularly well-suited for exerting a more tight and precise monitoring of the surrounding neuropil. First, NG2 glia are physically integrated in neuronal circuitries, since they receive synapses from collaterals of axons simultaneously impinging on nearby excitatory or inhibitory neurons [50,51,54]. Thus, by receiving paired/synchronous synaptic signals, NG2 glia can operate a real-time control of the incoming inputs onto surrounding neurons. Further, the expression of high levels of “leak” potassium channels and the connection with neurons through conventional synapses make NG2 glia able to sense even small changes in neuronal activity with an extremely high temporal and spatial resolution. Of note, NG2 glia have been reported to detect fine changes of extracellular potassium concentration due to the discharge of a single neuron via the inward-rectifier Kir4.1 potassium channels [78,80]. Similarly, by employing two-photon-based glutamate uncaging to produce very localized and brief release of glutamate onto NG2 glia processes segments, Sun et al. [81] showed that small neurotransmitter release events at neuron-to-NG2 cell synapses can be sensed via the generation of local depolarizations and, consequentially, local Ca2+ signals in NG2 glia processes. The incoming electrical synaptic inputs are temporally and spatially summed and integrated by NG2 glia, by using Ca2+ levels to proportionally encode the number of the active synapses [81]. Interestingly, by analysing the properties of ionic currents elicited by repetitive axonal stimulation at glutamatergic neuron-to-NG2 glia synapses, Nagy et al. [77] recently demonstrated that, similar to neurons, NG2 glia are also able to discriminate different patterns of presynaptic axonal activity (i.e., stimulation trains with distinct frequency of pulses). Since in neurons different patterns of incoming inputs induce Ca2+ signals with distinct temporal and spatial distribution and the activation of diverse signaling pathways, such discriminative ability is thought to subserve the engagement of NG2 glia in either proliferation or differentiation following stimulation at different frequencies [77].
On the whole, these findings indicate that NG2 glia are specialized “listeners and integrators” of neuronal activity, and adjust their behavior in response to its changes. In this context, intracellular Ca2+ transients—generated in different spatial domains of NG2 glia depending on the pattern/type of incoming activity (including synaptic and extrasynaptic inputs)—likely serve distinct roles. Namely, large Ca2+ signals involving the entire cell arborization and soma are particularly apt to induce gene expression changes and global cellular actions such as cell division, differentiation, survival or motility [78]. Conversely, beyond allowing the integration of the synaptic activity received by distinct axons, Ca2+ transients restricted to specific segments of individual processes may be implicated in more compartmentalized functions, such as the stabilization of contacts between NG2 glia processes and axons [82,83], process motility [31], local protein synthesis [84,85], or secretion. Indeed, NG2 glia produce a wide range of “neuroactive” factors, and the release of some of them is activity-dependent (see below). This suggests that the integration of NG2 glia in neuronal circuitries and their exquisite ability to perceive activity changes/patterns may be instrumental for a neuron-to-NG2 glia-to-neuron communication loop contributing to homeostasis and/or plasticity.
4. Maintenance of NG2 Glia Is Required for Central Nervous System Homeostasis and Development
To unveil the specific contribution of NG2 glia to CNS functions/ontogeny, a wide range of approaches have been developed to study the consequences of their selective ablation from the adult or developing brain. Pioneer studies have exploited the exposure to high-doses of X-rays or the infusion of mitotic blockers (such as Arabinofuranosyl cytidine (AraC)) to ablate cycling cells, including NG2 glia. These approaches lacked cell-type specificity and suffered from side-effects, making it difficult to attribute a phenotype exclusively to NG2 glia loss. Thus, in the most recent studies, NG2 glia ablation has been achieved by the generation of mouse lines expressing a “suicide” gene under a NG2 glia specific promoter. Suicide genes typically encode for an essential protein, for a toxin, or for an enzyme that converts an exogenous drug into a toxic agent [86]. Of note, in all cases the effectiveness of the control mechanisms by which NG2 glia density/distribution are maintained in space and time hampered a long-lasting NG2 glia ablation. Even in the most efficient systems, cells escaping the ablation immediately reacted, entering the cell cycle and replacing the lost cells. Thus, no study has succeeded so far in ablating NG2 glia for long periods, and available data only refer to phenotypes emerging from transient NG2 glia loss. However, despite this limitation, ablation strategies provided important evidence for a NG2 glia specific contribution in CNS physiology and development, beyond their role as OL progenitors.
By using a transgenic mouse model expressing the diphtheria toxin receptor (DTR) under the control of the NG2 promoter (NG2Cre-R26DTR mice), Birey and colleagues [87] showed that the selective ablation of about 50% of NG2 glia in adult mice caused deficits in the glutamatergic neurotransmission (i.e., decreased amplitude and increased decay of the miniature excitatory postsynaptic currents (mEPSC), and an altered postsynaptic glutamate receptor trafficking in pyramidal excitatory neurons), negatively affected the astrocytic extracellular glutamate uptake, and induced depressive-like behaviors in mice. Of note, these effects could be observed only in the prefrontal cortex. No changes were observed in the dorsal striatum, while different electrophysiological phenotypes have been found in the somatosensory cortex (where both mEPSC frequency and decay were increased), indicating a differential contribution of NG2 glia in the regulation of glutamatergic neurotransmission in distinct brain regions. By using this ablation strategy, NG2 glia loss did not result in brain vasculature alterations, inflammation, microglia activation, or neurodegeneration [87].
Conversely, in a transgenic rat model expressing thymidine kinase (TK) (that converts the prodrug ganciclovir into a toxic triphosphate molecule that can be incorporated into the genome during DNA synthesis, leading to the selective ablation of proliferative cells) under the NG2 promoter, Nakano et al. [88] found that NG2 glia loss induced neurodegeneration, microglia activation and neuroinflammation in the adult hippocampus. Reduced levels of the immunomodulatory factor hepatocyte growth factor (HGF) have been proposed to mediate such effects. The different outcomes of NG2 glia ablation in the two studies may be interpreted again as the result of the loss of region-specific functions exerted by NG2 glia, or may be related to the different extent and timing of cell ablation (about 50% of NG2 glia in the cortex after seven days in [87]; about 80% in the hippocampus after one day in [88]). Milder effects reported in Birey et al. [87] may be indicative of the persistence of a sufficient number of NG2 glia contributing to neuronal support in the mouse cortex. However, even in those regions where NG2 ablation reached higher percentages (i.e., 80% in the subcortical white matter and 90% in the striatum after seven days), no sign of neurodegeneration was detected in NG2Cre-R26DTR mice. An alternative explanation could be that the huge mass of NG2 glia simultaneously undergoing cell death in [88] may have triggered a robust microglia response that eventually impacted on hippocampal neuron survival per se (i.e., independently of the possible effects of NG2 glia loss); further studies will clarify this issue. In any case, the inconsistency of the results of these two studies points to the intrinsic limitations of all cell ablation approaches, that must be taken into account in the interpretation of the results obtained by these experimental strategies.
A third recent study [42] used three different methods (i.e., X-irradiation, AraC infusion into the third ventricle and a novel mouse line where Esco2, a protein necessary for cell cycle progression, could be deleted in NG2 glia, inducing cell death in cycling NG2 cells; see also [12]) to ablate NG2 glia in the adult mouse median eminence. Here, NG2 glia ablation caused the degeneration of the dendrites of hypothalamic neurons expressing the leptin receptor. This led to the loss of neuronal responsiveness to leptin, and consequentially, to mouse overeating and obesity. These data showed a specific NG2 glia contribution to the body weight control, likely exerted by providing trophic or structural support to neuronal dendrites.
Cell ablation strategies have also been used to investigate NG2 glia roles during CNS ontogenesis. Recently, Minocha et al. [63] developed two Cre-mouse lines expressing a ’floxed’ diphtheria toxin gene under either the Nkx2.1 (Nkx2.1-Cre -R26DTA) or the NG2 (Cspg4-Cre-R26DTA) promoter to ablate the first wave of NG2 glia produced in the forebrain during the embryonic life. In the mouse, these cells originate at E12.5 from Nkx2.1-expressing progenitors of the medial ganglionic eminence and anterior entopeduncular area, and transiently populate the entire telencephalon by E14.5 before disappearing at around postnatal day eight (P8) [84,89,90]. Thus, they do not contribute to myelination and their transient nature raises numerous questions about their possible role/s during CNS development. Interestingly, the ablation of such first wave of NG2 glia severely affected the formation of the blood vessel network by reducing vascular ramifications and connections during the embryonic life. Of note, analyses at postnatal stages did not reveal any such defects, indicating compensatory proangiogenic actions of later appearing NG2 glia or other cell types.
In another study, the ablation of NG2 glia in a transgenic myelin basic protein (MBP)-TK mouse line during the early phases of postnatal development (i.e., at P1) resulted in a rapid increase of axonal sprouting in the cerebellum, changes in the expression of molecules involved in axon plasticity and guidance (i.e., increased levels of the growth-associated protein GAP43, reduced expression of Semaphorin 3a and Netrin 1), and altered localization and function of ionotropic glutamate receptors in Purkinje neurons [91]. These studies showed that during the postnatal and embryonic life, NG2 glia populations are engaged in developmental events well before the onset of myelination, and indicated their active participation in shaping neuronal circuits and blood vessel networks.
5. NG2 Glia-Derived Signals Can Modulate Neuronal and Non-Neuronal Cell Functions in the Central Nervous System
Traditionally, NG2 glia have been considered only as a target for factors produced by neighboring cells: a large repertoire of paracrine signals produced by neurons, astrocytes, microglia, and endothelial cells has been shown to influence NG2 glia during myelination and after injury [68,92]. However, accumulating evidence shows that, depending on the context, NG2 glia can serve as a source for a plethora of secreted molecules which are able to support or interfere with the functions of other CNS cell types. Namely, Sypecka and Sarnowska [93] reported a neuroprotective effect of rat NG2 glia conditioned medium (CM) on organotypic hippocampal slices subjected to oxygen-glucose deprivation. Consistently, rat embryonic cortical neurons showed a marked increase in survival when co-cultured with NG2 glia or exposed to their CM [94,95]. Further, the CM of NG2 glia displayed remarkable pro-angiogenic effects by stimulating proliferation, tip sprouting, tube formation, and expressions of tight-junctions proteins in endothelial cells [65,96,97] and promoting proliferation/survival of pericytes [66]. In contrast, the CM obtained from NG2 glia exposed to inflammatory cytokines weakened endothelial barrier tightness in vitro [67]. These findings indicated that, overall, NG2 glia can provide functionally relevant paracrine signals. However, mechanistic insights about (i) their molecular identity, (ii) the spatio-temporal regulation of their production/release, and (iii) their targeting and relevance in vivo, started to be unveiled only very recently. Indeed, the physical apposition of the tiny NG2 glia processes at specific sites of neuronal and non-neuronal CNS cells suggests a focal release and a precise targeting of NG2 glia-derived signals. Synaptophysin-positive clusters indicative of a ‘classical’ secretory vesicle accumulation and release machinery have been observed at defined sites along the NG2 glia processes [41]. Other mechanisms of signal transfer have been also reported, including cleavage of fragments of membrane-bound molecules [98] or exosome shedding [99]. Signal transfer through exosomes allows cell-specific targeting by receptor-ligand interactions [100] and assures that a well defined concentration of the signal may be delivered to the target. Thus, this latter form of communication again suggests a specific control of the distribution and intensity of the biological effects elicited by NG2-derived signals.
As regards the molecular identity of NG2 glia secreted factors, in vitro/in vivo analyses of gene expression or of NG2 glia CM showed that they include growth factors, neurotrophins, neuromodulatory and neurosupportive signals, cell adhesion and extracellular matrix (ECM) molecules, matrix metalloproteases and metalloprotease inhibitors, inflammatory cytokines/immunomodulatory factors, morphogens (Table 1).
It is noteworthy that NG2 glia express and secrete a large repertoire of ‘neuroactive’ factors, that can contribute to regulate neuronal survival, maturation and functions. The bioactivity and functional relevance of some these signals have been validated by in vitro or in vivo approaches. Namely, the neutralization of either brain derived growth factor (BDNF) or interleukin (Il)-10 in NG2 glia CM showed that these factors contribute to NG2 glia neuroprotective function in vitro [93]. Birey and colleagues showed that the specific knock-down of fibroblast growth factor 2 (FGF2) in the NG2 glia of the prefrontal cortex suffices to induce depressive-like behaviors in mice, and suggested that the loss of this signal mediates the effects of NG2 cell ablation on glutamatergic transmission [87]. Similarly, by pharmacological and lentiviral inhibition of the retinoic acid (RA) synthesis, Goncalves et al., showed that NG2 glia support axon growth and regeneration by contributing to the conversion of retinal to RA [99].
The enrichment of ECM components and matrix metalloproteases (Table 1) suggests unique interactions of NG2 glia with the ECM compared with other CNS cell types, and specific roles in tissue maintenance and remodeling. Beside this, the regulation of ECM integrity and composition is important for neurotransmitter receptor clustering at synapses [200,201]. Of note, a relatively large repertoire of cell adhesion and ECM-associated molecules expressed by NG2 glia are well known secreted pro-synaptogenic factors and synaptic organizers (e.g., thrombospondins, cerebellin 1, olfactomedin 2, neurexophilin 1, neuronal pentraxin 2). These components may be instrumental to build and maintain neuron-to-NG2 glia synaptic contacts. However, since NG2 glia processes have been observed in close apposition to neuronal dendrites and somata or interdigitating within the synaptic cleft of neuron-to-neuron synapses, this enrichment may be also related to a role of NG2 glia in neuronal synaptogenesis and synaptic plasticity. Similarly, ECM molecules expressed by NG2 glia (i.e., brevican, versican, tenascin R) importantly contribute to the assembly, maintenance and function of the nodes of Ranvier [46], thereby suggesting a specific NG2 glia contribution to neuronal maturation and action potential transmission.
In this context, recent data indicated that the NG2 chondroitin sulfate proteoglycan, whose expression is restricted to NG2 glia among the neural cell types, can serve as a polyvalent regulator of neuronal synaptic activity and action potential conduction. Sakry et al. [98] showed that the ectodomain of the NG2 protein, constitutively released from the NG2 glia surface into the ECM by the ADAM10 secretase, influences AMPA receptor currents and kinetics at glutamatergic synapses of cortical pyramidal neurons. Moreover, the pharmacological inhibition of NG2 cleavage and release from NG2 glia resulted in a large reduction of NMDAR-dependent, long-term potentiation (LTP) at these synapses, indicating a specific role of NG2 in synaptic plasticity [98]. A further neuromodulatory function of NG2 has been attributed to its intracellular domain that regulates the expression of prostaglandin D2 synthase, a secreted enzyme that catalyzes the conversion of prostaglandin H2 to the neuromodulatory form prostaglandin D2 [154]. Finally, in the injured spinal cord, secreted NG2 has been observed to block action potential conduction at the nodes of Ranvier [202].
Transforming growth factor β (TGF-β) and Wingless-type MMTV integration site family 7 (Wnt7) have recently been shown to mediate NG2 glia pro-angiogenetic effects in vivo. The pharmacological inhibition of TGF-b signaling abolished the effect of NG2 glia CM on endothelial cell expression of tight junction proteins in vitro. Consistently, the specific ablation of TGF-b1 in NG2 glia resulted in cerebral hemorrhage and loss of blood brain barrier integrity in neonatal mice [65]. Similar approaches have been used by Yuen et al. [96] to demonstrate that NG2 glia-derived Wnt7 mediates their pro-angiogenic effects in vitro.
Notably, as a part of their plastic nature, NG2 glia dynamically modulate the expression/secretion of specific signals in response to external factors. Importantly, signals inducing NG2 glia depolarization reportedly affected their secretion. Exposure to GABA largely increased the production and secretion of BDNF from adult rat NG2 glia [113]. Consistently, the application of a moderate intensity static magnetic field, with its associated electric field, on human NG2 glia cultures resulted in increased secretion of BDNF and neurotrophin 3 (NT-3) [114]. The release of the NG2 ectodomain from NG2 glia was augmented by treatments with depolarizing agents or glutamate in vitro [98]. Along this line, the expression of the neuromodulatory/neuroprotective peptide galanin significantly increased in NG2 glia upon experimental cortical spreading depression [169]. These observations suggest that activity dependent/synapse-mediated signals converging on NG2 glia may dynamically regulate their secretory activity and homeostatic functions.
In conclusion, although the repertoire of factors expressed/secreted by NG2 glia has not yet been explored in depth, these findings clearly indicate that they can serve as providers of relevant signals for neighboring cells—in particular for neurons—in a context-dependent manner.
6. NG2 Glia Upon Central Nervous System Injury and Stress
The cellular roles of NG2 glia upon pathological conditions remain largely unclear. Notably, in vivo/in vitro exposure to different kinds of stresses or injury-related signals remarkably affects NG2 glia expression profile and secretion. Some of these changes can be interpreted as compensatory attempts aimed at sustaining cell survival/functions and restoring the homeostasis. For example, early after an acute brain injury, such as an ischemic event, BDNF protein expression largely increased in NG2 glia, likely amplifying their neuroprotective potential [113]. However, when the exposure to danger signals or stresses takes place over longer periods, gene expression/secretion alterations in NG2 glia make them sustain or even worsen the pathological condition. In vivo exposure to chronic stress induces profound transcriptional alterations in NG2 glia in specific brain regions (i.e., prefrontal cortex and nucleus accumbens [87,203]), and largely reduces the levels of growth factors (i.e., FGF2) and neurotrophins (i.e., NGF and NT4/5) [87]. After prolonged cerebral hypoperfusion stress, perivascular NG2 glia in the adult white matter increased matrix metalloprotease 9 (MMP9) expression and contributed to blood brain barrier opening [67]. Treatment with sublethal concentrations of the inflammatory cytokine Il-1β also increased MMP9 secretion by cultured NG2 glia [67]. Similarly, exposure to Il-17 or Tnf increased MMP9 and MMP3 expression and elicited a robust inflammatory response in cultured NG2 glia [158]. Consistently, sustained Il-17 signaling in NG2 glia incorporated these cells in the inflammatory pathogenesis of experimental autoimmune encephalomyelitis (EAE) [158]. Complementarily, experimental attenuation of NG2 glia proliferative response after spinal cord lesion was associated with reduced accumulation of activated microglia/macrophages, diminished astrocyte hypertrophy, and eventually with the establishment of a post-injury environment more supportive for tissue repair [204]. This suggests a role for NG2 cells in orchestrating reactive gliosis.
Thus, beyond their role in myelin repair and glial scar formation [205], depending on the circumstances, NG2 glia have been observed either to limit CNS damage or to actively contribute to neuroinflammation/neurotoxicity, as also shown in Amyotrophic Lateral Sclerosis models [206]. To further increase the complexity of this scenario, consistent with previous findings [207,208], in vivo two-photon imaging analyses revealed an assortment of distinct NG2 glia reactions to injury, with some cells starting migration toward the lesion site, others entering cell cycle, and others displaying only hypertrophy and morphological changes [31,86]. Single-cell messenger-RNA (mRNA) profiling of NG2 glia after focal cerebral ischemia also identified at least three subpopulations of cells that could be distinguished according to changes in their expression patterns [19]. Such behavioral and molecular heterogeneity suggests the existence of diverse NG2 glia subsets which respond differently to CNS injury.
7. Concluding Remarks and Open Issues
In this review, we have summarized the advancements obtained over recent years toward the understanding of the roles of NG2 glia in CNS homeostasis and development. Though it is widely accepted that these cells act as precursors for OLs in the developing and mature CNS, accumulating evidence indicates that NG2 glia (or subsets of NG2 glia) can exert additional/alternative functions. Recent studies have shown that NG2 glia establish physical interactions with the other CNS cell types, are integrated in neuronal networks, and possess a repertoire of ion channels and neurotransmitter receptors apt to constantly monitor the activity of the surrounding neurons. In addition to operating as sensors, NG2 glia have features that are expected for active modulators of the neuronal activity, including the expression and release of a complex array of neuromodulatory and neuroprotective factors. Consistently, cell ablation strategies targeting NG2 glia demonstrate that the maintenance of their density contributes to the normal function and development of different CNS regions, beyond their role in myelination.
Although showing some similarities with astrocytes—as regards their physical/functional relationships with neurons and blood vessels, and to a certain extent, gene expression—data available so far point to a distinct contribution of these two glial cell types in CNS homeostasis. NG2 glia specificity relies on the ability to monitor, and perhaps modulate, the activity of the neuronal networks with a resolution in time, space and intensity that cannot be achieved by astroglia. This appears to be related to NG2 glia typical morphology (with fine processes establishing contacts at well defined sites onto neurons) and functional properties (including the ability to sense neuronal activity at the quantal level through neuron-to-NG2 glia synapses). Further, since the roles/features of NG2 glia described above do not emerge as the by-product of the metabolic/gene expression control of typical progenitor functions (i.e., proliferation and differentiation in myelin-producing cells), they cannot simply be interpreted as epiphenomena (i.e., secondary functions) of the role of NG2 glia as OL precursors. Thus, NG2 glia not only represent a transitional stage along the oligodendroglial lineage, but rather, a specific glial cell type endowed with typical properties and functions.
However, this field of research is still in its infancy and the mechanistic nature of NG2 glia functions beyond re-/myelination remains still unclear. Further efforts are needed to overcome the limitations of the experimental strategies employed so far, and to identify the molecular and cellular processes subserving NG2 glia participation in CNS homeostasis, ontogeny, and disease. Namely, the in vivo NG2 glia ablation approaches showed important intrinsic caveats, e.g., the achievement of a partial and transient cell ablation due to the fast repopulation of NG2 glia; region-specific differences in the ablation efficiency due to different cell cycle dynamics and repopulation efficiency of NG2 glia in distinct CNS areas [87]; selective/predominant ablation of specific NG2 glia subsets (i.e., those that are more prone to enter cell cycle; [42,88]); possible interference/superimposition of the reactions of other CNS cell types responding to NG2 glia loss [88]. Such limitations restrict the time window/extent/types of phenotypes that could be attributed to NG2 glia loss, and may also hamper investigation into the underlying molecular mechanisms. Recent studies have started to unveil the cellular processes through which NG2 glia can transfer functionally relevant signals (i.e., conventional vesicle release, exosome shedding, cleavage of extracellular fragments of membrane-bound proteins). Targeting such processes (e.g., by a genetic-based deletion of specific components of the secretory/shedding/cleavage machineries in NG2 glia) may provide an alternative experimental setting to unveil the impact and relevance of NG2 glia to other cell types communication. Further, validation, perhaps using cell type-specific subtractive/deletion approaches (as in [87]), of the effects exerted by NG2 glia-derived factors in vivo is required to define the regulatory signals that are truly unique in mediating NG2 glia functions. The identification of such factors has recently taken advantage of RNA sequencing technologies [131]. The progresses of these strategies will make it possible to study the transcriptomic landscape at the single cell level, thereby possibly unveiling also the heterogeneous nature of NG2 glia [209]. Moreover, since these cells establish contacts with neurons and other cell types at specific sites along processes and somata, and local mRNA localization and translation may be instrumental for their crosstalk (as recently shown for astrocytes, [210]), we envisage that novel approaches of spatial transcriptomics may be also fruitfully exploited to perform high-throughput quantifications while preserving spatial information about the subcellular localization of the analysed transcripts [211].
Indeed, such studies may also open translational perspectives for the implementation of the endogenous neurosupportive/neuroprotective potential of the CNS. In this context, more research is also needed to fully understand NG2 glia participation in pathology. A detailed and comprehensive examination of NG2 glia behaviors/molecular changes upon lesion will likely unveil some elements of cell heterogeneity (see above). Indeed, diverse NG2 glia subpopulations have also been described in the intact CNS based on their origin, proliferative activity, division modality, differentiation potential, expression of diverse transcription factors, Ca2+-binding molecules, neurotransmitter receptors, and ionic currents [5,23,48,69,212]. Thus, it is conceivable that subsets of functionally specialized NG2 glia may be differentially tailored to contribute to distinct aspects of CNS homeostasis, including oligodendrogenesis and additional/alternative functions. This issue deserves to be the focus of future investigations, and adds a further element of complexity on the functioning of this fascinating cell type.
Funding
Our work is supported by the “Cariplo Ricerca Biomedica Giovani Ricercatori” grant from the Cariplo Foundation (ID: 2014-1207) and Individual funding for basic research (Ffabr) granted by the Italian Agency for the Evaluation of University and Research, to EB. This study was also supported by Ministero dell’Istruzione, dell’Università e della Ricerca—MIUR project “Dipartimenti di Eccellenza 2018–2022” to Dept. of Neuroscience “Rita Levi Montalcini”.
Acknowledgments
We thank Annalisa Buffo (University of Turin) for insightful comments.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the interpretation of data or in the writing of the manuscript.
References
- Boda, E.; Buffo, A. Glial cells in non-germinal territories: Insights into their stem/progenitor properties in the intact and injured nervous tissue. Arch. Ital. Biol. 2010, 148, 119–136. [Google Scholar] [PubMed]
- Dawson, M.R.L.; Polito, A.; Levine, J.M.; Reynolds, R. NG2-expressing glial progenitor cells: An abundant and widespread population of cycling cells in the adult rat CNS. Mol. Cell. Neurosci. 2003, 24, 476–488. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Plemel, J.R.; Lee, M.J.; Stratton, J.A.; Manesh, S.B.; Liu, J.; Ramer, L.M.; Kang, S.H.; Bergles, D.E.; et al. Myelinogenic plasticity of oligodendrocyte precursor cells following spinal cord contusion injury. J. Neurosci. 2017, 37, 8635–8654. [Google Scholar] [CrossRef] [PubMed]
- Baxi, E.G.; DeBruin, J.; Jin, J.; Strasburger, H.J.; Smith, M.D.; Orthmann-Murphy, J.L.; Schott, J.T.; Fairchild, A.N.; Bergles, D.E.; Calabresi, P.A. Lineage tracing reveals dynamic changes in oligodendrocyte precursor cells following cuprizone-induced demyelination. Glia 2017, 65, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Boda, E.; Di Maria, S.; Rosa, P.; Taylor, V.; Abbracchio, M.P.; Buffo, A. Early phenotypic asymmetry of sister oligodendrocyte progenitor cells after mitosis and its modulation by aging and extrinsic factors. Glia 2015, 63, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.; Patel, K.D.; Goncalves, C.M.; Grutzendler, J.; Nishiyama, A. Modulation of oligodendrocyte generation during a critical temporal window after NG2 cell division. Nat. Neurosci. 2014, 17, 1518–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, I.A.; Ohayon, D.; Li, H.; Paes de Faria, J.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Ohayon, D.; McKenzie, I.A.; Sinclair-Wilson, A.; Wright, J.L.; Fudge, A.D.; Emery, B.; Li, H.; Richardson, W.D. Rapid production of new oligodendrocytes is required in the earliest stages of motor-skill learning. Nat. Neurosci. 2016, 19, 1210–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, E.G.; Orthmann-Murphy, J.L.; Langseth, A.J.; Bergles, D.E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nat. Neurosci. 2018, 21, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Li, A.M.; Grutzendler, J. Lifelong cortical myelin plasticity and age-related degeneration in the live mammalian brain. Nat. Neurosci. 2018, 21, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Young, K.M. White matter plasticity in adulthood. Neuroscience 2014, 276, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Gruart, A.; Grade, S.; Zhang, Y.; Kröger, S.; Kirchhoff, F.; Eichele, G.; Delgado García, J.M.; Dimou, L. Decrease in newly generated oligodendrocytes leads to motor dysfunctions and changed myelin structures that can be rescued by transplanted cells. Glia 2016, 64, 2201–2218. [Google Scholar] [CrossRef] [PubMed]
- Psachoulia, K.; Jamen, F.; Young, K.M.; Richardson, W.D. Cell cycle dynamics of NG2 cells in the postnatal and ageing brain. Neuron Glia Biol. 2009, 5, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boda, E.; Buffo, A. Beyond cell replacement: Unresolved roles of NG2-expressing progenitors. Front. Neurosci. 2014, 8, 122. [Google Scholar] [CrossRef] [PubMed]
- Birey, F.; Kokkosis, A.; Aguirre, A. Oligodendroglia-lineage cells in brain plasticity, homeostasis and psychiatric disorders. Curr. Opin. Neurobiol. 2017, 47, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Boda, E.; Viganò, F.; Rosam, P.; Fumagalli, M.; Labat-Gest, V.; Tempia, F.; Abbracchio, M.P.; Dimou, L.; Buffo, A. The GPR17 receptor in NG2 expressing cells: Focus on in vivocell maturation and participation in acute trauma and chronic damage. Glia 2011, 59, 1958–1973. [Google Scholar] [CrossRef] [PubMed]
- Levine, J. The reactions and role of NG2 glia in spinal cord injury. Brain Res. 2016, 1638, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Riew, T.R.; Kim, H.L.; Choi, J.H.; Lee, M.Y. Morphological characterization of NG2 glia and their association with neuroglial cells in the 3-nitropropionic acid–lesioned striatum of rat. Sci. Rep. 2018, 8, 5942. [Google Scholar] [CrossRef] [PubMed]
- Valny, M.; Honsa, P.; Waloschkova, E.; Matuskova, H.; Kriska, J.; Kirdajova, D.; Androvic, P.; Valihrach, L.; Kubista, M.; Anderova, M. A single-cell analysis reveals multiple roles of oligodendroglial lineage cells during post-ischemic regeneration. Glia 2018, 66, 1068–1081. [Google Scholar] [CrossRef] [PubMed]
- Kazanis, I.; Evans, K.A.; Andreopoulou, E.; Dimitriou, C.; Koutsakis, C.; Karadottir, R.T.; Franklin, R.J.M. Subependymal zone-derived oligodendroblasts respond to focal demyelination but fail to generate myelin in young and aged mice. Stem Cell Rep. 2017, 8, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, C.; Praet, J.; Rangarajan, J.R.; Vreys, R.; De Vocht, N.; Maes, F.; Verhoye, M.; Ponsaerts, P.; Van der Linden, A. Multimodal imaging of subventricular zone neural stem/progenitor cells in the cuprizone mouse model reveals increased neurogenic potential for the olfactory bulb pathway, but no contribution to remyelination of the corpus callosum. Neuroimage 2014, 86, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.L.; Roth, P.T.; Stratton, J.A.S.; Chuang, B.H.A.; Danne, J.; Ellis, S.L.; Ng, S.W.; Kilpatrick, T.J.; Merson, T.D. Adult neural precursor cells from the subventricular zone contribute significantly to oligodendrocyte regeneration and remyelination. J. Neurosci. 2014, 34, 14128–14146. [Google Scholar] [CrossRef] [PubMed]
- Van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Hill, R.A.; Nishiyama, A. NG2 cells generate oligodendrocytes and gray matter astrocytes in the spinal cord. Neuron Glia Biol. 2009, 4, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Bergles, D.E.; Nishiyama, A. NG2 cells generate both oligodendrocytes and gray matter astrocytes. Development 2007, 135, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Zhao, N.; Bai, X.; Karram, K.; Trotter, J.; Goebbels, S.; Scheller, A.; Kirchhoff, F. Novel NG2-CreERT2 knock-in mice demonstrate heterogeneous differentiation potential of NG2 glia during development. Glia 2014, 62, 896–913. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.B.; Rivers, L.E.; Young, K.M.; Jamen, F.; Richardson, W.D. NG2 glia generate new oligodendrocytes but few astrocytes in a murine experimental autoimmune encephalomyelitis model of demyelinating disease. J. Neurosci. 2010, 30, 16383–16390. [Google Scholar] [CrossRef] [PubMed]
- Boda, E.; Nato, G.; Buffo, A. Emerging pharmacological approaches to promote neurogenesis from endogenous glial cells. Biochem. Pharmacol. 2017, 141, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.; Wood, W.M.; Sherafat, A.; Hill, R.A.; Lu, Q.R.; Nishiyama, A. Age-dependent decline in fate switch from NG2 cells to astrocytes after Olig2 deletion. J. Neurosci. 2018, 38, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Valny, M.; Honsa, P.; Kriska, J.; Anderova, M. Multipotency and therapeutic potential of NG2 cells. Biochem. Pharmacol. 2017, 141, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Hughes, E.G.; Kang, S.H.; Fukaya, M.; Bergles, D.E. Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat. Neurosci. 2013, 16, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, B.B.; Takada, N.; Latimer, A.J.; Shin, J.; Carney, T.J.; Kelsh, R.N.; Appel, B. In vivo time-lapse imaging shows dynamic oligodendrocyte progenitor behavior during zebrafish development. Nat. Neurosci. 2006, 9, 1506–1511. [Google Scholar] [CrossRef] [PubMed]
- Birey, F.; Aguirre, A. Age-dependent netrin-1 signaling regulates NG2+ glial cell spatial homeostasis in normal adult gray matter. J. Neurosci. 2015, 35, 6946–6951. [Google Scholar] [CrossRef] [PubMed]
- Dufour, A.; Gontran, E.; Deroulers, C.; Varlet, P.; Pallud, J.; Grammaticos, B.; Badoual, M. Modeling the dynamics of oligodendrocyte precursor cells and the genesis of gliomas. PLoS Comput. Biol. 2018, 14, e1005977. [Google Scholar] [CrossRef] [PubMed]
- Sugiarto, S.; Persson, A.I.; Munoz, E.G.; Waldhuber, M.; Lamagna, C.; Andor, N.; Hanecker, P.; Ayers-Ringler, J.; Phillips, J.; Siu, J.; et al. Asymmetry-defective oligodendrocyte progenitors are glioma precursors. Cancer Cell 2011, 20, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Balia, M.; Benamer, N.; Angulo, M.C. A specific GABAergic synapse onto oligodendrocyte precursors does not regulate cortical oligodendrogenesis. Glia 2017, 65, 1821–1832. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkhratsky, A.; Bush, N.; Nedergaard, M.; Butt, A. The special case of human astrocytes. Neuroglia 2018, 1, 4. [Google Scholar] [CrossRef]
- Casano, A.M.; Peri, F. Microglia: Multitasking specialists of the brain. Dev. Cell 2015, 32, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Réu, P.; Khosravi, A.; Bernard, S.; Mold, J.E.; Salehpour, M.; Alkass, K.; Perl, S.; Tisdale, J.; Possnert, G.; Druid, H.; et al. The lifespan and turnover of microglia in the human brain. Cell Rep. 2017, 20, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Wigley, R.; Butt, A.M. Integration of NG2-glia (synantocytes) into the neuroglial network. Neuron Glia Biol. 2009, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Djogo, T.; Robins, S.C.; Schneider, S.; Kryzskaya, D.; Liu, X.; Mingay, A.; Gillon, C.J.; Kim, J.H.; Storch, K.F.; Boehm, U.; et al. Adult NG2-glia are required for median eminence-mediated leptin sensing and body weight control. Cell Metab. 2016, 23, 797–810. [Google Scholar] [CrossRef] [PubMed]
- März, M.; Schmidt, R.; Rastegar, S.; Strähle, U. Expression of the transcription factor Olig2 in proliferating cells in the adult zebrafish telencephalon. Dev. Dyn. 2010, 239, 3336–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, J.J.; Messier, C. Oligodendrocyte progenitor cells are paired with GABA neurons in the mouse dorsal cortex: Unbiased stereological analysis. Neuroscience 2017, 362, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.M.; Duncan, A.; Hornby, M.F.; Kirvell, S.L.; Hunter, A.; Levine, J.M.; Berry, M. Cells expressing the NG2 antigen contact nodes of Ranvier in adult CNS white matter. Glia 1999, 26, 84–91. [Google Scholar] [CrossRef]
- Serwanski, D.R.; Jukkola, P.; Nishiyama, A. Heterogeneity of astrocyte and NG2 cell insertion at the node of Ranvier. J. Comp. Neurol. 2017, 525, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Ong, W.Y.; Levine, J.M. A light and electron microscopic study of NG2 chondroitin sulfate proteoglycan-positive oligodendrocyte precursor cells in the normal and kainate-lesioned rat hippocampus. Neuroscience 1999, 92, 83–95. [Google Scholar] [CrossRef]
- Maldonado, P.P.; Angulo, M.C. Multiple modes of communication between neurons and oligodendrocyte precursor cells. Neuroscientist 2015, 21, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Orduz, D.; Maldonado, P.P.; Balia, M.; Vélez-Fort, M.; de Sars, V.; Yanagawa, Y.; Emiliani, V.; Angulo, M.C. Interneurons and oligodendrocyte progenitors form a structured synaptic network in the developing neocortex. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Huck, J.H.J.; Roberts, J.D.B.; Macklin, W.B.; Somogyi, P.; Bergles, D.E. Climbing fiber innervation of NG2-expressing glia in the mammalian cerebellum. Neuron 2005, 46, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Reyes-Haro, D.; Pivneva, T.; Nolte, C.; Schaette, R.; Lübke, J.; Kettenmann, H. The principal neurons of the medial nucleus of the trapezoid body and NG2+ glial cells receive coordinated excitatory synaptic input. J. Gen. Physiol. 2009, 134, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Mangin, J.M.; Li, P.; Scafidi, J.; Gallo, V. Experience-dependent regulation of NG2 progenitors in the developing barrel cortex. Nat. Neurosci. 2012, 15, 1192–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Bergles, D.E. Synaptic signaling between GABAergic interneurons and oligodendrocyte precursor cells in the hippocampus. Nat. Neurosci. 2004, 7, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Mangin, J.M.; Kunze, A.; Chittajallu, R.; Gallo, V. Satellite NG2 progenitor cells share common glutamatergic inputs with associated interneurons in the mouse Dentate gyrus. J. Neurosci. 2008, 28, 7610–7623. [Google Scholar] [CrossRef] [PubMed]
- Balia, M.; Vélez-Fort, M.; Passlick, S.; Schäfer, C.; Audinat, E.; Steinhäuser, C.; Seifert, G.; Angulo, M.C. Postnatal down-regulation of the GABAA receptor γ2 subunit in neocortical NG2 cells accompanies synaptic-to-extrasynaptic switch in the GABAergic transmission mode. Cereb. Cortex 2015, 25, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, N.; Vayro, S.; Wigley, R.; Butt, A.M. Axons and astrocytes release ATP and glutamate to evoke calcium signals in NG2-glia. Glia 2010, 58, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wang, W.; Zhou, M. Spatial organization of NG2 glial cells and astrocytes in rat hippocampal CA1 region. Hippocampus 2014, 24, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Yu, M.; Drazba, J.A.; Tuohy, V.K. Normal and reactive NG2+ glial cells are distinct from resting and activated microglia. J. Neurosci. Res. 1997, 48, 299–312. [Google Scholar] [CrossRef]
- Wallraff, A.; Odermatt, B.; Willecke, K.; Steinhäuser, C. Distinct types of astroglial cells in the hippocampus differ in gap junction coupling. Glia 2004, 48, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.M.; Hamilton, N.; Hubbard, P.; Pugh, M.; Ibrahim, M. Synantocytes: The fifth element. J. Anat. 2005, 207, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Melanson-Drapeau, L.; Beyko, S.; Davé, S.; Hebb, A.L.O.; Franks, D.J.; Sellitto, C.; Paul, D.L.; Bennett, S.A. Oligodendrocyte progenitor enrichment in the connexin32 null-mutant mouse. J. Neurosci. 2003, 23, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Maglione, M.; Tress, O.; Haas, B.; Karram, K.; Trotter, J.; Willecke, K.; Kettenmann, H. Oligodendrocytes in mouse corpus callosum are coupled via gap junction channels formed by connexin47 and connexin32. Glia 2010, 58, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Minocha, S.; Valloton, D.; Brunet, I.; Eichmann, A.; Hornung, J.P.; Lebrand, C. NG2 glia are required for vessel network formation during embryonic development. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.H.; Niu, J.; Munji, R.; Davalos, D.; Chang, J.; Zhang, H.; Tien, A.C.; Kuo, C.J.; Chan, J.R.; Daneman, R.; et al. Oligodendrocyte precursors migrate along vasculature in the developing nervous system. Science 2016, 351, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.H.; Maki, T.; Maeda, M.; Miyamoto, N.; Liang, A.C.; Hayakawa, K.; Pham, L.D.; Suwa, F.; Taguchi, A.; Matsuyama, T.; et al. Oligodendrocyte precursor cells support blood-brain barrier integrity via TGF-β signaling. PLoS ONE 2014, 9, e103174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, T.; Maeda, M.; Uemura, M.; Lo, E.K.; Terasaki, Y.; Liang, A.C.; Shindo, A.; Choi, Y.K.; Taguchi, A.; Matsuyama, T.; et al. Potential interactions between pericytes and oligodendrocyte precursor cells in perivascular regions of cerebral white matter. Neurosci. Lett. 2015, 597, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.H.; Miyamoto, N.; Hayakawa, K.; Pham, L.D.D.; Maki, T.; Ayata, C.; Kim, K.W.; Lo, E.H.; Arai, K. Oligodendrocyte precursors induce early blood-brain barrier opening after white matter injury. J. Clin. Investig. 2013, 123, 782–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, N.; Pham, L.D.D.; Seo, J.H.; Kim, K.W.; Lo, E.H.; Arai, K. Crosstalk between cerebral endothelium and oligodendrocyte. Cell. Mol. Life Sci. 2014, 71, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Nishiyama, A. NG2 cells (polydendrocytes): Listeners to the neural network with diverse properties. Glia 2014, 62, 1195–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, D.; Eto, K.; Nabekura, J.; Wake, H. Activity-dependent functions of non-electrical glial cells. J. Biochem. 2018, 163, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A.; Raff, M.C. Proliferation of oligodendrocyte precursor cells depends on electrical activity in axons. Nature 1993, 361, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Brus-Ramer, M.; Martin, J.H.; McDonald, J.W. Electrical stimulation of the medullary pyramid promotes proliferation and differentiation of oligodendrocyte progenitor cells in the corticospinal tract of the adult rat. Neurosci. Lett. 2010, 479, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, E.M.; Purger, D.; Mount, C.W.; Goldstein, A.K.; Lin, G.L.; Wood, L.S.; Inema, I.; Miller, S.E.; Bieri, G.; Zuchero, J.B.; et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science 2014, 344, 1252304. [Google Scholar] [CrossRef] [PubMed]
- Ehninger, D.; Wang, L.P.; Klempin, F.; Römer, B.; Kettenmann, H.; Kempermann, G. Enriched environment and physical activity reduce microglia and influence the fate of NG2 cells in the amygdala of adult mice. Cell Tissue Res. 2011, 345, 69–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, C.; Götz, M.; Dimou, L. Progenitors in the adult cerebral cortex: Cell cycle properties and regulation by physiological stimuli and injury. Glia 2011, 59, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, L.; Huang, P.H.; Colognato, H. Prefrontal cortex NG2 glia undergo a developmental switch in their responsiveness to exercise. Dev. Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nagy, B.; Hovhannisyan, A.; Barzan, R.; Chen, T.J.; Kukley, M. Different patterns of neuronal activity trigger distinct responses of oligodendrocyte precursor cells in the corpus callosum. PLoS Biol. 2017, 15, e2001993. [Google Scholar] [CrossRef] [PubMed]
- Larson, V.A.; Zhang, Y.; Bergles, D.E. Electrophysiological properties of NG2+ cells: Matching physiological studies with gene expression profiles. Brain Res. 2016, 1638, 138–160. [Google Scholar] [CrossRef] [PubMed]
- Ziskin, J.L.; Nishiyama, A.; Rubio, M.; Fukaya, M.; Bergles, D.E. Vesicular release of glutamate from unmyelinated axons in white matter. Nat. Neurosci. 2007, 10, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, P.P.; Velez-Fort, M.; Levavasseur, F.; Angulo, M.C. Oligodendrocyte precursor cells are accurate sensors of local K+ in mature gray matter. J. Neurosci. 2013, 33, 2432–2442. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Matthews, E.A.; Nicolas, V.; Schoch, S.; Dietrich, D. NG2 glial cells integrate synaptic input in global and dendritic calcium signals. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Hines, J.H.; Ravanelli, A.M.; Schwindt, R.; Scott, E.K.; Appel, B. Neuronal activity biases axon selection for myelination in vivo. Nat. Neurosci. 2015, 18, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mensch, S.; Baraban, M.; Almeida, R.; Czopka, T.; Ausborn, J.; El Manira, A.; Lyons, D.A. Synaptic vesicle release regulates myelin sheath number of individual oligodendrocytes in vivo. Nat. Neurosci. 2015, 18, 628–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, H.; Lee, P.R.; Fields, R.D. Control of local protein synthesis and initial events in myelination by action potentials. Science 2011, 333, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Haberlandt, C.; Derouiche, A.; Wyczynski, A.; Haseleu, J.; Pohle, J.; Karram, K.; Trotter, J.; Seifert, G.; Frotscher, M.; Steinhäuser, C.; et al. Gray matter NG2 cells display multiple Ca2+-signaling pathways and highly motile processes. PLoS ONE 2011, 6, e17575. [Google Scholar] [CrossRef] [PubMed]
- Jäkel, S.; Dimou, L. Glial cells and their function in the adult brain: A journey through the history of their ablation. Front. Cell. Neurosci. 2017, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Birey, F.; Kloc, M.; Chavali, M.; Hussein, I.; Wilson, M.; Christoffel, D.J.; Chen, T.; Frohman, M.A.; Robinson, J.K.; Russo, S.J.; et al. Genetic and stress-induced loss of NG2 glia triggers emergence of depressive-like behaviors through reduced secretion of FGF2. Neuron 2015, 88, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Tamura, Y.; Yamato, M.; Kume, S.; Eguchi, A.; Takata, K.; Watanabe, Y.; Kataoka, Y. NG2 glial cells regulate neuroimmunological responses to maintain neuronal function and survival. Sci. Rep. 2017, 7, 42041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessaris, N.; Fogarty, M.; Iannarelli, P.; Grist, M.; Wegner, M.; Richardson, W.D. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nat. Neurosci. 2006, 9, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Van Tilborg, E.; Achterberg, E.J.M.; van Kammen, C.M.; van der Toorn, A.; Groenendaal, F.; Dijkhuizen, R.M.; Heijnen, C.J.; Vanderschuren, L.J.M.J.; Benders, M.N.J.L.; Nijboer, C.H.A. Combined fetal inflammation and postnatal hypoxia causes myelin deficits and autism-like behavior in a rat model of diffuse white matter injury. Glia 2018, 66, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Doretto, S.; Malerba, M.; Ramos, M.; Ikrar, T.; Kinoshita, C.; De Mei, C.; Tirotta, E.; Xu, X.; Borrelli, E. Oligodendrocytes as regulators of neuronal networks during early postnatal development. PLoS ONE 2011, 6, e19849. [Google Scholar] [CrossRef] [PubMed]
- Clemente, D.; Ortega, M.C.; Melero-Jerez, C.; de Castro, F. The effect of glia-glia interactions on oligodendrocyte precursor cell biology during development and in demyelinating diseases. Front. Cell. Neurosci. 2013, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Sypecka, J.; Sarnowska, A. The neuroprotective effect exerted by oligodendroglial progenitors on ischemically impaired hippocampal cells. Mol. Neurobiol. 2014, 49, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.; Chandran, S.; Compston, A. A role for oligodendrocyte-derived IGF-1 in trophic support of cortical neurons. Glia 2001, 36, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.; Majed, H.; Layfield, R.; Compston, A.; Chandran, S. Oligodendrocytes promote neuronal survival and axonal length by distinct intracellular mechanisms: A novel role for oligodendrocyte-derived glial cell line-derived neurotrophic factor. J. Neurosci. 2003, 23, 4967–4974. [Google Scholar] [CrossRef] [PubMed]
- Yuen, T.J.; Silbereis, J.C.; Griveau, A.; Chang, S.M.; Daneman, R.; Fancy, S.P.J.; Zahed, H.; Maltepe, E.; Rowitch, D.H. Oligodendrocyte-encoded HIF function couples postnatal myelination and white matter angiogenesis. Cell 2014, 158, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xiao, L.; Gong, X.; Shao, W.; Yin, Y.; Liao, Q.; Meng, Y.; Zhang, Y.; Ma, D.; Qiu, X. Cytokine-like molecule CCDC134 Contributes to CD8+ T-cell effector functions in cancer immunotherapy. Cancer Res. 2014, 74, 5734–5745. [Google Scholar] [CrossRef] [PubMed]
- Sakry, D.; Neitz, A.; Singh, J.; Frischknecht, R.; Marongiu, D.; Binamé, F.; Perera, S.S.; Endres, K.; Lutz, B.; Radyushkin, K.; et al. Oligodendrocyte precursor cells modulate the neuronal network by activity-dependent ectodomain cleavage of glial NG2. PLoS Biol. 2014, 12, e1001993. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, M.B.; Wu, Y.; Trigo, D.; Clarke, E.; Malmqvist, T.; Grist, J.; Hobbs, C.; Carlstedt, T.P.; Corcoran, J.P.T. Retinoic acid synthesis by NG2 expressing cells promotes a permissive environment for axonal outgrowth. Neurobiol. Dis. 2018, 111, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.H.; Shin, U.S.; Kim, H.W. Fibroblast growth factors: Biology, function, and application for tissue regeneration. J. Tissue Eng. 2010, 2010, 218142. [Google Scholar] [CrossRef] [PubMed]
- Zechel, S.; Unsicker, K.; von Bohlen und Halbach, O. Fibroblast growth factor-2 deficiency affects hippocampal spine morphology, but not hippocampal catecholaminergic or cholinergic innervation. Dev. Dyn. 2009, 238, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, D.R.; Berry, M.; Kirvell, S.L.; Butt, A.M. Fibroblast growth factor-2 induces astroglial and microglial reactivity in vivo. J. Anat. 2002, 200, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.; Mattson, M.P. PDGFs protect hippocampal neurons against energy deprivation and oxidative injury: Evidence for induction of antioxidant pathways. J. Neurosci. 1995, 15, 7095–7104. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Yao, H.; Bai, X.; Zhu, X.; Reiner, B.C.; Beazely, M.; Funa, K.; Xiong, H.; Buch, S. Platelet-derived growth factor-mediated induction of the synaptic plasticity gene Arc/Arg3.1. J. Biol. Chem. 2010, 285, 21615–21624. [Google Scholar] [CrossRef] [PubMed]
- Risau, W.; Drexler, H.; Mironov, V.; Smits, A.; Siegbahn, A.; Funa, K.; Heldin, C.H. Platelet-derived growth factor is angiogenic in vivo. Grow. Factors 1992, 7, 261–266. [Google Scholar] [CrossRef]
- Sasahara, A.; Kott, J.N.; Sasahara, M.; Raines, E.W.; Ross, R.; Westrum, L.E. Platelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brain. Dev. Brain Res. 1992, 68, 41–53. [Google Scholar] [CrossRef]
- Rosenstein, J.M.; Krum, J.M.; Ruhrberg, C. VEGF in the nervous system. Organogenesis 2010, 6, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovar-y-Romo, L.B.; Tapia, R. Delayed administration of VEGF rescues spinal motor neurons from death with a short effective time frame in excitotoxic experimental models in vivo. ASN Neuro 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Estévez, V.; Defterali, Ç.; Vicario-Abejón, C. IGF-I: A key growth factor that regulates neurogenesis and synaptogenesis from embryonic to adult stages of the brain. Front. Neurosci. 2016, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Rivkees, S.A. Hepatocyte growth factor stimulates the proliferation and migration of oligodendrocyte precursor cells. J. Neurosci. Res. 2002, 69, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Funakoshi, H.; Kadoyama, K.; Noma, S.; Kanai, M.; Ohya-Shimada, W.; Mizuno, S.; Doe, N.; Taniguchi, T.; Nakamura, T. Hepatocyte growth factor overexpression in the nervous system enhances learning and memory performance in mice. J. Neurosci. Res. 2012, 90, 1743–1755. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Tozuka, Y.; Takata, T.; Shimazu, N.; Matsumura, N.; Ohta, A.; Hisatsune, T. Excitatory GABAergic activation of cortical dividing glial cells. Cereb. Cortex 2009, 19, 2181–2195. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; The, D.B.L.; Blasiak, A.; Chai, C.; Wu, Y.; Gharibani, P.M.; Yang, I.H.; Phan, T.T.; Lim, K.L.; Yang, H.; et al. Static magnetic field stimulation enhances oligodendrocyte differentiation and secretion of neurotrophic factors. Sci. Rep. 2017, 7, 6743. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Byravan, S.; Foster, L.M.; Phan, T.; Verity, A.N.; Campagnoni, A.T. Murine oligodendroglial cells express nerve growth factor. Proc. Natl. Acad. Sci. USA 1994, 91, 8812–8816. [Google Scholar] [CrossRef] [PubMed]
- Varon, S.; Conner, J.M. Nerve growth factor in CNS repair. J. Neurotrauma 1994, 11, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, S.I.; Harvey, A.R. Neurotrophic factors used to treat spinal cord injury. Vitam. Horm. 2017, 104, 405–457. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.Q.; Dybdal, N.; Shinsky, N.; Murnane, A.; Schmelzer, C.; Siegel, M.; Keller, G.; Hefti, F.; Phillips, H.S.; Winslow, J.W. Neurotrophin-3 reverses experimental cisplatin-induced peripheral sensory neuropathy. Ann. Neurol. 1995, 38, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Rabacchi, S.A.; Kruk, B.; Hamilton, J.; Carney, C.; Hoffman, J.R.; Meyer, S.L.; Springer, J.E.; Baird, D.H. BDNF and NT4/5 promote survival and neurite outgrowth of pontocerebellar mossy fiber neurons. J. Neurobiol. 1999, 40, 254–269. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.F.; Doherty, D.H.; Lile, J.D.; Bektesh, S.; Collins, F. GDNF: A glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A.; Burne, J.F.; Holtmann, B.; Thoenen, H.; Sendtner, M.; Raff, M.C. Ciliary neurotrophic factor enhances the rate of oligodendrocyte generation. Mol. Cell. Neurosci. 1996, 8, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Giess, R.; Holtmann, B.; Braga, M.; Grimm, T.; Müller-Myhsok, B.; Toyka, K.V.; Sendtner, M. Early onset of Severe Familial Amyotrophic Lateral Sclerosis with a SOD-1 mutation: Potential impact of CNTF as a candidate modifier gene. Am. J. Hum. Genet. 2002, 70, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Talbott, J.F.; Cao, Q.; Bertram, J.; Nkansah, M.; Benton, R.L.; Lavik, E.; Whittemore, S.R. CNTF promotes the survival and differentiation of adult spinal cord-derived oligodendrocyte precursor cells in vitro but fails to promote remyelination in vivo. Exp. Neurol. 2007, 204, 485–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, F.J.; Lang, J.K.; Waldau, B.; Roy, N.S.; Schwartz, T.E.; Pilcher, W.H.; Chandross, K.J.; Natesan, S.; Merrill, J.E.; Goldman, S.A. Complementary patterns of gene expression by human oligodendrocyte progenitors and their environment predict determinants of progenitor maintenance and differentiation. Ann. Neurol. 2006, 59, 763–779. [Google Scholar] [CrossRef] [PubMed]
- Amor, V.; Feinberg, K.; Eshed-Eisenbach, Y.; Vainshtein, A.; Frechter, S.; Grumet, M.; Rosenbluth, J.; Peles, E. Long-term maintenance of Na+ channels at nodes of ranvier depends on glial contact mediated by gliomedin and NrCAM. J. Neurosci. 2014, 34, 5089–5098. [Google Scholar] [CrossRef] [PubMed]
- Demyanenko, G.P.; Mohan, V.; Zhang, X.; Brennaman, L.H.; Dharbal, K.E.S.; Tran, T.S.; Manis, P.B.; Maness, P.F. Neural cell adhesion molecule NrCAM regulates Semaphorin 3F-induced dendritic spine remodeling. J. Neurosci. 2014, 34, 11274–11287. [Google Scholar] [CrossRef] [PubMed]
- Rønn, L.C.; Bock, E.; Linnemann, D.; Jahnsen, H. NCAM-antibodies modulate induction of long-term potentiation in rat hippocampal CA1. Brain Res. 1995, 677, 145–151. [Google Scholar] [CrossRef]
- Tessier-Lavigne, M.; Goodman, C.S. The molecular biology of axon guidance. Science 1996, 274, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Gascon, E.; Vutskits, L.; Kiss, J.Z. The role of PSA-NCAM in adult neurogenesis. Adv. Exp. Med. Biol. 2010, 663, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Hattori, D.; Demir, E.; Kim, H.W.; Viragh, E.; Zipursky, S.L.; Dickson, B.J. Dscam diversity is essential for neuronal wiring and self-recognition. Nature 2007, 449, 223–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, D.; Chen, Y.; Matthews, B.J.; Salwinski, L.; Sabatti, C.; Grueber, W.B.; Zipursky, S.L. Robust discrimination between self and non-self neurites requires thousands of Dscam1 isoforms. Nature 2009, 461, 644–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soba, P.; Zhu, S.; Emoto, K.; Younger, S.; Yang, S.J.; Yu, H.H.; Lee, T.; Jan, L.Y.; Jan, Y.N. Drosophila sensory neurons require dscam for dendritic self-avoidance and proper dendritic field organization. Neuron 2007, 54, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.E.; Bortnick, R.; Tsubouchi, A.; Bäumer, P.; Kondo, M.; Uemura, T.; Schmucker, D. Homophilic Dscam interactions control complex dendrite morphogenesis. Neuron 2007, 54, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Bosse, F.; D’Urso, D.; Muller, H.; Sereda, M.W.; Nave, K.; Niehaus, A.; Kempf, T.; Schnolzer, M.; Trotter, J. The AN2 protein is a novel marker for the Schwann cell lineage expressed by immature and nonmyelinating Schwann cells. J. Neurosci. 2001, 21, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Hunanyan, A.S.; Garcia-Alias, G.; Alessi, V.; Levine, J.M.; Fawcett, J.W.; Mendell, L.M.; Arvanian, V.L. Role of chondroitin sulfate proteoglycans in axonal conduction in mammalian spinal cord. J. Neurosci. 2010, 30, 7761–7769. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.H.; Katagiri, Y.; Susarla, B.; Figge, D.; Symes, A.J.; Geller, H.M. Alterations in sulfated chondroitin glycosaminoglycans following controlled cortical impact injury in mice. J. Comp. Neurol. 2012, 520, 3295–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dours-Zimmermann, M.T.; Maurer, K.; Rauch, U.; Stoffel, W.; Fassler, R.; Zimmermann, D.R. Versican V2 assembles the extracellular matrix surrounding the nodes of Ranvier in the CNS. J. Neurosci. 2009, 29, 7731–7742. [Google Scholar] [CrossRef] [PubMed]
- Bekku, Y.; Rauch, U.; Ninomiya, Y.; Oohashi, T. Brevican distinctively assembles extracellular components at the large diameter nodes of Ranvier in the CNS. J. Neurochem. 2009, 108, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dityatev, A.; Schachner, M. Extracellular matrix molecules and synaptic plasticity. Nat. Rev. Neurosci. 2003, 4, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Hagino, S.; Iseki, K.; Mori, T.; Zhang, Y.; Sakai, N.; Yokoya, S.; Hikake, T.; Kikuchi, S.; Wanaka, A. Expression pattern of glypican-1 mRNA after brain injury in mice. Neurosci. Lett. 2003, 349, 29–32. [Google Scholar] [CrossRef]
- Hagino, S.; Iseki, K.; Mori, T.; Zhang, Y.; Hikake, T.; Yokoya, S.; Takeuchi, M.; Hasimoto, H.; Kikuchi, S.; Wanaka, A. Slit and glypican-1 mRNAs are coexpressed in the reactive astrocytes of the injured adult brain. Glia 2003, 42, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, M.; Nakayama, T.; Sato, N.; Naganuma, T.; Yamaguchi, M.; Aoi, N.; Sato, M.; Izumi, Y.; Soma, M.; Matsumoto, K. Association Study of the elastin microfibril interfacer 1 (EMILIN1) gene in essential hypertension. Am. J. Hypertens. 2010, 23, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, M.; Pavlov, I.; Võikar, V.; Lauri, S.E.; Hienola, A.; Riekki, R.; Lakso, M.; Taira, T.; Rauvala, H. Syndecan-3-deficient mice exhibit enhanced LTP and impaired hippocampus-dependent memory. Mol. Cell. Neurosci. 2002, 21, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, Y.; Borrell, V.; Garcia, C.; Burstyn-Cohen, T.; Tzarfaty, V.; Frumkin, A.; Nose, A.; Okamoto, H.; Higashijima, S.; Soriano, E.; et al. F-spondin and mindin: Two structurally and functionally related genes expressed in the hippocampus that promote outgrowth of embryonic hippocampal neurons. Development 1999, 126, 3637–3648. [Google Scholar] [PubMed]
- Wang, B.; Guo, W.; Huang, Y. Thrombospondins and synaptogenesis. Neural Regen. Res. 2012, 7, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Sawyer, A.; Kocaoglu, F.B.; Kyriakides, T.R. Astrocyte-derived thrombospondin-2 is critical for the repair of the blood-brain barrier. Am. J. Pathol. 2011, 179, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Yuzaki, M. Cbln family proteins promote synapse formation by regulating distinct neurexin signaling pathways in various brain regions. Eur. J. Neurosci. 2011, 33, 1447–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuzaki, M. Two classes of secreted synaptic organizers in the central nervous system. Annu. Rev. Physiol. 2018, 80, 243–262. [Google Scholar] [CrossRef] [PubMed]
- Awwad, K.; Hu, J.; Shi, L.; Mangels, N.; Abdel Malik, R.; Zippel, N.; Fisslthaler, B.; Eble, J.A.; Pfeilschifter, J.; Popp, R.; et al. Role of secreted modular calcium-binding protein 1 (SMOC1) in transforming growth factor β signalling and angiogenesis. Cardiovasc. Res. 2015, 106, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettem, K.L. New Synaptic Organizing Proteins and Their Roles in Excitatory and Inhibitory Synapse Development. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2012. [Google Scholar]
- Dean, C.; Dresbach, T. Neuroligins and neurexins: Linking cell adhesion, synapse formation and cognitive function. Trends Neurosci. 2006, 29, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Sakry, D.; Yigit, H.; Dimou, L.; Trotter, J. Oligodendrocyte precursor cells synthesize neuromodulatory factors. PLoS ONE 2015, 10, e0127222. [Google Scholar] [CrossRef] [PubMed]
- Tucsek, Z.; Noa Valcarcel-Ares, M.; Tarantini, S.; Yabluchanskiy, A.; Fülöp, G.; Gautam, T.; Orock, A.; Csiszar, A.; Deak, F.; Ungvari, Z. Hypertension-induced synapse loss and impairment in synaptic plasticity in the mouse hippocampus mimics the aging phenotype: Implications for the pathogenesis of vascular cognitive impairment. GeroScience 2017, 39, 385. [Google Scholar] [CrossRef] [PubMed]
- Pelkey, K.A.; Barksdale, E.; Craig, M.T.; Yuan, X.; Sukumaran, M.; Vargish, G.A.; Mitchell, R.M.; Wyeth, M.S.; Petralia, R.S.; Chittajallu, R.; et al. Pentraxins coordinate excitatory synapse maturation and circuit integration of parvalbumin interneurons. Neuron 2016, 90, 661. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, I.; Lerer-Goldshtein, T.; Okamoto, H.; Appelbaum, L. Reduced synaptic density and deficient locomotor response in neuronal activity-regulated pentraxin 2a mutant zebrafish. FASEB J. 2015, 29, 1220–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Z.; Wang, C.; Zepp, J.; Wu, L.; Sun, K.; Zhao, J.; Chandrasekharan, U.; DiCorleto, P.E.; Trapp, B.D.; Ransohoff, R.M.; et al. Act1 mediates IL-17–induced EAE pathogenesis selectively in NG2+ glial cells. Nat. Neurosci. 2013, 16, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Van Hove, I.; Lemmens, K.; Van de Velde, S.; Verslegers, M.; Moons, L. Matrix metalloproteinase-3 in the central nervous system: A look on the bright side. J. Neurochem. 2012, 123, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Pham, L.D.D.; Hayakawa, K.; Seo, J.H.; Nguyen, M.N.; Som, A.T.; Lee, B.J.; Guo, S.; Kim, K.W.; Lo, E.H.; Arai, K. Crosstalk between oligodendrocytes and cerebral endothelium contributes to vascular remodeling after white matter injury. Glia 2012, 60, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, S.M.; Razak, K.; Ethell, I.M. A delicate balance: Role of MMP-9 in brain development and pathophysiology of neurodevelopmental disorders. Front. Cell. Neurosci. 2015, 9, 280. [Google Scholar] [CrossRef] [PubMed]
- Jourquin, J.; Tremblay, E.; Bernard, A.; Charton, G.; Chaillan, F.A.; Marchetti, E.; Roman, F.S.; Soloway, P.D.; Dive, V.; Yiotakis, A.; et al. Tissue inhibitor of metalloproteinases-1 (TIMP-1) modulates neuronal death, axonal plasticity, and learning and memory. Eur. J. Neurosci. 2005, 22, 2569–2578. [Google Scholar] [CrossRef] [PubMed]
- Leco, K.J.; Apte, S.S.; Taniguchi, G.T.; Hawkes, S.P.; Khokha, R.; Schultz, G.A.; Edwards, D.R. Murine tissue inhibitor of metalloproteinases-4 (Timp-4): CDNA isolation and expression in adult mouse tissues. FEBS Lett. 1997, 401, 213–217. [Google Scholar] [CrossRef]
- Melendez-Zajgla, J.; Del Pozo, L.; Ceballos, G.; Maldonado, V. Tissue inhibitor of metalloproteinases-4. The road less traveled. Mol. Cancer 2008, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Raabe, T.D.; Clive, D.R.; Wen, D.; DeVries, G.H. Neonatal oligodendrocytes contain and secrete neuregulins in vitro. J. Neurochem. 2002, 69, 1859–1863. [Google Scholar] [CrossRef]
- Buonanno, A.; Kwon, O.B.; Yan, L.; Gonzalez, C.; Longart, M.; Hoffman, D.; Vullhorst, D. Neuregulins and neuronal plasticity: Possible relevance in Schizophrenia. Novartis Found. Symp. 2008, 289, 165–177, discussion 177–179, 193–195. [Google Scholar] [PubMed]
- Canoll, P.D.; Musacchio, J.M.; Hardy, R.; Reynolds, R.; Marchionni, M.A.; Salzer, J.L. GGF/neuregulin is a neuronal signal that promotes the proliferation and survival and inhibits the differentiation of oligodendrocyte progenitors. Neuron 1996, 17, 229–243. [Google Scholar] [CrossRef]
- Bermingham-McDonogh, O.; McCabe, K.L.; Reh, T.A. Effects of GGF/neuregulins on neuronal survival and neurite outgrowth correlate with erbB2/neu expression in developing rat retina. Development 1996, 122, 1427–1438. [Google Scholar] [PubMed]
- Shen, K.Z.; Zhu, Z.T.; Munhall, A.; Johnson, S.W. Dopamine receptor supersensitivity in rat subthalamus after 6-hydroxydopamine lesions. Eur. J. Neurosci. 2003, 18, 2967–2974. [Google Scholar] [CrossRef] [PubMed]
- Hökfelt, T.; Broberger, C.; Diez, M.; Xu, Z.Q.; Shi, T.; Kopp, J.; Zhang, X.; Holmberg, K.; Landry, M.; Koistinaho, J. Galanin and NPY, two peptides with multiple putative roles in the nervous system. Horm. Metab. Res. 1999, 31, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Biswas, N.; Gayen, J.R.; Miramontes-Gonzalez, J.P.; Hightower, C.M.; Mustapic, M.; Mahata, M.; Huang, C.T.; Hook, V.Y.; Mahata, S.K.; et al. Chromogranin B: Intra- and extra-cellular mechanisms to regulate catecholamine storage and release, in catecholaminergic cells and organisms. J. Neurochem. 2014, 129, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.G.; Kim, J.H.; Park, C.S.; Kim, B.J.; Kim, J.W.; Choi, I.G.; Hwang, J.; Shin, H.D.; Woo, S.I. Gender-specific associations between CHGB genetic variants and Schizophrenia in a Korean population. Yonsei Med. J. 2017, 58, 619. [Google Scholar] [CrossRef] [PubMed]
- Gasser, M.C.; Berti, I.; Hauser, K.F.; Fischer-Colbrie, R.; Saria, A. Secretoneurin promotes pertussis toxin-sensitive neurite outgrowth in cerebellar granule cells. J. Neurochem. 2003, 85, 662–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchmair, R.; Gander, R.; Egger, M.; Hanley, A.; Silver, M.; Ritsch, A.; Murayama, T.; Kaneider, N.; Sturm, W.; Kearny, M.; et al. The neuropeptide secretoneurin acts as a direct angiogenic cytokine in vitro and in vivo. Circulation 2004, 109, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Milbrandt, J.; de Sauvage, F.J.; Fahrner, T.J.; Baloh, R.H.; Leitner, M.L.; Tansey, M.G.; Lampe, P.A.; Heuckeroth, R.O.; Kotzbauer, P.T.; Simburger, K.S.; et al. Persephin, a novel neurotrophic factor related to GDNF and neurturin. Neuron 1998, 20, 245–253. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: A new resource for understanding brain development and function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Farías, G.G.; Godoy, J.A.; Cerpa, W.; Varela-Nallar, L.; Inestrosa, N.C. Wnt signaling modulates pre- and postsynaptic maturation: Therapeutic considerations. Dev. Dyn. 2009, 239, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.C.; Lucas, F.R.; Salinas, P.C. Axonal remodeling and synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell 2000, 100, 525–535. [Google Scholar] [CrossRef]
- Oliva, C.A.; Vargas, J.Y.; Inestrosa, N.C. Wnts in adult brain: From synaptic plasticity to cognitive deficiencies. Front. Cell. Neurosci. 2013, 7, 224. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Annuar, A.; Ciani, L.; Simeonidis, I.; Herreros, J.; Fredj, N.B.; Rosso, S.B.; Hall, A.; Brickley, S.; Salinas, P.C. Signaling across the synapse: A role for Wnt and Dishevelled in presynaptic assembly and neurotransmitter release. J. Cell Biol. 2006, 174, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Katayama, M.; Iwasaki, S.; Ishii, K.; Tsujimoto, M.; Kohno, M. Bone morphogenetic protein-2 promotes survival and differentiation of striatal GABAergic neurons in the absence of glial cell proliferation. J. Neurochem. 2002, 72, 2264–2271. [Google Scholar] [CrossRef]
- Kusakawa, Y.; Mikawa, S.; Sato, K. BMP7 expression in the adult rat brain. IBRO Rep. 2017, 3, 72–86. [Google Scholar] [CrossRef]
- Nguyen-Ba-Charvet, K.T.; Picard-Riera, N.; Tessier-Lavigne, M.; Baron-Van Evercooren, A.; Sotelo, C.; Chédotal, A. Multiple roles for slits in the control of cell migration in the rostral migratory stream. J. Neurosci. 2004, 24, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Wehrle, R.; Camand, E.; Chedotal, A.; Sotelo, C.; Dusart, I. Expression of netrin-1, slit-1 and slit-3 but not of slit-2 after cerebellar and spinal cord lesions. Eur. J. Neurosci. 2005, 22, 2134–2144. [Google Scholar] [CrossRef] [PubMed]
- Hewett, S.J.; Jackman, N.A.; Claycomb, R.J. Interleukin-1β in central nervous system injury and repair. Eur. J. Neurodegener. Dis. 2012, 1, 195–211. [Google Scholar] [PubMed]
- Bauer, S.; Kerr, B.J.; Patterson, P.H. The neuropoietic cytokine family in development, plasticity, disease and injury. Nat. Rev. Neurosci. 2007, 8, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Deverman, B.E.; Patterson, P.H. Cytokines and CNS development. Neuron 2009, 64, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Strle, K.; Zhou, J.H.; Shen, W.H.; Broussard, S.R.; Johnson, R.W.; Freund, G.G.; Dantzer, R.; Kelley, K.W. Interleukin-10 in the brain. Crit. Rev. Immunol. 2001, 21, 427–449. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Stillings, S.A.; Leclerc, J.L.; Phillips, H.; Edwards, N.J.; Robicsek, S.A.; Hoh, B.L.; Blackburn, S.; Doré, S.L. Role of interleukin-10 in acute brain injuries. Front. Neurol. 2017, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Dobolyi, A.; Vincze, C.; Pál, G.; Lovas, G. The neuroprotective functions of transforming growth factor βproteins. Int. J. Mol. Sci. 2012, 13, 8219–8258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.E.; Lipsky, B.P.; Russell, C.; Ketchem, R.R.; Kirchner, J.; Hensley, K.; Huang, Y.; Friedman, W.J.; Boissonneault, V.; Plante, M.M.; et al. A central nervous system-restricted isoform of the interleukin-1 receptor accessory protein modulates neuronal responses to interleukin-1. Immunity 2009, 30, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, C.J.; Martin, B.N.; Bulek, K.; Kang, Z.; Zhao, J.; Bian, G.; Carman, J.A.; Gao, J.; Dongre, A.; et al. IL-17 induced NOTCH1 activation in oligodendrocyte progenitor cells enhances proliferation and inflammatory gene expression. Nat. Commun. 2017, 8, 15508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marro, B.S.; Grist, J.J.; Lane, T.E. Inducible expression of CXCL1 within the central nervous system amplifies viral-induced demyelination. J. Immunol. 2016, 196, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.S. Regulation of neuroinflammation: The role of CXCL10 in lymphocyte infiltration during autoimmune encephalomyelitis. J. Cell. Biochem. 2004, 92, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Bisht, K.; Tremblay, M.È. Fractalkine regulation of microglial physiology and consequences on the brain and behavior. Front. Cell. Neurosci. 2014, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Hesselgesser, J.; Horuk, R. Chemokine and chemokine receptor expression in the central nervous system. J. Neurovirol. 1999, 5, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Tani, M. Do chemokines mediate leukocyte recruitment in post-traumatic CNS inflammation? Trends Neurosci. 1998, 21, 154–159. [Google Scholar] [CrossRef]
- Mennicken, F.; Maki, R.; de Souza, E.B.; Quirion, R. Chemokines and chemokine receptors in the CNS: A possible role in neuroinflammation and patterning. Trends Pharmacol. Sci. 1999, 20, 73–78. [Google Scholar] [CrossRef]
- Schäbitz, W.R.; Krüger, C.; Pitzer, C.; Weber, D.; Laage, R.; Gassler, N.; Aronowski, J.; Mier, W.; Kirsch, F.; Dittgen, T.; et al. A neuroprotective function for the hematopoietic protein granulocyte-macrophage colony stimulating factor (GM-CSF). J. Cereb. Blood Flow Metab. 2008, 28, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, R.; Heine, M.; Perrais, D.; Seidenbecher, C.I.; Choquet, D.; Gundelfinger, E.D. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat. Neurosci. 2009, 12, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Michaluk, P.; Mikasova, L.; Groc, L.; Frischknecht, R.; Choquet, D.; Kaczmarek, L. Matrix metalloproteinase-9 controls NMDA receptor surface diffusion through integrin β1 signaling. J. Neurosci. 2009, 29, 6007–6012. [Google Scholar] [CrossRef] [PubMed]
- Petrosyan, H.A.; Hunanyan, A.S.; Alessi, V.; Schnell, L.; Levine, J.; Arvanian, V.L. Neutralization of inhibitory molecule NG2 improves synaptic transmission, retrograde transport, and locomotor function after spinal cord injury in adult rats. J. Neurosci. 2013, 33, 4032–4043. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dietz, K.; Hodes, G.E.; Russo, S.J.; Casaccia, P. Widespread transcriptional alternations in oligodendrocytes in the adult mouse brain following chronic stress. Dev. Neurobiol. 2018, 78, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.P.; Coulter, M.; Miotke, J.; Meyer, R.L.; Takemaru, K.I.; Levine, J.M. Abrogation of catenin signaling in oligodendrocyte precursor cells reduces glial scarring and promotes axon regeneration after CNS Injury. J. Neurosci. 2014, 34, 10285–10297. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Gallo, V. The diversity and disparity of the glial scar. Nat. Neurosci. 2018, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Li, Y.; Fukaya, M.; Lorenzini, I.; Cleveland, D.W.; Ostrow, L.W.; Rothstein, J.D.; Bergles, D.E. Degeneration and impaired regeneration of gray matter oligodendrocytes in amyotrophic lateral sclerosis. Nat. Neurosci. 2013, 16, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampton, D.; Rhodes, K.; Zhao, C.; Franklin, R.J.; Fawcett, J. The responses of oligodendrocyte precursor cells, astrocytes and microglia to a cortical stab injury, in the brain. Neuroscience 2004, 127, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Lytle, J.M.; Chittajallu, R.; Wrathall, J.R.; Gallo, V. NG2 cell response in the CNP-EGFP mouse after contusive spinal cord injury. Glia 2009, 57, 270–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, S.; Zeisel, A.; Codeluppi, S.; van Bruggen, D.; Mendanha Falcão, A.; Xiao, L.; Häring, M.; Hochgerner, H.; Romanov, R.A.; Gyllborg, D.; et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 2016, 352, 1326–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakers, K.; Lake, A.M.; Khazanchi, R.; Ouwenga, R.; Vasek, M.J.; Dani, A.; Dougherty, J.D. Astrocytes locally translate transcripts in their peripheral processes. Proc. Natl. Acad. Sci. USA 2017, 114, E3830–E3838. [Google Scholar] [CrossRef] [PubMed]
- Crosetto, N.; Bienko, M.; van Oudenaarden, A. Spatially resolved transcriptomics and beyond. Nat. Rev. Genet. 2015, 16, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Dimou, L.; Simons, M. Diversity of oligodendrocytes and their progenitors. Curr. Opin. Neurobiol. 2017, 47, 73–79. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the mechanisms assuring NG2 glia maintenance and of their physical interactions with neurons and other central nervous system (CNS) cell types (A) NG2 glia are distributed in a grid-like manner and occupy non-overlapping domains. When an individual NG2-expressing cell is lost, a neighboring NG2 cell divide and migrate to fill the unoccupied space. (B) In the adult and juvenile cortex, NG2 glia can divide asymmetrically and give rise to a mixed progeny either keeping a progenitor phenotype or proceeding to differentiation. (C) NG2 glia establish physical contacts with neuronal dendrites, somata, nodes of Ranvier and synapses. NG2 glia and astrocytes often contact the same node of Ranvier or synapse, but the relative processes show distinct localization. NG2 glia also receive functional synaptic contacts from unmyelinated axons and axonal terminals. In the adult CNS, NG2 glia display a perivascular distribution or extend some processes to physically interact with pericytes and blood vessels. NT: Neurotransmitter; OL: Oligodendrocyte.
Figure 1.
Schematic representation of the mechanisms assuring NG2 glia maintenance and of their physical interactions with neurons and other central nervous system (CNS) cell types (A) NG2 glia are distributed in a grid-like manner and occupy non-overlapping domains. When an individual NG2-expressing cell is lost, a neighboring NG2 cell divide and migrate to fill the unoccupied space. (B) In the adult and juvenile cortex, NG2 glia can divide asymmetrically and give rise to a mixed progeny either keeping a progenitor phenotype or proceeding to differentiation. (C) NG2 glia establish physical contacts with neuronal dendrites, somata, nodes of Ranvier and synapses. NG2 glia and astrocytes often contact the same node of Ranvier or synapse, but the relative processes show distinct localization. NG2 glia also receive functional synaptic contacts from unmyelinated axons and axonal terminals. In the adult CNS, NG2 glia display a perivascular distribution or extend some processes to physically interact with pericytes and blood vessels. NT: Neurotransmitter; OL: Oligodendrocyte.


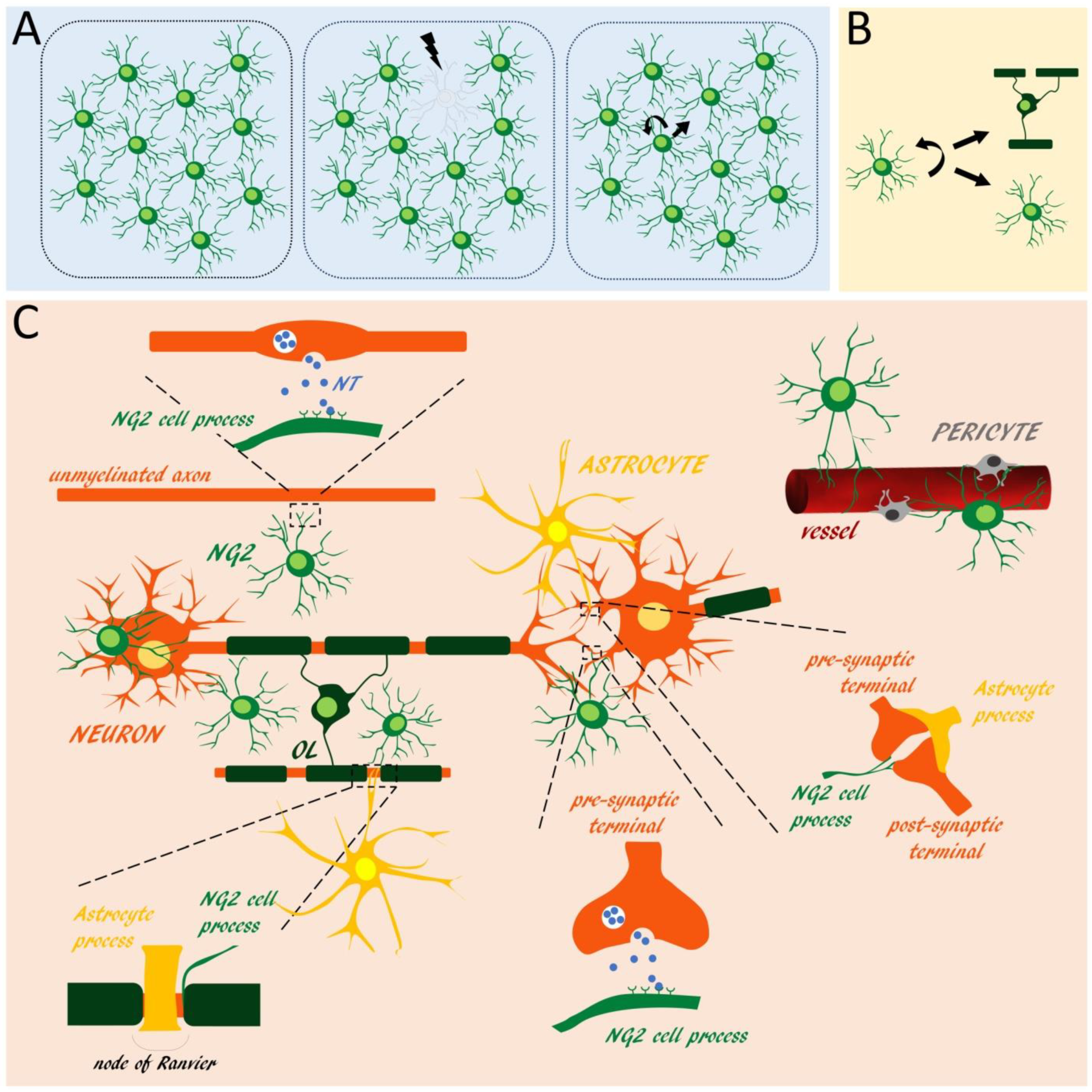{kind=link}
Table 1.
List of the signals produced/released by NG2 glia.
| Type of Signal | Name and Abbreviation | Exp. Approach (Source) | Function |
|---|---|---|---|
| Growth factors | Fibroblast growth factor 2, FGF2 | Gene expr. study of cultured mouse NG2 glia and expr. in adult mouse NG2 glia in vivo [87] | NG2 glia modulation of glutamatergic neurotransmission and astrocytic extracellular glutamate uptake [87], neuronal protection and CNS repair [101], modulation of synaptic plasticity [102] and astroglial and microglial reactivity [103] |
| Platelet-derived growth factor AA, PDGF-AA | CM of cultured mouse NG2 glia [97] | Neuronal protection [104], synaptic plasticity [105] and angiogenesis [106] | |
| Platelet-derived growth factor BB, PDGF-BB | CM of cultured mouse NG2 glia [97] | Neurotrophic and neuroregulatory functions [107], synaptic plasticity [105] | |
| Vascular endothelial growth factor, VEGF | CM of cultured mouse NG2 glia [97] | Neurotrophic function [108,109], regulation of adult neurogenesis [108] | |
| Insulin-like growth factor 1, IGF-1 | CM of cultured rat NG2 glia [94]; Gene expr. study of cultured rat NG2 glia [93] | Synaptic maturation [110] | |
| Hepatocyte growth factor, HGF | In vivo expr. in rat NG2 glia [88]; Protein expr. in cultured rat NG2 glia [111] | Neuronal plasticity [112] | |
| Neurotrophins | Brain derived growth factor, BDNF | CM of cultured rat NG2 glia and protein expr. in adult mouse NG2 glia in vivo [113]; CM of cultured human NG2 glia [114]; Gene expr. study and CM of cultured rat NG2 glia [93] | Neurotrophic function, neurotransmitter modulation and in neuronal plasticity [115] |
| Nerve growth factor, NGF | Gene expr. study of cultured mouse NG2 glia [87,116] | Neurotrophic function, brain plasticity [117] | |
| Neurotrophin-3, NT-3 | CM of cultured human NG2 glia [114]; Gene expr. study of cultured rat NG2 glia [93] | Neurotrophic function, neuronal survival and differentiation [118,119] | |
| Neurotrophin 4/5, NT-4/5 | Gene expr. study of cultured mouse NG2 glia [87] | Neurotrophic function, neuronal survival and differentiation [120] | |
| Glial cell-derived neurotrophic factor, GDNF | Gene expr. study of cultured mouse NG2 glia [87]; Gene expr. study of cultured rat NG2 glia [93] | Neurotrophic function, survival and morphological differentiation of dopaminergic neurons [121] | |
| Ciliary neurotrophic factor, CNTF | Gene expr. study of cultured rat NG2 glia [93] | Neuronal and oligodendroglial survival [122,123,124] | |
| Cell adhesion and extracellular matrix molecules | Neuronal cell adhesion molecule, NrCAM | Gene expr. study of freshly sorted adult human NG2 glia [125] | Maintenance of Na+ channels at nodes of Ranvier [126], dendritic spine remodelling [127] |
| NCAM | Gene expr. study of freshly sorted adult human NG2 glia [125] | Neuronal plasticity [128], neuritogenesis and synaptogenesis [129], adult neurogenesis [130] | |
| Down Syndrome Cell Adhesion Molecule, Dscam | Gene expr. study of freshly sorted mouse NG2 glia [131] and freshly sorted adult human NG2 glia [125] | Neurite repulsion [132,133,134,135] | |
| Chondroitin sulphate proteoglycan4, Cspg4/NG2 | Gene expr. study of freshly sorted adult human NG2 glia [125,136] | Modulation of synaptic activity and action potential conduction [137], axonal guidance and regenerative processes [138] | |
| Versican, Vcan | Gene expr. study of freshly sorted adult human NG2 glia [125] and freshly sorted mouse NG2 glia [131] | Assembly, maintenance and function of the nodes of Ranvier [139] | |
| Brevican | Gene expr. study of freshly sorted mouse NG2 glia [131] | Assembly, maintenance and function of the nodes of Ranvier [140] | |
| Tenascins | Gene expr. study of freshly sorted adult human NG2 glia [125]; Gene expr. study of freshly sorted mouse NG2 glia [131] | Synaptic plasticity [141] | |
| Glypican 5, Gpc5 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Neuronal plasticity [142,143] | |
| Emilin And Multimerin Domain-Containing Protein 1, Emid1 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Blood vessel maintenance [144] | |
| Syndecan 3, sdc3 | Gene expr. study of freshly sorted adult human NG2 glia [125] | Synaptic plasticity [145] | |
| Spondin 1, Spon1 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Neural cell adhesion and neurite outgrowth [146] | |
| Thrombospondin 2 and 4 | Gene expr. study of freshly sorted adult human NG2 glia [125] | Synaptogenesis and synaptic plasticity [147], inflammatory response and repair of the blood brain barrier [148] | |
| Cerebellin 1, cbln 1 | Gene expr. study of cultured mouse NG glia [87] | Synaptic organizer, synapse integrity and synaptic plasticity [149,150] | |
| SPARC-Related Modular Calcium-Binding Protein 1, SMOC1 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Angiogenesis [151] | |
| Olfactomedin 2, Olfm2 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Synaptogenesis and synaptic plasticity [152] | |
| Neurexophilin 1, Nxph1 | Gene expr. study of freshly sorted mouse NG2 glia [131] | Synaptogenesis and synaptic plasticity [150,153] | |
| Neuronal Pentraxin 2, Nptx2 | Gene expr. study of freshly sorted mouse NG2 glia [131] and of cultured and freshly sorted mouse NG2 glia [154] | Synaptogenesis and synaptic plasticity [150,153,155,156], trafficking of glutamate receptors [157] | |
| Matrix metalloproteases and metalloprotease inhibitors | Matrix Metalloprotease 3, MMP3 | CM of cultured mouse NG2 glia [26]; Gene expr. study of cultured mouse NG2 glia exposed to Il17 or Tnf [158] | Neuronal and synaptic plasticity [159] |
| Matrix Metalloprotease 9, MMP9 | CM of cultured mouse NG2 glia [26]; CM of cultured rat NG2 glia and protein expr. in mouse NG2 glia in vivo—adult white matter [67] Gene expr. study of cultured mouse NG2 glia exposed to Il17 or Tnf [158] | Blood vessel remodeling after white matter injury [160]. Remodeling of synaptic networks in adult brain [161] | |
| Tissue Inhibitor of Metalloprotease 1, TIMP1 | CM of cultured mouse NG2 glia [26] | Neuronal death and axonal plasticity [162] | |
| Tissue Inhibitor of Metalloprotease 4, TIMP4 | Gene expr. study of freshly sorted mouse NG2 glia [131]; Gene expr. study of freshly sorted adult human NG2 glia [125] | Tissue remodelling [163,164] | |
| Neuromodulatory/neurosupportive factors | Neuregulins | Gene expr. study of cultured mouse NG glia [87]; Gene expr. study and CM of cultured rat NG glia [165] | Synaptogenesis and synaptic plasticity [166], oligodendroglial and neuronal survival [167,168] |
| Galanin | mRNA and protein expr. in adult mouse NG2 glia [169] | Modulation of neuronal activity [170] | |
| Chromogranin B, Chgb | Gene expr. study of freshly sorted adult human NG2 glia [125] | Neurotransmission [171,172] | |
| Chromogranin C, Chgc | Gene expr. study of freshly sorted adult human NG2 glia [125] | Neurite outgrowth [173] and angiogenesis [174]. | |
| Persephin, PSPN | Gene expr. study of cultured mouse NG glia [87] | Neuronal survival [175] | |
| Morphogens | Wingless-type MMTV integration site family 4, Wnt4 | Gene expr. study of freshly sorted mouse NG2 glia [176] | Synaptic plasticity [177] |
| Wingless-type MMTV integration site family 7a, Wnt7a | Gene expr. study of freshly sorted mouse NG2 glia [176]; Gene expr. study and CM of cultured mouse NG2 glia [96]; In vivo expr. in embryonic NG2 glia [64] | Synaptic plasticity [178,179]; NG2 glia regulation of angiogenesis [96] | |
| Wingless-type MMTV integration site family 7b, Wnt7b | Gene expr. study of freshly sorted mouse NG2 glia [176]; Gene expr. study and CM of cultured mouse NG2 glia [96]; In vivo expr. in embryonic NG2 glia [64] | Synaptic plasticity [180] | |
| Bone morphogenic protein 2, BMP2 | Gene expr. study of freshly sorted adult human NG2 glia [125] | Neuronal survival [181] | |
| Bone morphogenic protein 7, BMP7 | Gene expr. study of freshly sorted adult human NG2 glia [125] | Maintainance of the identity of catecholaminergic neurons and differentiation of astrocytes [182] | |
| Retinoic acid, RA | Application of pharmacological inhibitors in cultured mouse NG2 glia and in vivo assays in a model of rat spinal cord injury [99] | NG2 glia regulation of axonal outgrowth [99] | |
| Slit1 | Gene expr. study of freshly sorted adult human NG2 glia [125] | Regulation of adult SVZ neurogenesis [183], axon outgrowth and glial scar formation [184] | |
| Inflammatory cytokines/immunomodulatory factors | Interleukin 1 beta, Il-1b | Gene expr. study of cultured mouse NG2 glia [87]; Gene expr. study of cultured rat NG2 glia [93] | Synaptic plasticity [185] |
| Interleukin 6, Il-6 | Gene expr. study of cultured mouse NG2 glia [87]; Gene expr. study of cultured rat NG2 glia [93] Gene expr. study of cultured mouse NG2 glia exposed to Il17 or Tnf [158] | Regulation of adult neurogenesis [186,187], neuronal protection [187] | |
| Interleukin 10, Il-10 | Gene expr. study of cultured mouse NG2 glia [87]; Gene expr. study and CM of cultured rat NG2 glia [93] | Neuronal and glial cell survival [88,188]. Anti-inflammatory functions [189] | |
| Transforming growth factor beta, Tgf-b | Gene expr. study of cultured mouse NG2 glia [87]; Gene expr. study of cultured rat NG2 glia and protein expr. in mouse neonatal NG2 glia in vivo [65] | Neuronal survival and modulation of synaptic transmission [190] | |
| Interleukin 1 Receptor Accessory Protein, Il1rap | Gene expr. study of freshly sorted mouse NG2 glia [131] | Neuronal survival [191] | |
| C-X-C Motif Chemokine Ligand 10, CxCl1 | Gene expr. study of cultured mouse NG2 glia exposed to Il17 or Tnf [158,192] | Neuroinflammation [193] | |
| C-X-C Motif Chemokine Ligand 10, CxCl10 | CM of cultured mouse NG2 glia [97] | Neuroinflammation [194] | |
| C-X3-C Motif Chemokine Ligand 1, Cx3Cl1 | CM of cultured mouse NG2 glia [97] | Regulation of synapses activity and plasticity, brain functional connectivity and adult hippocampal neurogenesis [195] | |
| C-C Motif Chemokine Ligand 7, Ccl7 | Gene expr. study of cultured mouse NG2 glia exposed to Il17 [192] | Neuroinflammation [196,197,198] | |
| Granulocyte-Macrophage Colony Stimulating Factor, GM-CSF or Csf2 | Gene expr. study of cultured mouse NG2 glia exposed to Il17 or Tnf [158] | Neurotrophic function [199] |
Exp: Experimental; expr: Expression; CM: Conditioned medium; FGF2: Fibroblast growth factor 2; NG2: Neuron-glia antigen 2.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Parolisi, R.; Boda, E. NG2 Glia: Novel Roles beyond Re-/Myelination. Neuroglia 2018, 1, 151-175. https://0-doi-org.brum.beds.ac.uk/10.3390/neuroglia1010011
AMA Style
Parolisi R, Boda E. NG2 Glia: Novel Roles beyond Re-/Myelination. Neuroglia. 2018; 1(1):151-175. https://0-doi-org.brum.beds.ac.uk/10.3390/neuroglia1010011
Chicago/Turabian StyleParolisi, Roberta, and Enrica Boda. 2018. "NG2 Glia: Novel Roles beyond Re-/Myelination" Neuroglia 1, no. 1: 151-175. https://0-doi-org.brum.beds.ac.uk/10.3390/neuroglia1010011