The Vitamin D System in Humans and Mice: Similar but Not the Same


Department of Biotechnology, University of Wroclaw, Joliot-Curie 14a, 50-383 Wroclaw, Poland
Reports 2020, 3(1), 1; https://0-doi-org.brum.beds.ac.uk/10.3390/reports3010001
Submission received: 7 November 2019
/
Revised: 2 January 2020
/
Accepted: 7 January 2020
/
Published: 10 January 2020
Abstract
:Vitamin D is synthesized in the skin from 7-dehydrocholesterol subsequently to exposure to UVB radiation or is absorbed from the diet. Vitamin D undergoes enzymatic conversion to its active form, 1,25-dihydroxyvitamin D (1,25D), a ligand to the nuclear vitamin D receptor (VDR), which activates target gene expression. The best-known role of 1,25D is to maintain healthy bones by increasing the intestinal absorption and renal reuptake of calcium. Besides bone maintenance, 1,25D has many other functions, such as the inhibition of cell proliferation, induction of cell differentiation, augmentation of innate immune functions, and reduction of inflammation. Significant amounts of data regarding the role of vitamin D, its metabolism and VDR have been provided by research performed using mice. Despite the fact that humans and mice share many similarities in their genomes, anatomy and physiology, there are also differences between these species. In particular, there are differences in composition and regulation of the VDR gene and its expression, which is discussed in this article.
{kind=link}
{kind=link}
1. Introduction
Different animal models have been used in biomedical research. Simple organisms, such as yeast, fruit flies, C. elegans and zebrafish are useful to study function of particular genes and their roles in development. However, when it comes to drug research, nothing compares to mice (Mus musculus). Mice are the most widely used animals for finding drugs for human diseases. This is because both mice and humans are mammals, and there is a high homology between their genomes, anatomy and physiology. In addition, mice are cheap and easy to breed, they reproduce quickly and genetic manipulations are relatively simple to conduct. However, there are also obvious and striking differences between humans and mice. Humans are approximately 2500 times larger than mice, the murine metabolic rate per gram of tissue in mice is seven times higher than that of an average man, mice preferentially consume grains and cereals, while contemporary people have plenty of meat in their diet. Mice lead a nocturnal life, while natural human behavior is diurnal, and lastly, mice are covered in fur, in contrast to humans [1].
2. Murine Model in Vitamin D Research
2.1. Short Overview of Vitamin D Physiology
Vitamin D is a derivative of cholesterol, as is similar with other steroid hormones. However, in contrast to other steroid hormones, it requires sun to be produced in the human body. In 7-dehydrocholesterol deposited in the skin, exposure to UVB radiation causes a break between C-9 and C-10 of the B ring, resulting in the formation of pre-vitamin D. Then, thermal isomerisation occurs which results in the formation of vitamin D, a biologically inert compound [2]. During the industrialization period, when people started to live in the overcrowded cities, decreased access to the sun resulted in a massive outbreak of rickets, due to vitamin D deficiency. It has been proposed by Jedrzej Śniadecki in 1822 that the exposure of children to the sun has an antirachitic effect [3]. Nowadays, rickets is rare, but since most of the people live in the areas where the access to sun is limited for approximately half a year, and in sunny days the use of sunscreens is advised, most people are not vitamin D sufficient. Therefore, the alternative sources of vitamin D are used, such as a vitamin-D-rich diet or vitamin D supplementation. The activation of vitamin D is performed in the body by cytochrome P450 oxygenases (CYPs) and occurs in two steps, and in two organs: 25-hydroxylation by CYP2R1 or CYP27A1 in the liver, followed by 1α-hydroxylation by CY27B1 in the kidneys (Figure 1) [4].
Double hydroxylated vitamin D, 1,25-dihydroxyvitamin D (1,25D), is a highly active compound, whose concentration in the human body must be strictly controlled. The mechanism of this control is performed by CYP24A1, encoding 24-hydroxylase of 1,25D, which catabolizes 1,25D to water soluble and biologically inactive calcitroic acid [4]. 1,25D, similarly to other steroid hormones, exerts its biological activity by binding to the nuclear vitamin D receptor (VDR), which forms a dimer with retinoid X receptor (RXR) after ligation. The 1,25D/VDR/RXR complex is then translocated to the cell nucleus where it acts as a ligand activated transcription factor [5]. There are hundreds of VDR-regulated genes, but the most strongly regulated one is CYP24A1, this way providing a negative feedback loop, which controls effective concentration of 1,25D [6]. The best known role of 1,25D is the control over calcium-phosphate homeostasis in the body [7]. However, VDR is present also in cells and tissues not implicated in calcium and phosphate regulation therefore the roles other than calcemic have been proposed for 1,25D. It has been accepted that 1,25D is important for regulation of immune system, however the clear and precise picture of this regulation is still not available [8]. The presence of VDR in developing blood cells has also been documented, and it has been shown that in human hematopoietic stem cells (HSCs) physiological concentrations of 1,25D induce monocytic differentiation [9].
2.2. Vitamin D Analogs
VDR was found not only in normal cells, but also in many of their cancerous counterparts [10]. In some of these cancers, 1,25D is capable to induce differentiation [11,12]. Thus, a number of studies have investigated the therapeutic potential of 1,25D. However, the use of 1,25D as a drug is limited by possible development of hypercalcemia [13]. This is why numerous 1,25D analogs have been produced, which are less calcemic than 1,25D, but still capable to induce cell differentiation [14,15]. Some of 1,25D analogs have been registered as drugs for psoriasis [16] or hyperparathyroidism [17], other are still in preclinical trials. The hundreds of available analogs have been tested first for their affinity to VDR, then in in vitro cell cultures, and eventually for toxicity and calcemic effects in vivo in murine models. Some of these analogs have been rejected from further studies because of general toxicity for mice [18].
2.3. Vitamin D System in Humans and Mice
The source of vitamin D in mice is the same as in humans. Despite being covered by fur, rodents produce vitamin D from 7-dehydrocholesterol after exposure to sun [19], but due to their nocturnal life, mice have less opportunities than humans to expose themselves to sun. In addition, both species can absorb vitamin D from the diet and humans also use supplements [20]. Interestingly, vitamin D is a very common component of rodenticides, which not only kill mice and rats, but also cause accidental toxicosis in dogs and cats [21]. As reported, 600 IU of vitamin D (cholecalciferol) is enough to kill a mouse [22]. It is hard to say whether the equivalent doses of vitamin D are lethal for people, since such doses have not been attempted in humans (1,500,000 IU).
The importance of 1,25D for living organisms can be learned not only from deficient individuals, but also from those who have mutations in genes encoding proteins that are important for vitamin D actions. There are some naturally occurring mutations that affect the vitamin D system in humans. Hereditary vitamin-D-resistant rickets (HVDRR) is caused by mutations in the VDR gene [23]. The predominant symptoms are severe rickets, growth retardation and hypoplasia of the tooth enamel. However, in some patients, depending on the localization of mutation in VDR gene, hair loss (alopecia) is an accompanying symptom. Mutations in the region coding ligand-binding domain (LBD) do not affect hair growth, while missense mutations in the DNA binding domain (DBD) or mutations that interfere with RXR dimerization cause alopecia in patients [23].
Vitamin-D-dependent rickets type 1 (VDDR-I) is caused by mutations in the CYP27B1 gene. This disease is an autosomal recessive disorder characterized by the early rickets with hypocalcemia due to inability to convert 25-hydroxyvitamin D to 1,25D [24]. In this other disease, hair growth has not been reported to be affected. The relevance of the above proteins for mice can be learnt from VDR-null and CYP27B1-null mice [25]. VDR-null mice develop hypocalcemia in infancy, followed by rickets, osteomalacia, growth retardation, secondary hyperparathyroidism and alopecia [26], while CYP27B1-null mice develop hypocalcemia, secondary hyperparathyroidism, retarded growth, and the skeletal abnormalities characteristic of rickets, but no alopecia. In addition, CYP27B1-null mice have enlarged lymph nodes in the vicinity of the thyroid gland and a reduced number of peripheral T lymphocytes. Female CYP27B1-null mice are infertile, exhibit uterine hypoplasia and absent corpora lutea [27]. The data described above, observed in patients and in mice, indicate that VDR has roles that are either ligand-dependent or ligand-independent. Regulation of hair growth does not depend on 1,25D, and most probably is caused by inhibition of transcription of certain genes by unliganded VDR [23]. Regulation of calcium-phosphate homeostasis, female reproduction and of some immune functions are 1,25D-dependent.
2.4. Vitamin D Receptor in Blood Development
It has been reported that VDR regulates transcription of around 3% of genes of human and murine genomes [25]. Among these genes, many are characteristic for blood cells’ development and function [28]. Some of them encode transcription factors involved in hematopoiesis [29,30], or proteins important for function of blood cells [31]. It has been also reported that VDR is present in multiple blood cells, including hematopoietic stem cells (HSC) in humans and in mice [9,32]. In VDR knockout mice, haematopoiesis and blood cell counts are normal, as long as the mice are kept in a pathogen-free environment. Exposure of these mice to pathogens led to the blood disorder characterized by an appearance of chronic myeloid leukemia-like cells, splenomegaly, granulocytosis, thrombocytosis and reduced erythropoiesis [33]. The above data suggest that 1,25D, VDR, or both, might take part in hematopoiesis.
2.5. Vitamin D Receptor Gene in Humans and in Mice
Vitamin D is produced after exposure to sun in wide variety of organisms. Phytoplankton, zooplankton, most plants and animals, invertebrate and vertebrate, have the capacity to make vitamin D [34]. The role of 1,25D and of its receptor is well understood in case of terrestrial vertebrates who need to maintain healthy skeleton. It is less clear why vitamin D and VDR are ubiquitous also in lower organisms. One hypothesis says that the primary role of VDR was not in calcium-phosphate homeostasis, but in detoxification [35]. The VDR-LBD binds not only 1,25D, but also litocholic acid (LCA) [36,37], which is a toxic and carcinogenic product of bile acids metabolism. There have been three rounds of whole genome duplications throughout the evolution of eukaryotes and common evolutionary path of humans and mice emerged after the second round. It is believed that before the second duplication, the functions of VDR and pregnane X receptor (PXR) have been combined in one protein. The function of PXR in humans is to detoxify xenobiotics by activating the enzyme CYP3A4 [38]. The second round of duplication gave rise to separate VDR and PXR genes and proteins with separate functions. It is noteworthy that in teleost fish, which were subjected to the third round of whole-genome duplication, there is one more copy of the VDR gene [39]. Separation of VDR and PXR allowed for the specification of VDR’s function in bone maintenance.
Despite the common evolution of murine and human VDR genes, their compositions are diverse. The VDR gene in humans is located on chromosome 12 and spans about 100 kb of genomic DNA [40]. Its 5’ regulatory region is very complex, and consists of seven untranslated exons (1a–1g) and three promoter regions [32]. Exon 1a is associated with a strong promoter and transcripts initiated in that site are the most common. Expression of transcripts originating in exons 1c, 1f and 1g is tissue-specific: exon 1f transcripts were detected in kidney, parathyroid adenoma tissue and in intestinal cell line LIM1863 [41], exon 1g transcripts in normal blood and in KG1 leukemic cell line [32,42], while the activity of 1c promoter was identified only in breast cancer cells [43]. Since no distinct promoter activity was recognized for the region adjacent to exons 1d and 1g, it has been postulated that a single promoter upstream of 1a [44] or overlapping regions [41] regulate transcription from these exons. Translation of human VDR protein starts from exon 2 and ends in exon 9. Due to the T to C polymorphism, which eliminates the most 5’-located ATG codon, translation starts from the second in-frame ATG codon in some individuals. As a result, two variants of the VDR protein exist, one of which is three amino-acids shorter (424 aa) than the other (427 aa) [45]. Two more isoforms of VDR protein: VDRB1 (477 aa) and VDRB2 (450 aa) can be generated by alternative splicing of transcripts originating in exon 1d, which contains an ATG translation start codon. The VDRB1 protein was shown to be expressed in several human tissues and may be functionally different from the main VDR isoform [46]. In addition to the polymorphism in exon 2, there are five more polymorphic sites located between the 8 and 9 exons [47,48,49,50] with no obvious functions. There were thousands of studies performed in order to determine associations of given polymorphisms with different phenotypes, but they produced conflicting results reviewed in [51].
The murine Vdr gene is located on chromosome 15 and is less complex than in humans. In the 5′ regulatory region of the murine Vdr gene exons 1 and 2 were identified, with strong homology to human 1a and 1c, respectively [44]. Though exon 1d is well conserved (1d-like), transcripts containing this exon have not been reported in mice [52]. There is one murine VDR protein which consists of 422 aa, its translation starts from exon 3 and ends on exon 10 [53]. Since a polymorphism is a genetic variant that appears in at least 1% of the population, we cannot learn anything about murine polymorphisms, because studies on the representative groups of the murine population have not been performed. However, some differences between inbred strains of mice have been reported. Single nucleotide polymorphism located in exon 8, has been discovered in murine Vdr gene, which changes A to T. This polymorphism is responsible for an amino acid change at position 276. Individuals with an A allele have methionine amino acid and those with T allele have a lysine. It was proposed that mice with A allele have airway resistance (an indication of airway damage) lower than mice with T allele [54].
The regulation of VDR expression in humans is also complex. The promoter associated with the 1c exon (proximal to the coding region of VDR) was shown to be regulated by 1,25D, retinoic acid (RA), estrogens, and phytoestrogens in breast cancer cell lines [43]. The VDR transcripts originating from exon 1a in blood cells are also RA-responsive. Moreover, retinoic acid receptor α (RARα)—as long as it is not activated by the ligand—acts as a suppressor of VDR expression [42]. Interestingly, the sequences of 1a and 1c promoters do not contain any apparent estrogen or RA response elements (ERE or RARE). However, the known positions of RAREs are highly variable—they range from over 10,000 bases upstream to about 8000 bases downstream of the 5’ end of transcription start site [55], or may be located in introns [56]. In mice, Vdr expression is autoregulated by the 1,25D/VDR complex through binding to three conserved regions located 20, 27, and 29 kb downstream of the transcription start site [57]. In normal human hematopoietic cells, the expression of VDR is moderately upregulated (less than in leukemic cells) by RA, and not by 1,25D. In the normal murine hematopoietic cell, the expression of Vdr is robustly autoregulated by 1,25D, but not by RA [32].
3. Conclusions
In recent years, vitamin D and its active metabolite attracted much more attention than just as antirachitic agents. 1,25D has multiple functions, including inhibiting of cell proliferation, enhancing cell differentiation, augmentation of innate immune functions, and reduction of inflammation [28,58]. This is why 1,25D has been proposed as a drug in hyperproliferative and autoimmune diseases. A main limitation to the use of 1,25D as a drug is its calcemic action. The solution to this problem has been in the development of analogs with lowered calcemic and retained prodifferentiating activities [14,15]. Some of the analogs have been already registered for treatment of psoriasis [16,59], which is a hyperproliferative and an autoimmune disease [60]. Any new drug before being introduced to the market must undergo systematic preclinical testing, including experiments in nonhuman subjects to test efficacy, toxicity and pharmacokinetics. Usually, the first species to be used in such experiments are mice, and in fact, there is no comparable replacement to these animals. Despite humans and mice share many similarities in their genomes, anatomy and physiology, one should be aware that there are also differences. It seems that the regulation of VDR expression is one of these differences. In humans, other nuclear receptors and their ligands regulate the expression of VDR gene and as a consequence, an amount of VDR protein, while in mice, only the liganded VDR autoregulates the expression of the Vdr gene and level of VDR protein. The cause of this difference is difficult to assume, but the fact that humans in their natural environment were able to abundantly synthesize vitamin D in their skin, while mice covered with fur and spending days underground had this ability limited, may be important. Thus, if a certain vitamin D analog, or any combination therapy including vitamin D analogs, appears to be toxic in mice, it may be worth examination in other species.
Funding
This research was funded by the National Science Centre of Poland (grant OPUS 2015/17/B/NZ4/02632).
Acknowledgments
The author wants to express her gratitude to all researchers who participated in the research project OPUS 2015/17/B/NZ4/02632. These were Małgorzata Cebrat, Urszula Nowak, Sylwia Janik, Aleksandra Marchwicka, Agnieszka Łaszkiewicz, Michał Majkowski, Łukasz Śnieżewski and obstetricians Magdalena Drychusz and Beata Jabłońska from Medfemina Hospital in Wrocław.
Conflicts of Interest
The author declares no conflict of interest.
References
- Perlman, R. Mouse models of human disease. An evolutionary perspective. Evol. Med. Public Health 2016, 1, 170–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.; MacLaughlin, J.; Clark, M.; Holick, S.; Potts, J.J.; Anderson, R.; Blank, I.; Parrish, J.; Elias, P. Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science 1980, 210, 203–205. [Google Scholar] [CrossRef]
- Mozolowski, W. Jędrzej Śniadecki (1768–1838) on the cure of rickets. Nature 1939, 143, 121–124. [Google Scholar] [CrossRef]
- Prosser, D.; Jones, G. Enzymes involved in the activation and inactivation of vitamin D. Trends Biochem. Sci. 2004, 29, 664–673. [Google Scholar] [CrossRef]
- Aranda, A.; Pascual, A. Nuclear hormone receptors and gene expression. Physiol. Rev. 2001, 81, 1269–1304. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, S.; Dunlop, T.; Sinkkonen, L.; Frank, C.; Carlberg, C. Spatio-temporal activation of chromatin on the human CYP24 gene promoter in the presence of 1alpha,25-dihydroxyvitamin D3. J. Mol. Biol. 2005, 350, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Holick, M. Vitamin D and bone health. J. Nutr. 1996, 126, S1159–S1164. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, E.; Mathieu, C. Immunoregulation by 1,25-dihydroxyvitamin D3: Basic concepts. J. Steroid Biochem. Mol. Biol. 2005, 97, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Grande, A.; Montanari, M.; Tagliafico, E.; Manfredini, R.; Zanocco Marani, T.; Siena, M.; Tenedini, E.; Gallinelli, A.; Ferrari, S. Physiological levels of 1alpha, 25 dihydroxyvitamin D3 induce the monocytic commitment of CD34+ hematopoietic progenitors. J. Leukoc. Biol. 2002, 71, 641–651. [Google Scholar] [PubMed]
- Anderson, M.; Nakane, M.; Ruan, X.; Kroeger, P.; Wu-Wong, J. Expression of VDR and CYP24A1 mRNA in human tumors. Cancer Chemother. Pharmacol. 2006, 57, 234–240. [Google Scholar] [CrossRef]
- Abe, E.; Miamura, C.; Sakagami, H.; Takeda, M.; Konno, K.; Yamazaki, T.; Yoshiki, S.; Suda, T. Differentiation of mouse myeloid leukemia cells induced by 1-alpha,25-dihydroxyvitamin D3. Proc. Natl. Acad. Sci. USA 1981, 78, 4990–4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, V.; Feldman, D. 1,25-dihydroxyvitamin D3 decreases human prostate cancer cell adhesion and migration. Mol. Cell Endocrinol. 2000, 164, 133–143. [Google Scholar] [CrossRef]
- Koeffler, H.; Hirji, K.; Itri, L. 1,25-dihydroxyvitamin D3: In vivo and in vitro effects on human preleukemic and leukemic cells. Cancer Treat Rep. 1985, 69, 1399–1407. [Google Scholar] [PubMed]
- Bouillion, R.; Okamura, W.; Norman, A. Structure-function relationships in the vitamin D endocrine system. Endocr. Rev. 1995, 16, 200–216. [Google Scholar]
- Nadkarni, S.; Chodynski, M.; Corcoran, A.; Marcinkowska, E.; Brown, G.; Kutner, A. Double point modified analogs of vitamin D as potent activators of vitamin D receptor. Curr. Pharm. Des. 2015, 21, 1741–1763. [Google Scholar] [CrossRef]
- Devaux, S.; Castela, A.; Archier, E.; Gallini, A.; Joly, P.; Misery, L.; Aractingi, S.; Aubin, F.; Bachelez, H.; Cribier, B.; et al. Topical vitamin D analogues alone or in association with topical steroids for psoriasis: A systematic review. J. Eur. Acad. Dermatol. Venereol. 2012, 26 (Suppl. 3), 52–60. [Google Scholar] [CrossRef]
- Nuijten, M.; Andress, D.; Marx, S.; Sterz, R. Chronic kidney disease Markov model comparing paricalcitol to calcitriol for secondary hyperparathyroidism: A US perspective. Curr. Med. Res. Opin. 2009, 25, 1221–1234. [Google Scholar] [CrossRef]
- Trynda, J.; Turlej, E.; Milczarek, M.; Pietraszek, A.; Chodyński, M.; Kutner, A.; Wietrzyk, J. Antiproliferative activity and in vivo toxicity of double-point modified analogs of 1,25-dihydroxyergocalciferol. Int. J. Mol. Sci. 2015, 16, 24873–24894. [Google Scholar] [CrossRef] [Green Version]
- AliAbulmeat, M. Sunlight exposure vs. Vitamin D supplementation on bone homeostasis of vitamin D deficient rats. Clin. Nutr. Exp. 2017, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Płudowski, P.; Karczmarewicz, E.; Bayer, M.; Carter, G.; Chlebna-Sokół, D.; Czech-Kowalska, J.; Dębski, R.; Decsi, T.; Dobrzańska, A.; Franek, E.; et al. Practical guidelines for the supplementation of vitamin D and the treatment of deficits in Central Europe—Recommended vitamin D intakes in the general population and groups at risk of vitamin D deficiency. Endokrynologia Polska 2013, 64, 319–327. [Google Scholar] [CrossRef]
- Dee, T.; Hovda, L. Toxicology brief: Cholecalciferol rodenticide toxicosis. Vet. Tech. 2012, E1–E4. [Google Scholar]
- Brown, L.; Marshall, E. Field Evaluation of Quintox (Cholecalciferol) for Controlling Commensal Rodents; University of Nebraska-Lincoln: Lincoln, NE, USA, 1988. [Google Scholar]
- Malloy, P.; Feldman, D. The role of vitamin D receptor mutations in the development of alopecia. Mol. Cell Endocrinol. 2011, 347, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Kaplan, L.; Perwad, F.; Huang, N.; Sharma, A.; Choi, Y.; Miller, W.; Portale, A. Vitamin D 1alpha-hydroxylase gene mutations in patients with 1alpha-hydroxylase deficiency. J. Clin. Endocrinol. Metab. 2007, 92, 3177–3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pirro, A.; Amling, M.; Delling, G.; Baron, R.; Bronson, R.; Demay, M. Targeted ablation of the vitamin D receptor: An animal model of vitamin D-dependent rickets type II with alopecia. Proc. Natl. Acad. Sci. USA 1997, 94, 9831–9835. [Google Scholar] [CrossRef] [Green Version]
- Panda, D.; Miao, D.; Tremblay, M.; Sirois, J.; Farookhi, R.; Hendy, G.; Goltzman, D. Targeted ablation of the 25-hydroxyvitamin D 1alpha-hydroxylase enzyme: Evidence for skeletal, reproductive, and immune dysfunction. Proc. Natl. Acad. Sci. USA 2001, 98, 7498–7503. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, C. The physiology of vitamin D-far more than calcium and bone. Front. Physiol. 2014, 5, 335. [Google Scholar] [CrossRef] [Green Version]
- Studzinski, G.; Wang, X.; Ji, Y.; Wang, Q.; Zhang, Y.; Kutner, A.; Harrison, J. The rationale for deltanoids in therapy for myeloid leukemia: Role of KSR-MAPK-C/EBP pathway. J. Steroid Biochem. Mol. Biol. 2005, 97, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Marcinkowska, E.; Garay, E.; Gocek, E.; Chrobak, A.; Wang, X.; Studzinski, G. Regulation of C/EBPbeta isoforms by MAPK pathways in HL60 cells induced to differentiate by 1,25-dihydroxyvitamin D3. Exp. Cell Res. 2006, 312, 2054–2065. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, C.; Seuter, S.; de Mello, V.; Schwab, U.; Voutilainen, S.; Pulkki, K.; Nurmi, T.; Virtanen, J.; Tuomainen, T.; Uusitupa, M. Primary vitamin D target genes allow a categorization of possible benefits of vitamin D3 supplementation. PLoS ONE 2013, 8, e71042. [Google Scholar] [CrossRef] [Green Version]
- Janik, S.; Nowak, U.; Łaszkiewicz, A.; Satyr, A.; Majkowski, M.; Marchwicka, A.; Śnieżewski, Ł.; Berkowska, K.; Gabryś, M.C.M.; Marcinkowska, E. Diverse regulation of vitamin D receptor gene expression by 1,25-dihydroxyvitamin D and ATRA in murine and human blood cells at early stages of their differentiation. Int. J. Mol. Sci. 2017, 18, 1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erben, R.; Zeitz, U.; Weber, K.; Stierstorfer, B.; Wolf, G.; Schmahl, W.; Balling, R.; Quintanilla-Martinez, L. A non-functioning vitamin D receptor predisposes to leukaemoid reactions in mice. Hematol. Oncol. 2010, 28, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Holick, M. Vitamin D: A millenium perspective. J. Cell Biochem. 2003, 88, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Newmark, H.; Dantoft, W.; Ghazal, P. Evolutionary origin of the interferon–immune metabolic axis: The sterol–vitamin D link. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Kollitz, E.; Zhang, G.; Hawkins, M.; Whitfield, G.; Reif, D.; Kullman, S. Evolutionary and functional diversification of thevitamin D receptor-lithocholic acid partnership. PLoS ONE 2016, 11, e0168278. [Google Scholar] [CrossRef] [Green Version]
- Makishima, M.; Lu, T.; Xie, W.; Whitfield, G.; Domoto, H.; Evans, R.; Haussler, M.; Mangelsdorf, D. Vitamin D receptor as an intestinal bile acid sensor. Science 2002, 296, 1313–1316. [Google Scholar] [CrossRef] [Green Version]
- Cruzeiro, C.; Lopes-Marques, M.; Ruivo, R.; Rodrigues-Oliveira, N.; Santos, M.; Rocha, M.; Rocha, E.; Castro, L. A mollusk VDR/PXR/CAR-like (NR1J) nuclear receptor provides insight into ancient detoxification mechanisms. Aquat. Toxicol. 2016, 174, 61–69. [Google Scholar] [CrossRef]
- Kollitz, E.; Zhang, G.; Hawkins, M.; Whitfield, G.; Reif, D.; Kullman, S. Molecular cloning, functional characterization, and evolutionary analysis of vitamin D receptors isolated from basal vertebrates. PLoS ONE 2015, 10, e0122853. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.; McDonnell, D.; Hughes, M.; Crisp, T.; Mangelsdorf, D.; Haussler, M.; Pike, J.; Shine, J.; O’Malley, B. Cloning and expression of full-length cDNA encoding human vitamin D receptor. Proc. Natl. Acad. Sci. USA 1988, 85, 3294–3298. [Google Scholar] [CrossRef] [Green Version]
- Crofts, L.; Hancock, M.; Morrison, N.; Eisman, J. Multiple promoters direct the tissue-specific expression of novel N-terminal variant human vitamin D receptor gene transcripts. Proc. Natl. Acad. Sci. USA 1998, 95, 10529–10534. [Google Scholar] [CrossRef] [Green Version]
- Marchwicka, A.; Cebrat, M.; Łaszkiewicz, A.; Śnieżewski, Ł.; Brown, G.; Marcinkowska, E. Regulation of vitamin D receptor expression by retinoic acid receptor alpha in acute myeloid leukemia cells. J. Steroid Biochem. Mol. Biol. 2016, 159, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, I.; Flanagan, L.; Tenniswood, M.; Welsh, J. Identification of a hormone-responsive promoter immediately upstream of exon 1c in the human vitamin D receptor gene. Endocrinology 2000, 141, 2829–2836. [Google Scholar] [CrossRef] [PubMed]
- Halsall, J.; Osborne, J.; Hutchinson, P.; Pringle, J. In silico analysis of the 5’ region of the vitamin D receptor gene: Functional implications of evolutionary conservation. J. Steroid Biochem. Mol. Biol. 2007, 103, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Kesterson, R.; Yamamoto, H.; Taketani, Y.; Nishiwaki, E.; Tatsumi, S.; Inoue, Y.; Morita, K.; Takeda, E.; Pike, J. Structural organization of the human vitamin D receptor chromosomal gene and its promoter. Mol. Endocrinol. 1997, 11, 1165–1179. [Google Scholar] [CrossRef]
- Sunn, K.; Cock, T.; Crofts, L.; Eisman, J.; Gardiner, E. Novel N-terminal variant of human VDR. Mol. Endocrinol. 2001, 15, 1599–1609. [Google Scholar] [CrossRef]
- Ye, W.; Reis, A.; Velho, G. Identification of a novel Tru9 I polymorphism in the human vitamin D receptor gene. J. Hum. Genet. 2000, 45, 56–57. [Google Scholar] [CrossRef] [Green Version]
- Morrison, N.; Qi, J.; Tokita, A.; Kelly, P.; Crofts, L.; Nguyen, T.; Sambrook, P.; Eisman, J. Prediction of bone density from vitamin D receptor alleles. Nature 1994, 367, 284–287. [Google Scholar] [CrossRef]
- Morrison, N.; Yeoman, R.; Kelly, P.; Eisman, J. Contribution of trans-acting factor alleles to normal physiological variability—Vitamin-D receptor gene polymorphisms and circulating osteocalcin. Proc. Natl. Acad. Sci. USA 1992, 89, 6665–6669. [Google Scholar] [CrossRef] [Green Version]
- Faraco, J.; Morrison, N.; Baker, A.; Shine, J.; Frossard, P. ApaI dimorphism at the human vitamin-D receptor gene locus. Nucleic Acids Res. 1989, 17, 2150. [Google Scholar] [CrossRef]
- Valdivielsoa, J.; Fernandez, E. Vitamin D receptor polymorphisms and diseases. Clin. Chim. Acta 2006, 371, 1–12. [Google Scholar] [CrossRef]
- Gardiner, E.; Esteban, L.; Fong, C.; Allison, S.; Flanagan, J.; Kouzmenko, A.; Eisman, J. Vitamin D receptor B1 and exon 1d: Functional and evolutionary analysis. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Burmester, J.; Maeda, K.; DeLuca, H. Isolation of expression of rat 1,25-dihydroxyvitamin D3 receptor cDNA. Proc. Natl. Acad. Sci. USA 1988, 85, 1005–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndt, A.; Savage, H.; Stearns, T.; Paigen, B. Genetic analysis of lung function in inbred mice suggests vitamin D receptor as a candidate gene. Mol. Genet. Genom. 2011, 286, 237. [Google Scholar] [CrossRef] [Green Version]
- Balmer, J.; Blomhoff, R. A robust characterization of retinoic acid response elements based on a comparison of sites in three species. J. Steroid Biochem. Mol. Biol. 2005, 96, 347–354. [Google Scholar] [CrossRef]
- Hua, S.; Kittler, R.; White, K. Genomic antagonism between retinoic acid and estrogen signaling in breast cancer. Cell 2009, 137, 1259–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zella, L.; Kim, S.; Shevde, N.; Pike, J. Enhancers located within two introns of the vitamin D receptor gene mediate transcriptional autoregulation by 1,25-dihydroxyvitamin D3. Mol. Endocrinol. 2006, 20, 1231–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christakos, S.; Hewison, M.; Gardner, D.; Wagner, C.; Sergeev, I.; Rutten, E.; Pittas, A.; Boland, R.; Ferrucci, L.; Bikle, D. Vitamin D: Beyond bone. Ann. N. Y. Acad. Sci. 2013, 1287, 45–58. [Google Scholar] [CrossRef]
- Girolomoni, G.; Vena, G.; Ayala, F.; Cannavò, S.; De Pità, O.; Chimenti, S.; Peserico, A. Consensus on the use of the fixed combination calcipotriol/betamethasone dipropionate in the treatment of plaque psoriasis. G. Ital. Dermatol. Venereol. 2012, 147, 609–624. [Google Scholar]
- Ayala-Fontánez, N.; Soler, D.; McCormick, T. Current knowledge on psoriasis and autoimmune diseases. Psoriasis (Auckl) 2016, 6, 7–32. [Google Scholar]
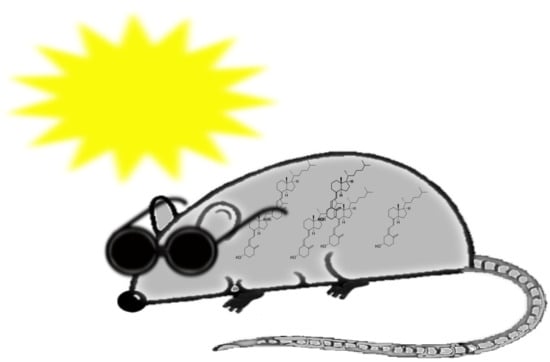
Figure 1.
Outline of vitamin D metabolism. After exposure to sun, vitamin D is synthesized in the skin from 7-dehydrocholesterol. Alternatively, vitamin D may be obtained from diet. Vitamin D is converted to 25-hydroxyvitamin D in the liver, and then to 1,25-dihydroxyvitamin D in the kidneys.
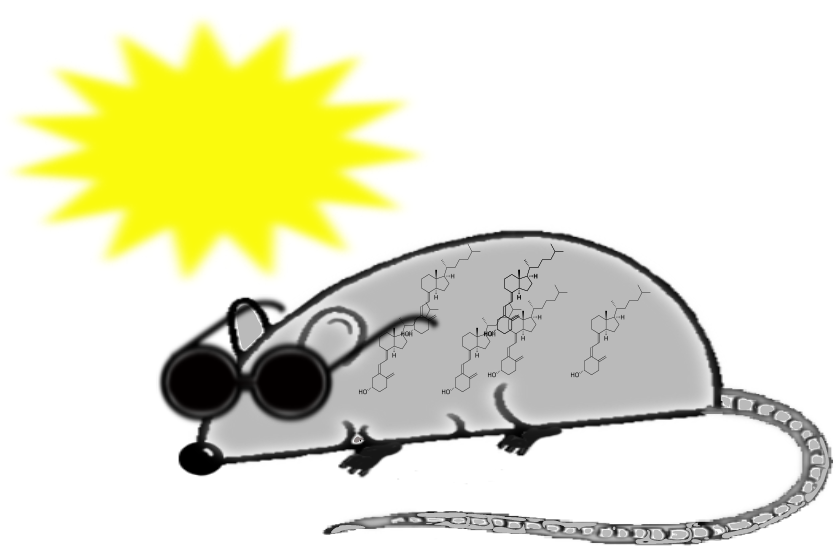
Figure 1.
Outline of vitamin D metabolism. After exposure to sun, vitamin D is synthesized in the skin from 7-dehydrocholesterol. Alternatively, vitamin D may be obtained from diet. Vitamin D is converted to 25-hydroxyvitamin D in the liver, and then to 1,25-dihydroxyvitamin D in the kidneys.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marcinkowska, E. The Vitamin D System in Humans and Mice: Similar but Not the Same. Reports 2020, 3, 1. https://0-doi-org.brum.beds.ac.uk/10.3390/reports3010001
AMA Style
Marcinkowska E. The Vitamin D System in Humans and Mice: Similar but Not the Same. Reports. 2020; 3(1):1. https://0-doi-org.brum.beds.ac.uk/10.3390/reports3010001
Chicago/Turabian StyleMarcinkowska, Ewa. 2020. "The Vitamin D System in Humans and Mice: Similar but Not the Same" Reports 3, no. 1: 1. https://0-doi-org.brum.beds.ac.uk/10.3390/reports3010001