Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health
by
 ,
,
Zahida H. Pervaiz
1, 
Javed Iqbal
2,
Qingming Zhang
3,
Dima Chen
4,
Hui Wei
5 and
Muhammad Saleem
6,* 1
Department of Biological Sciences, Auburn University, Auburn, AL 36849, USA
2
Department of Agronomy and Horticulture, University of Nebraska-Lincoln, Lincoln, NE 68583, USA
3
College of Plant Health and Medicine, Qingdao Agricultural University, Qingdao 266109, China
4
College of Biological and Pharmaceutical Sciences, China Three Gorges University, Yichang 443002, China
5
Department of Ecology, College of Natural Resources and Environment, South China Agricultural University, Guangzhou 510642, China
6
Department of Biological Sciences, Alabama State University, Montgomery, AL 36104, USA
*
Author to whom correspondence should be addressed.
Soil Syst. 2020, 4(4), 59; https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems4040059
Submission received: 20 July 2020
/
Revised: 14 September 2020
/
Accepted: 15 September 2020
/
Published: 23 September 2020
(This article belongs to the Special Issue Role of Soil Organisms in Modification of their Environment, Nutrient Cycling and Soil Formation)
Abstract
:The continuous cropping (CC) of major agricultural, horticultural, and industrial crops is an established practice worldwide, though it has significant soil health-related concerns. However, a combined review of the effects of CC on soil health indicators, in particular omics ones, remains missing. The CC may negatively impact multiple biotic and abiotic indicators of soil health, fertility, and crop yield. It could potentially alter the soil biotic indicators, which include but are not limited to the composition, abundance, diversity, and functioning of soil micro- and macro-organisms, microbial networks, enzyme activities, and soil food web interactions. Moreover, it could also alter various soil abiotic (physicochemical) properties. For instance, it could increase the accumulation of toxic metabolites, salts, and acids, reduce soil aggregation and alter the composition of soil aggregate-size classes, decrease mineralization, soil organic matter, active carbon, and nutrient contents. All these alterations could accelerate soil degradation. Meanwhile, there is still a great need to develop quantitative ranges in soil health indicators to mechanistically predict the impact of CC on soil health and crop yield gaps. Following ecological principles, we strongly highlight the significance of inter-, mixture-, and rotation-cropping with cover crops to sustain soil health and agricultural production.
1. Introduction
The existence of humanity is intertwined with the future of Earth’s soil health. There is an emerging consensus that the food security over the next century will be severely threatened by the destructive agronomic practices, which may amplify the adverse effects of climate and land-use change on soil ecosystems [1,2]. Therefore, maintaining soil health is central to meeting the demands of the growing population. However, the CC of major agricultural, horticultural, and industrial crops has been associated with significant soil health-related concerns. It is defined as the plantation of the same crop species over the years, mainly as a monoculture in the same agricultural field [3,4]. The agricultural crops, if grown continuously, could suffer from reduced plant growth and yield [5]. However, the impacts of CC on soil health are context-dependent in the sense that they depend on crop types (food, energy, annual, perennial), duration, soil, and climatic factors. Nevertheless, CC could lead to crop yield penalties due to the alteration in soil physicochemical properties and microbial communities with time.
Generally, in CC systems, growers employ standard and persistent agronomic practices over the years. For instance, the use of similar agrochemicals (pesticides, fertilizers), farm machinery, tillage depth, irrigation, and harvesting approaches [3,6]. Therefore, the standard farm management practices could lead to the selection and evolution of certain weeds, insect pests, and phytopathogens. Moreover, this type of cropping system not only depletes major soil nutrients such as nitrogen (N), phosphorus (P), and potassium (K), but it also reduces the contents of essential minerals in the agricultural soils [7,8]. Meanwhile, the CC could cause the accumulation of a similar type of plant-derived exudates. This phenomenon may lead to soil autotoxicity and reduced plant growth [9,10]. Furthermore, this practice could also reduce the diversity and inputs of crop residues into soil, which may also alter soil aggregation and physicochemical properties. Overall, changes in soil physicochemical properties under CC could ultimately alter soil biological properties. For instance, it may reduce the abundance of beneficial soil organisms over time [11,12], which are essential to soil fertility, health, and crop yield.
The CP may increase and decrease the abundance of soil-borne pathogens and beneficial microbes, respectively, in agricultural soils. Moreover, it may change the soil physicochemical properties, such as nutrient balance, pH, and structure, thus leading to overall soil degradation and reduction in crop yield [13,14,15]. For instance, the CC of corn and soybean caused yield penalties across hundreds of thousands of agricultural fields in the U.S., and researchers attributed this phenomenon to changes in soil and other environmental conditions [16]. Similarly, in another study, a reduction in sorghum yield under CC was attributed to the alteration in soil microbial communities, particularly, due to an increased abundance of pathogenic taxa in the soil [17]. It may also alter below-ground soil–plant interactions by exerting similar rhizospheric effects. For instance, a similar type of root exudation may lead to the accumulation of toxic metabolites, and ultimately soil auto-toxicity with time. The soil auto-toxicity may negatively affect soil health and crop yield [10,18,19,20].
In this review article, we review the effects of CC on soil biotic and abiotic properties, while advocating that it may alter soil health (Figure 1). Overall, we attempt to develop a broader understanding of the effects of CC on soil biome, followed by its impacts on soil physicochemical properties. Meanwhile, we also highlight the significance of alternative cropping strategies, such as inter-, mixture-, and rotation-cropping with cover crops in soil health.
2. Indicators of Soil Health
Soil health is a very a broad concept, and it is defined as the sustained capacity of agricultural soils to function and thrive as a healthy living ecosystem that supports microbes, plants, insects, and animals in a way which is desirable to and meets the demands of human beings [21,22,23,24,25]. Below, we discuss the impact of CC on soil biotic and abiotic health indicators. Meanwhile, we also provide a comprehensive list of soil health indicators and their responses to CC in the context of soil conservation (Table 1).
3. Biotic Indicators of Soil Health
The biotic indicators of soil health may include but are not limited to total biomass, activities, functioning, community composition, and interactions of soil-inhabiting macro-organisms and microorganisms that determine the trophic or food web complexity of soil ecosystems. At present, some studies have reported the impacts of CC on soil bacteria and fungi. However, we know little about the responses of other microbial groups such as viruses and protists to CC under a broad range of soil and environmental conditions. Both viruses and protists are micro-predators of microbial ecosystems, and they regulate and control soil microbial communities and their functions, which are essential to soil health and crop yield.
3.1. Microbiomic Indicators
Soil microbial communities regulate several biological functions, such as disease or pest suppression, crop plant nutrition, and resilience to anthrophonic and climatic changes [26,27]. Moreover, they also improve several abiotic indicators of soil health, such as soil aggregation, breakdown of crop residues, nutrient mineralization, and fixation. However, CC could negatively affect several parameters of soil microbial communities and their functions, as discussed below.
3.1.1. Microbial Biomass or Abundance
The microbial biomass or abundance is an essential component of the healthy soil ecosystem, and its contents determine the soil quality and crop yields [103,104]. A reduction in the soil microbial biomass as a function of CC not only decreases the abundance of beneficial microbial taxa, but it may also lead to a decline in general soil functions, which are critical for maintaining soil health and productivity [105]. For instance, there is an emerging consensus that the abundance of soil fungi and bacteria increases and decreases, respectively, under CC with time [106]. Specifically, the CC reduces the abundance of important beneficial bacterial taxa that deliver the vital soil ecosystem services such as N-fixing and disease suppression [107]. Regarding soil fungi, for instance, a continuous plantation of commodity crops reduces the abundance of important soil fungal taxa (e.g., Gliocladium and Trichoderma spp.), which act as biological control agents against soil-borne pathogens (e.g., Fusarium spp.) [108,109]. Similarly, some studies have also reported that long-term CC may also decrease the microbial biomass P [110] and C contents [111].
Moreover, in general, some classical comparative studies have also shown relatively lower soil microbial biomass under a continuous rather than rotation cropping system [112,113,114], and these effects were attributed to lower and higher resources or crop residues for microbes under monoculture rather than mixture cropping [114]. Overall, a reduction in microbial biomass under CC may predict a reduced input of organic materials and crop residues into soil, and it highlights the need for increasing cropping diversity to improve the soil organic matter and microbial biomass contents.
3.1.2. Microbial Community Composition and Diversity
The composition and diversity of soil microbes may influence microbial-driven soil functions, which determine soil fertility and crop productivity [26,115,116,117]. Previous research has demonstrated that agricultural intensification may alter the diversity, composition, and functioning of microbial communities in soils under different crops [108,118,119]. For instance, CC of tea plants from five to eight years increased the microbial community composition, though it was negatively affected in the long term (50-year-old to 90-year-old) [120]. In another study, authors showed that the relative abundance of important bacterial phyla such as Proteobacteria, Actinobacteria, and Firmicutes increased in the soils under wheat-corn-soybean crop rotations than in soil under continuous soybean cropping [121]. Moreover, the CC of tea plants also reduced Shannon’s diversity index of soil microbial communities [122].
Moreover, the CC may also alter microbial community structure by changing the abundance of certain microbial taxa; for instance, Tang et al., (2009) reported that Actinobacteria became more abundant in agricultural soils under soybean-corn rotation than under continuous soybean cropping [123]. They reported significant changes in the composition of soil microbial communities under canola rather than wheat monocultures, which suggested a monoculture-specific effect of CC on the soil microbial diversity [124]. In another study, the continuous banana cropping tended to decrease the soil bacterial diversity, and it altered the bacterial community structure [79]. Similarly, the CC of legume pea plants changed the composition of soil microbial communities by negatively affecting the abundance of arbuscular mycorrhizal fungi and gram-positive bacteria [125]. Similarly, the CC of cotton increased the abundance of certain soil fungal taxa such as Ascomycota [126], while the differential effects of monoculture cropping systems such as soybean [127], peanut [128], and vanilla [129] on soil microbial community composition and diversity were also reported. A recent study investigated the impact of Bacillus amyloliquefaciens on soil microbial communities under sorghum–maize rotation, sorghum, and maize CC systems [17]. The microbial communities of rhizosphere soil under continuous sorghum cropping showed lower and higher bacterial and fungal diversity, respectively, than the soil under a sorghum–maize rotation system. They reported that some bacterial phyla such as Acidobacteria and Gemmatimonadetes, in addition to some fungal taxa such as Basidiomycota, showed a greater abundance under the CC system. Though fungal taxa belonging to class Tremellomycetes was the most dominant group under CC, the use of B. amyloliquefaciens significantly reduced its abundance in the soil under CC [17]. Another study investigated the root-endophytic and rhizospheric bacteria under the CC of Panax notoginseng [130]. They reported that the CC of P. notoginseng reduced the abundance of rhizospheric bacteria, though it did not affect the endophytic bacteria significantly. Moreover, they reported that the bacterial diversity was higher in healthy than infected rhizospheric soils under plantation of P. notoginseng. But in general, the CC increased the abundance of some bacterial phyla such as Proteobacteria, Cyanobacteria, Actinobacteria, and Acidobacteria, and the alterations in microbial community composition were determined by the changes in the soil properties such as total P, pH, and soil organic matter contents [130].
Though several studies have reported the impact of CC on the composition of soil microbial communities, some studies did not find differences between the composition of soil microbial communities under monoculture or rotation cropping [131,132,133]. The variable impacts of CC on soil microbial communities could be attributed to different research methods, types of cropping systems, planting years, soil, and climatic conditions [133]. We suggest that the continuous plantation of crops, either legumes or non-legumes (agricultural, horticultural, medicinal, industrial), may negatively affect the species diversity, composition, and functioning of soil microbial communities. In general, CC may increase the abundance of soil fungi while the converse is true for soil bacteria, though further research is needed to elucidate its impacts on microbial functions. Moreover, little is known about the impact of CC on other important soil microbial groups such as soil protists and viruses. Therefore, future research should also focus on these microbial groups since they determine microbial trophic (predation, grazing) and non-trophic (competition) interactions, as these microorganisms are highly sensitives to the land-use changes.
3.2. Soil Enzyme Activities
Soil microbes and plant roots produce a tremendous diversity of extracellular enzymes [134], and these enzymes play a major role in catalyzing biochemical reactions in the soil. Therefore, the activities of enzymes determine the intensity and magnitude of soil biogeochemical processes [134,135]. The activities of soil enzymes regulate several properties of soil ecosystems depending on soil, vegetation, and climatic conditions. Other than influencing soil edaphic properties, soil enzymes may also predict the biological health of soil ecosystems. For instance, the soil sucrase activities demonstrated a strong negative correlation with the abundance of the important soil-borne pathogen, such as Fusarium spp. [136]. That is why the soil enzymes are considered key indicators of soil health, sustainability, and functioning [134,137]. The soil enzyme-driven processing of organic matter is critical for nutrient cycling and sequestration of essential elements, such as soil C. Given that soil enzyme activities are very sensitive to land-use changes, a better understanding of the sensitivity of soil enzymes to continuous monoculture cropping is essential for the management of agricultural soils in present scenarios of agricultural intensification and climate warming [138]. For instance, soil dehydrogenase activity is known to decrease in the soil under CC [111]. The CC of important crops such as potato and cotton could substantially reduce the activities of important soil enzymes such as urease, alkaline phosphatase, and sucrose [139,140]. Dou et al. (2016) investigated the impact of CC of sorghum, cotton, corn, and cotton/sorghum rotations on soil enzyme activities after 26 years of cropping. They reported that the activities of soil enzymes, such as arylsulfatase, alkaline phosphatase, and -d-glucosidase, were highest and lowest in soils under sorghum and cotton monoculture cropping. Overall, the soil enzyme activities were lower in soils under monoculture than rotation cropping [141]. Similarly, in another study, the CC also reduced the activities of urease, dehydrogenase, and catalase in agricultural soil [142]. Though several examples can be found regarding the impact of CC on soil enzyme activities, we suggest that monoculture cropping may reduce the activities of vital soil enzymes, which are essential for soil biological and biogeochemical health.
3.3. Abundance, Composition, and Diversity of Soil Macro-Organisms
Soil macro-organisms consist of several invertebrate animal groups, such as mites, nematodes, and earthworms, while these organisms demonstrate a remarkable complexity in their biotic and abiotic interactions in the soil environment. These organisms drive many trophic (predation) and non-trophic (competition, facilitation) interactions, which overall determine their top-down effects on soil microbial communities in the soil food web. These organisms may influence several vital soil processes such as the decomposition of crop residues, mineralization, breakdown of humic substance, bioturbation, bioengineering of soil structure, immobilization of nutrients, nutrient cycling, and biological N-fixation. However, unfortunately, the CC coupled with destructive and homogenous farm management practices may negatively affect the composition, diversity, and functioning of these organisms since they are more sensitive to agriculture-driven soil disturbances [143,144,145]. Moreover, at present, the impact of CC on soil macro-organisms is mainly limited to soil nematodes and earthworms, whereas further work on other groups of soil macro-organisms such as mites, soil-dwelling insects, and other animals is limited and merits future research.
3.3.1. Soil Nematodes
Nematodes are one of the dominant groups of soil macro-organisms, and their significance in soil food web interactions and soil ecosystem functioning is established. The community composition and functional attributes of nematodes play an important role in nutrient recycling, processing of crop residues, soil aggregation, aeration, and priming [146,147]. The agronomic practices and soil disturbances are considered to negatively affect the soil nematodes [148,149]. For instance, the CC of strawberry reduced the diversity of soil nematodes, while the accumulation of toxic metabolites inhibited the abundance of some nematode taxa, such as Acrobeloides spp. [18]. In another study, Tian et al. (2019) investigated the relative abundance of bacterial-feeding and plant-parasitic nematodes after 2 and 10 years of CC under greenhouse conditions. Regarding α-diversity, the bacterial eating nematodes were dominant in the two years across the whole growing season, whereas the plant-parasitic nematodes were dominant after 10 years in the late growing season [150]. Moreover, the meloidogyne and rhabditidae became dominant after 10- and 2-year cropping. Similarly, the β-diversity of soil nematodes also differed after the 10-year and 2-year CC. Some soil properties such as organic matter, N, and other biotic conditions also influenced the composition of soil nematodes under CC [150]. These examples suggest that CC may alter the diversity and composition of soil nematodes, while these effects may be driven by both top-down (plants and predator derived effects) and bottom-up effects under changing soil conditions. For instance, CC could potentially reduce soil pH due to, among others, N-enrichment, which could increase and decrease fungal and bacterial abundance, thus leading to an overabundance of fungal rather than bacterial eating nematodes [20].
3.3.2. Soil Earthworms
As one of the most common soil invertebrate organisms, earthworms are called the biological engineers of soil ecosystems because they can alter the soil biophysical properties to increase soil health and crop productions. They play a major role in improving the soil conditions, such as breakdown of organic substance, soil aggregation, aeration, nutrient cycling and sequestration, microbial community composition, and functioning [151,152]. Meanwhile, earthworms are considered to be sensitive to the climatic conditions [153], soil moisture, organic matter [154,155], physicochemical properties [156], nutrient resources [157], heavy metals [158], organic pollutants [159,160], and trophic interactions (predation) [161]. The repeated tillage practices under CC may also affect their abundance and functioning in the agricultural soils [162]. Moreover, it is also commonly perceived that conventional CC practices decrease the biodiversity of soil earthworms more than conservation cropping practices [163,164]. Another study also reported that continuous cropping of cotton reduced the abundance of soil earthworms; among these, the abundance of Diplocardia caroliniana and Diplocardia caroliniana decreased up to 55–68%, and 88%, respectively [59]. Moreover, the total earthworm numbers were lower in the cotton-corn than corn-soybean monoculture sequence cropping, which also suggested a negative effect of cotton soil earthworms. However, in cotton-soybean monoculture sequence cropping, the earthworm abundance did not differ among monoculture treatments, which may suggest that legume plants have a positive effect on soil earthworms [165]. The relatively positive and negative effects of soybean and cotton on earthworms could be attributed to the N availability and accumulation of toxic metabolites in soils, respectively. In another study, the soil under wheat-clover rather than wheat mono-cropping showed twice as large and more abundant earthworm species [166]. Moreover, the relatively higher abundance, biomass per individual, and species diversity of earthworms were seen in the soil under winter wheat plantation in an organic rather than conventional cropping system [167], which may also suggest that conventional agronomic practices may amplify the impact of monoculture cropping on soil earthworms. Moreover, higher abundance and diversity of earthworms were linked to higher amounts of organic matter in the monocultures of organic rather than conventional cropping [167]. Though there is not much research on the functional ecology of earthworms in monoculture cropping systems, recent research suggests that agricultural intensification in the form of monoculture cropping may negatively affect the composition, diversity, and functioning of soil earthworms, depending on soil, crop, and environmental conditions.
3.3.3. Soil Mites and Other Organisms
Mites (Acari) constitute the most abundant group of arthropods in soil throughout the world and can reach up to 100,000 individuals per m2 [168]. Because soil mites respond relatively quickly to land-use changes, these are considered as important indicators of soil quality and health [169,170]. Few studies have investigated the impact of monoculture cropping on soil mites, whereas legumes crops may favor certain groups of mites [171] due to greater N availability. Gruss et al. (2017) investigated the impact of a single crop, potato, or winter rye on the assemblage of soil-inhabiting mites after 90 years of CC. Different taxa of mites showed preferences for certain crops. For instance, a higher abundance mite (Prostigmata) was observed in potato, while Oribatida and Gamasida were more abundant in the winter rye [172]. Similarly, some studies have also investigated the impact of CC on other soil mesofauna groups such as springtails [173,174]. However, comparative studies are needed to investigate the effects of CC in comparison to other cropping systems (inter, mixture, rotation) on soil mites. Moreover, integrated studies involving different organisms at various soil trophic levels are needed to elucidate the impact of CC on the soil food web complexity and functioning.
3.4. Increased Incidences of Soil-Borne Diseases and Evolution of Disease Suppressive Soils
The soil-borne diseases are caused by several groups of soil microorganisms such as viruses, bacteria, fungi, and protists, while soil macro-organisms, such as nematodes, could also cause this phenomenon. Moreover, some free-living elements, such as prions and nucleic acids (micro-RNAs), may also cause plant diseases, though we know little about their role in soil-borne diseases. The CC may increase the incidents of soil-borne diseases, and in some cases, it may also lead to the evolution of disease-resistant soils, as discussed below.
3.4.1. Soil-Borne Fungal Diseases
Most soil-borne diseases are caused by fungi under the CC systems. For instance, pathogenic fungi caused soil-borne diseases in the continuous peanut [13] and soybean cropping [127] systems by reducing the abundance of anti-pathogenic taxa [119,175]. In another study, the soil fungal diversity was significantly reduced after three years of continuous notoginseng cropping. Moreover, a negative relationship between plant death rate and the fungal diversity was also observed, thus implying that higher fungal diversity is a key biotic indicator of soil health [176]. However, the relative abundance of Fusarium oxysporum and Phaeosphaeria rousseliana positively correlated with the death rate of P. notoginseng [176]. A reduction in soil fungal diversity under CC may also alter the community composition of soil fungi; for example, the continuous soybean cropping caused root rot by increasing the abundance of Fusarium in soil [177]. Though there are several examples of soil-borne fungal diseases under CC that we cannot discuss, we still can provide some examples to explain this topic. For instance, the take-all of wheat (Triticum aestivum) is an important fungal disease caused by Gaeumannomyces graminis (Sacc.) Arx and Olivier var. tritici Walker (Ggt) [178]. The soil sickness caused by this disease increased with the long-term monoculture cropping of wheat [5,179], which also reduced crop yield [180]. Similarly, an increased abundance of fungal pathogens coupled with simplification and reduction of soil beneficial fungal community decreased the growth and yield of continuous peanut crop [181]. In another study, a large-scale field study reported that the soybean root rot disease increased dramatically after less than three years as compared to the soil under crop rotation [182]. Similarly, under the CC of industrial food crops such as vanilla, the emergence of vanilla stem wilt was reported due to increased and decreased abundance of pathogenic (Fusarium) and beneficial microbes, respectively, in soil with the time [129]. Shen et al. (2018) investigated bacterial and fungal communities of soils under CC of banana monoculture [183]. The CC of banana was significantly linked to the emergence of Fusarium wilt. The soil fungal rather than bacterial communities were more responsive to the banana consecutive monoculture cropping. Unlike most other studies as discussed above, higher fungal species richness was significantly linked with the higher prevalence of banana Fusarium wilt and reduction in crop yield, thus suggesting a strong antagonistic effect of soil fungi on the banana crop. Specifically, the abundance Fusarium and Phyllosticta showed a strong correlation with the reduction in the banana yield [183]. Similarly, Gao et al. (2019) investigated the impact of continuous sweet potato cropping on soil fungal communities. They showed that the fungal diversity and richness significantly increased in soil under CC, while the abundance of fungal phyla such as Ascomycota decreased with time. Overall, the abundance of beneficial fungi such as Chaetomium decreased but that of pathogenic taxa such as Verticillium, Fusarium, and Colletotrichum increased under in soil under CC [184]. Both results of [184] and [183] are opposite to studies showing a positive role of relatively higher fungal diversity in soil disease suppression, which nevertheless suggests that fungal species identity rather than diversity may be important for soil health, though further research is needed to elucidate the role of fungal community parameters in the emergence and suppression of soil-borne diseases under a broad range of soil and crop types.
3.4.2. Soil-Borne Bacterial Diseases
Similar to soil fungi, alterations in the bacterial communities may also impair soil health under the CC systems. The soil-borne bacterial diseases may damage crop plant and overall yield, for instance, the bacterial wilt can reduce potato yield drastically [185]. The alterations in soil microbial communities are often linked to the occurrence of soil-borne bacterial diseases, such as bacterial wilt [186,187]. Moreover, some studies have also shown that microbial communities in the healthy rather than diseased rhizosphere soils were more richness in species and diverse, thus suggesting that species-rich soil microbiome may exclude pathogens and narrow down the ecological niches of soil-borne pathogens [26,188]. The continuous monoculture cropping may affect the composition of soil microbial communities. Zhang et al. (2011) investigated soil microbial communities of healthy and diseased cotton fields at different plant growth stages. They reported that the microbial communities in the rhizosphere of healthy rather than diseased cotton fields were more species-rich. The higher evenness of microbial communities was observed in the soil of diseased cotton plants, thus suggesting a variable relationship of microbial community composition with the soil sickness. Moreover, the soil of diseased cotton plants showed a higher abundance of bacterial phyla such as Deinococcus-Thermus and Firmicutes [189]. In another study, Wu et al. (2015) showed that the soil of sick rather than healthy P. notoginseng showed a reduction in the microbial diversity at the alpha diversity level. Moreover, soil bacterial community composition also differed between healthy and diseased soil [190]. The long-term CC of cash crops such as wheat, corn, soybean, tobacco, and sorghum could potentially cause soil degradation [191]. Soil-borne bacterial diseases could disturb soil-root interactions, which may lead to soil degradation. Therefore, as compared to healthy soils, the degraded soils may show a relatively lower diversity of beneficial microorganisms [192]. She et al. (2017) investigated bacterial communities in soil under tobacco CC. They showed that bacterial diversity indices such as species richness and evenness were reduced. However, the abundance of the soil-borne pathogen such as Ralstonia spp. increased, and it was positively correlated with the tobacco bacterial wilt disease [193]. In another study, Wang et al. (2018) investigated the effect of N addition and bacterial wilt disease on soil bacterial community parameters under continuous sesame cropping. Both N-addition and bacterial wilt altered bacterial composition and structure [194]. Though CC may induce soil-borne bacterial diseases with time, more research is needed to elucidate its impacts on soil-borne bacterial diseases.
3.4.3. Soil-Borne Nematode Infections
Soil-borne nematode infections may also affect soil health. The CC may increase nematode infections to crop plants with time. Among these, the soybean cyst nematode (SCN; Heterodera glycines Ichinohe) is an obligate sedentary parasite that causes substantial crop losses [195]. There is some evidence that suggests a relatively higher abundance of pathogenic fungi in SNC and their infections in soil under continuous monoculture cropping [6,196].
3.4.4. Evolution of Disease Resistance
Some studies have reported the evolution of disease resistance in soils due to an enrichment of disease-suppressing microbial taxa [197], though the magnitude of this phenomenon may be less effective against soil-borne diseases under the CC systems. Moreover, though some studies have reported the antagonistic effects of some bacterial (e.g., Pseudomonas, Bacillus) and fungal taxa (e.g., Trichoderma) against soil-borne pathogens, results from most studies are based on statistical correlations. For instance, Dong et al. (2016) suggested that the negative correlation of some microbial taxa such as Thermogemmatisporaceae, Actinosynnemataceae, Hydnodontaceae, Herpotrichiellaceae, and Coniosporium with soil-borne fungal pathogen and plant death rate might predict their disease-suppressing effects [176]. Similarly, in another study, the soil disease suppression was correlated with several bacterial phyla, such as Acidobacteria, Nitrospira, Chloroflexi, Planctomycetes, Alphaproteobacteria (notably Azospirillum), and Firmicutes (Thermoanaerobacter) after 10 years of sorghum CC [198]. Similarly, the enrichment of beneficial fungal taxa (Penicillium sp.) in diseased soils is also reported in some studies [176,199]. However, some studies have also reported the enrichment of beneficial bacterial and fungal taxa in the soils infected with nematodes under some crops, for instance, soybean [200]. Nevertheless, the evolution of disease resistance under monoculture cropping does not mean that it can overcome the soil-borne disease with time. However, some studies suggest that the adoption of conservation practices may also suppress soil-borne pathogens and improve soil disease suppression under CC [198,201]. Nevertheless, we need to discern the molecular and ecological mechanisms or processes that determine the emergence and evolution of disease-suppressive soils to sustain soil health under agricultural intensifications.
4. Abiotic Indicators of Soil Health
The abiotic indicators of soil health include but are not limited to soil aggregation, aggregate stability, organic C and organic matter contents, nutrient cycling and sequestration, the composition of soil exudates and metabolites, nutrient balance, and other essential properties such as pH and CEC. These properties are not only linked with each other and regulate soil biodiversity, but these may also influence other important physicochemical processes such as aeration, infiltration, gaseous exchange, soil bulk density, and strength, etc. The CC could potentially alter or disturb these properties, as discussed below.
4.1. Soil Aggregation and Structure
The soil’s physical health is determined by soil structure, which is defined as the aggregation of soil minerals and particles into both large and macro-aggregate-size classes. The soil aggregation and aggregate stability regulate several physical and chemical processes, such as soil compaction, pore geometry, nutrient cycling, water, air infiltration, erosion, drainage, nutrient leaching, root penetration, strength, biological activities, and crop productivity [202,203,204]. Therefore, soil aggregate stability is used as an important physical indicator of soil structure and health [205,206]. However, CC may lead to soil degradation by dismantling the soil structure. For instance, the continuous soybean cropping reduced the soil aggregate stability [207]. A recent study reported the impact of different cropping systems such as continuous corn (CC), soybean-corn rotation (SC), corn-soybean rotation (CS), fallow corn (FC), and fallow soybean (FS) on soil aggregates stability [208]. Their results showed that the CS and FS treatments significantly enhanced the mean weight diameter (MWD) and fractal dimension (D). These treatments also enhanced MWD and geometric mean diameter (GMD) in the water-stable aggregates (WSAs), as compared to the CC treatment. Moreover, rotation treatments also increased the water-stable aggregates stability rate (WSAR), in addition to reducing the aggregates destruction (PAD) more than the CC treatment. Furthermore, rotation rather than CC treatments enhanced the percentage of macro-aggregates [208], thus suggesting that CC may negatively affect soil aggregation, aggregate stability, and the composition of aggregate-size classes. However, the impact of CC on soil structure may differ among crops. For Instance, Acikgoz et al. (2017) studied the impact of 125 years of CC of corn, wheat, timothy, and a rotation of corn–wheat–red clover on soil physical properties. They collected intact soil cores from soils under continuous corn, continuous wheat, continuous timothy, and a rotation of corn–wheat–red clover cropping systems. Among these treatments, CC of timothy rather than wheat and corn showed three- to four-times better soil aggregate stability [209]. The CC of corn showed the lowest aggregate stability, soil strength, and highest bulk density [209], thus suggesting that CC may affect soil structure and strength. In another study, the CC of corn reduced and increased the proportion of macro-aggregates (>0.25 mm) and micro-aggregates (<0.25 mm), respectively, as compared to the initial composition of soil particles or that of fallow soil [210]. Similarly, several studies have reported the adverse effects of horticultural plants, such as tea plantation on soil structure [211,212]. Not only the CC but also the conversion of land from native vegetation to monoculture cropping could reduce soil aggregation [213]. In general, the CC may deteriorate soil aggregates, thus leading to a net release of C and nutrients from agricultural soils [214,215,216]. The differential effects of different CC systems on soil aggregation and structure may also emerge from differences in crop residue inputs to the soil ecosystems [207]. Soils under monoculture rather than diverse cropping systems may receive a lower level of crop residues, which in return may lead to less soil organic matter production and particle aggregation. Other than soil aggregate stability, more research is needed to elucidate the impact of CC on the composition of different soil aggregate-size classes regarding their relative significance in soil health and plant growth, as influenced by the crop identity.
4.2. Soil Organic Matter and Organic C Contents
The soil organic matter and C contents are the key drivers and indicators of soil health, as they play a central role in soil fertility, functioning, quality, and sustainability [217,218,219]. There is an emerging consensus that CC with destructive tillage practices may negatively affect soil organic matter in the agricultural soils [207]. For instance, Miao et al. (2017) showed that fallow soil showed relatively higher contents of soil organic C than the soil under a continuous corn cropping [210]. In another study, Qiao et al. (2018) studied the effect of maize-soybean rotation, continuous soybean, and continuous maize cropping on soil organic C contents and its chemical composition. They reported that a continuous rather than rotation cropping system altered the relative composition of two main classes of soil organic C, such as O-alkyl and aromatic C, across the soil depth profiles [220]. In another study, Börjesson et al. (2018) studied the impact of ley-dominated rotation and the cereal monoculture cropping systems on soil organic C contents. They found a significant increase in the soil organic C contents under ley-dominated rotation than under cereal monoculture cropping systems [221]. The relative impact of continuous monoculture cropping on soil organic contents may also vary depending on soil and plant types. For example, Fugen et al. (2016) showed that the soil under CC of sorghum (15%) and corn (11%) showed relatively higher soil C contents than soil under continuous cotton [141]. Similarly, in another study, a reduction in soil organic C contents was observed under continuous cotton [222]. The reduced C contents under cotton rather than other crops might be attributed to the nature of cotton crop residues and plant-derived toxic metabolites, which could limit the breakdown of the crop litter, thus limiting the amount of soil organic C in general. Similarly, a decrease in soil organic C contents under CC of sesame is also reported [142]. Understanding the impacts of CC on soil organic matter and C are difficult to comprehend at some level because of opposing results obtained from different soil and crop systems. Nevertheless, the soil organic matter and C contents may sustain or increase under the CC systems that follow conservational approaches (e.g., reduced tillage, manure input) [223,224]. Therefore, some soil and crop factors, such as tillage intensity, fertilizer use, crop residue input, crop root system, plant-derived metabolites, and soil biodiversity, could potentially determine the impact of CC on soil organic matter contents [225,226].
4.3. Mineralization and Cycling of C and N
The mineralization of soil organic C and N is an essential indicator of soil fertility and agroecosystem functioning. The impact of CC on either soil C or N mineralization may also depend on soil texture (sandy vs. clay soil), structure, amount of crop residues and fertilizer use, etc. For instance, the clay rather than sandy soil may have a more binding capacity for organic and inorganic compounds, which may limit mineralization. Moreover, an unpredictable nature of nutrient mineralization may also cause growers to overuse N fertilizers, which may lead to alteration in N mineralization and losses in soils under CC [227,228,229,230]. For instance, under a continuous corn cropping, the use of NH+4-N fertilizer suppressed the mineralization (i.e., 13–21% reduction in gross ammonification) of organic matter [231], thus liming nutrient cycling and affecting soil fertility. Moreover, a wider C/N ratio of some crop residues, such as corn rather than soybean, may also explain variations in C and N mineralization under the CC systems [232,233]. In a comprehensive study [229], authors compared the N mineralization at various N fertilizer rates in soil under continuous corn and corn-soybean cropping systems. In unfertilized soils of continuous corn rather than soybean-corn cropping, they observed a reduced N mineralization, while several other studies have also reported a reduced N mineralization under continuous corn than corn-soybean rotation cropping [233,234]. Similarly, in another study, fertilized rather than unfertilized control soil showed an increase in the C and N mineralization under a continuous corn cropping system [235]. Meanwhile, it is also anticipated that the soil under legume crops such as soybean may show increased nutrient contents and C mineralization due to more N and greater microbial activities [229,236,237]. Moreover, an increased coverage of corn rather than soybean residues and consequently reduced soil temperature may also affect nutrient mineralization [229]. Similarly, CC of sesame also showed a reduced soil total C and N content [142]. Regarding the comparison of nutrient mineralization among various CC systems, Dou et al. (2016) showed increased contents of soil C and N under continuous sorghum and corn rather than under cotton, which nevertheless suggests a greater breakdown and processing of crop residues and nutrient mineralization in these systems [222]. In addition to affecting nutrient cycling in the soil ecosystem, CC systems have relatively less capability to sequester nutrients, and thus more nutrient losses mainly through leaching or surface run are expected [238]. Moreover, continuous monoculture cropping systems may produce relatively less diverse crop residues, which may not only affect nutrient cycling but also other biological and physical components of soil ecosystems [239] that regulate nutrient cycling and mineralization. Therefore, an in-depth understanding of the impact of CC on nutrient cycling is important to predict its consequence on soil health and productivity.
4.4. Soil Physicochemical Properties and Nutrient Deficiencies
The soil physicochemical properties include but are not limited to: soil pH, moisture, nutrient contents and their ratios, water holding capacity, conductivity, cation exchange capacity, bulk density, compaction, strength, microclimate and temperature, saturation percentage, acidity, and alkalinity, porosity, buffering capacity, etc. The CC could potentially affect several soil physicochemical properties, particularly soil pH and levels of essential macro-and micro-nutrients. Among soil properties, the important ones are soil pH and CEC, which determine the soil biochemistry, biology, and plant growth. Nevertheless, the impact of CC on soil physicochemical properties may also depend, among others, on soil conditions and climatic factors. For instance, CC may increase soil pH by increasing salinity, alkalinity, and sodicity in semi-arid and arid agroecosystems, and correspondingly could affect soil nutrient availability to plants. Similarly, it may reduce soil pH and increase soil acidity in relatively acidic soils. At present, the impact of CC on soil physicochemical properties and nutrient availability is mostly tested in relatively acidic soils, while most studies have reported a reduction in soil pH and an increase in soil acidity under CC of several crops such as black pepper [240], tobacco, [241], tea [242], wheat [243], potato [244], corn [245] and soybean [246], including several other crops. A reduction in soil pH under CC is mostly attributed to the use of nitrogenous fertilizers, tillage intensity, biodegradation of crop residues, and resultantly increased production of organic acids in the soil. In addition to negatively affecting soil organisms as mentioned above, the soil acidity may also negatively affect the availability of essential macro- and micro-nutrients, such as N, P, K, calcium (Ca), magnesium (Mg), and Mo (molybdenum) [247]. It may also cause the toxicity of some heavy metals, for instance, Al or Mn, to plants [248]. Therefore, the negative impacts of soil acidity on yields of agricultural crops are also reported in several studies [207,249,250]. Moreover, the CC could also mine soil resources, thus leading to the depletion of essential nutrients and/or causing an imbalance in soil nutrient contents. For instance, the continuous corn showed relatively less soil K contents than the soils under crop rotation systems at three northern corn/soybean belt locations in the United States [207]. Jiao et al. (2019) reported that soil under American ginseng showed relatively higher micronutrients such as manganese (Mn), copper (Cu), and iron (Fe) being 113%, 99%, and 80% higher than in the soil of continuously cultivated maize crop [251]. Moreover, the CC under certain standard practices may create obnoxious conditions under which plants may not take up sufficient nutrients from the soil, which may also lead to reduced plant growth and crop yield [84,252]. Though readers can find several examples of soil nutrient deficiencies under CC, these kinds of cropping practices may also alter CEC and salt levels in soil, which may also lead to soil degradation [253]. Moreover, the CC may also increase soil bulk density and compaction, thus reducing the infiltration of water, nutrient movement, and aeration, which are important indicators of soil physical health [207,253]. Finally, among other changes, the CC may also alter a multitude of changes that determine the soil physicochemical environment, and with time, it may also lead to the accumulation of toxic substances and leaching of nutrients that may impair soil and water quality [212,254] in the agricultural neighborhoods.
4.5. Accumulation of Plant- and Microbe-Derived Exudates in Soil
The plant-derived exudates, metabolites, secondary metabolites, and other chemical compounds shape biotic and abiotic indicators of soil health and thus are an important predictor of soil- and root-associated microbial communities [255,256]. These metabolites accumulate in the soil through plant roots, flowers, residue incorporation, and biodegradation during and after crop harvesting [257,258]. The continuous monoculture cropping could reduce the composition and diversity of soil metabolites because monoculture rather than diverse cropping systems may produce and release less diverse metabolites into the soil. For instance, long-term CC of soybean may add similar types of metabolites with time, such as phenolic acids [259,260]. Similarly, the production of vanillic acid in soybean root may also affect the biotic component of soil [261,262]. Similarly, the accumulation of phenolic acids, such as cinnamic acids and p-hydroxybenzoic, under strawberry monoculture cropping is considered to cause autotoxicity in soil [18,263]. Likewise, long-term cropping of cotton plants may also increase the abundance of toxic compounds such as gossypol [264]. The cucumber roots also release a chemical metabolite named coumarin, while its accumulation under monoculture cropping may also affect soil health [265]. Given these examples, it is very likely that the accumulation of consistently similar types of plant- or microbe-derived metabolites such as alcohols, fatty acids, ketones, phenolic acids, aldehydes, and other organic and inorganic compounds could potentially lead to toxic conditions or auto-toxicity in agricultural soils. Therefore, auto-toxic conditions along with reduced soil pH, imbalanced nutrient contents and CEC, nutrient mining, and altered soil physical properties may lead to soil degradation and cropping penalties.
5. Conclusions: Alteration in Soil Physicochemical and Food Web Interactions
The soil ecosystems are characterized by physicochemical and biological components, while interactions between them determine the structure, complexity, and functioning of soil food webs. There is an established consensus that soil food web interactions regulate the productivity of agroecosystems. From the above-mentioned review, we suggest that CC may adversely affect the abiotic and biotic indicators of soil health. Regarding physical components, the CC may lead to degradation of soil physicochemical properties, such as the reduction in soil organic C, essential nutrients, aggregation, and pH, in addition to the accumulation of biochemically less diverse or similar types of metabolites, etc. It could also cause environmental penalties by increasing the surface runoff and leaching of nutrients, thus contaminating the surface and groundwaters. Consequently, the degraded soils with a reduced pH may alter the soil food structure or microbial interactions by promoting the abundance of soil fungi rather than bacteria and these effects are also translated to higher trophic-level organisms, for instance, increasing the abundance of fungal than bacterial feeders (nematodes). Overall, a reduction in beneficial microbial taxa and an enrichment of pathogenic microbes not only negatively impact crop production but also dismantle the beneficial soil food web and microbial loop interactions, which are essential for soil health and sustainability. Meanwhile, it is anticipated that breaking the continuity of monoculture cropping by introducing alternative cash or cover crops may restore soil physicochemical and biological properties, and thus might help in sustainable cropping in the long run. However, where necessary, the conservation or organic cropping practices may help in sustaining soil health under CC, though their cost may increase with time.
Finally, we summarized the adverse effects of CC on soil health indicators, in addition to predicting the positive impacts of conservation cropping practices such as inter, mixture, and alternative cropping with the inclusion of cover crops on soil health (Figure 2). As biodiversity and ecosystem function theory suggests that an increase in species diversity may increase ecosystem functioning [266,267], we conclude that enhancing cropping diversity may sustain or improve the health and fertility of soil ecosystems, which are essential for the provision of agroecosystem services.
Author Contributions
Conceptualization, Z.H.P. and M.S.; methodology, M.S.; software, Z.H.P.; validation, J.I., Q.Z., D.C. and H.W.; formal analysis, Z.H.P.; investigation, M.S.; resources, M.S.; data curation, Z.H.P.; writing—original draft preparation, Z.H.P.; writing—review and editing, M.S.; J.I., Q.Z., D.C. and H.W.; visualization, M.S.; supervision, M.S.; project administration, Z.H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amundson, R.; Berhe, A.A.; Hopmans, J.W.; Olson, C.; Sztein, A.E.; Sparks, D.L. Soil and human security in the 21st century. Science 2015, 348, 1261071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montanarella, L.; Pennock, D.J.; McKenzie, N.; Badraou, M.; Chude, V.; Baptista, I.; Mamo, T.; Yemefack, M.; Aulakh, M.S.; Yagi, K.; et al. World’s soils are under threat. Soil 2016. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.E.; Lemerle, D. Continuous cropping systems and weed selection. Euphytica 2006, 148, 61–73. [Google Scholar] [CrossRef]
- Plourde, J.D.; Pijanowski, B.C.; Pekin, B.K. Evidence for increased monoculture cropping in the Central United States. Agric. Ecosyst. Environ. 2013, 165, 50–59. [Google Scholar] [CrossRef]
- Shipton, P.J. Monoculture and soilborne plant pathogens. Monocult. Soilborne Plant Pathog. 1977, 15, 387–407. [Google Scholar] [CrossRef]
- Song, J.; Li, S.; Xu, Y.; Wei, W.; Yao, Q.; Pan, F. Diversity of parasitic fungi from soybean cyst nematode associated with long-term continuous cropping of soybean in black soil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 66, 432–442. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, H.; Bai, C.; Wu, D.; Song, Q.; Gao, D.; Dong, Z.; Cheng, X.; Dong, Q.; Zhang, Y.; et al. Effects of different mechanized soil fertilization methods on corn soil fertility under continuous cropping. IOP Conf. Ser. Earth Environ. Sci. 2017, 64, 012109. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Li, Z.-Z.; Zhao, Q.; Huang, X.-Y.; Huang, K.-F. Effect of continuous cropping on the rhizosphere soil and growth of common buckwheat. Plant Prod. Sci. 2020, 23, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Wang, Y.; Liu, Q.; Zhang, Y.; Li, X.; Li, H.; Li, W. Phase changes of continuous cropping obstacles in strawberry (Fragaria × ananassa Duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Chen, P.; Wang, Y.; Liu, Q.; Li, W.; Li, H.; Li, X.; Zhang, Y. Transcriptomic analysis reveals recovery strategies in strawberry roots after using a soil amendment in continuous cropping soil. BMC Plant Biol. 2020, 20, 5. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Qi, G.; Luo, T.; Zhang, H.; Jiang, Q.; Wang, R.; Zhao, X. Continuous-cropping tobacco caused variance of chemical properties and structure of bacterial network in soils. Land Degrad. Dev. 2018, 29, 4106–4120. [Google Scholar] [CrossRef]
- Treder, K.; Jastrzębska, M.; Kostrzewska, M.K.; Makowski, P. Do Long-Term Continuous Cropping and Pesticides Affect Earthworm Communities? Agronomy 2020, 10, 586. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Liu, T.; Wang, H.; Yang, Y.; Chen, X.; Zhu, S. Analysis of bacterial and fungal communities in continuous-cropping ramie (Boehmeria nivea L. Gaud) fields in different areas in China. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Li, T.; Zhao, D.; Liao, Y. Tillage practices with different soil disturbance shape the rhizosphere bacterial community throughout crop growth. Soil Tillage Res. 2020, 197, 104501. [Google Scholar] [CrossRef]
- Yang, J.; Ruegger, P.M.; McKenry, M.V.; Becker, J.O.; Borneman, J. Correlations between Root-Associated Microorganisms and Peach Replant Disease Symptoms in a California Soil. PLoS ONE 2012, 7, e46420. [Google Scholar] [CrossRef]
- Seifert, C.A.; Roberts, M.J.; Lobell, D.B. Continuous Corn and Soybean Yield Penalties across Hundreds of Thousands of Fields. Agron. J. 2017, 109, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.-L.; Jiao, X.-Y.; Fan, F.-F.; Wang, J.-S.; Guo, J.; Dong, E.-W.; Wang, L.-G.; Shen, X.-M. Effect of continuous sorghum cropping on the rhizosphere microbial community and the role of Bacillus amyloliquefaciens in altering the microbial composition. Plant Growth Regul 2019, 89, 299–308. [Google Scholar] [CrossRef]
- Li, X.; Lewis, E.E.; Liu, Q.; Li, H.; Bai, C.; Wang, Y. Effects of long-term continuous cropping on soil nematode community and soil condition associated with replant problem in strawberry habitat. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Chen, D.; Xing, W.; Lan, Z.; Saleem, M.; Wu, Y.; Hu, S.; Bai, Y. Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland. Funct. Ecol. 2019, 33, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, K.; Schindelbeck, R.; Kurtz, K.S.M.; van Es, H. Soil health assessment. In Soil Analysis: Recent Trends and Applications; Rakshit, A., Ghosh, S., Chakraborty, S., Philip, V., Datta, A., Eds.; Springer: Singapore, 2020; pp. 199–219. ISBN 978-981-15-2039-6. [Google Scholar]
- Lal, R. Soil health and carbon management. Food Energy Secur. 2016, 5, 212–222. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, M.D.; Grandy, A.S. Soil microbial biomass and function are altered by 12 years of crop rotation. Soil 2016, 2, 583–599. [Google Scholar] [CrossRef] [Green Version]
- Nunes, M.R.; Vas Es, H.M.; Schindelbeck, R.; Ristow, A.J.; Ryan, M. No-till and cropping system diversification improve soil health and crop yield. Geoderma 2018, 328, 30–43. [Google Scholar] [CrossRef]
- Saleem, M.; Hu, J.; Jousset, A. More Than the Sum of Its Parts: Microbiome Biodiversity as a Driver of Plant Growth and Soil Health. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 145–168. [Google Scholar] [CrossRef]
- Saleem, M. Microbiome Community Ecology: Fundamentals and Applications; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 978-3-319-11665-5. [Google Scholar]
- Weisberger, D.; Nichols, V.; Liebman, M. Does diversifying crop rotations suppress weeds? A meta-analysis. PLoS ONE 2019, 14, e0219847. [Google Scholar] [CrossRef] [Green Version]
- Abouziena, H.F.; Haggag, W.M.; Abouziena, H.F.; Haggag, W.M. Weed Control in Clean Agriculture: A Review 1. Planta Daninha 2016, 34, 377–392. [Google Scholar] [CrossRef]
- Titi, A.E. Soil Tillage in Agroecosystems; CRC Press: Boca Raton, FL, USA, 2002; ISBN 978-1-4200-4060-9. [Google Scholar]
- Hamblin, A.P. The effect of tillage on soil surface properties and the water balance of a xeralfic alfisol. Soil Tillage Res. 1984, 4, 543–559. [Google Scholar] [CrossRef]
- Ward, N.K.; Maureira, F.; Stöckle, C.O.; Brooks, E.S.; Painter, K.M.; Yourek, M.A.; Gasch, C.K. Simulating field-scale variability and precision management with a 3D hydrologic cropping systems model. Precis. Agric. 2018, 19, 293–313. [Google Scholar] [CrossRef] [Green Version]
- Kahimba, F.C.; Sri Ranjan, R.; Froese, J.; Entz, M.; Nason, R. Cover Crop Effects on Infiltration, Soil Temperature, and Soil Moisture Distribution in the Canadian Prairies. Appl. Eng. Agric. 2008, 24, 321–333. [Google Scholar] [CrossRef]
- Dexter, A.R. Soil physical quality: Part, I. Theory, effects of soil texture, density, and organic matter, and effects on root growth. Geoderma 2004, 120, 201–214. [Google Scholar] [CrossRef]
- Khademalrasoul, A.; Naveed, M.; Heckrath, G.; Kumari, K.G.I.D.; de Jonge, L.W.; Elsgaard, L.; Vogel, H.-J.; Iversen, B.V. Biochar Effects on Soil Aggregate Properties Under No-Till Maize. Soil Sci. 2014, 179, 273–283. [Google Scholar] [CrossRef]
- Lal, R. Soil quality changes under continuous cropping for seventeen seasons of an alfisol in western Nigeria. Land Degrad. Dev. 1998, 9, 259–274. [Google Scholar] [CrossRef]
- Hati, K.M.; Swarup, A.; Singh, D.; Misra, A.K.; Ghosh, P.K. Long-term continuous cropping, fertilisation, and manuring effects on physical properties and organic carbon content of a sandy loam soil. Soil Res. 2006, 44, 487–495. [Google Scholar] [CrossRef]
- Rieu, M.; Sposito, G. Fractal Fragmentation, Soil Porosity, and Soil Water Properties: I. Theory. Soil Sci. Soc. Am. J. 1991, 55, 1231–1238. [Google Scholar] [CrossRef] [Green Version]
- Głąb, T.; Kulig, B. Effect of mulch and tillage system on soil porosity under wheat (Triticum aestivum). Soil Tillage Res. 2008, 99, 169–178. [Google Scholar] [CrossRef]
- Srinivasarao, C.; Kundu, S.; Shanker, A.K.; Naik, R.P.; Vanaja, M.; Venkanna, K.; Maruthi Sankar, G.R.; Rao, V.U.M. Continuous cropping under elevated CO2: Differential effects on C4 and C3 crops, soil properties and carbon dynamics in semi-arid alfisols. Agric. Ecosyst. Environ. 2016, 218, 73–86. [Google Scholar] [CrossRef]
- Hansen, V.; Hauggaard-Nielsen, H.; Petersen, C.T.; Mikkelsen, T.N.; Müller-Stöver, D. Effects of gasification biochar on plant-available water capacity and plant growth in two contrasting soil types. Soil Tillage Res. 2016, 161, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chalise, K.S.; Singh, S.; Wegner, B.R.; Kumar, S.; Pérez-Gutiérrez, J.D.; Osborne, S.L.; Nleya, T.; Guzman, J.; Rohila, J.S. Cover Crops and Returning Residue Impact on Soil Organic Carbon, Bulk Density, Penetration Resistance, Water Retention, Infiltration, and Soybean Yield. Agron. J. 2019, 111, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Nouri, A.; Lee, J.; Yin, X.; Saxton, A.M.; Tyler, D.D.; Sykes, V.R.; Arelli, P. Crop species in no-tilled summer cropping rotation affect soil quality and yield in Alfisols, southeastern USA. Geoderma 2019, 345, 51–62. [Google Scholar] [CrossRef]
- Feng, G.; Sharratt, B.; Young, F. Influence of long-term tillage and crop rotations on soil hydraulic properties in the US Pacific Northwest. J. Soil Water Conserv. 2011, 66, 233–241. [Google Scholar] [CrossRef]
- Anwar, U.; Schulte, L.A.; Helmers, M.; Kolka, R.K. The Effect of Five Biomass Cropping Systems on Soil-Saturated Hydraulic Conductivity Across a Topographic Gradient. Bioenerg. Res. 2017, 10, 824–831. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, X.; Gregorich, E.G.; McLaughlin, N.B.; Zhang, X.; Guo, Y.; Liang, A.; Fan, R.; Sun, B. No-tillage with continuous maize cropping enhances soil aggregation and organic carbon storage in Northeast China. Geoderma 2018, 330, 204–211. [Google Scholar] [CrossRef]
- Stirzaker, R.J.; Passioura, J.B.; Wilms, Y. Soil structure and plant growth: Impact of bulk density and biopores. Plant Soil 1996, 185, 151–162. [Google Scholar] [CrossRef]
- Saleem, M.; Pervaiz, Z.H.; Contreras, J.; Lindenberger, J.H.; Hupp, B.M.; Chen, D.; Zhang, Q.; Wang, C.; Iqbal, J.; Twigg, P. Cover crop diversity improves multiple soil properties via altering root architectural traits. Rhizosphere 2020, 100248. [Google Scholar] [CrossRef]
- Ryu, J.; Na, J.-S.; Hwang, N.-Y. Study on the Cause of Injury by Continuous Cropping of Peanut. Korean J. Soil Sci. Fertil. 1992, 25, 270–274. [Google Scholar]
- Tsunekawa, A.; Kar, A.; Yanai, J.; Tanaka, U.; Miyazaki, T. Influence of continuous cultivation on the soil properties affecting crop productivity in the Thar Desert, India. J. Arid Environ. 1997, 36, 367–384. [Google Scholar] [CrossRef]
- Van Eerd, L.L.; Congreves, K.A.; Hayes, A.; Verhallen, A.; Hooker, D.C. Long-term tillage and crop rotation effects on soil quality, organic carbon, and total nitrogen. Can. J. Soil Sci. 2014, 94, 303–315. [Google Scholar] [CrossRef]
- Idowu, O.J.; Van Es, H.M.; Abawi, G.S.; Wolfe, D.W.; Schindelbeck, R.R.; Moebius-Clune, B.N.; Gugino, B.K. Use of an integrative soil health test for evaluation of soil management impacts. Renew. Agric. Food Syst. 2009, 24, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.F.; Anderson, R.L. Aggregate stability and glomalin in alternative crop rotations for the central Great Plains. Biol. Fertil. Soils 2000, 31, 249–253. [Google Scholar] [CrossRef]
- Sithole, N.J.; Magwaza, L.S.; Thibaud, G.R. Long-term impact of no-till conservation agriculture and N-fertilizer on soil aggregate stability, infiltration and distribution of C in different size fractions. Soil Tillage Res. 2019, 190, 147–156. [Google Scholar] [CrossRef]
- García, G.V.; Wyngaard, N.; Calvo, N.I.R.; San Martino, S.; Covacevich, F.; Studdert, G.A. Soil survey reveals a positive relationship between aggregate stability and anaerobically mineralizable nitrogen. Ecol. Indic. 2020, 117, 106640. [Google Scholar] [CrossRef]
- Heyman, H.; Bassuk, N.; Bonhotal, J.; Walter, T. Compost Quality Recommendations for Remediating Urban Soils. Int. J. Environ. Res. Public Health 2019, 16, 3191. [Google Scholar] [CrossRef] [Green Version]
- Declerck, S.; Risede, J.M.; Rufyikiri, G.; Delvaux, B. Effects of arbuscular mycorrhizal fungi on severity of root rot of bananas caused by Cylindrocladium spathiphylli. Plant Pathol. 2002, 51, 109–115. [Google Scholar] [CrossRef]
- Roarty, S.; Hackett, R.A.; Schmidt, O. Earthworm populations in twelve cover crop and weed management combinations. Appl. Soil Ecol. 2017, 114, 142–151. [Google Scholar] [CrossRef]
- Ashworth, A.J.; Allen, F.L.; Tyler, D.D.; Pote, D.H.; Shipitalo, M.J. Earthworm populations are affected from long-term crop sequences and bio-covers under no-tillage. Pedobiologia 2017, 60, 27–33. [Google Scholar] [CrossRef]
- Tian, X.; Zhao, X.; Mao, Z.; Xie, B. Variation and Dynamics of Soil Nematode Communities in Greenhouses with Different Continuous Cropping Periods. Hortic. Plant J. 2020. [Google Scholar] [CrossRef]
- Jordan, D.L.; Corbett, T.; Bogle, C.; Shew, B.; Brandenburg, R.; Ye, W. Effect of Previous Rotation on Plant Parasitic Nematode Population in Peanut and Crop Yield. Crop Forage Turfgrass Manag. 2017, 3. [Google Scholar] [CrossRef]
- Grabau, Z.J.; Vetsch, J.A.; Chen, S. Effects of Fertilizer, Nematicide, and Tillage on Plant–Parasitic Nematodes and Yield in Corn and Soybean. Agron. J. 2017, 109, 1651–1662. [Google Scholar] [CrossRef]
- Mahal, N.K.; Castellano, M.J.; Miguez, F.E. Potentially mineralizable nitrogen: A soil health indicator. Crop. Soils 2019, 52, 8–10. [Google Scholar] [CrossRef]
- Mahal, N.K.; Castellano, M.J.; Miguez, F.E. Conservation Agriculture Practices Increase Potentially Mineralizable Nitrogen: A Meta-Analysis. Soil Sci. Soc. Am. J. 2018, 82, 1270–1278. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Hayakawa, C.; Inagaki, Y.; Kosaki, T. Effects of land use change on turnover and storage of soil organic matter in a tropical forest. Plant Soil 2020, 446, 425–439. [Google Scholar] [CrossRef]
- Stakhurlova, L.D.; Stulin, A.F. Biodynamics of black soils leached under different agrotechnical practices in long-term field experiments. Russ. Agric. Sci. 2017, 43, 35–39. [Google Scholar] [CrossRef]
- Smith, J.D.; Strauss, J.A.; Hardie, A.G. Effects of long-term grazed crop and pasture systems under no-till on organic matter fractions and selected quality parameters of soil in the Overberg, South Africa. S. Afr. J. Plant Soil 2020, 37, 1–10. [Google Scholar] [CrossRef]
- Hua, K.; Wang, D.; Guo, Z. Effects of long-term application of various organic amendments on soil particulate organic matter storage and chemical stabilisation of vertisol soil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 505–514. [Google Scholar] [CrossRef]
- Hazra, K.K.; Nath, C.P.; Singh, U.; Praharaj, C.S.; Kumar, N.; Singh, S.S.; Singh, N.P. Diversification of maize-wheat cropping system with legumes and integrated nutrient management increases soil aggregation and carbon sequestration. Geoderma 2019, 353, 308–319. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhary, T.; Das, S.; Meena, S.K. Weed seed bank: Impacts and management for future crop production. In Agronomic Crops; Volume 2: Management Practices; Hasanuzzaman, M., Ed.; Springer: Singapore, 2019; pp. 207–223. ISBN 978-981-329-783-8. [Google Scholar]
- Pan, J.; Zhang, L.; Wang, L.; Fu, S. Effects of long-term fertilization treatments on the weed seed bank in a wheat-soybean rotation system. Glob. Ecol. Conserv. 2020, 21, e00870. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Liang, B.C.; Drury, C.F.; Mackenzie, A.F.; McGill, W.B. Elucidation of the source and turnover of water soluble and microbial biomass carbon in agricultural soils. Soil Biol. Biochem. 2000, 32, 581–587. [Google Scholar] [CrossRef]
- Li, L.; Wilson, C.B.; He, H.; Zhang, X.; Zhou, F.; Schaeffer, S.M. Physical, biochemical, and microbial controls on amino sugar accumulation in soils under long-term cover cropping and no-tillage farming. Soil Biol. Biochem. 2019, 135, 369–378. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, J.; Saleem, M.; Wang, B.; Hu, S.; Bai, Y.; Pan, Q.; Chen, D. Ecological clusters based on responses of soil microbial phylotypes to precipitation explain ecosystem functions. Soil Biol. Biochem. 2020, 142, 107717. [Google Scholar] [CrossRef]
- Guo, Z.C.; Zhang, Z.B.; Zhou, H.; Rahman, M.T.; Wang, D.Z.; Guo, X.S.; Li, L.J.; Peng, X.H. Long-term animal manure application promoted biological binding agents but not soil aggregation in a Vertisol. Soil Tillage Res. 2018, 180, 232–237. [Google Scholar] [CrossRef]
- Redmile-Gordon, M.; Gregory, A.S.; White, R.P.; Watts, C.W. Soil organic carbon, extracellular polymeric substances (EPS), and soil structural stability as affected by previous and current land-use. Geoderma 2020, 363, 114143. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, J.; Yu, Z.; Yao, Q.; Zhang, W.; Mi, G.; Liang, A.; Li, L.; Chen, X.; Jin, J.; et al. Continuous cropping of soybean induced a more fluctuating fungal network and intensive pathogenic fungal interactions in a Mollisol of Northeast China. Soil Sci. Soc. Am. J. 2020, 84, 775–783. [Google Scholar] [CrossRef]
- Sun, T.; Miao, J.; Saleem, M.; Zhang, H.; Yang, Y.; Zhang, Q. Bacterial compatibility and immobilization with biochar improved tebuconazole degradation, soil microbiome composition and functioning. J. Hazard Mater. 2020, 398, 122941. [Google Scholar] [CrossRef]
- Sun, J.; Zou, L.; Li, W.; Wang, Y.; Xia, Q.; Peng, M.; Sun, J.; Zou, L.; Li, W.; Wang, Y.; et al. Soil microbial and chemical properties influenced by continuous cropping of banana. Sci. Agric. 2018, 75, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Rankoth, L.M.; Udawatta, R.P.; Gantzer, C.J.; Jose, S.; Veum, K.; Dewanto, H.A. Cover Crops on Temporal and Spatial Variations in Soil Microbial Communities by Phospholipid Fatty Acid Profiling. Agron. J. 2019, 111, 1693–1703. [Google Scholar] [CrossRef]
- Finney, D.M.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Sun, T.; Li, M.; Saleem, M.; Zhang, Q.; Wang, C. Soil-applied biochar increases microbial diversity and wheat plant performance under herbicide fomesafen stress. Ecotoxicol. Environ. Saf. 2019, 171, 75–83. [Google Scholar] [CrossRef]
- Zhang, Q.; Saleem, M.; Wang, C. Probiotic strain Stenotrophomonas acidaminiphila BJ1 degrades and reduces chlorothalonil toxicity to soil enzymes, microbial communities and plant roots. AMB Express 2017, 7, 227. [Google Scholar] [CrossRef]
- Chen, W.; Teng, Y.; Li, Z.; Liu, W.; Ren, W.; Luo, Y.; Christie, P. Mechanisms by which organic fertilizer and effective microbes mitigate peanut continuous cropping yield constraints in a red soil of south China. Appl. Soil Ecol. 2018, 128, 23–34. [Google Scholar] [CrossRef]
- Kou, X.; Su, T.; Ma, N.; Li, Q.; Wang, P.; Wu, Z.; Liang, W.; Cheng, W. Soil micro-food web interactions and rhizosphere priming effect. Plant Soil 2018, 432, 129–142. [Google Scholar] [CrossRef]
- Hu, Z.; Zhu, C.; Chen, X.; Bonkowski, M.; Griffiths, B.; Chen, F.; Zhu, J.; Hu, S.; Hu, F.; Liu, M. Responses of rice paddy micro-food webs to elevated CO2 are modulated by nitrogen fertilization and crop cultivars. Soil Biol. Biochem. 2017, 114, 104–113. [Google Scholar] [CrossRef]
- De Vries, F.T.; Wallenstein, M.D. Below-ground connections underlying above-ground food production: A framework for optimising ecological connections in the rhizosphere. J. Ecol. 2017, 105, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Gou, X.; Cai, Y.; Wang, C.; Li, B.; Zhang, Y.; Tang, X.; Shen, J.; Cai, Z. Effects of different long-term cropping systems on phosphorus adsorption and desorption characteristics in red soils. J Soils Sediments 2020, 20, 1371–1382. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, J.; Zeng, S.; Wu, D.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J Soils Sediments 2017, 17, 2230–2238. [Google Scholar] [CrossRef]
- Li, Y.; Fang, F.; Wei, J.; Wu, X.; Cui, R.; Li, G.; Zheng, F.; Tan, D. Humic Acid Fertilizer Improved Soil Properties and Soil Microbial Diversity of Continuous Cropping Peanut: A Three-Year Experiment. Sci. Rep. 2019, 9, 12014. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R. The potassium paradox: Implications for soil fertility, crop production and human health. Renew. Agric. Food Syst. 2014, 29, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Srinivasarao, C.; Kundu, S.; Sharma, K.L.; Reddy, S.; Pharande, A.L.; Vijayasankarbabu, M.; Satish, A.; Singh, R.P.; Singh, S.R.; Chary, G.R.; et al. Magnesium balance in four permanent manurial experiments under rainfed agro-ecosystems of India. Crop Pasture Sci. 2016, 66, 1230–1240. [Google Scholar] [CrossRef]
- Behera, S.K.; Singh, D.; Dwivedi, B.S. Changes in Fractions of Iron, Manganese, Copper, and Zinc in Soil under Continuous Cropping for More Than Three Decades. Commun. Soil Sci. Plant Anal. 2009, 40, 1380–1407. [Google Scholar] [CrossRef]
- Haruna, S.I.; Nkongolo, N.V. Influence of Cover Crop, Tillage, and Crop Rotation Management on Soil Nutrients. Agriculture 2020, 10, 225. [Google Scholar] [CrossRef]
- Wacal, C.; Ogata, N.; Basalirwa, D.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Growth and K Nutrition of Sesame (Sesamum indicum L.) Seedlings as Affected by Balancing Soil Exchangeable Cations Ca, Mg, and K of Continuously Monocropped Soil from Upland Fields Converted Paddy. Agronomy 2019, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Dhaliwal, S.S. Effect of Sewage Sludge and Rice Straw Compost on Yield, Micronutrient Availability and Soil Quality under Rice–Wheat System. Commun. Soil Sci. Plant Anal. 2019, 50, 1943–1954. [Google Scholar] [CrossRef]
- Moreira, A.; Sfredo, G.J.; Moraes, L.A.C.; Fageria, N.K. Lime and Cattle Manure in Soil Fertility and Soybean Grain Yield Cultivated in Tropical Soil. Commun. Soil Sci. Plant Anal. 2015, 46, 1157–1169. [Google Scholar] [CrossRef]
- Kalkhoran, S.S.; Pannell, D.J.; Thamo, T.; White, B.; Polyakov, M. Soil acidity, lime application, nitrogen fertility, and greenhouse gas emissions: Optimizing their joint economic management. Agric. Syst. 2019, 176, 102684. [Google Scholar] [CrossRef]
- Rengel, Z. Handbook of Soil Acidity; CRC Press: Boca Raton, FL, USA, 2003; ISBN 978-0-203-91231-7. [Google Scholar]
- Ghafoor, A.; Qadir, M.; Murtaza, G. Salt-Affected Soils: Principles of Management; Allied Book Centre: Lahore, Pakistan, 2004. [Google Scholar]
- Liu, X.; Song, Q.; Tang, Y.; Li, W.; Xu, J.; Wu, J.; Wang, F.; Brookes, P.C. Human health risk assessment of heavy metals in soil–vegetable system: A multi-medium analysis. Sci. Total Environ. 2013, 463–464, 530–540. [Google Scholar] [CrossRef]
- Karim, Z.; Qureshi, B.A. Health Risk Assessment of Heavy Metals in Urban Soil of Karachi, Pakistan. Hum. Ecol. Risk Assess. Int. J. 2014, 20, 658–667. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef]
- Han, W.; Kemmitt, S.J.; Brookes, P.C. Soil microbial biomass and activity in Chinese tea gardens of varying stand age and productivity. Soil Biol. Biochem. 2007, 39, 1468–1478. [Google Scholar] [CrossRef]
- Liang, Y.; Lin, X.; Yamada, S.; Zhou, M.; Inoue, M.; Inosako, K. Cucumber Productivity and Soil Degradation in Recropping System in Greenhouse. Commun. Soil Sci. Plant Anal. 2012, 43, 1743–1748. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Sarwar, M.; Wong, P.T.W.; Mead, A.; Howe, G.; Newell, M. Field studies on the biofumigation of take-all by Brassica break crops. Aust. J. Agric. Res. 2000, 51, 445–456. [Google Scholar] [CrossRef]
- Meriles, J.M.; Gil, S.V.; Conforto, C.; Figoni, G.; Lovera, E.; March, G.J.; Guzmán, C.A. Soil microbial communities under different soybean cropping systems: Characterization of microbial population dynamics, soil microbial activity, microbial biomass, and fatty acid profiles. Soil Tillage Res. 2009, 103, 271–281. [Google Scholar] [CrossRef]
- Pérez-Brandán, C.; Huidobro, J.; Grümberg, B.; Scandiani, M.M.; Luque, A.G.; Meriles, J.M.; Vargas-Gil, S. Soybean fungal soil-borne diseases: A parameter for measuring the effect of agricultural intensification on soil health. Can. J. Microbiol. 2013, 60, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Katsalirou, E.; Deng, S.; Gerakis, A.; Nofziger, D.L. Long-term management effects on soil P, microbial biomass P, and phosphatase activities in prairie soils. Eur. J. Soil Biol. 2016, 76, 61–69. [Google Scholar] [CrossRef]
- Zhou, X.; Gao, D.; Liu, J.; Qiao, P.; Zhou, X.; Lu, H.; Wu, X.; Liu, D.; Jin, X.; Wu, F. Changes in rhizosphere soil microbial communities in a continuously monocropped cucumber (Cucumis sativus L.) system. Eur. J. Soil Biol. 2014, 60, 1–8. [Google Scholar] [CrossRef]
- Anderson, T.-H.; Domsch, K.H. Carbon assimilation and microbial activity in soil. Z. Pflanz. Bodenkd. 1986, 149, 457–468. [Google Scholar] [CrossRef]
- Insam, H.; Parkinson, D.; Domsch, K.H. Influence of macroclimate on soil microbial biomass. Soil Biol. Biochem. 1989, 21, 211–221. [Google Scholar] [CrossRef]
- Wardle, D.A. A Comparative Assessment of Factors Which Influence Microbial Biomass Carbon and Nitrogen Levels in Soil. Biol. Rev. 1992, 67, 321–358. [Google Scholar] [CrossRef]
- Sahib, M.R.; Pervaiz, Z.H.; Williams, M.A.; Saleem, M.; De Bolt, S. Rhizobacterial species richness improves sorghum growth and soil nutrient synergism in a nutrient-poor greenhouse soil. Sci. Rep. 2020, 13, 41598. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kätterer, T.; Thornton, B.; Börjesson, G. Dynamics of fungal and bacterial groups and their carbon sources during the growing season of maize in a long-term experiment. Biol. Fertil. Soils 2020. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Saleem, M.; Cheng, J.; Mi, J.; Chu, P.; Tuvshintogtokh, I.; Hu, S.; Bai, Y. Effects of aridity on soil microbial communities and functions across soil depths on the Mongolian Plateau. Funct. Ecol. 2019. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, Y.; Xu, X.; Liu, T.; Wu, D.; Zheng, X.; Tang, S.; Dai, Q. Potential use of high-throughput sequencing of soil microbial communities for estimating the adverse effects of continuous cropping on ramie (Boehmeria nivea L. Gaud). PLoS ONE 2018, 13, e0197095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liu, J.; Yu, Z.; Yao, Q.; Li, Y.; Liang, A.; Zhang, W.; Mi, G.; Jin, J.; Liu, X.; et al. Long-term continuous cropping of soybean is comparable to crop rotation in mediating microbial abundance, diversity and community composition. Soil Tillage Res. 2020, 197, 104503. [Google Scholar] [CrossRef]
- Xue, D.; Yao, H.; Huang, C. Microbial Biomass, N Mineralization and Nitrification, Enzyme Activities, and Microbial Community Diversity in Tea Orchard Soils. Plant Soil 2006, 288, 319–331. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Shi, F.Y.; Zhang, R.; Wu, Y.P. Comparison of bacterial diversity in rotational and continuous soybean cropping soils in Heilongjiang. Acta Phytophylacica Sin. 2014, 41, 403–409. [Google Scholar]
- Zhao, J.; Wu, X.; Nie, C.; Wu, T.; Dai, W.; Liu, H.; Yang, R. Analysis of unculturable bacterial communities in tea orchard soils based on nested PCR-DGGE. World J. Microbiol. Biotechnol. 2012, 28, 1967–1979. [Google Scholar] [CrossRef]
- Tang, H.; Xiao, C.; Ma, J.; Yu, M.; Li, Y.; Wang, G.; Zhang, L. Prokaryotic diversity in continuous cropping and rotational cropping soybean soil. FEMS Microbiol. Lett. 2009, 298, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Lay, C.-Y.; Bell, T.H.; Hamel, C.; Harker, K.N.; Mohr, R.; Greer, C.W.; Yergeau, É.; St-Arnaud, M. Canola Root–Associated Microbiomes in the Canadian Prairies. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.-M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Xi, H.; Shen, J.; Qu, Z.; Yang, D.; Liu, S.; Nie, X.; Zhu, L. Effects of Long-term Cotton Continuous Cropping on Soil Microbiome. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, X.; Kong, W.; Wu, Y.; Wang, J. Effect of monoculture soybean on soil microbial community in the Northeast China. Plant Soil 2010, 330, 423–433. [Google Scholar] [CrossRef]
- Li, X.; Ding, C.; Zhang, T.; Wang, X. Fungal pathogen accumulation at the expense of plant-beneficial fungi as a consequence of consecutive peanut monoculturing. Soil Biol. Biochem. 2014, 72, 11–18. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Zhao, J.; Xun, W.; Li, R.; Zhang, R.; Wu, H.; Shen, Q. Different Continuous Cropping Spans Significantly Affect Microbial Community Membership and Structure in a Vanilla-Grown Soil as Revealed by Deep Pyrosequencing. Microb. Ecol. 2015, 70, 209–218. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Diversity and composition of rhizospheric soil and root endogenous bacteria in Panax notoginseng during continuous cropping practices. J. Basic Microbiol. 2017, 57, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Schmidt, T.M.; Coleman, D.C.; Whitman, W.B. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol. Biochem. 2011, 43, 2184–2193. [Google Scholar] [CrossRef]
- Jiang, X.; Wright, A.L.; Wang, J.; Li, Z. Long-term tillage effects on the distribution patterns of microbial biomass and activities within soil aggregates. Catena 2011, 87, 276–280. [Google Scholar] [CrossRef]
- Venter, Z.S.; Jacobs, K.; Hawkins, H.-J. The impact of crop rotation on soil microbial diversity: A meta-analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Schloter, M.; Dilly, O.; Munch, J.C. Indicators for evaluating soil quality. Agric. Ecosyst. Environ. 2003, 98, 255–262. [Google Scholar] [CrossRef]
- Hussain, S.; Siddique, T.; Saleem, M.; Arshad, M.; Khalid, A. Impact of Pesticides on Soil Microbial Diversity, Enzymes, and Biochemical Reactions; Academic Press: Cambridge, MA, USA, 2009; Chapter 5; Volume 102, pp. 159–200. ISBN 0065-2113. [Google Scholar]
- Wang, S.; Ren, J.; Huang, T.; Qiao, G.; Hu, H.; Li, W.; Wu, F.; Pan, K. Evaluation of soil enzyme activities and microbial communities in tomato continuous cropping soil treated with Jerusalem artichoke residues. Commun. Soil Sci. Plant Anal. 2018, 49, 2727–2740. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, X.; Wang, X.; Shao, H.; Yang, J.; Wang, X. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Whitaker, J.; Ostle, N.; Bardgett, R.D.; McNamara, N.P.; Salinas, N.; Meir, P. Temperature sensitivity of soil enzymes along an elevation gradient in the Peruvian Andes. Biogeochemistry 2016, 127, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Martínez, V.; Burow, G.; Zobeck, T.M.; Allen, V.G. Soil Microbial Communities and Function in Alternative Systems to Continuous Cotton. Soil Sci. Soc. Am. J. 2010, 74, 1181–1192. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Yeboah, S.; Cao, L.; Zhang, J.; Shi, S.; Liu, Y. Breaking continuous potato cropping with legumes improves soil microbial communities, enzyme activities and tuber yield. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Dou, F.; Wright, A.L.; Mylavarapu, R.S.; Jiang, X.; Matocha, J.E. Soil Enzyme Activities and Organic Matter Composition Affected by 26 Years of Continuous Cropping. Pedosphere 2016, 26, 618–625. [Google Scholar] [CrossRef]
- Wacal, C.; Ogata, N.; Basalirwa, D.; Sasagawa, D.; Ishigaki, T.; Handa, T.; Kato, M.; Tenywa, M.M.; Masunaga, T.; Yamamoto, S.; et al. Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy. Agronomy 2019, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Bünemann, E.K.; Schwenke, G.D.; Zwieten, L.V. Impact of agricultural inputs on soil organisms—A review. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef] [Green Version]
- Lata, J.-C.; Barot, S.; Leloup, J.; Lerch, T.; Nunan, N.; Raynaud, X. Biodiversity and ecological functioning of soils. In Soils as a Key Component of the Critical Zone 6; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 39–56. ISBN 978-1-119-43827-4. [Google Scholar]
- Ney, L.; Franklin, D.; Mahmud, K.; Cabrera, M.; Hancock, D.; Habteselassie, M.; Newcomer, Q.; Dahal, S.; Subedi, A. Sensitivity of Nematode Community Analysis to Agricultural Management Practices and Inoculation with Local Effective Microorganisms in the Southeastern United States. Soil Syst. 2019, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Pulavarty, A.; Horgan, K.; Kakouli-Duarte, T. Effect of an Alltech soil health product on entomopathogenic nematodes, root-knot nematodes and on the growth of tomato plants in the greenhouse. J. Nematol. 2020, 52, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, W.; Wang, K.; Song, T.; Du, H. Responses of the soil nematode community to management of hybrid napiergrass: The trade-off between positive and negative effects. Appl. Soil Ecol. 2014, 75, 134–144. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, Q.Z.; Wang, Y.Z.; Sun, H.Y.; Bai, C.Q.; Lewis, E.E. Different changes of soil nematode communities in replant and continuous-planting peach orchards and their indicative value for peach replant problem. Helminthologia 2015, 52, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zhao, X.; Mao, Z.; Xie, B. Dynamics of Soil Nematode Communities Revealed Significant Variation in Greenhouse with Different Continuous Cropping Years. bioRxiv 2019, 593541. [Google Scholar] [CrossRef] [Green Version]
- Amador, J.A.; Görres, J.H. Microbiological characterization of the structures built by earthworms and ants in an agricultural field. Soil Biol. Biochem. 2007, 39, 2070–2077. [Google Scholar] [CrossRef]
- Hickman, Z.A.; Reid, B.J. Increased microbial catabolic activity in diesel contaminated soil following addition of earthworms (Dendrobaena veneta) and compost. Soil Biol. Biochem. 2008, 40, 2970–2976. [Google Scholar] [CrossRef]
- Edwards, C.A. Earthworm ecology in cultivated soils. In Earthworm Ecology: From Darwin to Vermiculture; Satchell, J.E., Ed.; Springer: Dordrecht, The Netherlands, 1983; pp. 123–137. ISBN 978-94-009-5965-1. [Google Scholar]
- Curry, J.P.; Byrne, D. The role of earthworms in straw decomposition and nitrogen turnover in arable land in ireland. Soil Biol. Biochem. 1992, 24, 1409–1412. [Google Scholar] [CrossRef]
- Crittenden, S.J.; Eswaramurthy, T.; De Goede, R.G.M.; Brussaard, L.; Pulleman, M.M. Effect of tillage on earthworms over short- and medium-term in conventional and organic farming. Appl. Soil Ecol. 2014, 83, 140–148. [Google Scholar] [CrossRef]
- Lavelle, P. Earthworm activities and the soil system. Biol. Fert. Soils 1988, 6, 237–251. [Google Scholar] [CrossRef]
- Fraser, P.M.; Haynes, R.J.; Williams, P.H. Effects of pasture improvement and intensive cultivation on microbial biomass, enzyme activities, and composition and size of earthworm populations. Biol. Fertil. Soils 1994, 17, 185–190. [Google Scholar] [CrossRef]
- Spurgeon, D.J.; Hopkin, S.P. Risk assessment of the threat of secondary poisoning by metals to predators of earthworms in the vicinity of a primary smelting works. Sci. Total Environ. 1996, 187, 167–183. [Google Scholar] [CrossRef]
- Chen, J.; Saleem, M.; Wang, C.; Liang, W.; Zhang, Q. Individual and combined effects of herbicide tribenuron-methyl and fungicide tebuconazole on soil earthworm Eisenia fetida. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Zhang, Q.; Saleem, M.; Wang, C. Effects of biochar on the earthworm (Eisenia foetida) in soil contaminated with and/or without pesticide mesotrione. Sci. Total Environ. 2019, 671, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Churchfield, S.; Hollier, J.; Brown, V.K. The Effects of Small Mammal Predators on Grassland Invertebrates, Investigated by Field Exclosure Experiment. Oikos 1991, 60, 283–290. [Google Scholar] [CrossRef]
- Emmerling, C. Response of earthworm communities to different types of soil tillage. Appl. Soil Ecol. 2001, 17, 91–96. [Google Scholar] [CrossRef]
- Henneron, L.; Bernard, L.; Hedde, M.; Pelosi, C.; Villenave, C.; Chenu, C.; Bertrand, M.; Girardin, C.; Blanchart, E. Fourteen years of evidence for positive effects of conservation agriculture and organic farming on soil life. Agron. Sustain. Dev. 2015, 35, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef] [Green Version]
- Bilalis, D.; Sidiras, N.; Vavoulidou, E.; Konstantas, A. Earthworm populations as affected by crop practices on clay loam soil in a Mediterranean climate. Acta Agric. Scand. Sect. B Soil Plant Sci. 2009, 59, 440–446. [Google Scholar] [CrossRef]
- Schmidt, O.; Curry, J.P. Population dynamics of earthworms (Lumbricidae) and their role in nitrogen turnover in wheat and wheatclover cropping systems. Pedobiologia 2001, 45, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Feledyn-Szewczyk, B.; Radzikowski, P.; Stalenga, J.; Matyka, M. Comparison of the Effect of Perennial Energy Crops and Arable Crops on Earthworm Populations. Agronomy 2019, 9, 675. [Google Scholar] [CrossRef] [Green Version]
- Gulvik, M. Mites (Acari) as indicators of soil biodiversity and land use monitoring—A review. Pol. J. Ecol. 2007, 55, 415–440. [Google Scholar]
- Koehler, H.H. Predatory mites (Gamasina, Mesostigmata). Agric. Ecosyst. Environ. 1999, 74, 395–410. [Google Scholar] [CrossRef]
- Neher, D.A.; Barbercheck, M.E. Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems. Insects 2019, 10, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badejo, M.A.; Woas, S.; Beck, L. Redescription of Archegozetes magnus [Sellinick, 1925] [Trhypochthonioidea] from Brazil and description of two new species of nanhermanniid mites: Bicyrthermannia nigeriana and Masthermannia seropedica [Nanhermannioidea] [Acari: Oribatida]. Genus. Int. J. Invertebr. Taxon. 2003, 14, 125–149. [Google Scholar]
- Gruss, I.; Twardowski, J.P.; Hurej, M. Influence of 90-year potato and winter rye monocultures under different fertilisation on soil mites. Plant Prot. Sci. 2017, 54, 31–38. [Google Scholar] [CrossRef]
- Gruss, I.; Twardowski, J. The assemblages of soil-dwelling springtails (Collembola) in winter rye under long-term monoculture and crop rotation. Žemdirbystė Agric. 2016, 103, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Twardowski, J.P.; Hurej, M.; Gruss, I. Diversity and abundance of springtails (Hexapoda: Collembola) in soil under 90-year potato monoculture in relation to crop rotation. Arch. Agron. Soil Sci. 2016, 62, 1158–1168. [Google Scholar] [CrossRef]
- Liu, D.; Sun, H.; Ma, H. Deciphering Microbiome Related to Rusty Roots of Panax ginseng and Evaluation of Antagonists against Pathogenic Ilyonectria. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Feng, G.; Li, X.; Chen, S. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jie, W.; Bai, L.; Yu, W.; Cai, B. Analysis of interspecific relationships between Funneliformis mosseae and Fusarium oxysporum in the continuous cropping of soybean rhizosphere soil during the branching period. Biocontrol Sci. Technol. 2015, 25, 1036–1051. [Google Scholar] [CrossRef]
- Hornby, D. Take-All Disease of Cereals: A Regional Perspective; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial Populations Responsible for Specific Soil Suppressiveness to Plant Pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J. Take-all of wheat. Physiol. Mol. Plant Pathol. 2003, 62, 73–86. [Google Scholar] [CrossRef]
- Chen, M.; Li, X.; Yang, Q.; Chi, X.; Pan, L.; Chen, N.; Yang, Z.; Wang, T.; Wang, M.; Yu, S. Soil Eukaryotic Microorganism Succession as Affected by Continuous Cropping of Peanut—Pathogenic and Beneficial Fungi were Selected. PLoS ONE 2012, 7, e40659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Cui, J.; Jie, W.; Cai, B. Analysis of the community compositions of rhizosphere fungi in soybeans continuous cropping fields. Microbiol. Res. 2015, 180, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Penton, C.R.; Lv, N.; Xue, C.; Yuan, X.; Ruan, Y.; Li, R.; Shen, Q. Banana Fusarium Wilt Disease Incidence Is Influenced by Shifts of Soil Microbial Communities Under Different Monoculture Spans. Microb. Ecol 2018, 75, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Han, M.; Hu, Y.; Li, Z.; Liu, C.; Wang, X.; Tian, Q.; Jiao, W.; Hu, J.; Liu, L.; et al. Effects of Continuous Cropping of Sweet Potato on the Fungal Community Structure in Rhizospheric Soil. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ciampi-Panno, L.; Fernandez, C.; Bustamante, P.; Andrade, N.; Ojeda, S.; Contreras, A. Biological control of bacterial wilt of potatoes caused by Pseudomonas solanacearum. Am. Potato J. 1989, 66, 315–332. [Google Scholar] [CrossRef]
- Niu, J.; Rang, Z.; Zhang, C.; Chen, W.; Tian, F.; Yin, H.; Dai, L. The succession pattern of soil microbial communities and its relationship with tobacco bacterial wilt. BMC Microbiol. 2016, 16, 233. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.; Zhang, S.; Liu, X.; Jiang, Q.; Ding, W. Soil acidification amendments change the rhizosphere bacterial community of tobacco in a bacterial wilt affected field. Appl. Microbiol. Biotechnol. 2018, 102, 9781–9791. [Google Scholar] [CrossRef] [Green Version]
- Luan, F.G.; Zhang, L.L.; Lou, Y.Y.; Wang, L.; Liu, Y.N.; Zhang, H.Y. Analysis of microbial diversity and niche in rhizosphere soil of healthy and diseased cotton at the flowering stage in southern Xinjiang. Genet. Mol. Res. 2015, 14, 1602–1611. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, B.-H.; Jin, Z.; Li, Z.; Song, H.; Ding, Y.-Q. Analysis of bacterial communities in rhizosphere soil of healthy and diseased cotton (Gossypium sp.) at different plant growth stages. Plant Soil 2011, 339, 447–455. [Google Scholar] [CrossRef]
- Wu, Z.; Hao, Z.; Zeng, Y.; Guo, L.; Huang, L.; Chen, B. Molecular characterization of microbial communities in the rhizosphere soils and roots of diseased and healthy Panax notoginseng. Antonie Van Leeuwenhoek 2015, 108, 1059–1074. [Google Scholar] [CrossRef]
- Govers, G.; Merckx, R.; Wesemael van, B.; Oost van, K. Soil conservation in the 21st century: Why we need smart agricultural intensification. Soil 2017, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, R.; Chen, S.; Qi, G.; He, Z.; Zhao, X. Microbial taxa and functional genes shift in degraded soil with bacterial wilt. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- She, S.; Niu, J.; Zhang, C.; Xiao, Y.; Chen, W.; Dai, L.; Liu, X.; Yin, H. Significant relationship between soil bacterial community structure and incidence of bacterial wilt disease under continuous cropping system. Arch. Microbiol. 2017, 199, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xiao, Y.; Lv, F.; Hu, L.; Wei, L.; Yuan, Z.; Lin, H. Bacterial community structure and functional potential of rhizosphere soils as influenced by nitrogen addition and bacterial wilt disease under continuous sesame cropping. Appl. Soil Ecol. 2018, 125, 117–127. [Google Scholar] [CrossRef]
- Niblack, T.L. Soybean Cyst Nematode Management Reconsidered. Plant Dis. 2005, 89, 1020–1026. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Li, S.; Wei, W.; Xu, Y.; Yao, Q. Assessment of parasitic fungi for reducing soybean cyst nematode with suppressive soil in soybean fields of northeast China. Acta Agric. Scand. Sect. B Soil Plant Sci. 2017, 67, 730–736. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Soil immune responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef]
- Sanguin, H.; Sarniguet, A.; Gazengel, K.; Moënne-Loccoz, Y.; Grundmann, G.L. Rhizosphere bacterial communities associated with disease suppressiveness stages of take-all decline in wheat monoculture. New Phytol. 2009, 184, 694–707. [Google Scholar] [CrossRef]
- Wei, W.; Xu, Y.; Li, S.; Zhu, L.; Song, J. Developing suppressive soil for root diseases of soybean with continuous long-term cropping of soybean in black soil of Northeast China. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 279–285. [Google Scholar] [CrossRef]
- Hamid, M.I.; Hussain, M.; Wu, Y.; Zhang, X.; Xiang, M.; Liu, X. Successive soybean-monoculture cropping assembles rhizosphere microbial communities for the soil suppression of soybean cyst nematode. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Van Veen, J.A.; Van Elsas, J.D. Microbial Diversity in Soil: Selection of Microbial Populations by Plant and Soil Type and Implications for Disease Suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Dexter, A.R. Advances in characterization of soil structure. Soil Tillage Res. 1988, 11, 199–238. [Google Scholar] [CrossRef]
- Nunan, N.; Wu, K.; Young, I.M.; Crawford, J.W.; Ritz, K. Spatial distribution of bacterial communities and their relationships with the micro-architecture of soil. FEMS Microbiol. Ecol. 2003, 44, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.-J. Soil structure as an indicator of soil functions—A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Amézketa, E. Soil Aggregate Stability—A review. J. Sustain. Agric. 1999, 14, 83–151. [Google Scholar] [CrossRef]
- Almajmaie, A.; Hardie, M.; Acuna, T.; Birch, C. Evaluation of methods for determining soil aggregate stability. Soil Tillage Res. 2017, 167, 39–45. [Google Scholar] [CrossRef]
- Karlen, D.L.; Hurley, E.G.; Andrews, S.S.; Cambardella, C.A.; Meek, D.W.; Duffy, M.D.; Mallarino, A.P. Crop Rotation Effects on Soil Quality at Three Northern Corn/Soybean Belt Locations. Agron. J. 2006, 98, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Liu, C.; Wang, J.; Meng, Q.; Yuan, Y.; Ma, X.; Liu, X.; Zhu, Y.; Ding, G.; Zhang, J.; et al. Soil aggregates stability and storage of soil organic carbon respond to cropping systems on Black Soils of Northeast China. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Acikgoz, S.; Anderson, S.H.; Gantzer, C.J.; Thompson, A.L.; Miles, R.J. 125 Years of Soil and Crop Management on Sanborn Field: Effects on Soil Physical Properties Related to Soil Erodibility. Soil Sci. 2017, 182, 172–180. [Google Scholar] [CrossRef]
- Miao, S.; Qiao, Y.; Li, P.; Han, X.; Tang, C. Fallow associated with autumn-plough favors structure stability and storage of soil organic carbon compared to continuous maize cropping in Mollisols. Plant Soil 2017, 416, 27–38. [Google Scholar] [CrossRef]
- Abe, S.S.; Hashi, I.; Masunaga, T.; Yamamoto, S.; Honna, T.; Wakatsuki, T. Soil Profile Alteration in a Brown Forest Soil under High-Input Tea Cultivation. Plant Prod. Sci. 2006, 9, 457–461. [Google Scholar] [CrossRef]
- Oh, K.; Kato, T.; Li, Z.-P.; Li, F.-Y. Environmental Problems From Tea Cultivation in Japan and a Control Measure Using Calcium Cyanamide1 1Project supported by the National Natural Science Foundation of China (No. 40471066) and the Knowledge Innovation Program of the Chinese Academy of Sciences (No. KZCX3-SW-417). Pedosphere 2006, 16, 770–777. [Google Scholar] [CrossRef]
- Zhang, Y.; Bhattacharyya, R.; Dalal, R.C.; Wang, P.; Menzies, N.W.; Kopittke, P.M. Impact of land use change and soil type on total phosphorus and its fractions in soil aggregates. Land Degrad. Dev. 2020, 31, 828–841. [Google Scholar] [CrossRef]
- Hati, K.M.; Swarup, A.; Dwivedi, A.K.; Misra, A.K.; Bandyopadhyay, K.K. Changes in soil physical properties and organic carbon status at the topsoil horizon of a vertisol of central India after 28 years of continuous cropping, fertilization and manuring. Agric. Ecosyst. Environ. 2007, 119, 127–134. [Google Scholar] [CrossRef]
- Aboim, M.C.R.; Coutinho, H.L.C.; Peixoto, R.S.; Barbosa, J.C.; Rosado, A.S. Soil bacterial community structure and soil quality in a slash-and-burn cultivation system in Southeastern Brazil. Appl. Soil Ecol. 2008, 38, 100–108. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, B.; Wang, X.; Gao, X.; Liu, G. Erosion effect on the productivity of black soil in Northeast China. Sci. China Ser. D Earth Sci. 2009, 52, 1005–1021. [Google Scholar] [CrossRef]
- Jia, Z.; Kuzyakov, Y.; Myrold, D.; Tiedje, J. Soil Organic Carbon in a Changing World. Pedosphere 2017, 27, 789–791. [Google Scholar] [CrossRef]
- Kay, B.D. Soil Structure and Organic Carbon—A Review. Available online: https://www.taylorfrancis.com/ (accessed on 8 April 2020).
- Nunan, N.; Schmidt, H.; Raynaud, X. The ecology of heterogeneity: Soil bacterial communities and C dynamics. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190249. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Miao, S.; Li, Y.; Zhong, X. Chemical composition of soil organic carbon changed by long-term monoculture cropping system in Chinese black soil. Plant Soil Environ. 2018, 64, 557–563. [Google Scholar] [CrossRef] [Green Version]
- Börjesson, G.; Bolinder, M.A.; Kirchmann, H.; Kätterer, T. Organic carbon stocks in topsoil and subsoil in long-term ley and cereal monoculture rotations. Biol. Fertil. Soils 2018, 54, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, A.J.; Allen, F.L.; Wight, J.P.; Saxton, A.M.; Tyler, D.D.; Sams, C.E. Soil Organic Carbon Sequestration Rates under Crop Sequence Diversity, Bio-Covers, and No-Tillage. Soil Sci. Soc. Am. J. 2014, 78, 1726–1733. [Google Scholar] [CrossRef]
- Lal, R. Enhancing crop yields in the developing countries through restoration of the soil organic carbon pool in agricultural lands. Land Degrad. Dev. 2006, 17, 197–209. [Google Scholar] [CrossRef]
- Fiorini, A.; Boselli, R.; Maris, S.C.; Santelli, S.; Ardenti, F.; Capra, F.; Tabaglio, V. May conservation tillage enhance soil C and N accumulation without decreasing yield in intensive irrigated croplands? Results from an eight-year maize monoculture. Agric. Ecosyst. Environ. 2020, 296, 106926. [Google Scholar] [CrossRef]
- Kätterer, T.; Bolinder, M.A.; Andrén, O.; Kirchmann, H.; Menichetti, L. Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric. Ecosyst. Environ. 2011, 141, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Kirchmann, H.; Schön, M.; Börjesson, G.; Hamnér, K.; Kätterer, T. Properties of soils in the Swedish long-term fertility experiments: VII. Changes in topsoil and upper subsoil at Örja and Fors after 50 years of nitrogen fertilization and manure application. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 25–36. [Google Scholar] [CrossRef]
- Blackmer, A.M. Nitrogen Fertilizer Recommendations for Corn in Iowa; FAO: Roma, Italy, 1997. [Google Scholar]
- Ding, W.; Hume, D.J.; Vyn, T.J.; Beauchamp, E.G. N credit of soybean to a following corn crop in central Ontario. Can. J. Plant Sci. 1998, 78, 29–33. [Google Scholar] [CrossRef]
- Gentry, L.F.; Ruffo, M.L.; Below, F.E. Identifying Factors Controlling the Continuous Corn Yield Penalty. Agron. J. 2013, 105, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Ney, L.; Franklin, D.; Mahmud, K.; Cabrera, M.; Hancock, D.; Habteselassie, M.; Newcomer, Q.; Dahal, S. Impact of inoculation with local effective microorganisms on soil nitrogen cycling and legume productivity using composted broiler litter. Appl. Soil Ecol. 2020, 154, 103567. [Google Scholar] [CrossRef]
- Mahal, N.; Osterholz, W.; Miguez, F.; Poffenbarger, H.; Sawyer, J.; Olk, D.; Archontoulis, S.; Castellano, M. Nitrogen Fertilizer Suppresses Mineralization of Soil Organic Matter in Maize Agroecosystems. Front. Ecol. Evol. 2019, 7, 59. [Google Scholar] [CrossRef] [Green Version]
- Kaboneka, S.; Sabbe, W.E.; Mauromoustakos, A. Carbon decomposition kinetics and nitrogen mineralization from corn, soybean, and wheat residues. Commun. Soil Sci. Plant Anal. 1997, 28, 1359–1373. [Google Scholar] [CrossRef]
- Gentry, L.E.; Below, F.E.; David, M.B.; Bergerou, J.A. Source of the soybean N credit in maize production. Plant Soil 2001, 236, 175–184. [Google Scholar] [CrossRef]
- Meng, Y.; Chua-Ona, T.; Thompson, M.L. Short-term Nitrogen Mineralization Potential in Soils of Biofuel Cropping Systems. Soil Sci. 2016, 181, 503–512. [Google Scholar] [CrossRef]
- Cai, A.; Xu, H.; Shao, X.; Zhu, P.; Zhang, W.; Xu, M.; Murphy, D.V. Carbon and Nitrogen Mineralization in Relation to Soil Particle-Size Fractions after 32 Years of Chemical and Manure Application in a Continuous Maize Cropping System. PLoS ONE 2016, 11, e0152521. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, W.W.; Wortmann, C.S. Tillage and Rotation Interactions for Corn and Soybean Grain Yield as Affected by Precipitation and Air Temperature. Agron. J. 2004, 96, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.J.; Russell, A.E.; Moore, A.R. Do corn-soybean rotations enhance decomposition of soil organic matter? Plant Soil 2019, 444, 427–442. [Google Scholar] [CrossRef]
- Hussain, M.Z.; Bhardwaj, A.K.; Basso, B.; Robertson, G.P.; Hamilton, S.K. Nitrate Leaching from Continuous Corn, Perennial Grasses, and Poplar in the US Midwest. J. Environ. Qual. 2019, 48, 1849–1855. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.E.; Willson, T.C.; Kizilkaya, K.; Parker, E.; Harwood, R.R. Enhancing the Mineralizable Nitrogen Pool Through Substrate Diversity in Long Term Cropping Systems. Soil Sci. Soc. Am. J. 2001, 65, 1442–1447. [Google Scholar] [CrossRef]
- Xiong, W.; Li, Z.; Liu, H.; Xue, C.; Zhang, R.; Wu, H.; Li, R.; Shen, Q. The Effect of Long-Term Continuous Cropping of Black Pepper on Soil Bacterial Communities as Determined by 454 Pyrosequencing. PLoS ONE 2015, 10, e0136946. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, Y.; Wang, J.; Yang, L.; Zhang, S.; Xu, C.; Ding, W. Soil Acidification Aggravates the Occurrence of Bacterial Wilt in South China. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Z.; Arafat, Y.; Lin, W. Studies on fungal communities and functional guilds shift in tea continuous cropping soils by high-throughput sequencing. Ann. Microbiol. 2020, 70, 7. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, J.; Corso, M.R.D.; Figueiredo, B.; Nambi, E.; Raun, W. Soil Organic Carbon, Total Nitrogen, and Soil pH, in a Long-Term Continuous Winter Wheat (Triticum aestivum, L.) Experiment. Commun. Soil Sci. Plant Anal. 2018, 49, 803–813. [Google Scholar] [CrossRef]
- Sakuma, F.; Maeda, M.; Takahashi, M.; Hashizume, K.; Kondo, N. Suppression of Common Scab of Potato Caused by Streptomyces turgidiscabies Using Lopsided Oat Green Manure. Plant Dis. 2011, 95, 1124–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juo, A.S.; Franzluebbers, K.; Dabiri, A.; Ikhile, B. Changes in soil properties during long-term fallow and continuous cultivation after forest clearing in Nigeria. Agric. Ecosyst. Environ. 1995, 56, 9–18. [Google Scholar] [CrossRef]
- Heidari, G.; Mohammadi, K.; Sohrabi, Y. Responses of Soil Microbial Biomass and Enzyme Activities to Tillage and Fertilization Systems in Soybean (Glycine max L.) Production. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agegnehu, G.; Nelson, P.N.; Bird, M.I. Crop yield, plant nutrient uptake and soil physicochemical properties under organic soil amendments and nitrogen fertilization on Nitisols. Soil Tillage Res. 2016, 160, 1–13. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011; ISBN 978-0-12-384906-9. [Google Scholar]
- Nyamadzawo, G.; Chikowo, R.; Nyamugafata, P.; Giller, K.E. Improved legume tree fallows and tillage effects on structural stability and infiltration rates of a kaolinitic sandy soil from central Zimbabwe. Soil Tillage Res. 2007, 96, 182–194. [Google Scholar] [CrossRef]
- Agegnehu, G.; Lakew, B.; Nelson, P.N. Cropping sequence and nitrogen fertilizer effects on the productivity and quality of malting barley and soil fertility in the Ethiopian highlands. Arch. Agron. Soil Sci. 2014, 60, 1261–1275. [Google Scholar] [CrossRef]
- Jiao, X.-L.; Zhang, X.-S.; Lu, X.-H.; Qin, R.; Bi, Y.-M.; Gao, W.-W. Effects of maize rotation on the physicochemical properties and microbial communities of American ginseng cultivated soil. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Malhi, S.S.; Nyborg, M.; Harapiak, J.T. Effects of long-term N fertilizer-induced acidification and liming on micronutrients in soil and in bromegrass hay. Soil Tillage Res. 1998, 48, 91–101. [Google Scholar] [CrossRef]
- Ashworth, A.J.; Owens, P.R.; Allen, F.L. Long-term cropping systems management influences soil strength and nutrient cycling. Geoderma 2020, 361, 114062. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Z.; Li, Z.W.; Jiang, Y.H.; Weng, B.Q.; Lin, W.X. Variations of rhizosphere bacterial communities in tea (Camellia sinensis L.) continuous cropping soil by high-throughput pyrosequencing approach. J. Appl. Microbiol. 2016, 121, 787–799. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Contreras, J.; Hupp, B.M.; Lindenberger, J.; Chen, D.; Zhang, Q.; Wang, C.; Twigg, P.; Saleem, M. Root microbiome changes with root branching order and root chemistry in peach rhizosphere soil. Rhizosphere 2020, 100249. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Chomel, M.; Fernandez, C.; Bousquet-Mélou, A.; Gers, C.; Monnier, Y.; Santonja, M.; Gauquelin, T.; Gros, R.; Lecareux, C.; Baldy, V. Secondary metabolites of Pinus halepensis alter decomposer organisms and litter decomposition during afforestation of abandoned agricultural zones. J. Ecol. 2014, 102, 411–424. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, G.; Cheng, Y.; Shi, P.; Yang, C.; Yang, H.; Xu, Z. Soil acidification in continuously cropped tobacco alters bacterial community structure and diversity via the accumulation of phenolic acids. Sci. Rep. 2019, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.-Y.; Kong, C.-H.; Wang, J.-G.; Wang, Y.-F. Rhizosphere isoflavones (daidzein and genistein) levels and their relation to the microbial community structure of mono-cropped soybean soil in field and controlled conditions. Soil Biol. Biochem. 2011, 43, 2257–2264. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Zhang, J.; Yao, T.; Wei, D.; Wang, Y.; Wang, J. Effect of root exudates on beneficial microorganisms—Evidence from a continuous soybean monoculture. Plant Ecol. 2012, 213, 1883–1892. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, Y.; Zhu, W.; Tang, J.; Hu, S.; Zhou, T.; Chen, X. Baicalin Released from Scutellaria baicalensis Induces Autotoxicity and Promotes Soilborn Pathogens. J. Chem. Ecol. 2010, 36, 329–338. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, H.; Ge, X.; Wu, F. Effects of vanillin on the community structures and abundances of Fusarium and Trichoderma spp. in cucumber seedling rhizosphere. J. Plant Interact. 2018, 13, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Kitazawa, H.; Asao, T.; Ban, T.; Pramanik, M.H.R.; Hosoki, T. Autotoxicity of root exudates from strawberry in hydroponic culture. J. Hortic. Sci. Biotechnol. 2005, 80, 677–680. [Google Scholar] [CrossRef]
- Gadelha, I.C.N.; Fonseca, N.B.S.; Oloris, S.C.S.; Melo, M.M.; Soto-Blanco, B. Gossypol Toxicity from Cottonseed Products. Available online: https://www.hindawi.com/journals/tswj/2014/231635/ (accessed on 28 March 2020).
- Zhou, X.; Wu, F. p-Coumaric Acid Influenced Cucumber Rhizosphere Soil Microbial Communities and the Growth of Fusarium oxysporum f. sp. cucumerinum Owen. PLoS ONE 2012, 7, e48288. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M. Bacteria-Protist Interactions in the Context of Biodiversity and Ecosystem Functioning Research. Ph.D. Thesis, University of Leipzig, Leipzig, Germany, 2012. [Google Scholar]
Figure 1.
Predicted hypothetical impact of continuous cropping (CC) on soil health.

Figure 2.
The summary of the comparative effects of continuous monoculture vs. alternative cropping (inter, mixture, cover-crop rotation) systems on multiple health indicators of soil health.
Figure 2.
The summary of the comparative effects of continuous monoculture vs. alternative cropping (inter, mixture, cover-crop rotation) systems on multiple health indicators of soil health.


{kind=link}
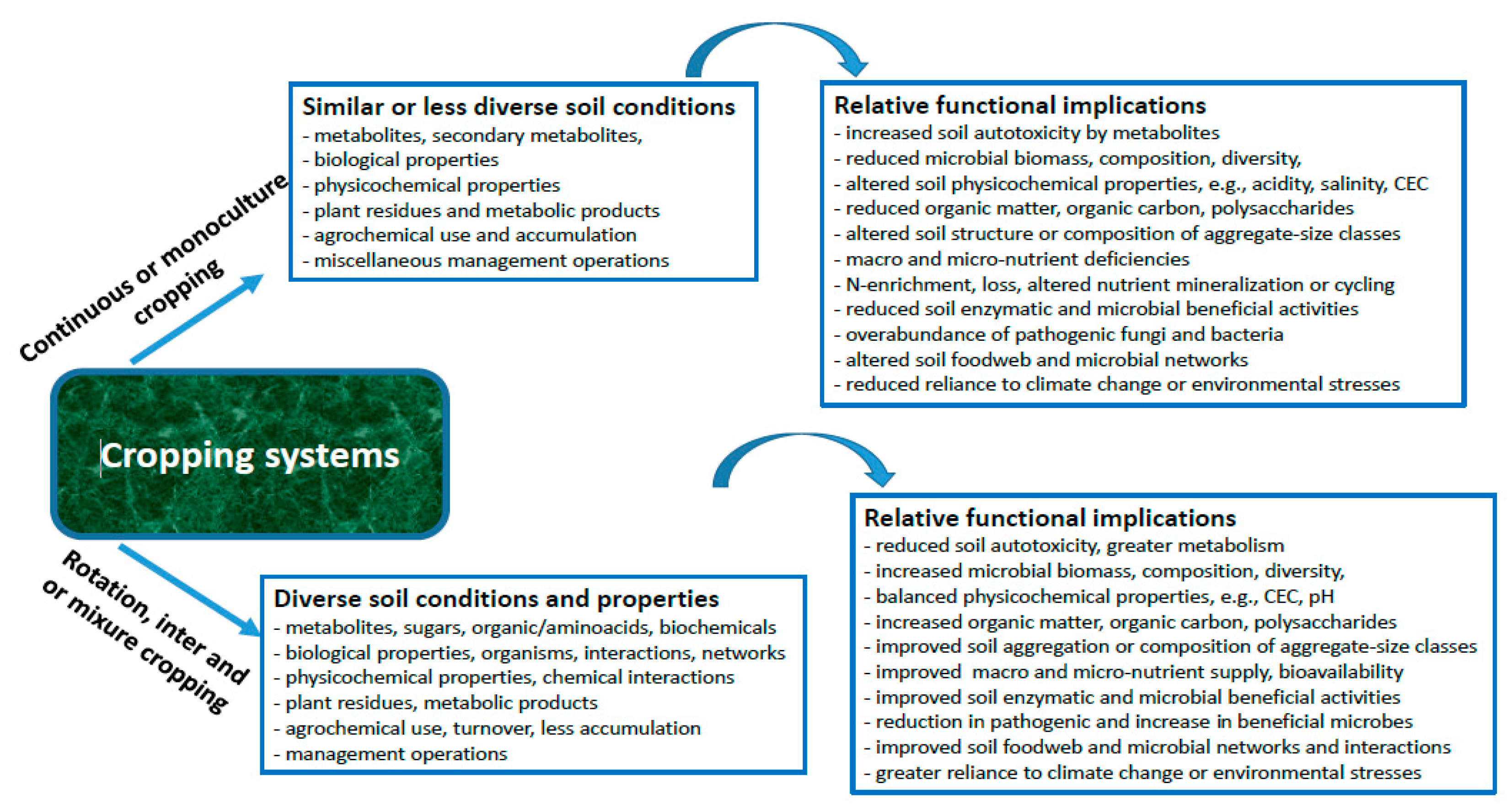{kind=link}
Table 1.
A brief overview of key indicators of soil health.
| Soil Indicators | Functions | Monoculture Cropping | Measures to Improve | Relevant Literature |
|---|---|---|---|---|
| Visual indicators | ||||
| Weeds | Low weed coverage or presence is an indicator of healthy soil. | Promotes weeds. | Weeding, cover crops, cleaned use of farm machinery, increasing cropping diversity. | [28,29] |
| Soil surface | Relatively even soil surface and less shallow patches are indicators of healthy soil. | Alters the degree of soil surface evenness. | Precise use of farm machinery, proper land leveling | [30,31] |
| Water or moisture distribution | Relatively even moisture and fewer water ponds are indicators of healthy soil. | May alter moisture distribution in soil. | Proper land leveling, cover cropping, even distribution of crop residues and snow. | [32,33] |
| Physical indicators | ||||
| Soil texture | Loam-textured soils are considered productive. Sandy or clay-textured soils challenge plant growth. | Though no effect on texture, it may intensify textural effects. | Organic amendments, crop residues, biochar, livestock, and poultry waste, could improve soil conditions. | [34] |
| Bulk density (BD) | Soil with BD 1.3–1.6 g/cm3 is considered good for roots. | May increase BD. | Same as above, reduced use of farm machinery. No-till agriculture. | [35,36,37] |
| Soil porosity | Macro-, meso-, and micro-porosities regulate nutrient, gas, and water mobility and improve soil functions. | May reduce soil porosity. | Same as above, reduced use of farm machinery. No-till agriculture. | [38,39] |
| Available water capacity (AWC) | The AWC determines soil functioning. | May alter AWC. | Water conservation practices, mulching, organic amendments, crop residues. | [40,41] |
| Penetration resistance | It determines root penetration into soil and root-microbe (e.g., mycorrhizae) interactions. | May increase penetration resistance. | Organic amendments, no-till agriculture, cover cropping. | [42,43] |
| Saturated hydraulic conductivity | It may influence water and nutrient movement. | May alter or reduce hydraulic conductivity. | Organic amendments, no-till agriculture, cover cropping. | [44,45] |
| Soil aggregate size classes, their composition | These determine soil structure, nutrient dynamics, microbial activities, plant growth, and carbon (C) storage. | May reduce soil aggregation or aggregate composition. | No-till, cover cropping, organic amendments, cropping diversity. | [46,47,48] |
| Soil surface hardness | It is a measure of the maximum soil surface (0–15 cm depth) penetration resistance (psi), and it is determined by a field penetrometer. It must not exceed 300 psi. Low values are good for soil functioning and root growth. | May increase soil hardiness | No-till, crop rotation, organic amendment, crop residues, and reduced use of farm machinery. | [49,50,51] |
| Subsurface Hardness: | It is also a measure of the maximum resistance (psi) in soil between 15 to 45 cm depths using a field penetrometer. | May increase soil hardiness | Same as above. | [49,50,51,52] |
| Soil aggregate Stability | It is a measure of soil aggregates’ resistance to disintegration under raindrop impact. It is determined by testing the impact of simulated rainfall on aggregates in the soil sieves (0.25–2.0 mm). The proportion of soil that stays on the sieve reflects the percentage of aggregate stability. Healthy soils should have >50% stable aggregates. | May reduce soil aggregate stability. | No-till, crop rotation, organic amendment, cover cropping mulching, and reduced use of machinery. | [53,54,55] |
| Biological indicators | ||||
| Root pathogen pressure assessment | It is an estimation of pathogen load on plant roots and it is rated from 2 to 9. The higher numbers show higher plant sensitivity and pathogen damage. | May increase root pathogen pressure. | The mixture, inter-cropping, cover cropping, organic amendments, and biopesticides. | [56,57] |
| Beneficial nematode population | A higher abundance of beneficial nematodes is considered as a good indicator of soil health. | May reduce their abundance. | The mixture, inter-cropping, cover cropping, cropping diversity, organic amendments, | [12,58,59] |
| Earthworms | A higher abundance of earthworms is considered a good indicator of soil health. | May reduce their abundance. | The mixture, inter-cropping, cover cropping, cropping diversity, and organic amendments. | [12,58,59] |
| Parasitic nematode population | A low abundance of parasitic nematodes is considered a good indicator of soil health. | May increase their abundance | Biopesticides, mixture, inter-cropping, cover cropping, and crop rotation. | [60,61,62] |
| Potentially mineralizable N (PMN) | It is defined as the fraction of organic N that is available to plants. | May reduce PMN. | Cropping diversity, crop rotation, cover cropping, organic amendments, and crop residues. | [63,64] |
| Cellulose decomposition rate | Higher decomposition leads to greater C and nutrient availability to soil organisms and health. | May reduce decomposition. | Organic amendments, crop rotation, cover cropping, crop residues, and organic amendments. | [65,66] |
| Particulate organic matter (POM) | POM is a fundamental indicator of soil health. | May reduce POM. | The mixture, inter-cropping, cover cropping, and organic amendments. | [67,68] |
| Active C | It is a small portion of SOM that is readily available for soil organisms and regulates soil food web interactions. | May activate C. | Organic amendments, cropping diversity, cover cropping, and crop residues. | [25,69] |
| Weed seed bank | A higher weed seed bank may lead to weed invasion, and it is detrimental to soil health. | May increase weed seed bank. | Cropping diversity, cover cropping, and crop rotation | [70,71] |
| Microbial respiration rate | It is an indicator of microbial activities and soil functions. It also determines soil C storage and sequestration. | May alter respiration. | Organic amendments, crop residues, crop rotation, cover cropping, and cropping diversity. | [72,73,74] |
| Soil protein contents. | These determine the quality of SOM, and the level of soil biological activities. | Crop residues, organic amendments, cover cropping, and crop rotation. | [75,76] | |
| Microbiomic indicators | ||||
| Microbial networks, interactions | Microbial networks and teamwork regulate multiple soil functions. | May alter or deplete microbial networks. | Cropping diversity, crop rotation, cover cropping, biofertilizers, organic amendments, biochar, and crop residues. | [11,77,78] |
| Microbial abundance, biomass | Higher microbial biomass contents are considered an indicator of healthy soil. | May reduce or alter microbial biomass. | Same as above. | [79,80] |
| Community composition | Higher values of community composition such as species evenness are indicators of healthy soil. | May reduce or alter the microbial community structure. | Same as above. | [17,81] |
| Diversity | Higher values of microbial diversity such as species richness are indicators of healthy soil. | May reduce or alter microbial species richness. | Same as above. Microbial probiotics | [17,81,82,83] |
| Beneficial properties | Soils with microbes of diverse properties are considered healthy soils. | May reduce the abundance of beneficial microbes. | Same as above. Microbial probiotics. | [82,84] |
| Food web interactions | Soils with a greater abundance of different trophic level organisms and interactions are considered healthy soils. | May alter soil trophic complexity and interactions. | Same as above. Microbial probiotics. | [85,86,87] |
| Chemical indicators | ||||
| P | The optimum level of P (30–50 ppm) is also an indicator of healthy soil. | May reduce soil P contents. | Organic amendments, biofertilizers, crop residues, and cropping diversity. | [88,89,90] |
| N | The optimum level of N (50–75 mg-N/kg soil) is also an indicator of healthy soil. | May alter soil N contents. | Organic amendments, biofertilizers, crop residues, and legume cover cropping. | [88,89,90] |
| K | The optimum level of K (50–75 mg-N/kg soil) is also an indicator of healthy soil. | May reduce soil K contents. | Organic amendments, biofertilizers, crop residues, and cover crops. | [91] |
| pH | Near neutral pH is a sign of healthy soils while low and high pH damage soil health by causing acidity and salinity. | May alter soil pH. In slight acidic and saline soils, it may increase soil acidity and salinity, respectively. | Crop rotation, cover cropping, organic amendments, and crop residues. | [89] |
| Magnesium (Mg) | The optimum level of Mg is also an indicator of healthy soil. | May alter or reduce soil Mg contents. | Dolomitic limestone, quality irrigation water, cover cropping, and crop residues. | [92,93] |
| Calcium (Ca) | The optimum level of Ca determines soil structure and pH, and are an indicator of healthy soil. | May alter or reduce soil Ca contents. | Limestone, gypsum, irrigation water, and crop residues. | [93,94,95] |
| Iron (Fe) | The optimum level of soil Fe contents is an indicator of healthy soil. | May alter or reduce soil Fe contents. | Iron chelates, manure or sewage sludge, foliar spray of ferrous sulfate solution | [93,94,95,96] |
| Aluminum | [93,94,95,96] | |||
| Manganese (Mn) | The optimum levels of soil Mn contents are an indicator of healthy soil. | May alter or reduce soil Mn contents. | Crop residues, organic amendments, and conservation tillage. | [93,94,95,96,97] |
| Zinc (Zn) | The optimum levels of soil Zn contents are an indicator of healthy soil. | May alter or reduce soil Zn contents. | Crop residues, zinc sulfate, zinc chelates, biofertilizers, | [93,94,95,96,97] |
| Copper (Cu) | The optimum levels of soil Cu contents are an indicator of healthy soil. | May alter or reduce soil Cu contents. | Organic matter, organic amendments, biofertilizers, and crop residues. | [66,67,68,69] |
| Exchangeable acidity | It is defined as the sum of the concentrations of Al and hydrogen (H) ions in the soil exchange complex, and it is inversely related to the pH and base saturation of the soil. | May alter it and soil pH | Lime, organic matter, manure, crop rotation, and cover cropping. | [98,99] |
| Salinity/sodicity | Higher salt contents damage soil structure and salinity is linked to soil degradation. | May increase soil salinity in arid zones. | Organic amendments, cover cropping, crop residues, and crop rotation. | [100] |
| Heavy metals | Higher heavy metal contents damage soil health. Lower concentrations are considered as an indicator of healthy soil. | May increase or alter heavy metal contents. | Heavy metal-free organic amendments, crop residues, | [101,102] |
Note: Soil health indicator is a very broad term, and there are no universal values for them. Their significance and ranges may differ across different soil types, cropping systems, and climatic zones.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.; Chen, D.; Wei, H.; Saleem, M. Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health. Soil Syst. 2020, 4, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems4040059
AMA Style
Pervaiz ZH, Iqbal J, Zhang Q, Chen D, Wei H, Saleem M. Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health. Soil Systems. 2020; 4(4):59. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems4040059
Chicago/Turabian StylePervaiz, Zahida H., Javed Iqbal, Qingming Zhang, Dima Chen, Hui Wei, and Muhammad Saleem. 2020. "Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health" Soil Systems 4, no. 4: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems4040059