Essential Oil Content of Baccharis crispa Spreng. Regulated by Water Stress and Seasonal Variation

 ,
,  ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
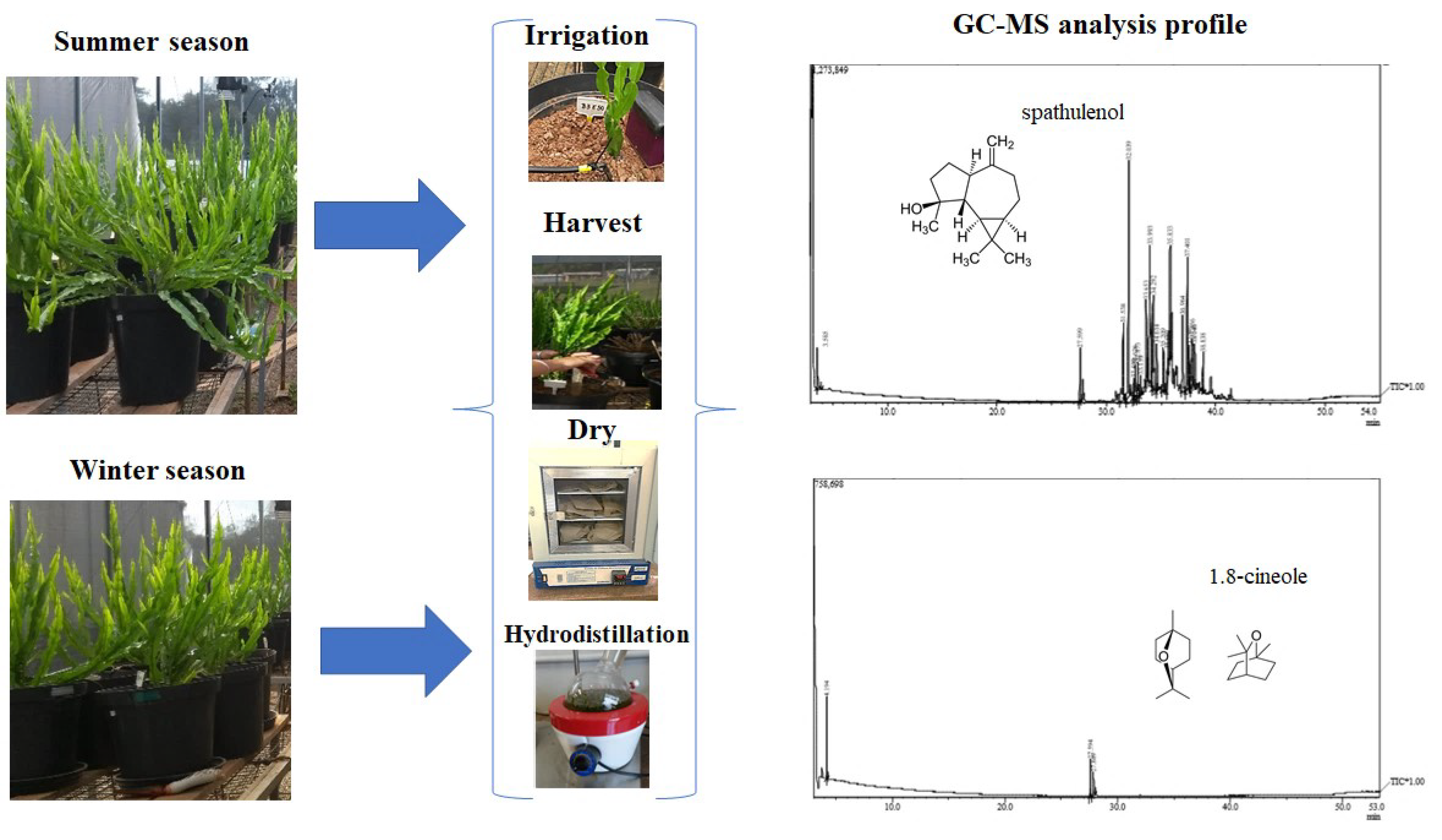
2.1. Plant Material
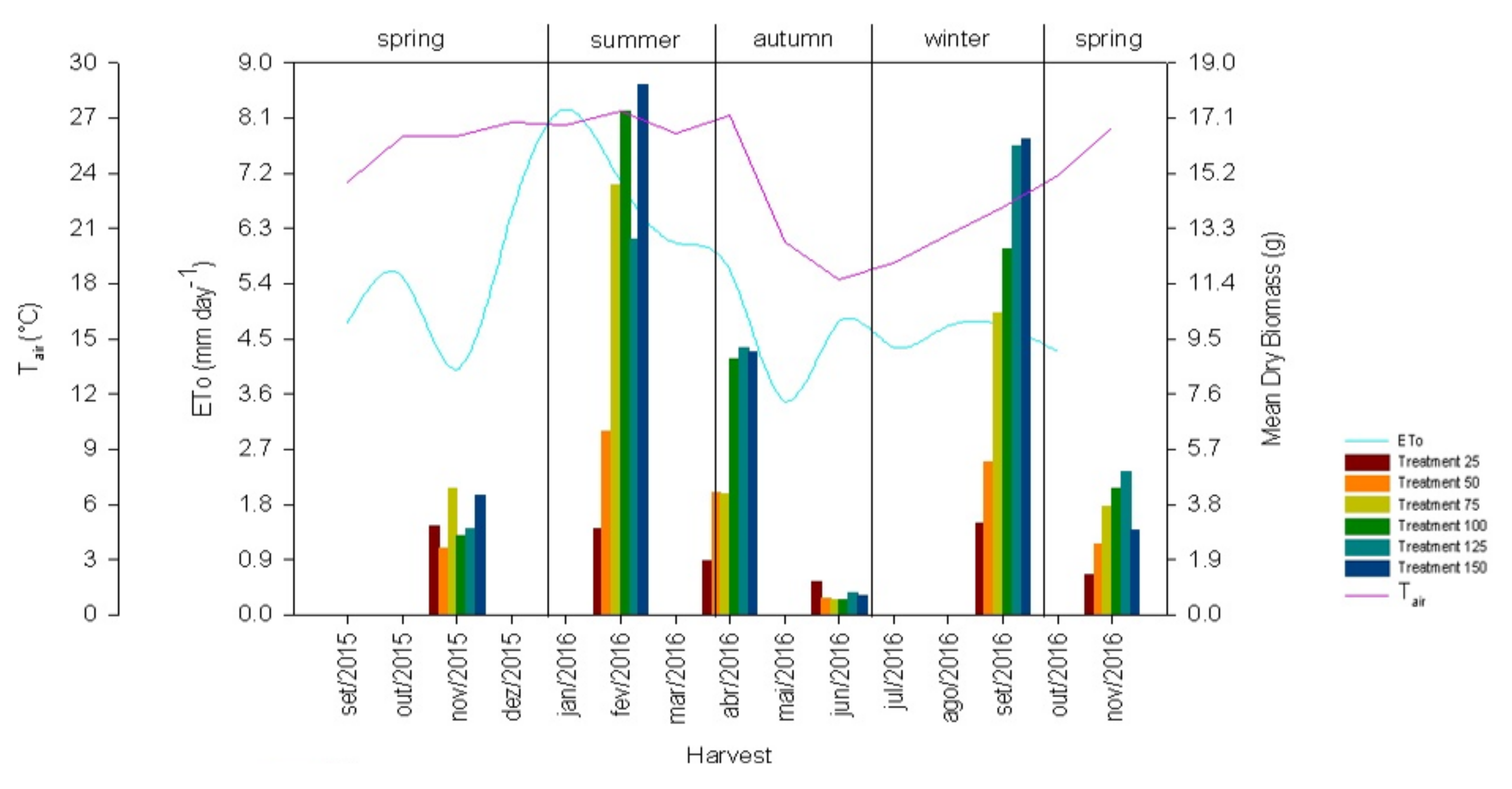
2.2. Water Deficit Treatments
2.3. Extraction of the Essential Oil (EO)
2.4. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Environmental Data
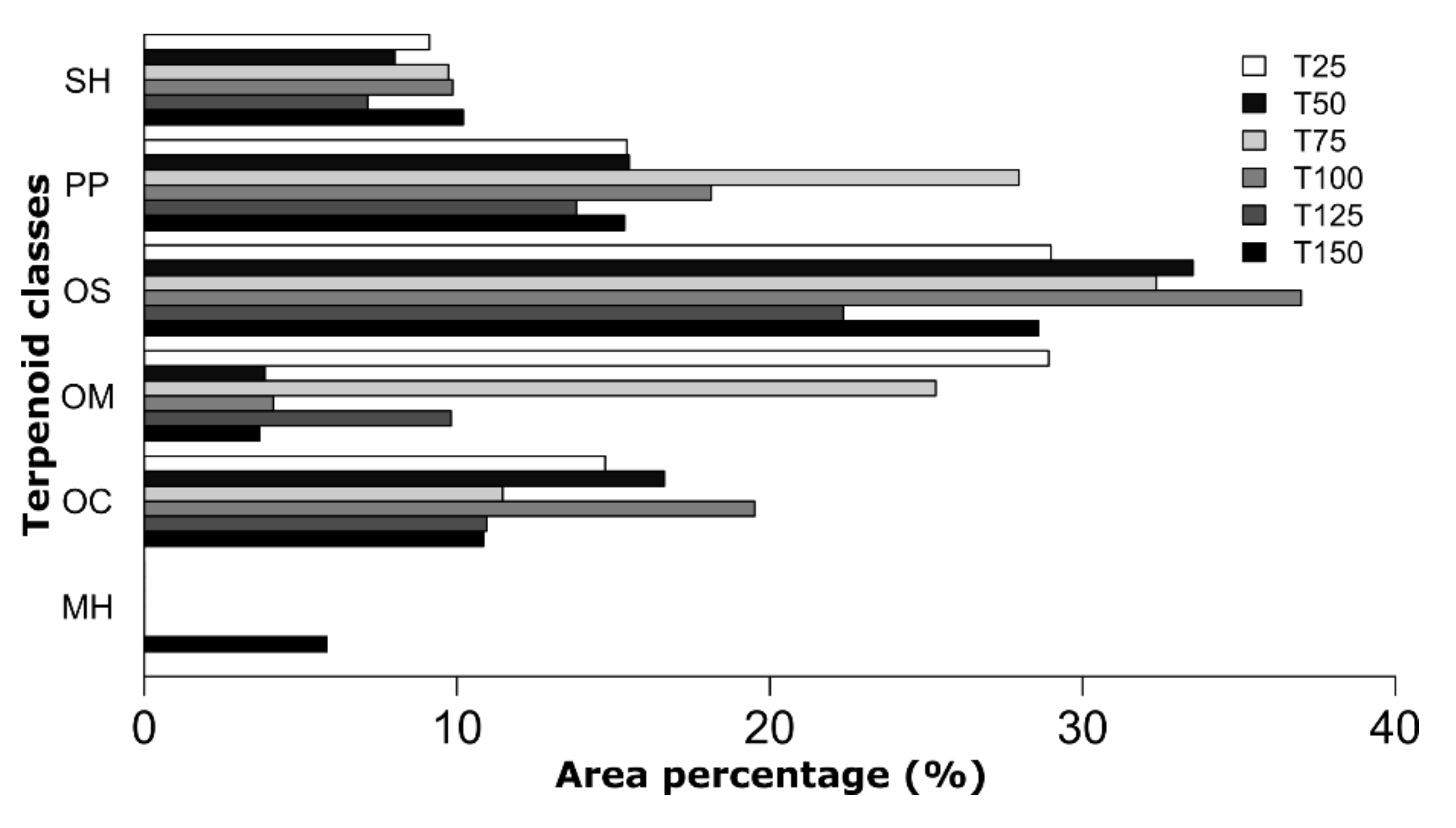
3.2. Essential Oil Components
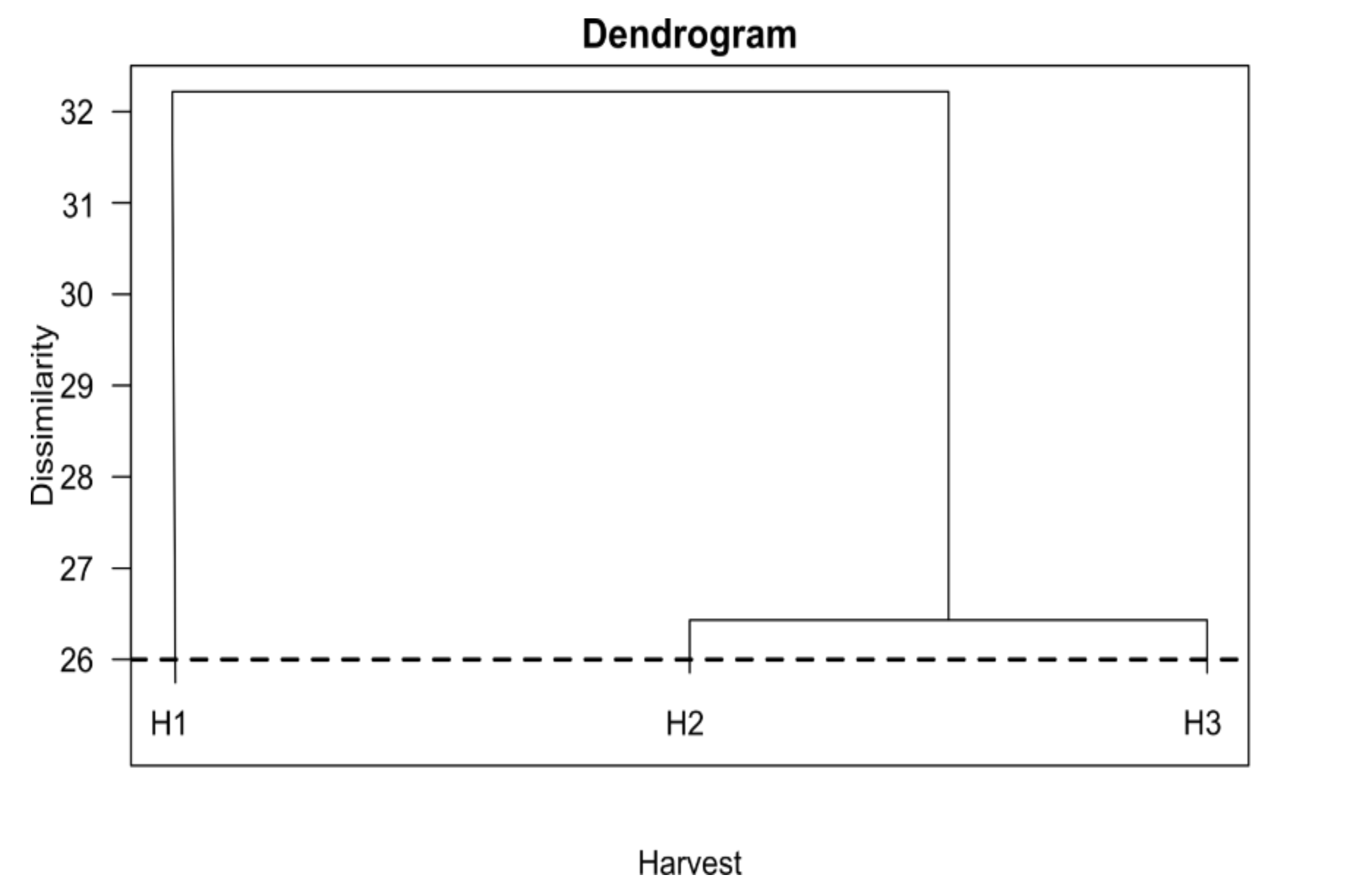
3.3. Differences in Content and Composition of Essential Oils between the Three Harvest Times
3.4. Volatile Profiles under Different Irrigation Conditions
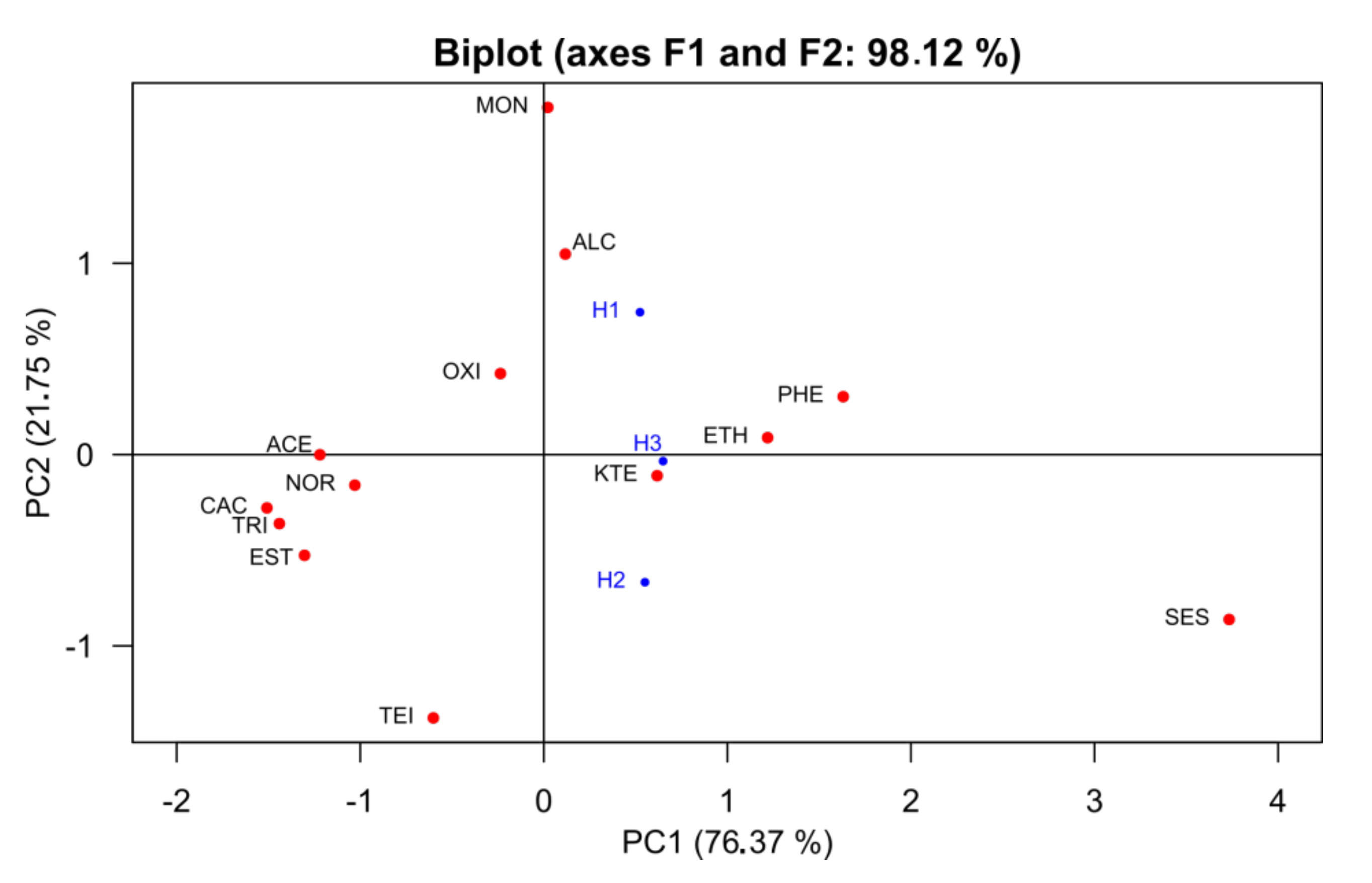
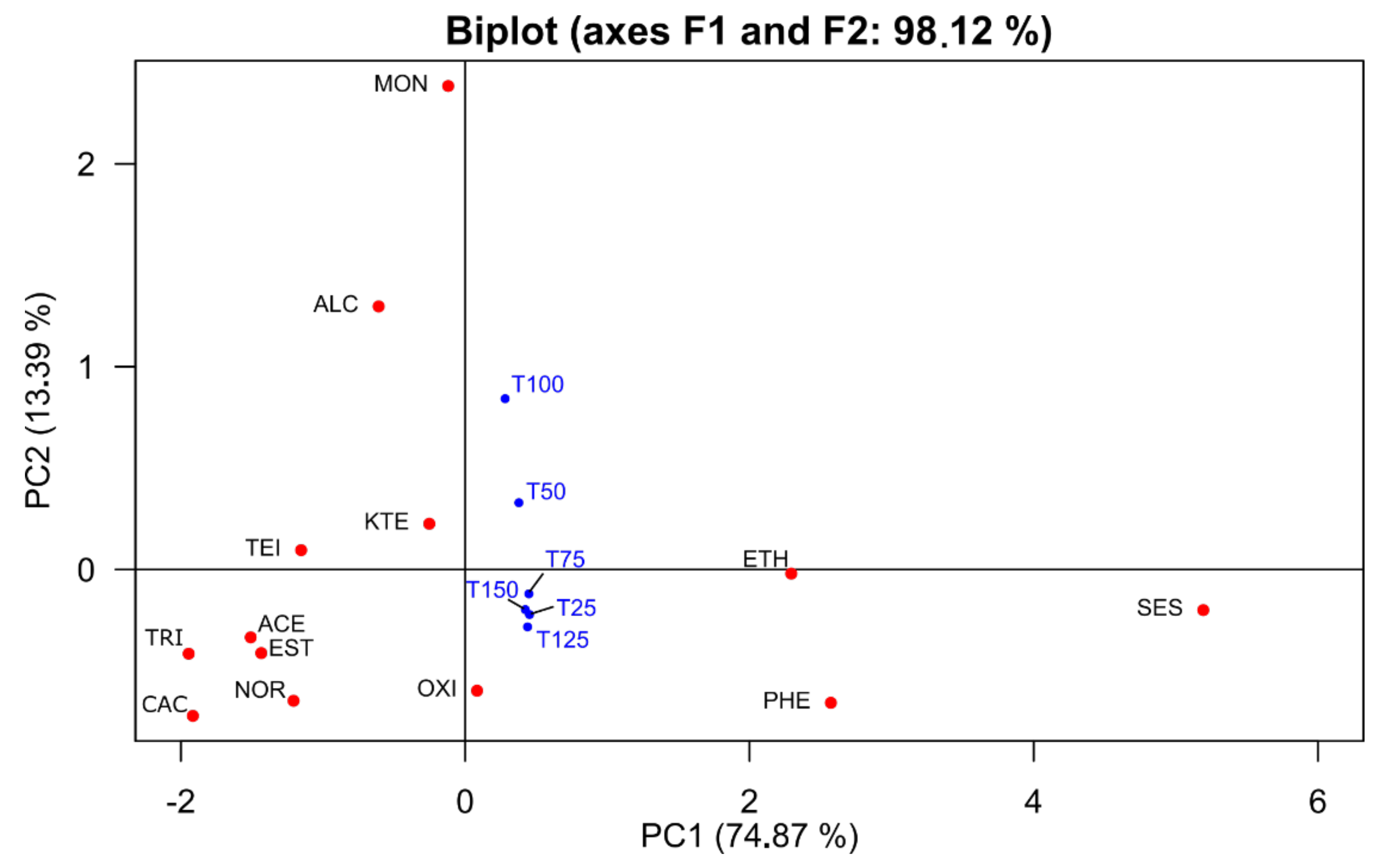
3.5. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dos Santos, D.A.; Fukui, M.D.J.; Nanayakkara, N.D.; Khan, S.I.; Sousa, J.P.B.; Bastos, J.K.; De Andrade, S.F.; Filho, A.A.D.S.; Quintão, N.L. Anti-inflammatory and antinociceptive effects of Baccharis dracunculifolia DC (Asteraceae) in different experimental models. J. Ethnopharmacol. 2010, 127, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, N.; Reis, P.; Laranja, G.; Pinto, A.; Aiub, C.; Felzenszwalb, I.; Paes, M.; Bastos, F.; Bastos, V.; Sabino, K.; et al. In vitro and in vivo toxicological evaluation of extract and fractions from Baccharis trimera with anti-inflammatory activity. J. Ethnopharmacol. 2011, 138, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.d.A. Desenvolvimento de Forma Farmacêutica Sólida à Base de Baccharis trimera (Less.) DC. Para o Tratamento da Artrite Reumatóide. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2012. [Google Scholar]
- Verdi, L.G.; Brighente, I.M.C.; Pizzolatti, M.G. Gênero Baccharis (Asteraceae): Aspectos químicos, económicos e biológicos. Química Nova 2005, 28, 85–94. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R.N.; Rehder, V.L.G.; Santos Oliveira, A.S.; Júnior, Í.M.; de Carvalho, J.E.; de Ruiz, A.L.T.G.; Sierpe Jeraldo, V.d.L.; Linhares, A.X.; Allegretti, S.M. Schistosoma mansoni: In vitro schistosomicidal activity of essential oil of Baccharis trimera (Less) DC. Exp. Parasitol. 2012, 132, 135–143. [Google Scholar] [CrossRef]
- Silva, F.G.; Oliveira, C.; Pinto, J.E.B.; Nascimento, V.E.; Santos, S.C.; Seraphin, J.C.; Ferri, P.H. Seasonal variability in the essenial oils of wild and cultivated Baccharis trimera. J. Braz. Chem. Soc. 2007, 18, 990–997. [Google Scholar] [CrossRef]
- De Oliveira, R.N.; Rehder, V.L.G.; Oliveira, A.S.S.; Jeraldo, V.d.L.S.; Linhares, A.X.; Allegretti, S.M. Anthelmintic activity in vitro and in vivo of Baccharis trimera (Less) DC. against immature and adult worms of Schistosoma mansoni. Exp. Parasitol. 2014, 139, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Zunino, M.P.; Newton, M.N.; Maestri, D.M.; Zygadlo, J.A. Essential oils of three Baccharis species. Planta Med. 1998, 64, 86–87. [Google Scholar] [CrossRef]
- De Bona, C.M.; Biasi, L.A.; Zanette, F.; Nakashima, T. Estaquia de três espécies de Baccharis. Ciênc. Rural 2005, 35, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Letchamo, W.; Xu, H.L.; Gosselin, A. Variations in photosynthesis and essential oil in thyme. J. Plant Physiol. 1995, 147, 29–37. [Google Scholar] [CrossRef]
- Bettaieb, I.; Zakhama, N.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Sci. Hortic. 2009, 120, 271–275. [Google Scholar] [CrossRef]
- Rioba, N.B.; Itulya, F.M.; Saidi, M.; Dudai, N.; Bernstein, N. Effects of nitrogen, phosphorus and irrigation frequency on essential oil content and composition of sage (Salvia officinalis L.). J. Appl. Res. Med. Aromat. Plants 2015, 2, 21–29. [Google Scholar] [CrossRef]
- Polanía Perdomo, J.A.; Rivera, M.; Ricaurte, J.; Rao, I.M. Phenotyping Common Beans for Adaptation to Drought: Protocol for Greenhouse Evaluation; International Center for Tropical Agriculture: Cali, Colombia, 2012. [Google Scholar]
- Putievsky, E.; Ravid, U.; Dudai, N. The influence of season and harvest frequency on essential oil and herbal yields from a pure clone of sage (Salvia officinalis) grown under cultivated conditions. J. Nat. Prod. 1986, 49, 326–329. [Google Scholar] [CrossRef]
- Verma, R.S.; Padalia, R.C.; Chauhan, A.; Upadhyay, R.K.; Singh, V.R. Productivity and essential oil composition of rosemary (Rosmarinus officinalis L.) harvested at different growth stages under the subtropical region of north India. Essent. Oil Res. 2019, 32, 144–149. [Google Scholar] [CrossRef]
- Kaltsa, O.; Grigorakis, S.; Lakka, A.; Bozinou, E.; Lalas, S.; Makris, D.P. Green Valorization of Olive Leaves to Produce Polyphenol-Enriched Extracts Using an Environmentally Benign Deep Eutectic Solvent. AgriEngineering 2020, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Giannoulis, K.D.; Kamvoukou, C.-A.; Gougoulias, N.; Wogiatzi, E. Irrigation and nitrogen application affect Greek oregano (Origanum vulgare ssp. hirtum) dry biomass, essential oil yield and composition. Ind. Crops Prod. 2020, 150, 112392. [Google Scholar] [CrossRef]
- Minteguiaga, M.; González, A.; Cassel, E.; Umpierrez, N.; Fariña, L.; Dellacassa, E. Volatile constituents from Baccharis spp. L. (Asteraceae): Chemical support for the conservation of threatened Species in Uruguay. Chem. Biodivers. 2018, 15, e1800017. [Google Scholar] [CrossRef]
- Embrapa Informação Tecnológica. Sistema Brasileiro de Classificação de Solos. Available online: www.cnps.embrapa.br/sac (accessed on 9 May 2016).
- United States Department of Agriculture NRCS; Soil Survey Staff. GNATSGO (Gridded National Soil Survey Geographic). Available online: https://nrcs.app.box.com/v/soils (accessed on 20 February 2020).
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper; FAO: Rome, Italy, 1998; Volume 300, p. D05109. [Google Scholar]
- Asensio, C.M.; Grosso, N.R.; Juliani, H.R. Quality preservation of organic cottage cheese using oregano essential oils. LWT Food Sci. Technol. 2015, 60, 664–671. [Google Scholar] [CrossRef]
- Clevenger, J.F. Apparatus for volatile oil determination, Description of New Type. Am. Perfum. Essent. Oil Rev. 1928, 17, 467–503. [Google Scholar]
- Ferhat, M.A.; Meklati, B.Y.; Smadja, J.; Chemat, F. An improved microwave Clevenger apparatus for distillation of essential oils from orange peel. J. Chromatogr. A 2006, 1112, 121–126. [Google Scholar] [CrossRef]
- Gavahian, M.; Farahnaky, A.; Javidnia, K.; Majzoobi, M. Comparison of ohmic-assisted hydrodistillation with traditional hydrodistillation for the extraction of essential oils from Thymus vulgaris L. Innov. Food Sci. Emerg. Technol. 2012, 14, 85–91. [Google Scholar] [CrossRef]
- El Gendy, A.; Leonardi, M.; Mugnaini, L.; Bertelloni, F.; Ebani, V.; Nardoni, S.; Mancianti, F.; Hendawy, S.; Omer, E.; Pistelli, L. Chemical composition and antimicrobial activity of essential oil of wild and cultivated Origanum syriacum plants grown in Sinai, Egypt. Ind. Crop. Prod. 2015, 67, 201–207. [Google Scholar] [CrossRef]
- Cazella, L.N.; Glamoclija, J.; Soković, M.; Gonçalves, J.E.; Linde, G.A.; Colauto, N.B.; Gazim, Z.C. Antimicrobial activity of essential oil of Baccharis dracunculifolia DC. (Asteraceae) aerial parts at flowering period. Front. Plant Sci. 2019, 10, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, F.; Torres, P.; Oomah, B.D.; De Alencar, S.M.; Massarioli, A.P.; Martin-Venegas, R.; Albarral-Ávila, V.; Burgos-Díaz, C.; Ferrer, R.; Rubilar, M. Volatile and non-volatile/semi-volatile compounds and in vitro bioactive properties of Chilean Ulmo (Eucryphia cordifolia Cav.) honey. Food Res. Int. 2017, 94, 20–28. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.r-project.org (accessed on 29 February 2020).
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Boil. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Fachinello, J.C.; da Silveira Pasa, M.; Schmtiz, J.D.; Betemps, D.L. Situação e perspectivas da fruticultura de clima temperado no Brasil. Rev. Bras. Frutic. 2011, 33, 109–120. [Google Scholar] [CrossRef]
- Andrião, M.A. Marcha de absorção de macronutrientes e acúmulo de fenólicos totais em [(Baccharis trimera (Less.) DC.] var. CPQBA-1, Sob Diferentes Podas no Plantio. Master’s Thesis, Universidade Estadual Paulista “Julio de Mesquita Filho” (UNESP), Botucatu, Brazil, 2010. [Google Scholar]
- Burmeister, J.; Guttenberg, H.V. Die ätherischen öle als produkt eines partiell anaeroben stoffwechsels. Planta Med. 1960, 8, 1–33. [Google Scholar] [CrossRef]
- García, D. Desenvolvimento, Rendimento, Teor e Componentes do Óleo Essencial em Função de Adubação Orgânica e Rebroto de Baccharis trimera (Less.) DC. cv. CPQBA-1. Master’s Thesis, Universidade Estadual Paulista “Julio de Mesquita Filho” (UNESP), Botucatu, Brazil, 2013. [Google Scholar]
- Palácio, C.P.A.M.; Biasi, L.A.; Nakashima, T.; Serrat, B.M. Biomassa e óleo essencial de carqueja (Baccharis trimera (Less.) DC.) sob influência de fontes e doses de nitrogênio. Rev. Bras. Plantas. Med. 2007, 9, 58–63. [Google Scholar]
- Budel, J.M.; Duarte, M.R.; Santos, C.A.M. Stem morpho-anatomy of Baccharis cylindrica (Less) DC. (Asteraceae). Rev. Bras. Ciênc. Farm. 2004, 40, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Bogo, C.A.; Andrade, M.H.; Paula, J.P.; Farago, P.V.; Döll-Boscardin, P.M.; Budel, J.M. Comparative analysis of essential oils of Baccharis L.: A review. Rev. Strict. Sensu. 2016, 1, 1–11. [Google Scholar] [CrossRef]
- Budel, J.M.; Wang, M.; Raman, V.; Zhao, J.; Khan, S.I.; Rehman, J.U.; Techen, N.; Tekwani, B.; Monteiro, L.M.; Heiden, G.; et al. Essential oils of five baccharis species: Investigations on the chemical composition and biological activities. Molecules 2018, 23, 2620. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, N.; Thiesen, F.V.; Alice, C.B. Aspectos farmacognósticos e perfil cromatográfico dos constituintes de Bacharis articulata Lam. Pers. Compositae. Cad. Farm. Porto Alegre RS 1988, 4, 63–67. [Google Scholar]
- Fachinetto, J.M.; Tedesco, S.B. Atividade antiproliferativa e mutagênica dos extratos aquosos de Baccharis trimera (Less.) AP de Candolle e Baccharis articulata (Lam.) Pers.(Asteraceae) sobre o sistema teste de Allium cepa. Rev. Bras. Plantas Med. 2009, 11, 360–367. [Google Scholar] [CrossRef]
- Tischer, B.; Vendruscolo, R.G.; Wagner, R.; Menezes, C.R.; Giacomelli, S.R.; Budel, J.M.; Barin, C.S.; Barin, J.S. Effect of grinding method on the analysis of essential oil from Baccharis articulata (Lam). Pers. Chem. Pap. 2017, 71, 753–761. [Google Scholar] [CrossRef]
- Zribi, I.; Bleton, J.; Moussa, F.; Abderrabba, M. GC-MS analysis of the volatile profile and the essential oil compositions of Tunisian Borago Officinalis L.: Regional locality and organ dependency. Ind. Crop. Prod. 2019, 129, 290–298. [Google Scholar] [CrossRef]
- Amiri, R.; Nikbakht, A.; Rahimmalek, M.; Hosseini, H. Variation in the Essential Oil Composition, Antioxidant Capacity, and Physiological Characteristics of Pelargonium graveolens L. Inoculated with Two Species of Mycorrhizal Fungi Under Water Deficit Conditions. J. Plant Growth Regul. 2017, 36, 502–515. [Google Scholar] [CrossRef]
- Xavier, V.; Vargas, R.; Cassel, E.; Lucas, A.; Santos, M.; Mondin, C.; Santarém, E.; Astarita, L.V.; Sartor, T. Mathematical modeling for extraction of essential oil from Baccharis spp. by steam distillation. Ind. Crop. Prod. 2011, 33, 599–604. [Google Scholar] [CrossRef]
- Rahman, A.; Shanta, Z.S.; Rashid, M.; Parvin, T.; Afrin, S.; Khatun, M.K.; Sattar, M. In vitro antibacterial properties of essential oil and organic extracts of Premna integrifolia Linn. Arab. J. Chem. 2016, 9, S475–S479. [Google Scholar] [CrossRef] [Green Version]
- Morais, L.A.S.; Castanha, R.F. Composição química do óleo essencial de duas amostras de carqueja (Baccharis sp.) coletadas em Paty do Alferes—Rio de Janeiro. Rev. Bras. Plantas Med. 2011, 13, 628–632. [Google Scholar] [CrossRef]
- Costa, D.P.; Santos, S.C.; Seraphin, J.C.; Ferri, P.H. Seasonal variability of essential oils of eugenia uniflora leaves. J. Braz. Chem. Soc. 2009, 20, 1287–1293. [Google Scholar] [CrossRef]
- Sartoratto, A.; Machado, A.L.M.; Delarmelina, C.; Figueira, G.M.; Duarte, M.C.T.; Rehder, V.L.G. Composition and Antimicrobial Activity of Essential Oils From Aromatic Plants Used in Brazil. Braz. J. Microbiol. 2004, 35, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, F.F.G.; Oliveira, L.G.S.; Rodrigues, F.F.G.; Saraiva, M.E.; Almeida, S.C.X.; Cabral, M.E.S.; Campos, A.R.; Costa, J.G.M. Chemical composition, antibacterial and antifungal activities of essential oil from Cordia verbenacea DC. leaves. Pharmacogn. Res. 2012, 4, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, J.; Lukas, B.; Franz, C. Temperature influences thymol and carvacrol differentially in Origanum spp.(Lamiaceae). J. Essent. Oil Res. 2010, 22, 412–415. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Pizani, M.A.; Paredes-Trejo, F.J.; Farias-Ramirez, A.J.; dos Santos, H.T.; Prado Massarioli, A.; Marin, F.R.; Takeyoshi, B.Y.; Alves Marques, P.A.; De Stefano Piedade, S.M.; de Alencar, S.M. Essential Oil Content of Baccharis crispa Spreng. Regulated by Water Stress and Seasonal Variation. AgriEngineering 2020, 2, 458-470. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering2030031
Moreno-Pizani MA, Paredes-Trejo FJ, Farias-Ramirez AJ, dos Santos HT, Prado Massarioli A, Marin FR, Takeyoshi BY, Alves Marques PA, De Stefano Piedade SM, de Alencar SM. Essential Oil Content of Baccharis crispa Spreng. Regulated by Water Stress and Seasonal Variation. AgriEngineering. 2020; 2(3):458-470. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering2030031
Chicago/Turabian StyleMoreno-Pizani, Maria Alejandra, Franklin Javier Paredes-Trejo, Asdrubal Jesus Farias-Ramirez, Hugo Thaner dos Santos, Adna Prado Massarioli, Fabio Ricardo Marin, Bruno Yukio Takeyoshi, Patricia Angelica Alves Marques, Sônia Maria De Stefano Piedade, and Severino Matias de Alencar. 2020. "Essential Oil Content of Baccharis crispa Spreng. Regulated by Water Stress and Seasonal Variation" AgriEngineering 2, no. 3: 458-470. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering2030031