
The Future of Anticancer Drugs: A Cytotoxicity Assessment Study of CdSe/ZnS Quantum Dots



Abstract
:1. Introduction
2. Experimental Section
2.1. CdSe/ZnS Quantum Dots (QDs)
2.2. Culturing HeLa Cells and Non-Cancerous Mouse-Derived Fibroblast Cells
2.3. Cell Viability Assay
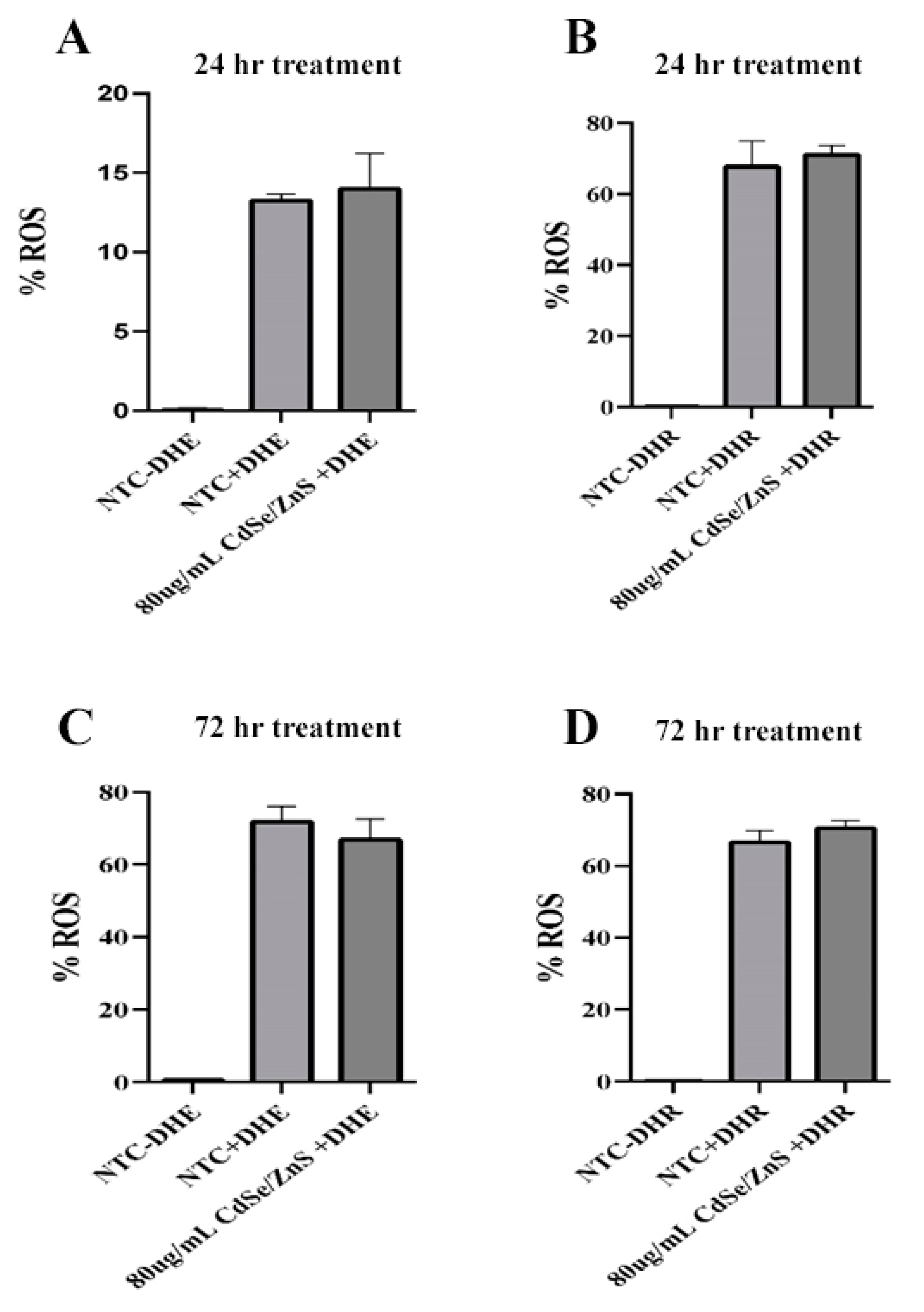
2.4. Reactive Oxygen Species (ROS) Assay
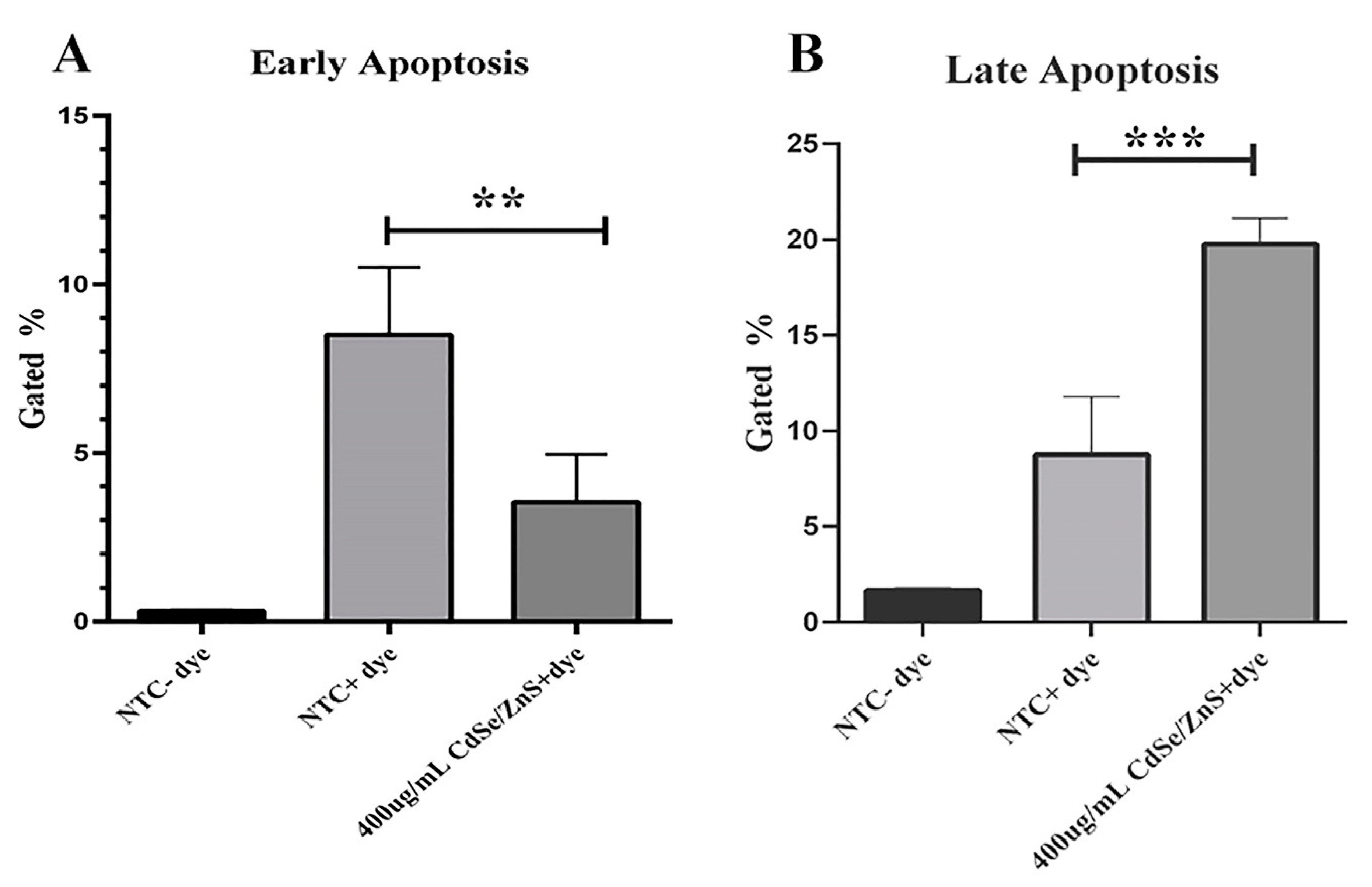
2.5. Apoptosis Assay
2.6. Total RNA Extraction, mRNA Isolation, and cDNA Synthesis
2.7. Basepair Transcriptome Analysis
2.8. Statistical Analysis
3. Results
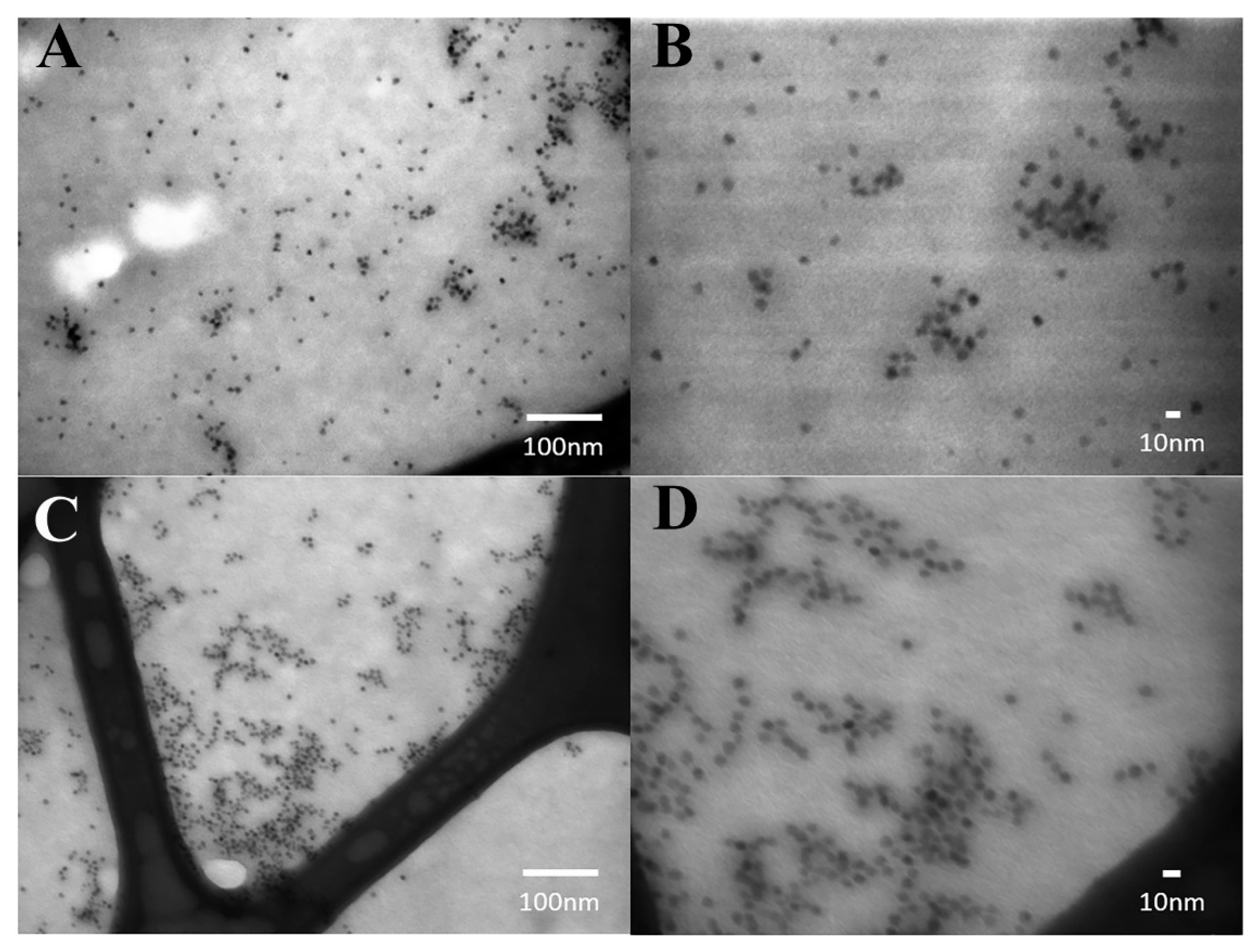
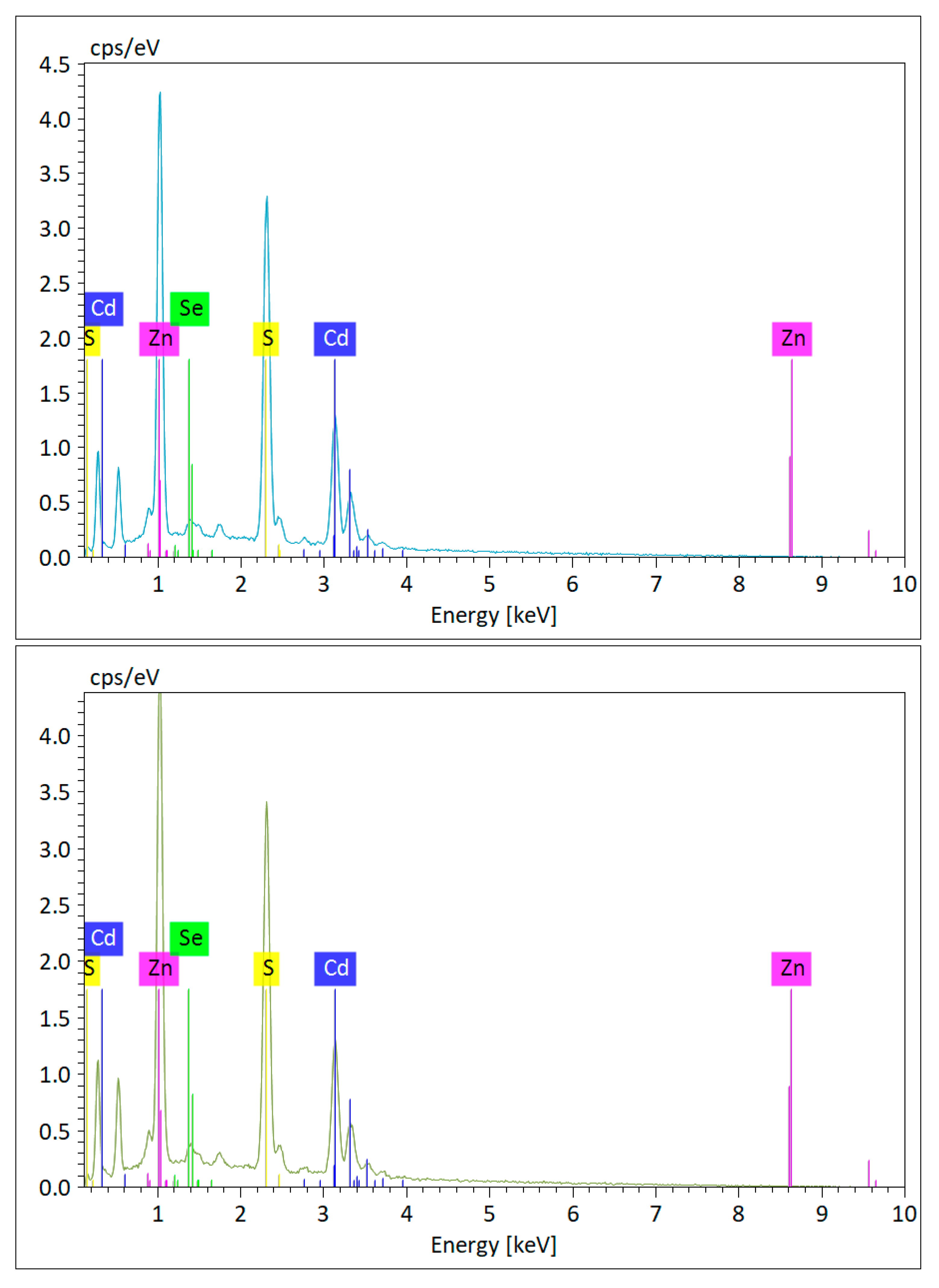
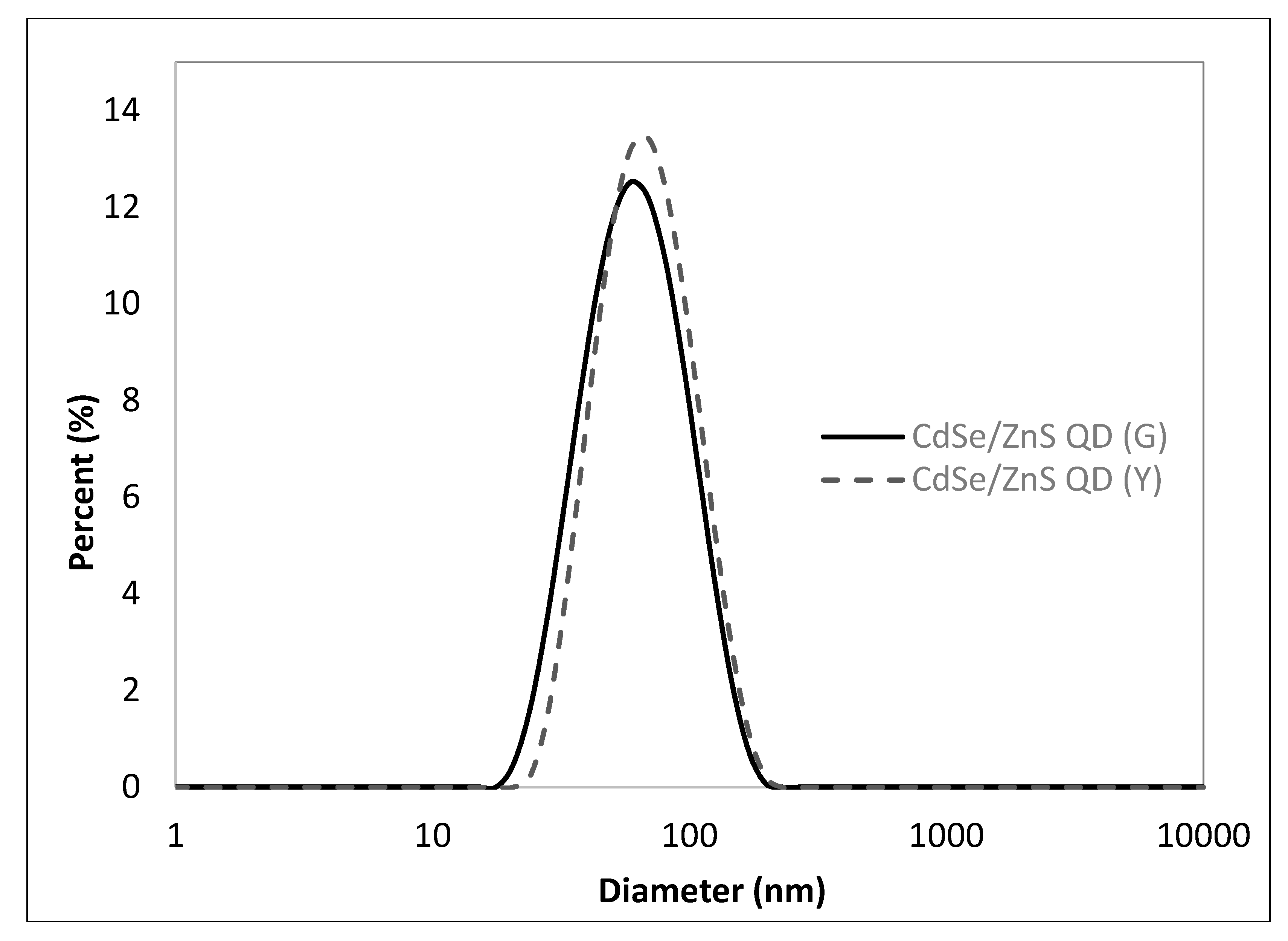
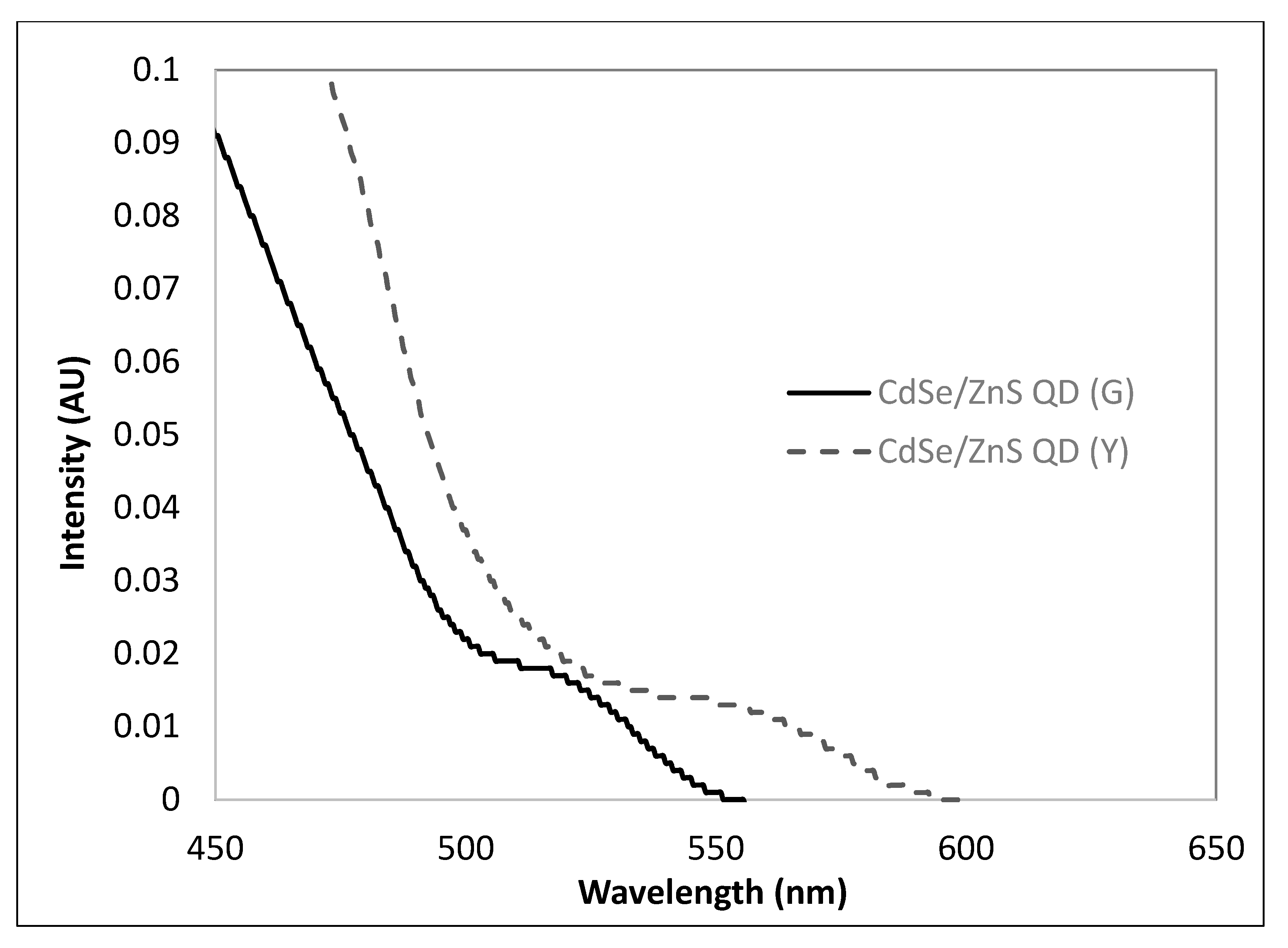
3.1. Chemophysical Properties of CdSe/ZnS QDs Used for This Study
3.2. Cell Viability Assay
3.3. ROS Assay
3.4. Apoptosis Assay
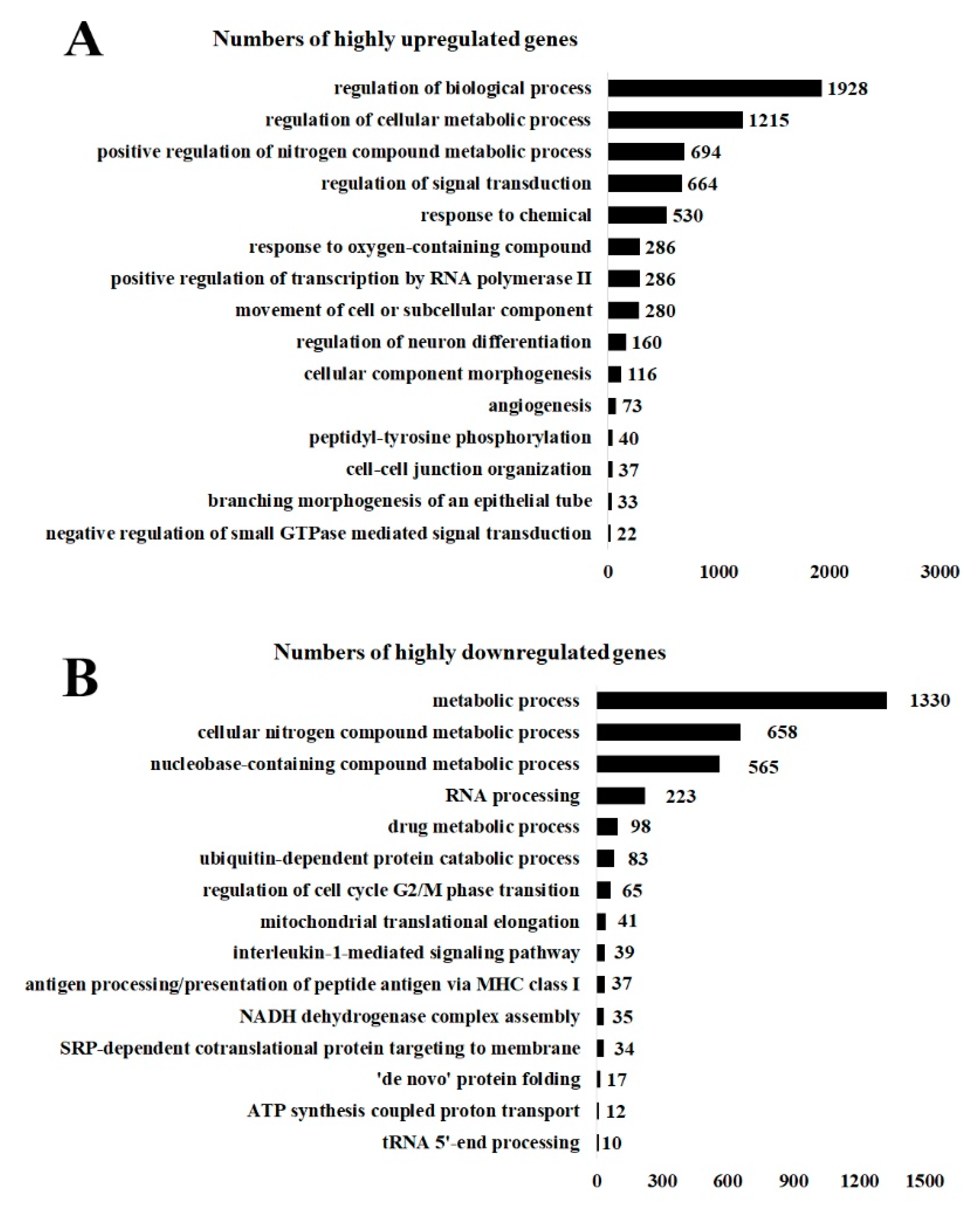
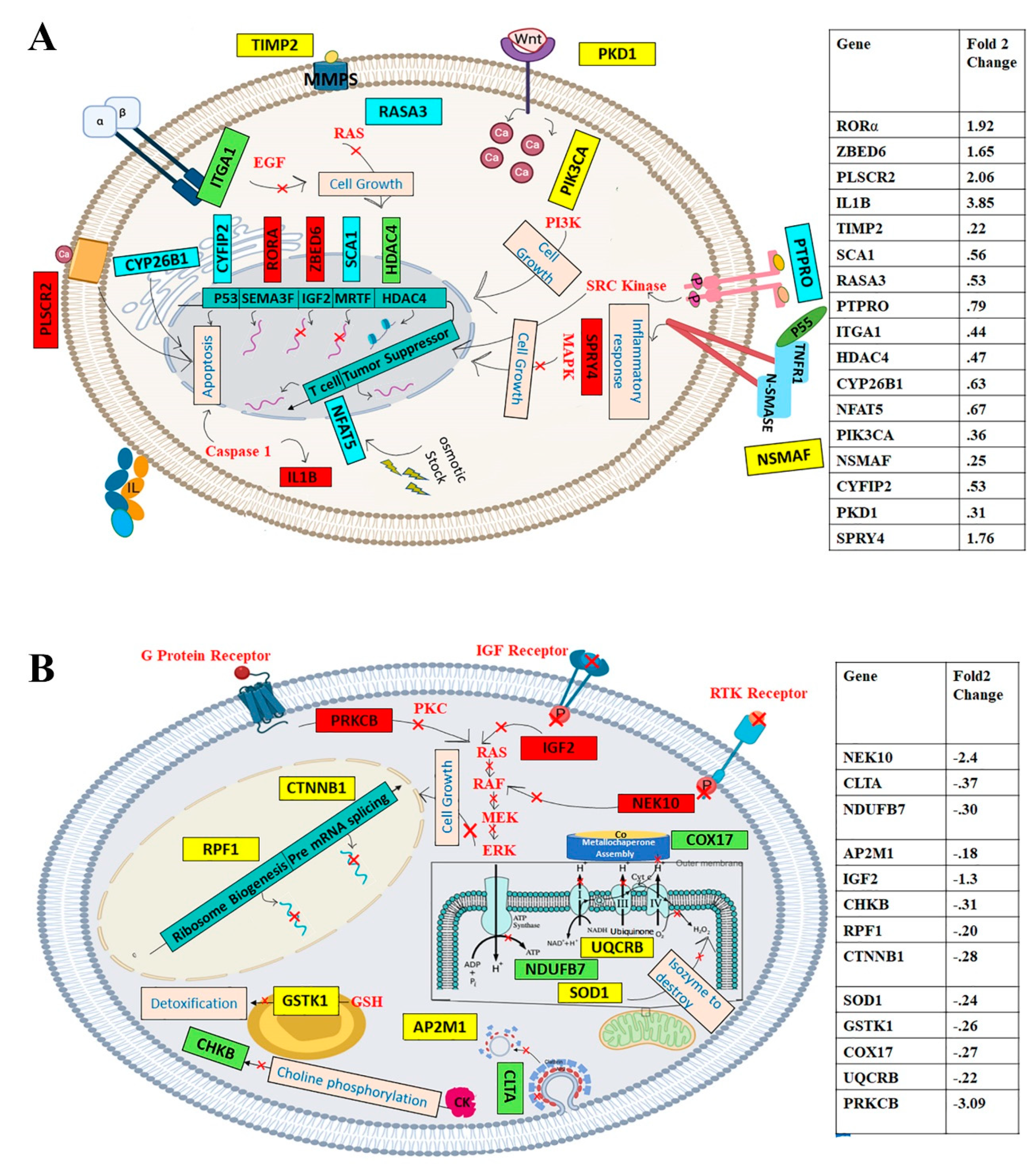
3.5. Upregulated and Downregulated Genes Analyzed by GoRilla: GoTerm
4. Discussion
4.1. Green CdSe/ZnS QDs Are More Effective in Reducing Viability of HeLa Cancer Cells than Yellow CdSe/ZnS QDs
4.2. Effects of Green CdSe/ZnS QDs on the Production of ROS
4.3. Green CdSe/ZnS and Its Impact on Apoptosis
4.4. Upregulated Gene Expression and the Subsequent Impacts upon Green CdSe/ZnS QD Treatment
4.5. Downregulated Cellular Functions Mediated by CdSe/ZnS QDs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gutiérrez-Hoya, A.; Zerecero-Carreón, O.; Valle-Mendiola, A.; Moreno-Lafont, M.; López-Santiago, R.; Weiss-Steider, B.; Soto-Cruz, I. Cervical Cancer Cells Express Markers Associated with Immunosurveillance. J. Immunol. Res. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- Dillner, J.; Rebolj, M.; Birembaut, P.; Petry, K.-U.; Szarewski, A.; Munk, C.; de Sanjose, S.; Naucler, P.; Lloveras, B.; Kjaer, S.; et al. Long term predictive values of cytology and human papillomavirus testing in cervical cancer screening: Joint European cohort study. BMJ 2008, 337, a1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucey, B.P.; Nelson-Rees, W.A.; Hutchins, G.M. Henrietta Lacks, HeLa Cells, and Cell Culture Contamination. Arch. Pathol. Lab. Med. 2009, 133, 1463–1467. [Google Scholar]
- Puck, T.T.; Marcus, P.I. A rapid method for viable cell titration and clone production with HeLa cells in tissue culture: The use of X-irradiated cells to supply conditioning factors. Proc. Natl. Acad. Sci. USA 1955, 41, 432–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiang, Y.; Hertzberg, R.; Hecht, S.; Liu, L. Camptothecin induces protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem. 1985, 260, 14873–14878. [Google Scholar]
- Blackburn, E.H. Telomere states and cell fates. Nature 2000, 408, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, I.; Gonel, A.; Durgun, M.; Kocyigit, A.; Yuksekdag, O.; Supuran, C.T. Assessment of the antiproliferative and apoptotic roles of sulfonamide carbonic anhydrase IX inhibitors in HeLa cancer cell line. J. Enzyme Inhib. Med. Chem. 2019, 34, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakkarainen, T.; Schramm, A.; Viheriälä, J.; Guina, M. Site-Controlled Epitaxy of InAs Quantum Dots on Nanoimprint Lithography Patterns. In Molecular Beam Epitaxy; Elsevier: Amsterdam, The Netherlands, 2018; pp. 277–292. [Google Scholar]
- Ekimov, A.I.; Efros, A.L.; Onushchenko, A.A. Quantum size effect in semiconductor microcrystals. Solid State Commun. 1985, 56, 921–924. [Google Scholar] [CrossRef]
- Lin, L.Y.; Al-Jamal, W.T.; Kostarelos, K. Quantum-Dot Toxicity. In Encyclopedia of Nanotechnology; Bhushan, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 2197–2200. [Google Scholar]
- Mahajan, S.; Rani, M.; Dubey, R.; Mahajan, J. Characteristics and properties of CdSe quantum dots. Int. J. Latest Res. Sci. Technol. 2013, 2, 518–521. [Google Scholar]
- Su, Y.; He, Y.; Lu, H.; Sai, L.; Li, Q.; Li, W.; Wang, L.; Shen, P.; Huang, Q.; Fan, C. The cytotoxicity of cadmium based, aqueous phase—Synthesized, quantum dots and its modulation by surface coating. Biomaterials 2009, 30, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, W.; Zhang, J.; Liu, J.; Chen, G.; Pope, C. In vitro and in vivo toxicity of CdTe nanoparticles. J. Nanosci. Nanotechnol. 2007, 7, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Ryman-Rasmussen, J.P.; Riviere, J.E.; Monteiro-Riviere, N.A. Penetration of Intact Skin by Quantum Dots with Diverse Physicochemical Properties. Toxicol. Sci. 2006, 91, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Liu, W.; Liang, J.; He, Z.; Xu, H.; Yang, X. Probing the cytotoxicity of CdSe quantum dots with surface modification. Mater. Lett. 2007, 61, 1641–1644. [Google Scholar] [CrossRef]
- Wang, L.; Nagesha, D.K.; Selvarasah, S.; Dokmeci, M.R.; Carrier, R.L. Toxicity of CdSe Nanoparticles in Caco-2 Cell Cultures. J. Nanobiotechnol. 2008, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Xing, T.; Zeng, J.; Wang, H.; Li, C.; Yin, S.; Yan, D.; Deng, H.; Liu, J.; Wang, M.; et al. Unmodified CdSe Quantum Dots Induce Elevation of Cytoplasmic Calcium Levels and Impairment of Functional Properties of Sodium Channels in Rat Primary Cultured Hippocampal Neurons. Environ. Health Perspect. 2008, 116, 915–922. [Google Scholar] [CrossRef]
- Mansuriya, B.D.; Altintas, Z. Applications of Graphene Quantum Dots in Biomedical Sensors. Sensors 2020, 20, 1072. [Google Scholar] [CrossRef] [Green Version]
- Fang, M.; Peng, C.; Pang, D.-W.; Li, Y. Quantum Dots for Cancer Research: Current Status, Remaining Issues, and Future Perspectives. Cancer Biol. Med. 2012, 9, 151–163. [Google Scholar]
- Liu, H.; Tang, W.; Li, C.; Lv, P.; Wang, Z.; Liu, Y.; Zhang, C.; Bao, Y.; Chen, H.; Meng, X.; et al. CdSe/ZnS Quantum Dots-Labeled Mesenchymal Stem Cells for Targeted Fluorescence Imaging of Pancreas Tissues and Therapy of Type 1 Diabetic Rats. Nanoscale Res. Lett. 2015, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Bharti, S.; Kaur Bhullar, G.; Tripathi, S.K. pH dependent drug release from drug conjugated PEGylated CdSe/ZnS nanoparticles. Mater. Chem. Phys. 2020, 240, 122162. [Google Scholar] [CrossRef]
- Kaviyarasu, K.; Kanimozhi, K.; Matinise, N.; Maria Magdalane, C.; Mola, G.T.; Kennedy, J.; Maaza, M. Antiproliferative effects on human lung cell lines A549 activity of cadmium selenide nanoparticles extracted from cytotoxic effects: Investigation of bio-electronic application. Mater. Sci. Eng. C 2017, 76, 1012–1025. [Google Scholar] [CrossRef] [PubMed]
- Balaiya, S.; Chalam, K. An In vitro Assay to Quantify Nitrosative Component of Oxidative Stress. J. Mol. Genet. Med. 2014, 8, 120. [Google Scholar] [PubMed] [Green Version]
- Lovrić, J.; Bazzi, H.S.; Cuie, Y.; Fortin, G.R.A.; Winnik, F.M.; Maysinger, D. Differences in subcellular distribution and toxicity of green and red emitting CdTe quantum dots. J. Mol. Med. 2005, 83, 377–385. [Google Scholar] [CrossRef]
- Horstmann, C.; Campbell, C.; Kim, D.S.; Kim, K. Transcriptome profile with 20 nm silver nanoparticles in yeast. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naderi, S.; Zare, H.; Taghavinia, N.; Irajizad, A.; Aghaei, M.; Panjehpour, M. Cadmium telluride quantum dots induce apoptosis in human breast cancer cell lines. Toxicol. Ind. Health 2018, 34, 339–352. [Google Scholar] [CrossRef]
- He, S.-J.; Cao, J.; Li, Y.-S.; Yang, J.-C.; Zhou, M.; Qu, C.-Y.; Zhang, Y.; Shen, F.; Chen, Y.; Li, M.-M.; et al. CdSe/ZnS quantum dots induce photodynamic effects and cytotoxicity in pancreatic cancer cells. World J. Gastroenterol. 2016, 22, 5012. [Google Scholar] [CrossRef]
- Lim, S.D.; Sun, C.; Lambeth, J.D.; Marshall, F.; Amin, M.; Chung, L.; Petros, J.A.; Arnold, R.S. Increased Nox1 and hydrogen peroxide in prostate cancer. Prostate 2005, 62, 200–207. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.; et al. SEER Cancer Statistics Review; National Cancer Institute: Bethesda, MD, USA, 2019.
- Zhang, L.W.; Bäumer, W.; Monteiro-Riviere, N.A. Cellular uptake mechanisms and toxicity of quantum dots in dendritic cells. Nanomedicine 2011, 6, 777–791. [Google Scholar] [CrossRef] [Green Version]
- Pathakoti, K.; Hwang, H.-M.; Xu, H.; Aguilar, Z.P.; Wang, A. In vitro cytotoxicity of CdSe/ZnS quantum dots with different surface coatings to human keratinocytes HaCaT cells. J. Environ. Sci. 2013, 25, 163–171. [Google Scholar] [CrossRef]
- Derfus, A.M.; Chan, W.C.W.; Bhatia, S.N. Probing the Cytotoxicity of Semiconductor Quantum Dots. Nano Lett. 2004, 4, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, C.; Liedl, T.; Kudera, S.; Pellegrino, T.; Muñoz Javier, A.; Gaub, H.E.; Stölzle, S.; Fertig, N.; Parak, W.J. Cytotoxicity of Colloidal CdSe and CdSe/ZnS Nanoparticles. Nano Lett. 2005, 5, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Wang, C.; Peng, F.; Wang, R.; Xiao, X.; Zeng, J.; Kang, H.; Fan, X.; Sha, L.; et al. Transcriptomic Profiles Reveal the Interactions of Cd/Zn in Dwarf Polish Wheat (Triticum polonicum L.) Roots. Front. Physiol. 2017, 8, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnik, F.M.; Maysinger, D. Quantum Dot Cytotoxicity and Ways to Reduce It. Acc. Chem. Res. 2013, 46, 672–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradburne, C.E.; Delehanty, J.B.; Boeneman Gemmill, K.; Mei, B.C.; Mattoussi, H.; Susumu, K.; Blanco-Canosa, J.B.; Dawson, P.E.; Medintz, I.L. Cytotoxicity of Quantum Dots Used for In Vitro Cellular Labeling: Role of QD Surface Ligand, Delivery Modality, Cell Type, and Direct Comparison to Organic Fluorophores. Bioconjug. Chem. 2013, 24, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Steinbrück, A.; Gao, J.; Doggett, N.; Hollingsworth, J.A.; Iyer, R. Comprehensive Analysis of the Effects of CdSe Quantum Dot Size, Surface Charge, and Functionalization on Primary Human Lung Cells. ACS Nano 2012, 6, 4748–4762. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.-T.; Li, X.; Wang, Q.-S.; Zhang, Z.-J.; Liu, P.; Zhang, C.-C. Toxicity evaluation of CdTe quantum dots with different size on Escherichia coli. Toxicology In Vitro 2012, 26, 1233–1239. [Google Scholar] [CrossRef]
- Manshian, B.B.; Soenen, S.J.; Al-Ali, A.; Brown, A.; Hondow, N.; Wills, J.; Jenkins, G.J.S.; Doak, S.H. Cell Type-Dependent Changes in CdSe/ZnS Quantum Dot Uptake and Toxic Endpoints. Toxicol. Sci. 2015, 144, 246–258. [Google Scholar] [CrossRef]
- Dimayuga, F.O.; Wang, C.; Clark, J.M.; Dimayuga, E.R.; Dimayuga, V.M.; Bruce-Keller, A.J. SOD1 overexpression alters ROS production and reduces neurotoxic inflammatory signaling in microglial cells. J. Neuroimmunol. 2007, 182, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Remacle, C.; Coosemans, N.; Jans, F.; Hanikenne, M.; Motte, P.; Cardol, P. Knock-down of the COX3 and COX17 gene expression of cytochrome c oxidase in the unicellular green alga Chlamydomonas reinhardtii. Plant Mol. Biol. 2010, 74, 223–233. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Y.; Kong, L.; Xue, Y.; Tang, M. Threshold Dose of Three Types of Quantum Dots (QDs) Induces Oxidative Stress Triggers DNA Damage and Apoptosis in Mouse Fibroblast L929 Cells. Int. J. Environ. Res. Public Health 2015, 12, 13435–13454. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Tang, M.; Kong, L.; Ying, J.; Wu, T.; Xue, Y.; Pu, Y. Liver Toxicity of Cadmium Telluride Quantum Dots (CdTe QDs) Due to Oxidative Stress in Vitro and in Vivo. Int. J. Mol. Sci. 2015, 16, 23279–23299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Gao, M.-H.; Zhang, X.-C.; Chu, X.-M. Molecular immune mechanism of C-phycocyanin from Spirulina platensis induces apoptosis in HeLa cells in vitro. Biotechnol. Appl. Biochem. 2006, 43, 155. [Google Scholar] [PubMed]
- Liu, C.C.; Wang, L.V.; Zhou, M.; Ye, J.; Chan, G.C.; Yang, M. Tashinone IIA Can Trigger the Apoptosis of a Megakaryocytic Cell Line CHRF- 288 through the TNFRSF9 Mediated Caspase-3 Activation Pathway. Blood 2008, 112, 4729. [Google Scholar] [CrossRef]
- Ruiz-Vela, A.; Opferman, J.T.; Cheng, E.H.; Korsmeyer, S.J. Proapoptotic BAX and BAK control multiple initiator caspases. EMBO Rep. 2005, 6, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nafisi, H.; Banihashemi, B.; Daigle, M.; Albert, P.R. GAP1(IP4BP)/RASA3 mediates Galphai-induced inhibition of mitogen-activated protein kinase. J. Biol. Chem. 2008, 283, 35908–35917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattila, E.; Pellinen, T.; Nevo, J.; Vuoriluoto, K.; Arjonen, A.; Ivaska, J. Negative regulation of EGFR signalling through integrin-α1β1-mediated activation of protein tyrosine phosphatase TCPTP. Nat. Cell Biol. 2005, 7, 78–85. [Google Scholar] [CrossRef]
- Akhtar Ali, M.; Younis, S.; Wallerman, O.; Gupta, R.; Andersson, L.; Sjöblom, T. Transcriptional modulator ZBED6 affects cell cycle and growth of human colorectal cancer cells. Proc. Natl. Acad. Sci. USA 2015, 112, 7743–7748. [Google Scholar] [CrossRef] [Green Version]
- Motiwala, T.; Kutay, H.; Ghoshal, K.; Bai, S.; Seimiya, H.; Tsuruo, T.; Suster, S.; Morrison, C.; Jacob, S.T. Protein tyrosine phosphatase receptor-type O (PTPRO) exhibits characteristics of a candidate tumor suppressor in human lung cancer. Proc. Natl. Acad. Sci. USA 2004, 101, 13844–13849. [Google Scholar] [CrossRef] [Green Version]
- Holditch, S.J.; Brown, C.N.; Lombardi, A.M.; Nguyen, K.N.; Edelstein, C.L. Recent Advances in Models, Mechanisms, Biomarkers, and Interventions in Cisplatin-Induced Acute Kidney Injury. Int. J. Mol. Sci. 2019, 20, 3011. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Xu, R. RORα, a Potential Tumor Suppressor and Therapeutic Target of Breast Cancer. Int. J. Mol. Sci. 2012, 13, 15755–15766. [Google Scholar] [CrossRef] [Green Version]
- Alsina, F.C.; Irala, D.; Fontanet, P.A.; Hita, F.J.; Ledda, F.; Paratcha, G. Sprouty4 Is an Endogenous Negative Modulator of TrkA Signaling and Neuronal Differentiation Induced by NGF. PLoS ONE 2012, 7, e32087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.; Li, N.; Cai, S.; Guo, H.; Wen, Y. Inhibition of CYFIP2 promotes gastric cancer cell proliferation and chemoresistance to 5-fluorouracil through activation of the Akt signaling pathway. Oncol. Lett. 2017, 13, 2133–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Yi, H.; Bao, Y.; Liu, Y.; Gao, X. Oestrogen inhibits PTPRO to prevent the apoptosis of renal podocytes. Exp. Ther. Med. 2019, 17, 2373–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montfort, A.; Martin, P.G.P.; Levade, T.; Benoist, H.; Ségui, B. FAN (factor associated with neutral sphingomyelinase activation), a moonlighting protein in TNF-R1 signaling. J. Leukoc. Biol. 2010, 88, 897–903. [Google Scholar] [CrossRef]
- Jackson II, R.S.; Cho, Y.-J.; Stein, S.; Liang, P. CYFIP2, a Direct p53 Target, is Leptomycin-B Sensitive. Cell Cycle 2007, 6, 95–103. [Google Scholar] [CrossRef]
- Su, J.; Su, B.; Xia, H.; Liu, F.; Zhao, X.; Li, J.; Zhang, J.; Shi, Y.; Zeng, Y.; Zeng, X.; et al. RORα Suppresses Epithelial-to-Mesenchymal Transition and Invasion in Human Gastric Cancer Cells via the Wnt/β-Catenin Pathway. Front. Oncol. 2019, 9, 1344. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y. Substantial reduction in risk of breast cancer associated with genetic polymorphisms in the promoters of the matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-2 genes. Carcinogenesis 2003, 25, 399–404. [Google Scholar] [CrossRef]
- Gao, X.; Nawaz, Z. Progesterone receptors-animal models and cell signaling in breast cancer: Role of steroid receptor coactivators and corepressors of progesterone receptors in breast cancer. Breast Cancer Res. 2002, 4, 182. [Google Scholar] [CrossRef]
- Saijo, K.; Mecklenbräuker, I.; Santana, A.; Leitger, M.; Schmedt, C.; Tarakhovsky, A. Protein kinase C beta controls nuclear factor kappaB activation in B cells through selective regulation of the IkappaB kinase alpha. J. Exp. Med. 2002, 195, 1647–1652. [Google Scholar] [CrossRef]
- Martins, A.S.; Olmos, D.; Missiaglia, E.; Shipley, J. Targeting the Insulin-Like Growth Factor Pathway in Rhabdomyosarcomas: Rationale and Future Perspectives. Sarcoma 2011, 2011, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Thomas, G.; Ghoussaini, M.; Healey, C.S.; Humphreys, M.K.; Platte, R.; Morrison, J.; Maranian, M.; Pooley, K.A.; Luben, R.; et al. Newly discovered breast cancer susceptibility loci on 3p24 and 17q23.2. Nat. Genet. 2009, 41, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Wang, J.; Yang, F.; Tao, Q.; Zhang, J.; Wang, L.; Yang, Y.; Liu, H.; Wang, Z.; Xu, Q.; et al. Long noncoding RNA DANCR increases stemness features of hepatocellular carcinoma by derepression of CTNNB1: HEPATOLOGY, Vol. XX, No. X, 2015. Hepatology 2016, 63, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-C.; Chang, J.; Lee, H.S.; Kwon, H.J. Mitochondrial UQCRB as a new molecular prognostic biomarker of human colorectal cancer. Exp. Mol. Med. 2017, 49, e391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimaki, M.; Wang, X.; McKenzie, M.; Thorburn, D.R.; Ryan, M.T. Understanding mitochondrial complex I assembly in health and disease. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 851–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Chen, M.; Wang, Z.; Zhao, G.; Liu, J.-X. Transcriptional profiles and copper stress responses in zebrafish cox17 mutants. Environ. Pollut. 2020, 256, 113364. [Google Scholar] [CrossRef] [PubMed]
- Whitson, J.A.; Zhang, X.; Medvedovic, M.; Chen, J.; Wei, Z.; Monnier, V.M.; Fan, X. Transcriptome of the GSH-Depleted Lens Reveals Changes in Detoxification and EMT Signaling Genes, Transport Systems, and Lipid Homeostasis. Investig. Opthalmol. Vis. Sci. 2017, 58, 2666. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Ontology Term | No. of Genes | Corresponding Highly Upregulated Genes |
|---|---|---|
| activation of phospholipase C activity | 2 | EGFR, GNA15, alpha 15 (gq class) |
| regulation of NIK/NF-kappaB signaling | 4 | EGFR; GPR97; IL1β; MIR30C2 |
| cellular response to tumor necrosis factor | 4 | KCNJ11; RORA; IL8; MIR30C2 |
| response to tumor necrosis factor | 4 | KCNJ11; RORA; IL8; MIR30C2 |
| signaling | 6 | HTR7; EREG; KCNJ4; PKP2; GDF15; IL1β |
| astrocyte activation | 2 | EGFR; IL1β |
| positive regulation of interleukin-6 biosynthetic process | 2 | EREG; IL1β |
| Gene Ontology Term | No. of Genes | Corresponding Highly Downregulated Genes |
|---|---|---|
| mesenchyme migration | 3 | ACTA1, ACTC1, FOXF1 |
| tissue migration | 3 | ACTA1, ACTC1, FOXF1 |
| skeletal muscle thin filament assembly | 2 | ACTA1, ACTC1 |
| epithelial cell differentiation involved in mammary gland alveolus development | 2 | ID2, FOXF1 |
| regulation of systemic arterial blood pressure by hormone | 2 | EDN2, NOX1 |
| endocrine process | 2 | EDN2, NOX1 |
| extracellular matrix organization | 5 | A2M, NOX1, JAM3, EMILIN1, FOXF1 |
| regulation of systemic arterial blood pressure mediated by a chemical signal | 2 | EDN2, NOX1 |
| embryonic digestive tract morphogenesis | 2 | ID2, FOXF1 |
| extracellular structure organization | 5 | A2M, NOX1, JAM3, EMILIN1, FOXF1 |
| regulation of blood pressure | 3 | ID2, EDN2, NOX1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hens, B.; Smothers, J.; Rizvanovic, H.; Patel, R.; Wu, Q.; Kim, K. The Future of Anticancer Drugs: A Cytotoxicity Assessment Study of CdSe/ZnS Quantum Dots. J. Nanotheranostics 2020, 1, 19-38. https://0-doi-org.brum.beds.ac.uk/10.3390/jnt1010003
Hens B, Smothers J, Rizvanovic H, Patel R, Wu Q, Kim K. The Future of Anticancer Drugs: A Cytotoxicity Assessment Study of CdSe/ZnS Quantum Dots. Journal of Nanotheranostics. 2020; 1(1):19-38. https://0-doi-org.brum.beds.ac.uk/10.3390/jnt1010003
Chicago/Turabian StyleHens, Basant, Jared Smothers, Husref Rizvanovic, Rishi Patel, Qihua Wu, and Kyoungtae Kim. 2020. "The Future of Anticancer Drugs: A Cytotoxicity Assessment Study of CdSe/ZnS Quantum Dots" Journal of Nanotheranostics 1, no. 1: 19-38. https://0-doi-org.brum.beds.ac.uk/10.3390/jnt1010003