Changes to the Oligosaccharide Profile of Bovine Milk at the Onset of Lactation

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design and Sample Collection
2.3. Oligosaccharide Analysis
2.4. Statistical Analysis
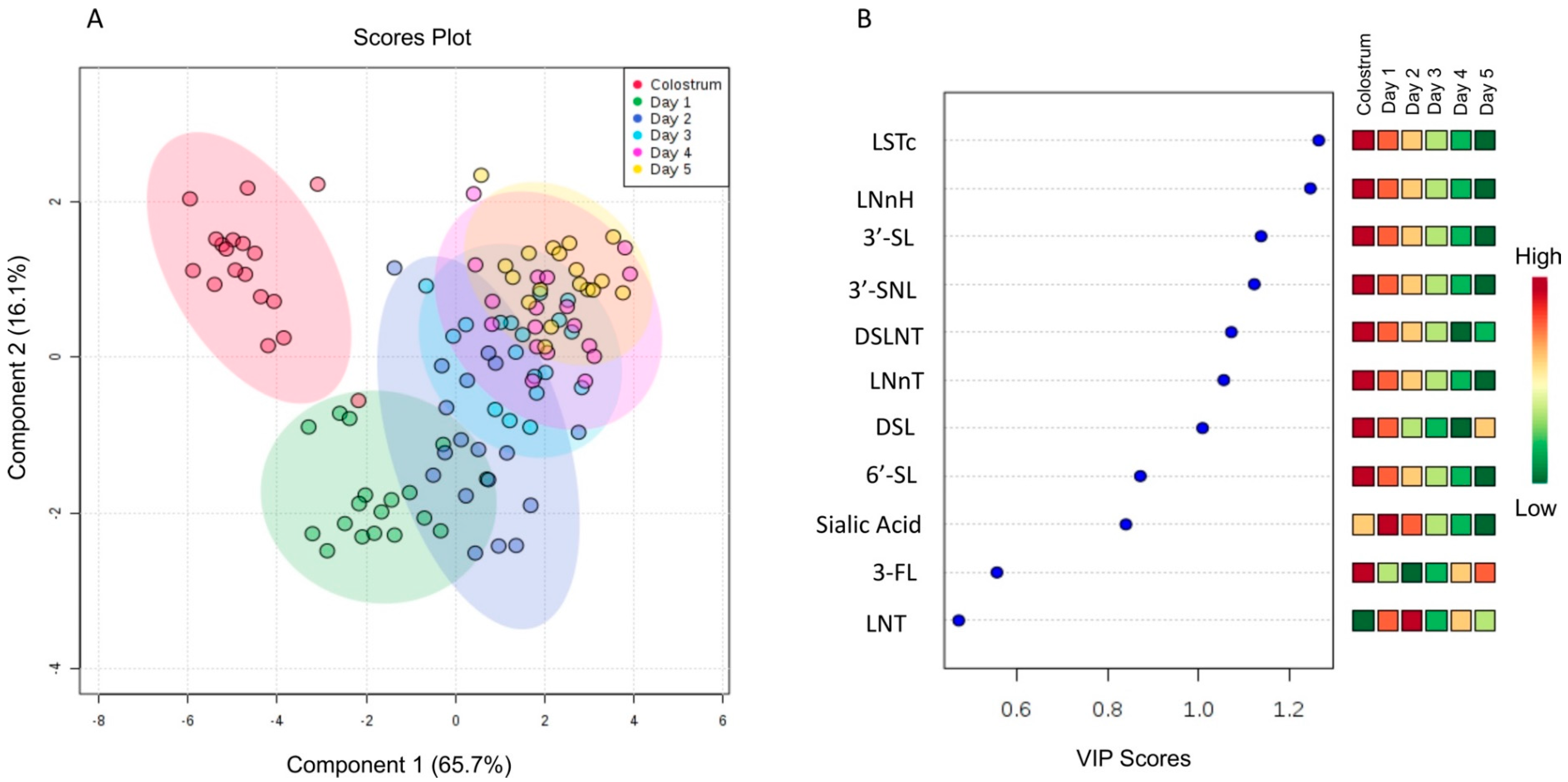
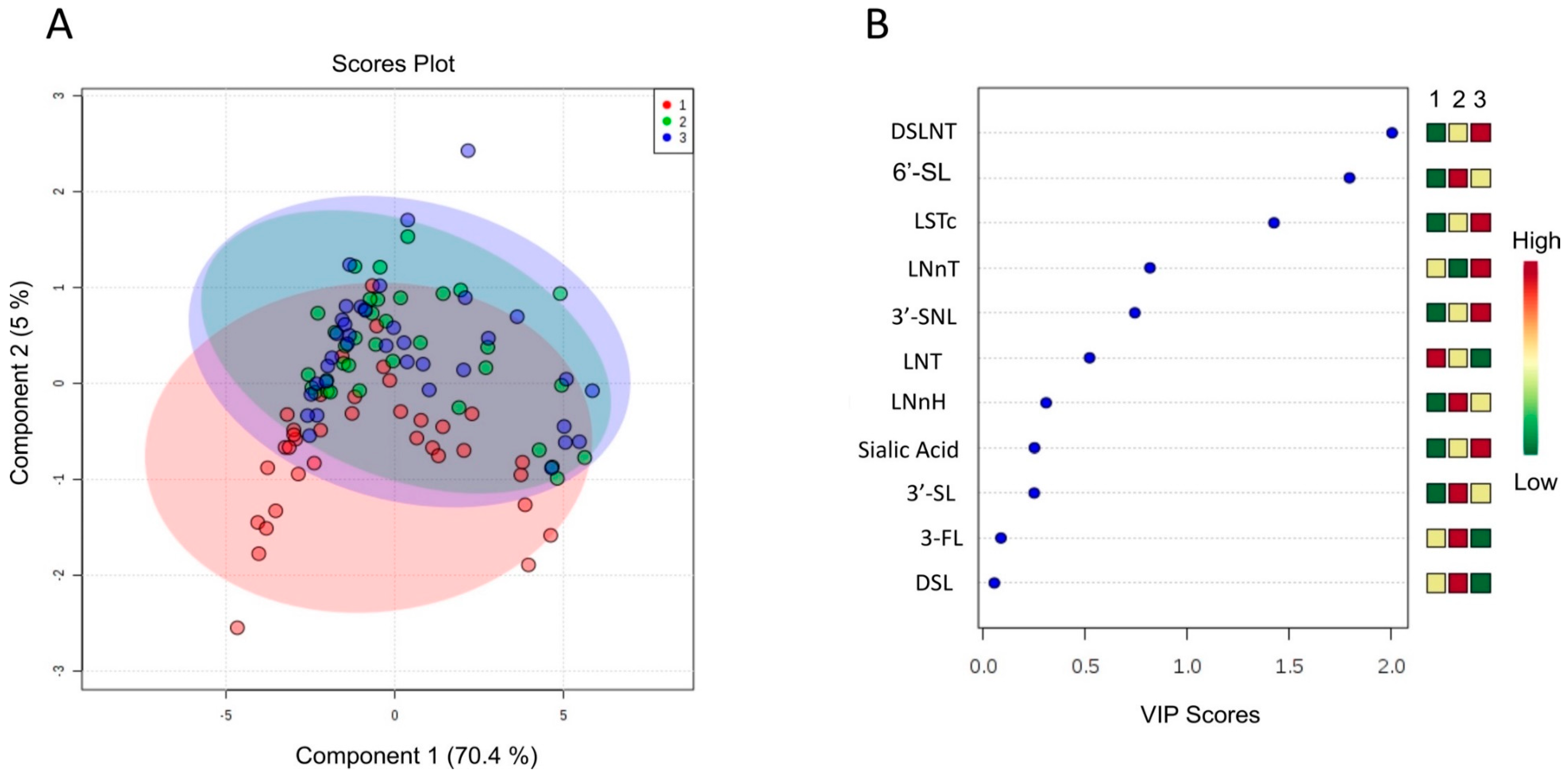
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kunz, C.; Rudloff, S.; Baier, W.; Klein, N.; Strobel, S. Oligosaccharides in Human Milk: Structural, Functional, and Metabolic Aspects. Annu. Rev. Nutr. 2000, 20, 699–722. [Google Scholar] [CrossRef] [PubMed]
- Quinn, E.M.; Joshi, L.; Hickey, R.M. Symposium Review: Dairy-Derived Oligosaccharides—Their Influence on Host–Microbe Interactions in the Gastrointestinal Tract of Infants. J. Dairy Sci. 2020, 103, 3816–3827. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; Lebrilla, C.B.; Mills, D.A.; German, J.B.; Freeman, S.L. Breast Milk Oligosaccharides: Structure-Function Relationships in the Neonate. Annu. Rev. Nutr. 2014, 34, 143–169. [Google Scholar] [CrossRef] [Green Version]
- Bienenstock, J.; Buck, R.H.; Linke, H.; Forsythe, P.; Stanisz, A.M.; Kunze, W.A. Fucosylated but Not Sialylated Milk Oligosaccharides Diminish Colon Motor Contractions. PLoS ONE 2013, 8, e76236. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, S.K.; Yatsunenko, T.; Li, D.; Dasgupta, S.; Yu, R.K.; Berg, B.M.; Chichlowski, M.; Odle, J. Dietary Isomers of Sialyllactose Increase Ganglioside Sialic Acid Concentrations in the Corpus Callosum and Cerebellum and Modulate the Colonic Microbiota of Formula-Fed Piglets. J. Nutr. 2016, 146, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeuner, B.; Teze, D.; Muschiol, J.; Meyer, A.S. Synthesis of Human Milk Oligosaccharides: Protein Engineering Strategies for Improved Enzymatic Transglycosylation. Molecules 2019, 24, 2033. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. From Lab Bench to Formulated Ingredient: Characterization, Production, and Commercialization of Human Milk Oligosaccharides. J. Funct. Foods 2020, 72, 104052. [Google Scholar] [CrossRef]
- Ninonuevo, M.R.; Park, Y.; Yin, H.; Zhang, J.; Ward, R.E.; Clowers, B.H.; German, J.B.; Freeman, S.L.; Killeen, K.; Grimm, R.; et al. A Strategy for Annotating the Human Milk Glycome. J. Agric. Food Chem. 2006, 54, 7471–7480. [Google Scholar] [CrossRef]
- Wu, S.; Tao, N.; German, J.B.; Grimm, R.; Lebrilla, C.B. Development of an Annotated Library of Neutral Human Milk Oligosaccharides. J. Proteome Res. 2010, 9, 4138–4151. [Google Scholar] [CrossRef] [Green Version]
- Barile, D.; Tao, N.; Lebrilla, C.B.; Coisson, J.-D.; Arlorio, M.; German, J.B. Permeate from Cheese Whey Ultrafiltration Is a Source of Milk Oligosaccharides. Int. Dairy J. 2009, 19, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Silanikove, N.; Leitner, G.; Merin, U.; Prosser, C.G. Recent Advances in Exploiting Goat’s Milk: Quality, Safety and Production Aspects. Small Rumin. Res. 2010, 89, 110–124. [Google Scholar] [CrossRef]
- Mariño, K.; Lane, J.A.; Abrahams, J.L.; Struwe, W.B.; Harvey, D.J.; Marotta, M.; Hickey, R.M.; Rudd, P.M. Method for Milk Oligosaccharide Profiling by 2-Aminobenzamide Labeling and Hydrophilic Interaction Chromatography. Glycobiology 2011, 21, 1317–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Nakamura, T.; Kawase, H.; Kimura, K.; Watanabe, Y.; Ohtani, M.; Arai, I.; Urashima, T. Concentrations of Sialyloligosaccharides in Bovine Colostrum and Milk during the Prepartum and Early Lactation. J. Dairy Sci. 2003, 86, 1315–1320. [Google Scholar] [CrossRef] [Green Version]
- Fong, B.; Ma, K.; McJarrow, P. Quantification of Bovine Milk Oligosaccharides Using Liquid Chromatography-Selected Reaction Monitoring-Mass Spectrometry. J. Agric. Food Chem. 2011, 59, 9788–9795. [Google Scholar] [CrossRef] [PubMed]
- Özcan, E.; Sela, D.A. Inefficient Metabolism of the Human Milk Oligosaccharides Lacto-N-Tetraose and Lacto-N-Neotetraose Shifts Bifidobacterium longum subsp. infantis Physiology. Front. Nutr. 2018, 5, 46. [Google Scholar] [CrossRef]
- Scammell, A. Production and Uses of Colostrum. Aust. J. Dairy Technol. 2001, 56, 74–82. [Google Scholar]
- McGrath, B.A.; Fox, P.F.; McSweeney, P.L.H.; Kelly, A.L. Composition and Properties of Bovine Colostrum: A Review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef]
- Sischo, W.M.; Short, D.M.; Geissler, M.; Bunyatratchata, A.; Barile, D. Comparative Composition, Diversity, and Abundance of Oligosaccharides in Early Lactation Milk from Commercial Dairy and Beef Cows. J. Dairy Sci. 2017, 100, 3883–3892. [Google Scholar] [CrossRef] [Green Version]
- Sundekilde, U.K.; Barile, D.; Meyrand, M.; Poulsen, N.A.; Larsen, L.B.; Lebrilla, C.B.; German, J.B.; Bertram, H.C. Natural Variability in Bovine Milk Oligosaccharides from Danish Jersey and Holstein-Friesian Breeds. J. Agric. Food Chem. 2012, 60, 6188–6196. [Google Scholar] [CrossRef]
- Levieux, D.; Ollier, A. Bovine Immunoglobulin G, β-Lactoglobulin, α-Lactalbumin and Serum Albumin in Colostrum and Milk during the Early Post Partum Period. J. Dairy Res. 1999, 66, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Godhia, M.L.; Patel, N. Colostrum—Its Composition, Benefits as a Nutraceutical: A Review. Curr. Res. Nutr. 2013, 1, 37–47. [Google Scholar] [CrossRef]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of Milk and Colostrum in Non-Human Mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef]
- Playford, R.J. Peptide Therapy and the Gastroenterologist: Colostrum and Milk-Derived Growth Factors. Clin. Nutr. 2001, 20, 101–106. [Google Scholar] [CrossRef]
- Playford, R.; Macdonald, C.; Johnson, W.S. Colostrum and Milk-Derived Peptide Growth Factors for the Treatment of Gastrointestinal Disorders. Am. J. Clin. Nutr. 2000, 72, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Otterby, D.E. Availability, Storage, Treatment, Composition, and Feeding Value of Surplus Colostrum: A Review. J. Dairy Sci. 1978, 61, 1033–1060. [Google Scholar] [CrossRef]
- Gopal, P.K.; Gill, H.S. Oligosaccharides and Glycoconjugates in Bovine Milk and Colostrum. Br. J. Nutr. 2000, 84, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, P.F.; Greenhill, N.S.; Rowan, A.M.; Schollum, L.M. The Safety of New Zealand Bovine Colostrum: Nutritional and Physiological Evaluation in Rats. Food Chem. Toxicol. 2007, 45, 229–236. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Wang, J.Q.; Yang, Y.X.; Bu, D.P.; Li, S.S.; Zhou, L.Y. Comparative Proteomic Analysis of Changes in the Bovine Whey Proteome during the Transition from Colostrum to Milk. Asian-Australas. J. Anim. Sci. 2011, 24, 272–278. [Google Scholar] [CrossRef]
- Georgiev, I.P. Differences in Chemical Composition Between Cow Colostrum and Milk. Bulg. J. Vet. Med. 2008, 11, 3–12. [Google Scholar]
- Zarcula, S.; Cernescu, H.; Mircu, C.; Tulcan, C.; Morvay, A.; Baul, S.; Popovici, D. Influence of Breed, Parity and Food Intake on Chemical Composition of First Colostrum in Cow. Anim. Sci. Biotechnol. 2010, 43, 43. [Google Scholar]
- Abd El-Fattah, A.M.; Abd Rabo, F.H.R.; EL-Dieb, S.M.; Elkashef, H.A.S. Changes in Composition of Colostrum of Egyptian Buffaloes and Holstein Cows. BMC Vet. Res. 2012, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, T.F.; O’Donovan, M.; Murphy, J.P.; Sugrue, K.; Mannion, D.; McCarthy, W.P.; Timlin, M.; Kilcawley, K.N.; Hickey, R.M.; Tobin, J.T. Evolution of the Bovine Milk Fatty Acid Profile—From Colostrum to Milk Five Days Post Parturition. Int. Dairy J. 2020, 104, 104655. [Google Scholar] [CrossRef]
- Quinn, E.; Slattery, H.; Thompson, A.; Kilcoyne, M.; Joshi, L.; Hickey, R. Mining Milk for Factors Which Increase the Adherence of Bifidobacterium longum subsp. infantis to Intestinal Cells. Foods 2018, 7, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Wishart, D.S. Using Metaboanalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 2016, 14.10.1–14.10.91. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, J.; Hammon, H. Bovine Colostrum: More than Just an Immunoglobulin Supplier. Schweiz. Arch. Tierheilkd. 2000, 142, 221–228. [Google Scholar]
- Uruakpa, F.O.; Ismond, M.A.H.; Akobundu, E.N.T. Colostrum and Its Benefits: A Review. Nutr. Res. 2002, 22, 755–767. [Google Scholar] [CrossRef]
- Newburg, D.S. Glycobiology of Human Milk. Biochemistry 2013, 78, 771–785. [Google Scholar] [CrossRef]
- Martín-Sosa, S.; Martín, M.J.; García-Pardo, L.A.; Hueso, P. Sialyloligosaccharides in Human and Bovine Milk and in Infant Formulas: Variations with the Progression of Lactation. J. Dairy Sci. 2003, 86, 52–59. [Google Scholar] [CrossRef]
- Tao, N.; DePeters, E.J.; German, J.B.; Grimm, R.; Lebrilla, C.B. Variations in Bovine Milk Oligosaccharides during Early and Middle Lactation Stages Analyzed by High-Performance Liquid Chromatography-Chip/Mass Spectrometry. J. Dairy Sci. 2009, 92, 2991–3001. [Google Scholar] [CrossRef] [PubMed]
- Takimori, S.; Shimaoka, H.; Furukawa, J.I.; Yamashita, T.; Amano, M.; Fujitani, N.; Takegawa, Y.; Hammarström, L.; Kacskovics, I.; Shinohara, Y.; et al. Alteration of the N-Glycome of Bovine Milk Glycoproteins during Early Lactation. FEBS J. 2011, 278, 3769–3781. [Google Scholar] [CrossRef] [PubMed]
- Feeney, S.; Gerlach, J.Q.; Slattery, H.; Kilcoyne, M.; Hickey, R.M.; Joshi, L. Lectin Microarray Profiling and Monosaccharide Analysis of Bovine Milk Immunoglobulin G Oligosaccharides during the First 10 Days of Lactation. Food Sci. Nutr. 2019, 7, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ Binding to Bovine Lactoferrin Enhances Protein Stability and Influences the Release of Bacterial Lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48. [Google Scholar] [CrossRef]
- Lis-Kuberka, J.; Orczyk-Pawiłowicz, M. Sialylated Oligosaccharides and Glycoconjugates of Human Milk. The Impact on Infant and Newborn Protection, Development and Well-Being. Nutrients 2019, 11, 306. [Google Scholar] [CrossRef] [Green Version]
- De Leoz, M.L.A.; Wu, S.; Strum, J.S.; Niñonuevo, M.R.; Gaerlan, S.C.; Mirmiran, M.; German, J.B.; Mills, D.A.; Lebrilla, C.B.; Underwood, M.A. A Quantitative and Comprehensive Method to Analyze Human Milk Oligosaccharide Structures in the Urine and Feces of Infants. Anal. Bioanal. Chem. 2013, 405, 4089–4105. [Google Scholar] [CrossRef] [Green Version]
- Padler-Karavani, V.; Varki, A. Potential Impact of the Non-Human Sialic Acid N-Glycolylneuraminic Acid on Transplant Rejection Risk. Xenotransplantation 2011, 18, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Varki, A. Are Humans Prone to Autoimmunity? Implications from Evolutionary Changes in Hominin Sialic Acid Biology. J. Autoimmun. 2017, 83, 134–142. [Google Scholar] [CrossRef]
- Alisson-Silva, F.; Kawanishi, K.; Varki, A. Human Risk of Diseases Associated with Red Meat Intake: Analysis of Current Theories and Proposed Role for Metabolic Incorporation of a Non-Human Sialic Acid. Mol. Asp. Med. 2016, 51, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Tangvoranuntakul, P.; Gagneux, P.; Diaz, S.; Bardor, M.; Varki, N.; Varki, A.; Muchmore, E. Human Uptake and Incorporation of an Immunogenic Nonhuman Dietary Sialic Acid. Proc. Natl. Acad. Sci. USA 2003, 100, 12045–12050. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.H.; Tangvoranuntakul, P.; Varki, A. Effects of Natural Human Antibodies against a Nonhuman Sialic Acid That Metabolically Incorporates into Activated and Malignant Immune Cells. J. Immunol. 2005, 175, 228–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okerblom, J.; Varki, A. Biochemical, Cellular, Physiological, and Pathological Consequences of Human Loss of N-Glycolylneuraminic Acid. ChemBioChem 2017, 18, 1155–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commission Implementing Regulation (EU) 2017/2470. Establishing the Union List of Novel Foods in Accordance with Regulation (EU) 2015/2283 of the European Parliament and of the Council on Novel Foods, Europe. 2017. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32017R2470 (accessed on 30 September 2020).
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of Infant Formula with Human Milk Oligosaccharides on Growth and Morbidity. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idänpään-Heikkilä, I.; Simon, P.M.; Zopf, D.; Vullo, T.; Cahill, P.; Sokol, K.; Tuomanen, E. Oligosaccharides Interfere with the Establishment and Progression of Experimental Pneumococcal Pneumonia. J. Infect. Dis. 1997, 176, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Autran, C.A.; Kellman, B.P.; Kim, J.H.; Asztalos, E.; Blood, A.B.; Spence, E.C.H.; Patel, A.L.; Hou, J.; Lewis, N.E.; Bode, L. Human Milk Oligosaccharide Composition Predicts Risk of Necrotising Enterocolitis in Preterm Infants. Gut 2018, 67, 1064–1070. [Google Scholar] [CrossRef]
- Albrecht, S.; Lane, J.A.; Mariño, K.; Al Busadah, K.A.; Carrington, S.D.; Hickey, R.M.; Rudd, P.M. A Comparative Study of Free Oligosaccharides in the Milk of Domestic Animals. Br. J. Nutr. 2014, 111, 1313–1328. [Google Scholar] [CrossRef]
- Ashida, H.; Miyake, A.; Kiyohara, M.; Wada, J.; Yoshida, E.; Kumagai, H.; Katayama, T.; Yamamoto, K. Two Distinct Alpha-L-Fucosidases from Bifidobacterium bifidum Are Essential for the Utilization of Fucosylated Milk Oligosaccharides and Glycoconjugates. Glycobiology 2009, 19, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.A.; Mariño, K.; Naughton, J.; Kavanaugh, D.; Clyne, M.; Carrington, S.D.; Hickey, R.M. Anti-Infective Bovine Colostrum Oligosaccharides: Campylobacter jejuni as a Case Study. Int. J. Food Microbiol. 2012, 157, 182–188. [Google Scholar] [CrossRef]
- Jung, S.M.; Park, Y.C.; Seo, J.H. Production of 3-Fucosyllactose in Engineered Escherichia coli with α-1,3-Fucosyltransferase from Helicobacter pylori. Biotechnol. J. 2019, 14, e1800498. [Google Scholar] [CrossRef]
- Robinson, R.C.; Poulsen, N.A.; Barile, D. Multiplexed Bovine Milk Oligosaccharide Analysis with Aminoxy Tandem Mass Tags. PLoS ONE 2018, 13, e0196513. [Google Scholar] [CrossRef]
- Asakuma, S.; Urashima, T.; Akahori, M.; Obayashi, H.; Nakamura, T.; Kimura, K.; Watanabe, Y.; Arai, I.; Sanai, Y. Variation of Major Neutral Oligosaccharides Levels in Human Colostrum. Eur. J. Clin. Nutr. 2008, 62, 488–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, K.M.; Townsend, S.D. The Human Milk Glycome as a Defense Against Infectious Diseases: Rationale, Challenges, and Opportunities. ACS Infect. Dis. 2018, 4, 77–83. [Google Scholar] [CrossRef] [PubMed]
- El-Hawiet, A.; Kitova, E.N.; Klassen, J.S. Recognition of Human Milk Oligosaccharides by Bacterial Exotoxins. Glycobiology 2015, 25, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. Human Milk Oligosaccharides: Prebiotics and Beyond. Nutr. Rev. 2009, 67, S183–S191. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, A.; Katoh, T.; Sakanaka, M.; Ling, Y.; Yamada, C.; Asakuma, S.; Urashima, T.; Tomabechi, Y.; Katayama-Ikegami, A.; Kurihara, S.; et al. Sharing of Human Milk Oligosaccharides Degradants within Bifidobacterial Communities in Faecal Cultures Supplemented with Bifidobacterium bifidum. Sci. Rep. 2018, 8, 13958. [Google Scholar] [CrossRef]
- Romero-Velarde, E.; Delgado-Franco, D.; García-Gutiérrez, M.; Gurrola-Díaz, C.; Larrosa-Haro, A.; Montijo-Barrios, E.; Muskiet, F.A.J.; Vargas-Guerrero, B.; Geurts, J. The Importance of Lactose in the Human Diet: Outcomes of a Mexican Consensus Meeting. Nutrients 2019, 11, 2737. [Google Scholar] [CrossRef] [Green Version]
- Antonowicz, I.; Lebenthal, E. Developmental Pattern of Small Intestinal Enterokinase and Disaccharidase Activities in the Human Fetus. Gastroenterology 1977, 72, 1299–1303. [Google Scholar] [CrossRef]
- Raul, F.; Lacroix, B.; Aprahamian, M. Longitudinal Distribution of Brush Border Hydfolases and Morphological Maturation in the Intestine of the Preterm Infant. Early Hum. Dev. 1986, 13, 225–234. [Google Scholar] [CrossRef]
- Weaver, L.T.; Laker, M.F.; Nelson, R. Neonatal Intestinal Lactase Activity. Arch. Dis. Child. 1986, 61, 896–899. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.C. Structures and Metabolic Properties of Bovine Milk Oligosaccharides and Their Potential in the Development of Novel Therapeutics. Front. Nutr. 2019, 6, 50. [Google Scholar] [CrossRef]
- Hassinger, D.; Clausen, D.M.; Nitka, S.; Herdt, A.; Griffin, I. Analysis of Disialyllacto-N-Tetraose (DSLNT) Content in Milk from Mothers of Preterm Infants. J. Hum. Lact. 2020, 36, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Simon, P.M.; Goode, P.L.; Mobasseri, A.; Zopf, D. Inhibition of Helicobacter pylori Binding to Gastrointestinal Epithelial Cells by Sialic Acid-Containing Oligosaccharides. Infect. Immun. 1997, 65, 750–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hester, S.N.; Chen, X.; Li, M.; Monaco, M.H.; Comstock, S.S.; Kuhlenschmidt, T.B.; Kuhlenschmidt, M.S.; Donovan, S.M. Human Milk Oligosaccharides Inhibit Rotavirus Infectivity In Vitro and in Acutely Infected Piglets. Br. J. Nutr. 2013, 110, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavanaugh, D.W.; O’Callaghan, J.; Buttó, L.F.; Slattery, H.; Lane, J.; Clyne, M.; Kane, M.; Joshi, L.; Hickey, R.M. Exposure of Bifidobacterium longum subsp. infantis to Milk Oligosaccharides Increases Adhesion to Epithelial Cells and Induces a Substantial Transcriptional Response. PLoS ONE 2013, 8, e67224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dionex Corporation. Technical Note 42: Glycoprotein Oligosaccharide Analysis Using High-Performance Anion-Exchange Chromatography. Available online: https://assets.thermofisher.com/TFS-Assets/CMD/Application-Notes/5054-TN42_LPN0932.pdf (accessed on 30 September 2020).
- De Moura Bell, J.M.L.N.; Cohen, J.L.; de Aquino, L.F.M.C.; Lee, H.; de Melo Silva, V.L.; Liu, Y.; Domizio, P.; Barile, D. An Integrated Bioprocess to Recover Bovine Milk Oligosaccharides from Colostrum Whey Permeate. J. Food Eng. 2018, 216, 27–35. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinn, E.M.; O’Callaghan, T.F.; Tobin, J.T.; Murphy, J.P.; Sugrue, K.; Slattery, H.; O’Donovan, M.; Hickey, R.M. Changes to the Oligosaccharide Profile of Bovine Milk at the Onset of Lactation. Dairy 2020, 1, 284-296. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy1030019
Quinn EM, O’Callaghan TF, Tobin JT, Murphy JP, Sugrue K, Slattery H, O’Donovan M, Hickey RM. Changes to the Oligosaccharide Profile of Bovine Milk at the Onset of Lactation. Dairy. 2020; 1(3):284-296. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy1030019
Chicago/Turabian StyleQuinn, Erinn M., Tom F. O’Callaghan, John T. Tobin, John Paul Murphy, Katie Sugrue, Helen Slattery, Michael O’Donovan, and Rita M. Hickey. 2020. "Changes to the Oligosaccharide Profile of Bovine Milk at the Onset of Lactation" Dairy 1, no. 3: 284-296. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy1030019