Effects of Energy Supply from Roughage and Concentrates and the Occurrence of Subclinical Ketosis on Blood Chemistry and Liver Health in Lactating Dairy Cows during Early Lactation

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Feeding Experiment
2.2. Sample Collection
2.3. Analyses
2.4. Statistics
3. Results
3.1. Effects of Lactation Stage and Postpartum Energy Supply from Roughage and Concentrates
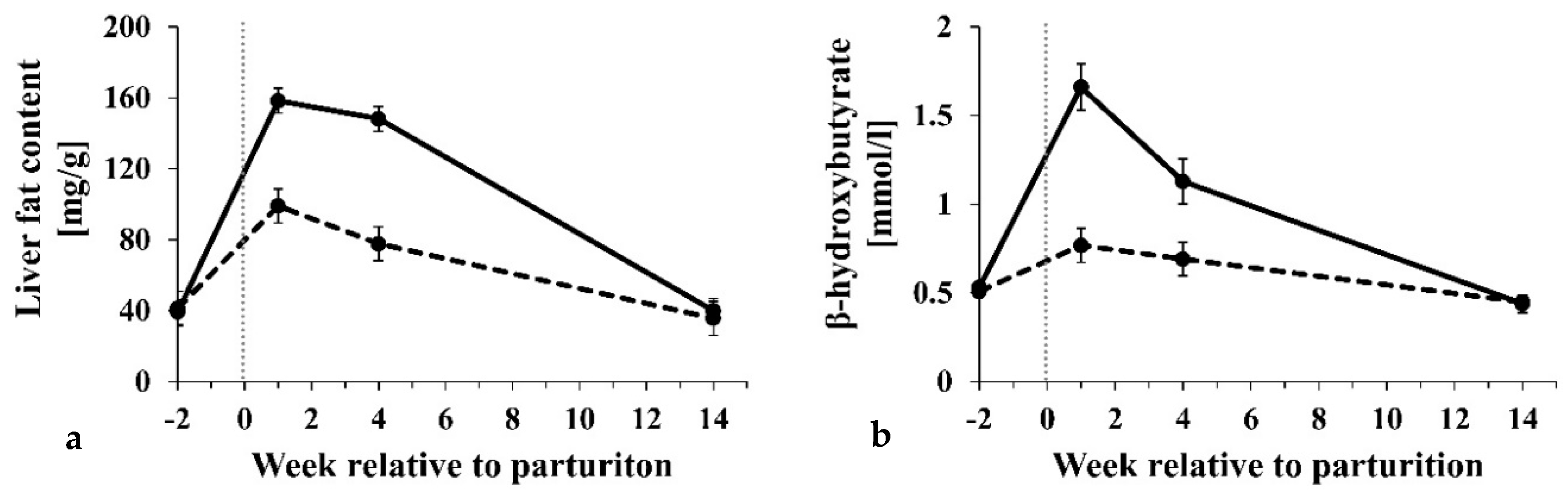
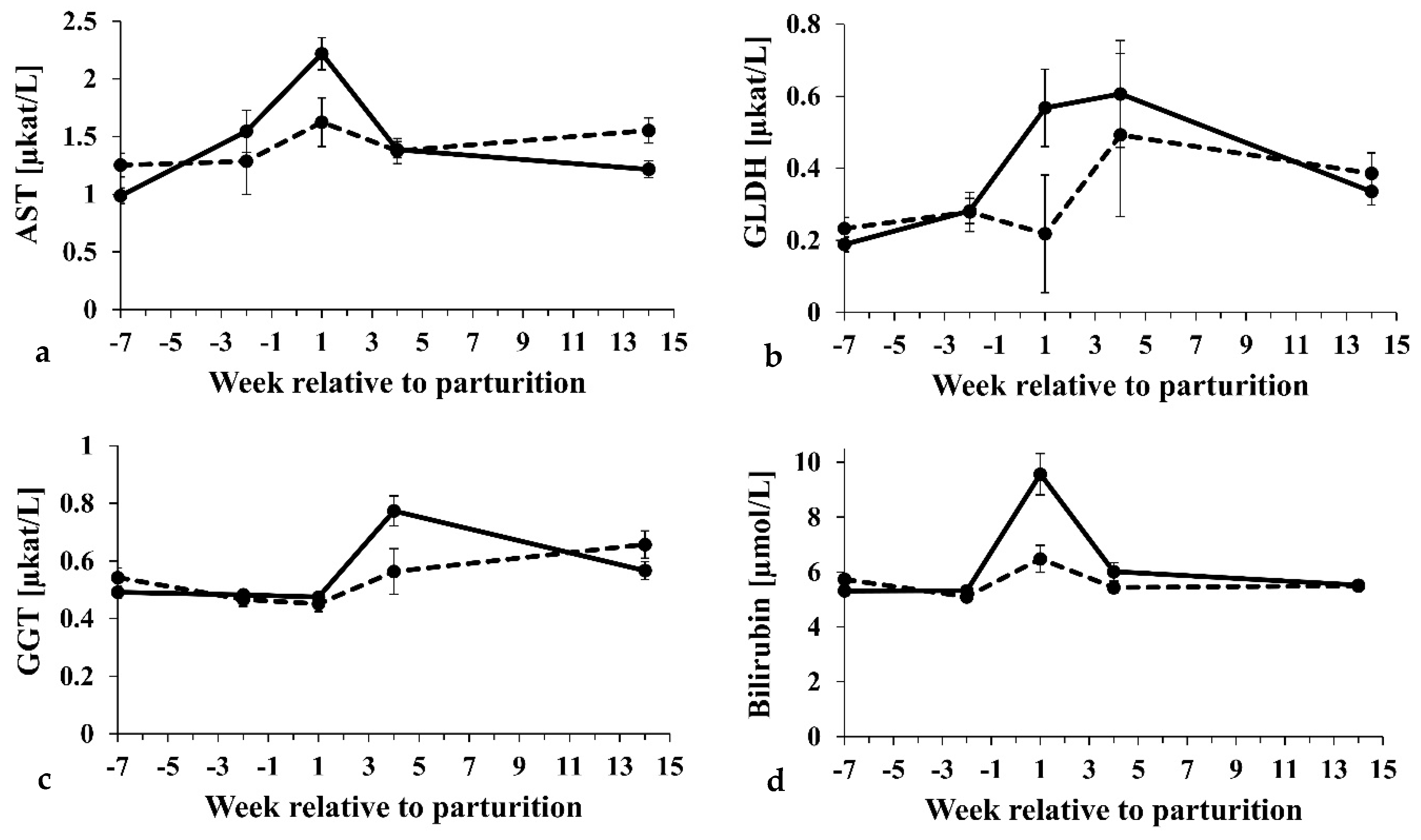
3.2. Influence of Subclinical Ketosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grummer, R.R. Impact of Changes in Organic Nutrient Metabolism on Feeding the Transition Dairy-Cow. J. Anim. Sci. 1995, 73, 2820–2833. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Hametner, C.; Tuchscherer, A.; Losand, B.; Kanitz, E.; Otten, W.; Singh, S.P.; Bruckmaier, R.M.; Becker, F.; Kanitz, W.; et al. Variation in fat mobilization during early lactation differently affects feed intake, body condition, and lipid and glucose metabolism in high-yielding dairy cows. J. Dairy Sci. 2013, 96, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited review: Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [Green Version]
- Zerbe, H.; Schneider, N.; Leibold, W.; Wensing, T.; Kruip, T.; Schuberth, H. Altered functional and immunophenotypical properties of neutrophilic granulocytes in postpartum cows associated with fatty liver. Theriogenology 2000, 54, 771–786. [Google Scholar] [CrossRef]
- Contreras, G.A.; Sordillo, L.M. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp. Immunol. Microbiol. Infect. 2011, 34, 281–289. [Google Scholar] [CrossRef]
- Uchida, E.; Katoh, N.; Takahashi, K. Appearance of haptoglobin in serum from cows at parturition. J. Vet. Med. Sci. 1993, 55, 893–894. [Google Scholar] [CrossRef] [Green Version]
- Schulz, K.; Frahm, J.; Kersten, S.; Meyer, U.; Reiche, D.; Sauerwein, H.; Dänicke, S. Effects of elevated parameters of subclinical ketosis on the immune system of dairy cows: In vivo and in vitro results. Arch. Anim. Nutr. 2015, 69, 113–127. [Google Scholar] [CrossRef]
- Ametaj, B.; Bradford, B.; Bobe, G.; Nafikov, R.; Lu, Y.; Young, J.; Beitz, D. Strong relationships between mediators of the acute phase response and fatty liver in dairy cows. Can. J. Anim. Sci. 2005, 85, 165–175. [Google Scholar] [CrossRef]
- de Jong, W.H.; Smit, R.; Bakker, S.J.; de Vries, E.G.; Kema, I.P. Plasma tryptophan, kynurenine and 3-hydroxykynurenine measurement using automated on-line solid-phase extraction HPLC–tandem mass spectrometry. J. Chromatogr. B 2009, 877, 603–609. [Google Scholar] [CrossRef]
- Drong, C.; Bühler, S.; Frahm, J.; Hüther, L.; Meyer, U.; von Soosten, D.; Gessner, D.; Eder, K.; Sauerwein, H.; Dänicke, S. Effects of body condition, monensin, and essential oils on ruminal lipopolysaccharide concentration, inflammatory markers, and endoplasmatic reticulum stress of transition dairy cows. J. Dairy Sci. 2017, 100, 2751–2764. [Google Scholar] [CrossRef] [Green Version]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.; Schnabel, K.; von Soosten, D.; Meyer, U.; Spiekers, H.; Rehage, J.; Dänicke, S. The effects of energy concentration in roughage and allowance of concentrates on performance, health and energy efficiency of pluriparous dairy cows during early lactation. Arch. Anim Nutr. 2018, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Dirksen, G.; Gründer, H.; Stöber, M. Die Klinische Untersuchung des Rindes [The Clinical Investigation of Bovine Animals], als “Krankheiten des Rindes” Begründet von Gustav Rosenberger; Enke: Stuttgart, Germany, 2012. [Google Scholar]
- Starke, A.; Haudum, A.; Busche, R.; Beyerbach, M.; Dänicke, S.; Rehage, J. Analysis of total lipid and triacylglycerol content in small liver biopsy samples in cattle 1. J. Anim. Sci. 2010, 88, 2741–2750. [Google Scholar] [CrossRef] [PubMed]
- Bühler, S.; Frahm, J.; Tienken, R.; Kersten, S.; Meyer, U.; Huber, K.; Dänicke, S. Effects of energy supply and nicotinic acid supplementation on serum anti-oxidative capacity and on expression of oxidative stress-related genes in blood leucocytes of periparturient primi-and pluriparous dairy cows. J. Anim. Physiol. Anim. Nutr. 2017, 102, e87–e98. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Hiss, S.; Mielenz, M.; Bruckmaier, R.; Sauerwein, H. Haptoglobin concentrations in blood and milk after endotoxin challenge and quantification of mammary Hp mRNA expression. J. Dairy Sci. 2004, 87, 3778–3784. [Google Scholar] [CrossRef]
- Hüther, L.; Hartwiger, J.; Drong, C.; Meyer, U.; Dänicke, S.J.J.V.S.M.D. Simultaneous determination of tryptophan, kynurenine and niacin in serum of periparturient dairy cows by high-performance liquid chromatography with diode array detection. J. Vet. Sci. Med. Diagn 2016, 5. [Google Scholar] [CrossRef]
- Littell, R.C.; Stroup, W.W.; Milliken, G.A.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models; SAS Institute: Cary, NC, USA, 2006. [Google Scholar]
- Kraft, W. Klinische Labordiagnostik in der Tiermedizin; Schattauer Verlag: Stuttgart, Germany, 2005. [Google Scholar]
- Drong, C.; Meyer, U.; Soosten, D.; Frahm, J.; Rehage, J.; Schirrmeier, H.; Beer, M.; Dänicke, S. Effects of monensin and essential oils on immunological, haematological and biochemical parameters of cows during the transition period. J. Anim. Physiol. Anim. Nutr. 2017, 101, 791–806. [Google Scholar] [CrossRef]
- Meglia, G.; Johannisson, A.; Petersson, L.; Waller, K.P. Changes in some blood micronutrients, leukocytes and neutrophil expression of adhesion molecules in periparturient dairy cows. Acta Vet. Scand. 2001, 42, 139. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Gohar, A.; Ahmad, N.; Ahmad, M. Haematological profile in cyclic, non-cyclic and endometritic cross-bred cattle. Int. J. Agric. Biol. 2003, 5, 332–334. [Google Scholar]
- Olmos, G.; Boyle, L.; Horan, B.; Berry, D.P.; Sayers, R.; Hanlon, A.; Mee, J.F. Effect of Holstein–Friesian genetic group on peripartum and early lactation haematological and acute phase proteins profiles, health and fertility. Animal 2009, 3, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, E.; Gross, J.J.; Bruckmaier, R.; Albrecht, C. Cholesterol metabolism, transport, and hepatic regulation in dairy cows during transition and early lactation. J. Dairy Sci. 2014, 97, 5481–5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroz-Rocha, G.F.; LeBlanc, S.J.; Duffield, T.F.; Wood, D.; Leslie, K.E.; Jacobs, R.M. Reference limits for biochemical and hematological analytes of dairy cows one week before and one week after parturition. Can. Vet. J. 2009, 50, 383. [Google Scholar] [PubMed]
- Glatz-Hoppe, J.; Boldt, A.; Spiekers, H.; Mohr, E.; Losand, B. Relationship between milk constituents from milk testing and health, feeding, and metabolic data of dairy cows. J. Dairy Sci. 2020, 11, 10175–10194. [Google Scholar] [CrossRef] [PubMed]
- Krogh, M.A.; Toft, N.; Enevoldsen, C. Latent class evaluation of a milk test, a urine test, and the fat-to-protein percentage ratio in milk to diagnose ketosis in dairy cows. J. Dairy Sci. 2011, 94, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, L.R.; de Oliveira Neto, C.B.; de Araújo, M.J.; Edvan, R.L.; de Oliveira, W.D.; Pereira, F.B. Major metabolic diseases affecting cows in transition period. Int. J. Biol. 2014, 6, 85. [Google Scholar] [CrossRef]
- Goff, J.P.; Horst, R.L. Physiological changes at parturition and their relationship to metabolic disorders. J. Dairy Sci. 1997, 80, 1260–1268. [Google Scholar] [CrossRef]
- Kessel, S.; Stroehl, M.; Meyer, H.H.D.; Hiss, S.; Sauerwein, H.; Schwarz, F.J.; Bruckmaier, R.M. Individual variability in physiological adaptation to metabolic stress during early lactation in dairy cows kept under equal conditions. J. Anim. Sci. 2008, 86, 2903–2912. [Google Scholar] [CrossRef]
- Bertoni, G.; Trevisi, E.; Han, X.; Bionaz, M. Effects of inflammatory conditions on liver activity in puerperium period and consequences for performance in dairy cows. J. Dairy Sci. 2008, 91, 3300–3310. [Google Scholar] [CrossRef] [Green Version]
- Hammon, D.; Evjen, I.; Dhiman, T.; Goff, J.; Walters, J. Neutrophil function and energy status in Holstein cows with uterine health disorders. Vet. Immunol. Immunopathol. 2006, 113, 21–29. [Google Scholar] [CrossRef]
- Trevisi, E.; Amadori, M.; Archetti, I.; Lacetera, N.; Bertoni, G. Inflammatory response and acute phase proteins in the transition period of high-yielding dairy cows. In Acute Phase Proteins as Early Non-Specific Biomarkers of Human and Veterinary Diseases; InTech: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy cows. Physiol. Genom. 2007, 32, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Shu, S.; Xia, C.; Wang, B.; Zhang, H.; Jun, B. Investigation on the relationship of insulin resistance and ketosis in dairy cows. Vet. Sci. Technol. 2014, 5. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Fatty acids as modulators of the cellular production of reactive oxygen species. Free Radic. Biol. Med. 2008, 45, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Urh, C.; Denißen, J.; Gerster, E.; Kraus, N.; Stamer, E.; Heitkönig, B.; Spiekers, H.; Sauerwein, H. Pro-and antioxidative indicators in serum of dairy cows during late pregnancy and early lactation: Testing the effects of parity, different dietary energy levels, and farm. J. Dairy Sci. 2019, 102, 6672–6678. [Google Scholar] [CrossRef] [PubMed]
- Credille, B.C.; Woolums, A.R.; Overton, M.W.; Hurley, D.J.; Giguère, S. Expression of inflammation-associated genes in circulating leukocytes and activity of indoleamine-2, 3-dioxygenase in dairy cattle with acute puerperal metritis and bacteremia. Res. Vet. Sci. 2015, 101, 6–10. [Google Scholar] [CrossRef]
- Bochniarz, M.; Kocki, T.; Dąbrowski, R.; Szczubiał, M.; Wawron, W.; Turski, W.A. Tryptophan, kynurenine, kynurenic acid concentrations and indoleamine 2, 3-dioxygenase activity in serum and milk of dairy cows with subclinical mastitis caused by coagulase-negative staphylococci. Reprod. Domest. Anim. 2018, 53, 1491–1497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Dry Period 1 | Postpartum Period 2 | ||||
|---|---|---|---|---|---|
| MRMC | MRHC | HRMC | HRHC | ||
| Dry matter [g/kg] | 347 | 547 | 601 | 523 | 580 |
| Nutrient [g/kg DM] | |||||
| Crude ash | 65 | 66 | 65 | 67 | 65 |
| Crude protein | 118 | 129 | 138 | 135 | 142 |
| Ether extract | 38 | 37 | 39 | 38 | 40 |
| Crude fiber | 189 | 205 | 184 | 188 | 173 |
| aNeutral detergent fiber om | 375 | 404 | 366 | 372 | 345 |
| Acid detergent fiber om | 215 | 241 | 221 | 221 | 207 |
| Energy [MJ/kg DM] | |||||
| Metabolisable energy | 11.2 | 11 | 11.5 | 11.3 | 11.7 |
| Net energy lactation | 6.9 | 6.7 | 7.1 | 6.9 | 7.2 |
| MR | HR | p-Value 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MC (n = 16) | HC (n = 16) | MC (n = 16) | HC (n = 15) | SEM 2 | R | C | W 3 | R × C | C × W | |
| White blood cell count | ||||||||||
| White blood cells [103/µL] | 7.89 | 7.54 | 8.37 | 7.80 | 0.34 | 0.277 | 0.174 | 0.069 | 0.751 | 0.995 |
| Granulocytes 4 [%] | 48.6 | 56.0 | 54.4 | 53.0 | 2.0 | 0.491 | 0.135 | 0.862 | 0.034 | 0.237 |
| Granulocytes 4 [103/µL] | 3.87 | 4.30 | 4.78 | 4.16 | 0.31 | 0.204 | 0.759 | 0.213 | 0.093 | 0.718 |
| Lymphocytes [%] | 41.9 | 37.7 | 38.4 | 40.6 | 1.7 | 0.859 | 0.554 | 0.448 | 0.070 | 0.327 |
| Lymphocytes [103/µL] | 3.28 | 2.78 | 3.05 | 3.16 | 0.15 | 0.614 | 0.196 | 0.837 | 0.046 | 0.261 |
| Eosinophils [%] | 8.49 | 5.94 | 6.23 | 6.24 | 0.90 | 0.268 | 0.156 | 0.064 | 0.153 | 0.099 |
| Eosinophils [103/µL] | 0.61 | 0.47 | 0.50 | 0.47 | 0.06 | 0.379 | 0.140 | 0.011 | 0.371 | 0.099 |
| Monocytes [%] | 0.86 | 0.64 | 0.59 | 0.53 | 0.10 | 0.169 | 0.311 | 0.849 | 0.560 | 0.732 |
| Monocytes [103/µL] | 0.07 | 0.04 | 0.04 | 0.03 | 0.01 | 0.124 | 0.147 | 0.830 | 0.384 | 0.313 |
| Red blood cell count | ||||||||||
| Haematocrit [%] | 28.3 | 28.3 | 28.1 | 28.5 | 0.5 | 0.936 | 0.664 | <0.001 | 0.702 | 0.087 |
| Haemoglobin [mmol/L] | 5.54 | 5.63 | 5.51 | 5.63 | 0.09 | 0.908 | 0.282 | <0.001 | 0.875 | 0.079 |
| Red blood cells [106/µL] | 6.15 | 5.72 | 5.85 | 5.93 | 0.12 | 0.714 | 0.163 | <0.001 | 0.042 | 0.027 |
| Mean corpuscular volume [fL] | 46.2 | 49.7 | 48.0 | 48.1 | 0.7 | 0.845 | 0.015 | <0.001 | 0.026 | 0.697 |
| Mean corpuscular haemoglobin, [pg] | 14.6 | 16.1 | 15.3 | 15.4 | 0.3 | 0.973 | 0.006 | <0.001 | 0.019 | 0.915 |
| MR | HR | p-Value 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| MC (n = 16) | HC (n = 16) | MC (n = 16) | HC (n = 15) | SEM 2 | R | C | W 3 | |
| Albumin [g/L] | 36.0 | 35.9 | 36.5 | 36.0 | 0.5 | 0.545 | 0.614 | <0.001 |
| Cholesterol [mmol/L] | 3.72 | 3.79 | 4.06 | 4.10 | 0.16 | 0.051 | 0.738 | <0.001 |
| Total protein [g/L] | 68.5 | 71.5 | 70.8 | 71.4 | 0.7 | 0.143 | 0.018 | <0.001 |
| Total bilirubin [μmol/L] | 5.88 | 6.03 | 6.49 | 6.30 | 0.34 | 0.186 | 0.947 | <0.001 |
| AST 4 [µkat/L] | 1.57 | 1.52 | 1.57 | 1.52 | 0.10 | 0.973 | 0.622 | 0.002 |
| GLDH 5 [µkat/L] | 0.45 | 0.43 | 0.32 | 0.43 | 0.12 | 0.578 | 0.701 | 0.373 |
| AP 6 [µkat/L] | 0.76 | 0.72 | 0.70 | 0.74 | 0.05 | 0.641 | 0.960 | <0.001 |
| GGT 7 [µkat/L] | 0.54 | 0.58 | 0.61 | 0.56 | 0.04 | 0.516 | 0.830 | <0.001 |
| Triglycerides [mmol/L] | 0.139 | 0.139 | 0.142 | 0.139 | 0.008 | 0.837 | 0.849 | 0.001 |
| Urea [mmol/L] | 3.50 | 3.29 | 3.15 | 3.16 | 0.15 | 0.128 | 0.543 | <0.001 |
| Uric acid [μmol/L] | 71.92 | 70.30 | 72.44 | 71.69 | 2.98 | 0.755 | 0.701 | <0.001 |
| MR | HR | p-Value 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| MC | HC | MC | HC | SEM 2 | R | C | W 3 | |
| Number of cows | 10 | 10 | 10 | 10 | ||||
| Liver fat content [mg/g] | 92.9 | 85.6 | 72.4 | 94.6 | 11.5 | 0.616 | 0.522 | <0.001 |
| GPx 4 [mU/mL haemoglobin] | 2349 | 2264 | 2257 | 2122 | 129 | 0.367 | 0.406 | <0.001 |
| SOD 5 [mU/mL haemoglobin] | 64.6 | 59.0 | 55.9 | 59.3 | 4.1 | 0.314 | 0.792 | <0.001 |
| FRAP 6 [µmol/L] | 297 | 303 | 320 | 310 | 11 | 0.157 | 0.856 | <0.001 |
| Haptoglobin [µg/mL] | 705 | 1044 | 536 | 450 | 326 | 0.251 | 0.701 | 0.009 |
| Number of cows | 5 | 5 | 5 | 5 | ||||
| Kynurenine (K) [mg/L] | 1.02 | 0.82 | 1.06 | 1.05 | 0.12 | 0.242 | 0.384 | <0.001 |
| Tryptophan (T) [mg/L] | 5.16 | 5.23 | 5.75 | 5.46 | 0.37 | 0.280 | 0.772 | <0.001 |
| K:T ratio 7 | 0.17 | 0.18 | 0.18 | 0.18 | 0.02 | 0.685 | 0.964 | 0.063 |
| Incidence of SCK 1 | p-Value | ||||
|---|---|---|---|---|---|
| Control Group | SK Group | SCK | Week | SCK × Week | |
| Number of cows | 44 | 19 | |||
| β-hydroxybutyrate (mmol/L) | 0.59 ± 0.2 | 0.87 ± 0.3 | <0.001 | <0.001 | <0.001 |
| White blood cells [103/µL] | 7.79 ± 0.2 | 7.68 ± 0.3 | 0.780 | <0.001 | 0.853 |
| Lymphocytes [%] | 40.46 ± 0.9 | 37.81 ± 1.4 | 0.119 | 0.107 | 0.568 |
| Eosinophiles [%] | 7.82 ± 0.43 | 7.16 ± 0.64 | 0.396 | 0.003 | 0.596 |
| Monocytes [%] | 0.97 ± 0.08 | 0.80 ± 0.11 | 0.236 | 0.004 | 0.513 |
| Granulocytes 2 [%] | 50.7 ± 0.9 | 54.2 ± 1.3 | 0.026 | <0.001 | 0.978 |
| Total bilirubin [µmol/L] | 5.64 ± 0.17 | 6.33 ± 0.17 | 0.005 | <0.001 | <0.001 |
| AST 3 [µkat/L] | 1.42 ± 0.07 | 1.47 ± 0.1 | 0.662 | <0.001 | <0.001 |
| GLDH 4 [µkat/L] | 0.32 ± 0.04 | 0.40 ± 0.06 | 0.315 | 0.001 | 0.330 |
| AP 5 [µkat/L] | 0.82 ± 0.03 | 0.76 ± 0.04 | 0.219 | <0.001 | 0.226 |
| GGT 6 [µkat/L] | 0.54 ± 0.02 | 0.56 ± 0.03 | 0.500 | <0.001 | 0.024 |
| Triglycerides [mmol/L] | 0.203 ± 0.001 | 0.199 ± 0.008 | 0.726 | <0.001 | 0.047 |
| Number of cows | 26 | 14 | |||
| Liver fat content [mg/g] | 63.4 ± 4.2 | 96.3 ± 5.7 | 0.616 | 0.522 | <0.001 |
| GPx 7 [mU/mL HG] | 2039 ± 77 | 2269 ± 106 | 0.088 | <0.001 | 0.961 |
| SOD 8 [mU/mL HG] | 54.8 ± 2.3 | 61.3 ± 3.2 | 0.105 | <0.001 | 0.201 |
| FRAP 9 [µmol/L] | 287 ± 6 | 285 ± 8 | 0.878 | <0.001 | 0.702 |
| Haptoglobin [µg/mL] | 420 ± 128 | 728 ± 174 | 0.155 | 0.002 | 0.265 |
| Number of cows | 12 | 8 | |||
| Kynurenine (K) [mg/L] | 0.96 ± 0.07 | 1.02 ± 0.08 | 0.599 | <0.001 | 0.133 |
| Tryptophan (T) [mg/L] | 5.44 ± 0.18 | 5.31 ± 0.20 | 0.623 | <0.001 | 0.277 |
| K:T ratio 10 | 0.17 ± 0.01 | 0.19 ± 0.01 | 0.296 | 0.157 | 0.798 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmitz, R.; Schnabel, K.; Frahm, J.; von Soosten, D.; Meyer, U.; Hüther, L.; Spiekers, H.; Rehage, J.; Sauerwein, H.; Dänicke, S. Effects of Energy Supply from Roughage and Concentrates and the Occurrence of Subclinical Ketosis on Blood Chemistry and Liver Health in Lactating Dairy Cows during Early Lactation. Dairy 2021, 2, 25-39. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2010003
Schmitz R, Schnabel K, Frahm J, von Soosten D, Meyer U, Hüther L, Spiekers H, Rehage J, Sauerwein H, Dänicke S. Effects of Energy Supply from Roughage and Concentrates and the Occurrence of Subclinical Ketosis on Blood Chemistry and Liver Health in Lactating Dairy Cows during Early Lactation. Dairy. 2021; 2(1):25-39. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2010003
Chicago/Turabian StyleSchmitz, Rolf, Karina Schnabel, Jana Frahm, Dirk von Soosten, Ulrich Meyer, Liane Hüther, Hubert Spiekers, Jürgen Rehage, Helga Sauerwein, and Sven Dänicke. 2021. "Effects of Energy Supply from Roughage and Concentrates and the Occurrence of Subclinical Ketosis on Blood Chemistry and Liver Health in Lactating Dairy Cows during Early Lactation" Dairy 2, no. 1: 25-39. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2010003