
Characterization of Chemical and Bacterial Compositions of Dairy Wastewaters

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Chemical Analysis of Dairy Wastewater
2.3. Metabarcoding Analysis of Dairy Wastewater
2.3.1. Preparation of Samples and Genomic DNA Extraction
2.3.2. Quantification of 16S Gene Copies, High-Throughput Amplicon Sequencing and Bioinformatics
2.4. Statistical Analysis
3. Results
3.1. Chemical Characteristics and Composition of Dairy Wastewaters
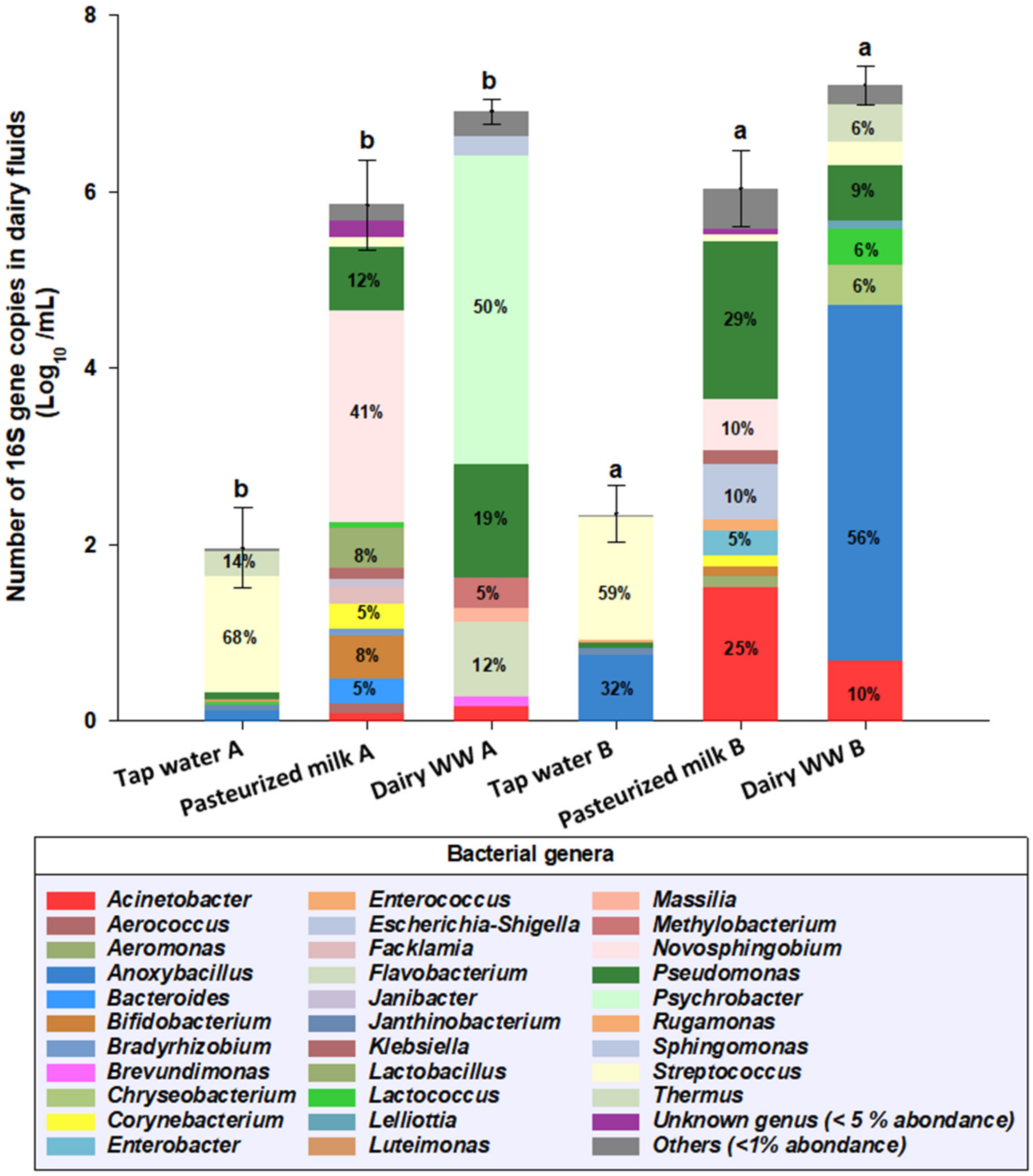
3.2. Bacterial Communities Founds in Dairy Fluids
4. Discussion
4.1. Differences in the Chemical Characteristics of White and Cleaning Wastewater
4.2. Bacterial Genera Found in Dairy White Wastewaters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vourch, M.; Balannec, B.; Chaufer, B.; Dorange, G. Treatment of Dairy Industry Wastewater by Reverse Osmosis for Water Reuse. Desalination 2008, 219, 190–202. [Google Scholar] [CrossRef]
- Balannec, B.; Vourch, M.; Rabiller-Baudry, M.; Chaufer, B. Comparative Study of Different Nanofiltration and Reverse Osmosis Membranes for Dairy Effluent Treatment by Dead-End Filtration. Sep. Purif. Technol. 2005, 42, 195–200. [Google Scholar] [CrossRef]
- Fernández, P.; Riera, F.A.; Álvarez, R.; Álvarez, S. Nanofiltration Regeneration of Contaminated Single-Phase Detergents Used in the Dairy Industry. J. Food Eng. 2010, 97, 319–328. [Google Scholar] [CrossRef]
- Seale, B.; Bremer, P.; Flint, S.; Brooks, J.; Palmer, J. Overview of the Problems Resulting from Biofilm Contamination in the Dairy Industry. In Biofilms in the Dairy Industry; Teh, K.H., Flint, S., Brooks, J., Knight, G., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 49–64. ISBN 978-1-118-87628-2. [Google Scholar]
- Daufin, G.; Escudier, J.-P.; Carrère, H.; Bérot, S.; Fillaudeau, L.; Decloux, M. Recent and Emerging Applications of Membrane Processes in the Food and Dairy Industry. Food Bioprod. Process. 2001, 79, 89–102. [Google Scholar] [CrossRef]
- Balannec, B.; Gésan-Guiziou, G.; Chaufer, B.; Rabiller-Baudry, M.; Daufin, G. Treatment of Dairy Process Waters by Membrane Operations for Water Reuse and Milk Constituents Concentration. Desalination 2002, 147, 89–94. [Google Scholar] [CrossRef]
- Slavov, A.K. Dairy Wastewaters—General Characteristics and Treatment Possibilities—A Review. Food Technol. Biotechnol. 2017, 55. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, J.P.; Srivastava, V.C.; Mall, I.D. An Overview of Various Technologies for the Treatment of Dairy Wastewaters. Crit. Rev. Food Sci. Nutr. 2011, 51, 442–452. [Google Scholar] [CrossRef]
- Chi, F.H.; Cheng, W.P. Use of Chitosan as Coagulant to Treat Wastewater from Milk Processing Plant. J. Polym. Environ. 2006, 14, 411–417. [Google Scholar] [CrossRef]
- Ghernaout, D. Water Reuse: Extenuating Membrane Fouling in Membrane Processes. Am. J. Chem. Eng. 2018, 6, 25. [Google Scholar] [CrossRef]
- Suárez, A.; Riera, F.A. Production of High-Quality Water by Reverse Osmosis of Milk Dairy Condensates. J. Ind. Eng. Chem. 2015, 21, 1340–1349. [Google Scholar] [CrossRef]
- Brião, V.B.; Vieira Salla, A.C.; Miorando, T.; Hemkemeier, M.; Cadore Favaretto, D.P. Water Recovery from Dairy Rinse Water by Reverse Osmosis: Giving Value to Water and Milk Solids. Resour. Conserv. Recycl. 2019, 140, 313–323. [Google Scholar] [CrossRef]
- Gésan-Guiziou, G.; Alvarez, N.; Jacob, D.; Daufin, G. Cleaning-in-Place Coupled with Membrane Regeneration for Re-Using Caustic Soda Solutions. Sep. Purif. Technol. 2007, 54, 329–339. [Google Scholar] [CrossRef]
- Vourch, M.; Balannec, B.; Chaufer, B.; Dorange, G. Nanofiltration and Reverse Osmosis of Model Process Waters Fromthe Dairy Industry to Produce Water for Reuse. Desalination 2005, 172, 245–256. [Google Scholar] [CrossRef]
- Bennani, C.F.; Ousji, B.; Ennigrou, D.J. Reclamation of Dairy Wastewater Using Ultrafiltration Process. Desalination Water Treat. 2015, 55, 297–303. [Google Scholar] [CrossRef]
- Luo, J.; Ding, L. Influence of PH on Treatment of Dairy Wastewater by Nanofiltration Using Shear-Enhanced Filtration System. Desalination 2011, 278, 150–156. [Google Scholar] [CrossRef]
- Chamberland, J.; Lessard, M.-H.; Doyen, A.; Labrie, S.; Pouliot, Y. A Sequencing Approach Targeting the 16S RRNA Gene Unravels the Biofilm Composition of Spiral-Wound Membranes Used in the Dairy Industry. Dairy Sci. Technol. 2017, 96, 827–843. [Google Scholar] [CrossRef] [Green Version]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic Identification and in Situ Detection of Individual Microbial Cells without Cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Pace, N.R. A Molecular View of Microbial Diversity and the Biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef]
- Delbès, C.; Ali-Mandjee, L.; Montel, M.-C. Monitoring Bacterial Communities in Raw Milk and Cheese by Culture-Dependent and -Independent 16S rRNA Gene-Based Analyses. Appl. Environ. Microbiol. 2007, 73, 1882–1891. [Google Scholar] [CrossRef] [Green Version]
- McGarvey, J.A.; Miller, W.G.; Sanchez, S.; Stanker, L. Identification of Bacterial Populations in Dairy Wastewaters by Use of 16S rRNA Gene Sequences and Other Genetic Markers. Appl. Environ. Microbiol. 2004, 70, 4267–4275. [Google Scholar] [CrossRef] [Green Version]
- Ni’am, M.F.; Othman, F. Experimental Design of Electrocoagulation and Magnetic Technology for Enhancing Suspended Solids Removal from Synthetic Wastewater. Int. J. Sci. Eng. 2014, 7, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Chamberland, J.; Beaulieu-Carbonneau, G.; Lessard, M.-H.; Labrie, S.; Bazinet, L.; Doyen, A.; Pouliot, Y. Effect of Membrane Material Chemistry and Properties on Biofouling Susceptibility during Milk and Cheese Whey Ultrafiltration. J. Membr. Sci. 2017, 542, 208–216. [Google Scholar] [CrossRef]
- Lemarchand, K.; Berthiaume, F.; Maynard, C.; Harel, J.; Payment, P.; Bayardelle, P.; Masson, L.; Brousseau, R. Optimization of Microbial DNA Extraction and Purification from Raw Wastewater Samples for Downstream Pathogen Detection by Microarrays. J. Microbiol. Methods 2005, 63, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Li, W.K.W.; Tremblay, J.-É.; Carmack, E.C.; Lovejoy, C. Arctic Ocean Microbial Community Structure before and after the 2007 Record Sea Ice Minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.-I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The Microbiome Extends to Subepidermal Compartments of Normal Skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Doebeli, M.; Parfrey, L.W. Correcting for 16S rRNA Gene Copy Numbers in Microbiome Surveys Remains an Unsolved Problem. Microbiome 2018, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, A.; Sobhey, M.; Badawy, M. Treatment of a Combined Dairy and Domestic Wastewater in an Up-Flow Anaerobic Sludge Blanket (UASB) Reactor Followed by Activated Sludge (AS System). Desalination 2008, 227, 167–177. [Google Scholar] [CrossRef]
- Janczukowicz, W.; Zieliński, M.; Dębowski, M. Biodegradability Evaluation of Dairy Effluents Originated in Selected Sections of Dairy Production. Bioresour. Technol. 2008, 99, 4199–4205. [Google Scholar] [CrossRef] [PubMed]
- Preininger, C.; Klimant, I.; Wolfbeis, O.S. Optical Fiber Sensor for Biological Oxygen Demand. Anal. Chem. 1994, 66, 1841–1846. Available online: https://0-pubs-acs-org.brum.beds.ac.uk/doi/pdf/10.1021/ac00083a011 (accessed on 13 March 2020). [CrossRef]
- Sarkar, B.; Chakrabarti, P.P.; Vijaykumar, A.; Kale, V. Wastewater Treatment in Dairy Industries—Possibility of Reuse. Desalination 2006, 195, 141–152. [Google Scholar] [CrossRef]
- Yapıcıoğlu, P.; Yeşilnacar, M.I. Energy Cost Assessment of a Dairy Industry Wastewater Treatment Plant. Environ. Monit. Assess. 2020, 192, 536. [Google Scholar] [CrossRef]
- Chamberland, J.; Benoit, S.; Doyen, A.; Pouliot, Y. Integrating Reverse Osmosis to Reduce Water and Energy Consumption in Dairy Processing: A Predictive Analysis for Cheddar Cheese Manufacturing Plants. J. Water Process Eng. 2020, 38, 101606. [Google Scholar] [CrossRef]
- Parente, E.; Ricciardi, A.; Zotta, T. The Microbiota of Dairy Milk: A Review. Int. Dairy J. 2020, 107, 104714. [Google Scholar] [CrossRef]
- De Oliveira, G.B.; Favarin, L.; Luchese, R.H.; McIntosh, D. Psychrotrophic Bacteria in Milk: How Much Do We Really Know? Braz. J. Microbiol. 2015, 46, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Nam, H.M.; Tamang, M.D.; Chae, M.H.; Jang, G.C.; Jung, S.C.; Lim, S.K. Prevalence and Antimicrobial Susceptibility of Acinetobacter from Raw Bulk Tank Milk in Korea. J. Dairy Sci. 2013, 96, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, M.L.; Boor, K.J. Short Communication: Bacterial Ecology of High-Temperature, Short-Time Pasteurized Milk Processed in the United States. J. Dairy Sci. 2009, 92, 4833–4840. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The Complex Microbiota of Raw Milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, R.; Liu, Y.; Yang, S.; Liu, Y.; Shi, H.; Yue, X.; Wu, R.; Wu, J. High-Throughput Sequencing Provides New Insights into the Roles and Implications of Core Microbiota Present in Pasteurized Milk. Food Res. Int. 2020, 137, 109586. [Google Scholar] [CrossRef] [PubMed]
- Dumalisile, P.; Witthuhn, R.C.; Britz, T.J. Impact of Different Pasteurization Temperatures on the Survival of Microbial Contaminants Isolated from Pasteurized Milk. Int. J. Dairy Technol. 2005, 58, 74–82. [Google Scholar] [CrossRef]
- Langsrud, S.; Seifert, L.; Møretrø, T. Characterization of the Microbial Flora in Disinfecting Footbaths with Hypochlorite. J. Food Prot. 2006, 69, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Cherif-Antar, A.; Moussa–Boudjemâa, B.; Didouh, N.; Medjahdi, K.; Mayo, B.; Flórez, A.B. Diversity and Biofilm-Forming Capability of Bacteria Recovered from Stainless Steel Pipes of a Milk-Processing Dairy Plant. Dairy Sci. Technol. 2016, 96, 27–38. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and Antibiotic Resistance in Pseudomonas Spp. from Drinking Water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.; Flint, S.; Palmer, J.; Love, R.; Biggs, P.; Beuger, A. Analysis of Culturable and Non-Culturable Bacteria and Their Potential to Form Biofilms in a Primary Treated Dairy Wastewater System. Environ. Technol. 2018, 39, 2185–2192. [Google Scholar] [CrossRef]
- Chiesa, F.; Lomonaco, S.; Nucera, D.; Garoglio, D.; Dalmasso, A.; Civera, T. Distribution of Pseudomonas Species in a Dairy Plant Affected by Occasional Blue Discoloration. Ital. J. Food Saf. 2014, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Sadiq, F.A.; Burmølle, M.; Wang, N.; He, G. Insights into Psychrotrophic Bacteria in Raw Milk: A Review. J. Food Prot. 2019, 82, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Denner, E.B.M.; Mark, B.; Mark, B.; Busse, H.-J.; Busse, H.-J.; Turkiewicz, M.; Lubitz, W. Psychrobacter Proteolyticus Sp. Nov., a Psychrotrophic, Halotolerant Bacterium Isolated from the Antarctic Krill Euphausia Superba Dana, Excreting a Cold-Adapted Metalloprotease. Syst. Appl. Microbiol. 2001, 24, 44–53. [Google Scholar] [CrossRef]
- Hantsis-Zacharov, E.; Halpern, M. Culturable Psychrotrophic Bacterial Communities in Raw Milk and Their Proteolytic and Lipolytic Traits. Appl. Environ. Microbiol. 2007, 73, 7162–7168. [Google Scholar] [CrossRef] [Green Version]
- Quigley, L.; McCarthy, R.; O’Sullivan, O.; Beresford, T.P.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C.; Cotter, P.D. The Microbial Content of Raw and Pasteurized Cow Milk as Determined by Molecular Approaches. J. Dairy Sci. 2013, 96, 4928–4937. [Google Scholar] [CrossRef]
- Techer, C. MC SPOILAGE OF ANIMAL PRODUCTS | Microbial Milk Spoilage. Encycl. Food Microbiol. 2014, 3, 446–452. [Google Scholar]
- Goh, K.M.; Kahar, U.M.; Chai, Y.Y.; Chong, C.S.; Chai, K.P.; Ranjani, V.; Illias, R.M.; Chan, K.-G. Recent Discoveries and Applications of Anoxybacillus. Appl. Microbiol. Biotechnol. 2013, 97, 1475–1488. [Google Scholar] [CrossRef]
- Rusiñol, M.; Martínez-Puchol, S.; Timoneda, N.; Fernández-Cassi, X.; Pérez-Cataluña, A.; Fernández-Bravo, A.; Moreno-Mesonero, L.; Moreno, Y.; Alonso, J.L.; Figueras, M.J.; et al. Metagenomic Analysis of Viruses, Bacteria and Protozoa in Irrigation Water. Int. J. Hyg. Environ. Health 2020, 224, 113440. [Google Scholar] [CrossRef] [PubMed]
- Dettling, A.; Wedel, C.; Huptas, C.; Hinrichs, J.; Scherer, S.; Wenning, M. High Counts of Thermophilic Spore Formers in Dairy Powders Originate from Persisting Strains in Processing Lines. Int. J. Food Microbiol. 2020, 108888. [Google Scholar] [CrossRef] [PubMed]
- Kable, M.E.; Srisengfa, Y.; Xue, Z.; Coates, L.C.; Marco, M.L. Viable and Total Bacterial Populations Undergo Equipment- and Time-Dependent Shifts during Milk Processing. Appl. Environ. Microbiol. 2019, 85, e00270-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, S.A.; Brooks, J.D.; Rakonjac, J.; Walker, K.M.; Flint, S.H. The Formation of Spores in Biofilms of Anoxybacillus flavithermus. J. Appl. Microbiol. 2009, 107, 1012–1018. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, D.J.; Daly, D.; O’Sullivan, O.; Burdikova, Z.; Vana, R.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; McSweeney, P.L.H.; et al. Thermus and the Pink Discoloration Defect in Cheese. mSystems 2016, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Indicator 1 | Plant A | Plant B |
|---|---|---|
| pH | 8.23 ± 0.07 b | 12.45 ± 0.19 a |
| TS (g/L) | 0.50 ± 0.04 b | 3.12 ± 0.24 a |
| Ash (g/L) | 0.16 ± 0.0 b | 0.73 ± 0.03 a |
| Lactose (g/L) | 0.33 ± 0.03 b | 2.22 ± 0.18 a |
| Protein (g/L) | 0.01 ± 0.00 b | 0.15 ± 0.00 a |
| Fat (g/L) | 0.00 ± 0.00 a | 0.03 ± 0.02 a |
| COD (mg/L) | 21.50 ± 0.71 b | 354.50 ± 2.12 a |
| BOD5 (mg/L) | 4.50 ± 0.71 | ND 2 |
| Component 1 | Acid CW | Alkaline CW | ||
|---|---|---|---|---|
| Plant A | Plant B | Plant A | Plant B | |
| pH | 1.82 ± 0.06 c | 1.17 ± 0.01 d | 12.59 ± 0.01 a | 12.31 ± 0.10 b |
| EC (μS/cm) | 5.35 ± 0.10 d | 14.25 ± 0.13 c | 67.97 ± 2.70 a | 30.06 ± 0.65 b |
| TS (g/L) | 0.74 ± 0.04 c | 0.93 ± 0.02 d | 28.06 ± 0.38 a | 9.59± 0.56 b |
| TSS (g/L) | 0.02 ± 0.00 d | 0.01 ± 0.00 c | 0.19 ± 0.06 b | 0.21± 0.02 a |
| Ca2+ (mg/L) | 12.22 ± 1.45 d | 177.04 ± 0.43 a | 45.15 ± 1.76 b | 18.81 ± 1.98 c |
| Mg2+ (mg/L) | 1.89 ± 0.17 b | 7.07 ± 0.05 a | 0.82 ± 0.10 c | 0.23 ± 0.03 d |
| K+ (mg/L) | 0.76 ± 0.09 d | 1.57 ± 0.06 c | 9.41 ± 0.26 a | 6.48 ± 0.07 b |
| Na+ (mg/L) | 155 ± 13 c | 23 ± 4 d | 10,665 ±191 a | 4033 ± 46 b |
| COD (mg/L) | 25 ± 1 d | 26 ± 0 c | 1500 ± 7 a | 475 ± 62 b |
| BOD5 (mg/L) | ND 2 | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alalam, S.; Ben-Souilah, F.; Lessard, M.-H.; Chamberland, J.; Perreault, V.; Pouliot, Y.; Labrie, S.; Doyen, A. Characterization of Chemical and Bacterial Compositions of Dairy Wastewaters. Dairy 2021, 2, 179-190. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2020016
Alalam S, Ben-Souilah F, Lessard M-H, Chamberland J, Perreault V, Pouliot Y, Labrie S, Doyen A. Characterization of Chemical and Bacterial Compositions of Dairy Wastewaters. Dairy. 2021; 2(2):179-190. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2020016
Chicago/Turabian StyleAlalam, Sabine, Farah Ben-Souilah, Marie-Hélène Lessard, Julien Chamberland, Véronique Perreault, Yves Pouliot, Steve Labrie, and Alain Doyen. 2021. "Characterization of Chemical and Bacterial Compositions of Dairy Wastewaters" Dairy 2, no. 2: 179-190. https://0-doi-org.brum.beds.ac.uk/10.3390/dairy2020016