Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics?
by
 ,
,
Richard K. F. Unsworth
1,2,* ,
,
Alex Higgs
1, 
Bettina Walter
1,
Leanne C. Cullen-Unsworth
2,3,
Isabella Inman
1 and
Benjamin L. Jones
1,4 1
Project Seagrass, 33 Park Place, Cardiff CF10 3BA, UK
2
Seagrass Ecosystem Research Group, College of Science, Wallace Building, Swansea University, Swansea SA2 8PP, UK
3
Sustainable Places Research Institute, Cardiff University, 33 Park Place, Cardiff CF10 3BA, UK
4
Department of Ecology, Environment and Plant Sciences, Stockholm University, 106 91 Stockholm, Sweden
*
Author to whom correspondence should be addressed.
Oceans 2021, 2(1), 162-178; https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010010
Submission received: 25 September 2020
/
Revised: 11 December 2020
/
Accepted: 9 February 2021
/
Published: 12 February 2021
(This article belongs to the Special Issue Microplastics in Marine Environments: Occurrence, Distribution and Effects)
Abstract
:A growing body of research is documenting the accumulation of microplastics within marine sediments around the world. The hydrodynamic influences of seagrasses in coastal environments are shown to increase sedimentation of finer particles and as a result there has been speculation that this attribute will lead to seagrass meadows acting as a site of elevated microplastic contamination. To date a range of localised studies have provided conflicting answers to this hypothesis. Seagrass meadows provide multiple ecosystem services including vital support roles for a range of fisheries; therefore, there are considerable human health implications for understanding their role as sinks of microplastics. This research investigated the abundance and diversity of microplastics present in temperate North Atlantic seagrass meadow sediments relative to unvegetated sediments and examined how they correlate with the meadow structure and the sediment type. We also placed this data in the context of the current knowledge of microplastics in seagrass sediments through a global meta-analysis of published data. Eight seagrass meadows and adjacent unvegetated sites around the UK were sampled to test for the abundance of microplastic particles in the sediment. Microplastics were found in 98% of the samples, with fibres making up 91.8% of all microplastics identified. Abundance was recorded to overall be 215 ± 163 microplastic particles (MP) kg−1 Dry Weight (DW) of sediment in seagrass and 221 ± 236 MP kg−1 DW of sediment in unvegetated habitats. There were no significant differences found between the number of MP with respect to vegetation. We report evidence of the almost ubiquitous contamination of seagrass sediments with microplastics both in the UK and globally but find that the contamination reflects a general build-up of microplastics in the wider environment rather than becoming concentrated within seagrass as an enhanced sink. Microplastic build up in sediments is hypothesised to be the result of local hydrodynamics and plastic sources rather than the result of elevated habitat level concentration. Although not of a higher abundance in seagrass, such contamination in seagrass is of cause for concern given the high dependency of many species of fish on these habitat types and the potential for plastics to move up the food chain.
Keywords:
Zostera; sediment; marine; microbeads; pollution; eelgrass; seagrass; seascape; coastal; intertidal1. Introduction
Seagrass meadows provide multiple ecosystem services, including creation of spawning grounds for a variety of fish species, uptake and storage of carbon and coastal protection [1,2]. A significant basis for many of these services is the creation of a three-dimensional environment that reduces the velocity of water whilst increasing sedimentation and aiding particle trapping [3,4]. However, these highly productive ecosystems are vulnerable to a plethora of anthropogenic threats, including increased nutrient enrichment, poor water quality, heightened disease prevalence, physical disturbances and marine pollution [5,6].
Simultaneously, there has been increased recognition and concern of plastic pollution in the marine environment and concerns that seagrasses may act as plastic traps [7]. Manufacturing of plastic has increased exponentially since its discovery, with annual production increasing from 1.5 million tonnes in the 1950′s to 359 million tonnes in 2018 [8]. Formed from synthetic organic polymers, plastics are highly durable and are now considered to be ubiquitous in the oceans [9]. Plastic pollution has risen with increasing production, with up to 90% of marine litter and an estimated 4.8–12.7 million tonnes of plastic waste entering the oceans every year, with river systems being identified as the primary vector for transport [10,11,12].
Fragmentation and weathering of plastic waste commonly leads to the formation of microplastics—plastic particles ≤5 mm diameter [13]. The primary source of microplastics appears to be the effluent waste of washing machines, with a single clothing item expelling up to 1900 fibres per wash [14]. Microplastics occupy the same size fraction as sediments and are now found in benthic habitats globally, with sediments being suggested to be a long-term sink for microplastics [9,15]. Although the first reports of microplastics in the oceans date back as far as the 1970′s, it was not until 2004 that the first case of microplastics was reported in sediments [13]; given this knowledge, alongside previous studies suggesting a relationship between sediment grain size and microplastic prevalence, and the enhanced capacity of seagrass to trap particles [4] it seemed appropriate to test whether seagrass habitats also lead to heightened abundance of microplastic particles [10,16].
A growing body of research exists looking into the accumulation of microplastics within sediments of a variety of habitat types around the world, cumulatively highlighting the spread of this pollutant [15]. Plastic particles that accumulate in sediments are readily accessible for ingestion by benthic organisms, which then have the potential to bio-accumulate in the food chain [17]. Studies have highlighted the implications of microplastics entering the food chain at lower trophic levels [18].
Seagrass beds form from flowering plants, and as a result of their thick tall canopy, are known to increase sedimentation of particles as well as reduce the chances of particle re-suspension, with one study highlighting seagrasses to buffer sediment re-suspension by three times as much when compared to a non-vegetated sea floor [3,19]. Sediment retention by seagrass beds is a result of the reduction in water velocity and energy dissipation by plant canopies. Seagrass beds in the UK are formed of two different Zostera species (marina and noltii) with both species being regarded as net sinks of suspended matter [20].
More specifically, the hydrodynamic influences of seagrasses are shown to increase sedimentation of finer particles, with a number of recent studies indicating the presence of microplastics in seagrass sediments, particularly in subtidal environments [7,21]. Other studies have also highlighted high-energy coastal waters’ tendency to accumulate microplastics, with greater concentrations in finer sediments [22]. Additionally, a number of studies have examined how seagrass leaves provide a means of collecting microplastic particles, offering an additional mechanism for their potential concentration [23,24]. Research on plastic trapping in marine coastal vegetated ecosystems is still in its infancy and mainly carried out in mangrove forests [7]. Within seagrass meadows specifically, although documented to be present, we still know little about the concentrations and variability of microplastics in their sediments and the mechanisms driving any variability. Research however has begun to show that this presence may have negative impacts upon the seagrass food web [25,26,27].
This research investigated the abundance and diversity of microplastics present in temperate North Atlantic seagrass meadow sediments relative to unvegetated sediments and examined how they correlate with the meadow structure and the sediment type. We also placed these data in the context of the current knowledge of microplastics in seagrass sediments through a global meta-analysis of published data.
2. Materials and Methods
2.1. Sampling Locations
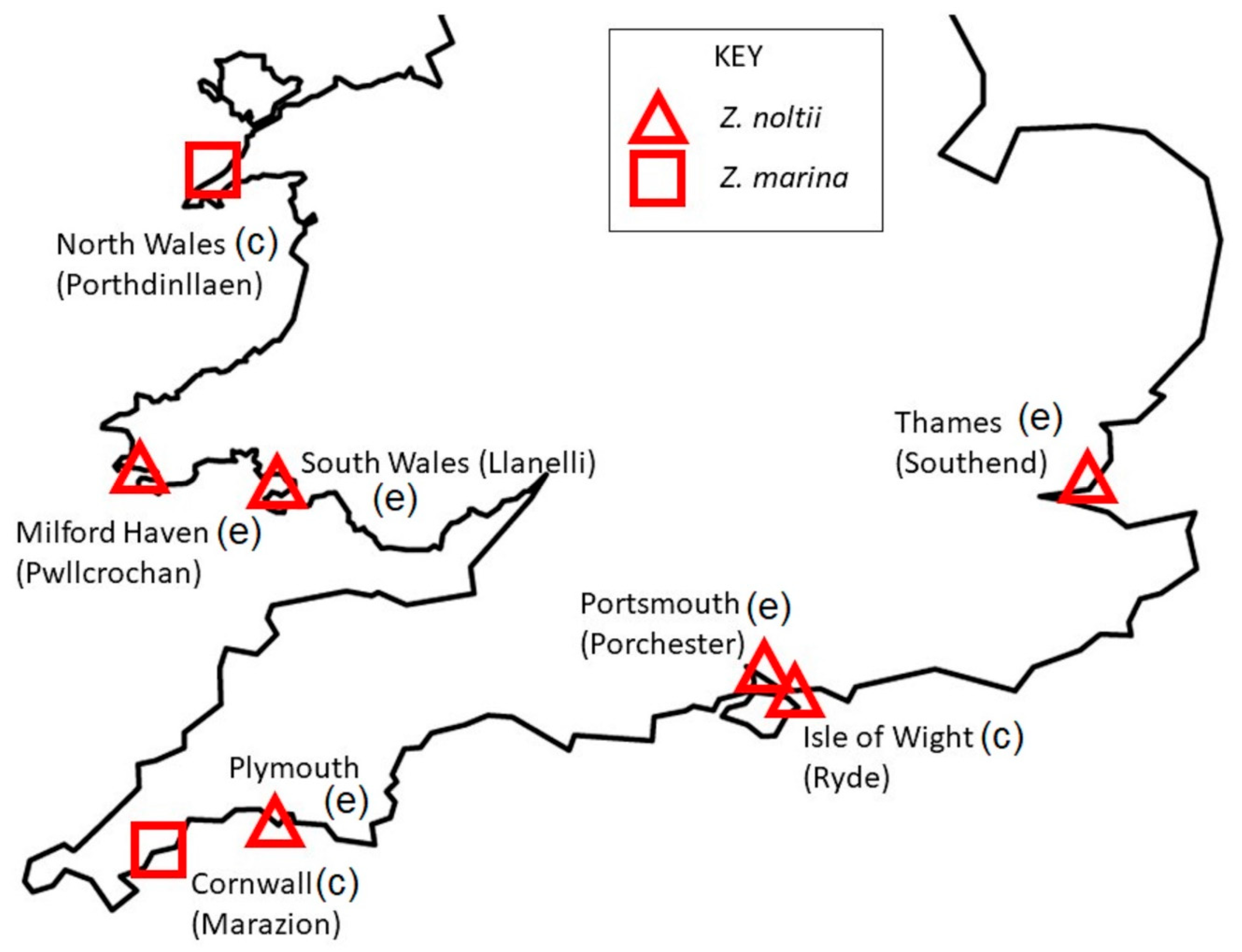
Between the 14th of August and 27th of October 2018, eight seagrass meadows and adjacent unvegetated sites in Southern England and Wales were sampled to test for the abundance of microplastic particles in the sediment (Figure 1). The study was conducted within meadows of two species of Eelgrass, Zostera marina (ZM) and Zostera noltii (ZN), and sites for the study were selected to all be inter-tidal and split between varied coastal or estuarine environments (Figure 1). In order to assess whether the frequency of microplastics is higher in seagrass habitats, a non-vegetative area at each site was sampled for comparison. It was critical that the non-vegetative area was distanced far enough from the hydrodynamic influences of the seagrass bed, whilst remaining close enough so that external conditions and influences are controlled. Selection of the non-vegetative area was determined on site and based on accessibility, potential external influences (e.g., river outputs) and distance from the high-tide mark. Distances between seagrass beds and non-vegetated areas was at least 75 m. Non vegetated sites were chosen based on locations of no previous evidence of seagrass at the site using available site data [28].
2.2. Sediment Sampling
At each location five 0.25-m2 quadrats were haphazardly placed (at least 5 m away from each other) within the centre of the seagrass meadow. In the centre of each quadrat one single pre-washed corer (5 cm diameter and 20 cm length) was used to collect the upper 5 cm of sediment. A further 5 cores were taken haphazardly in unvegetated habitat at least 75 m from the meadow using the same procedure. Previous studies have found flow attenuation and sediment deposition to be density dependent [29]. Sampling of all the seagrass was conducted in the centre of the meadow, where density is expected to be greater and theoretically where microplastic abundance should be highest [30]. A 5-cm sediment depth core was taken as previous studies have highlighted that a core must be between 4–5 cm deep in order to assess the standing stock of microplastics in marine sediments [31]. This method follows numerous similar analysis within marine sediments [22,23,32].
An additional core was taken from the centre of two of the five quadrats (in each habitat type). All cores were stored in a sealed and labelled zip-lock bag in laboratory freezers at −30 °C until analysis. To minimise contamination, sampling took place into the wind to avoid any synthetic fibres from clothing blowing into the sample [33] and the same corer (washed in between collections) was used for all samples.
2.3. Seagrass Assessment
Prior to the collection of the sediment cores, the percentage cover of seagrass was assessed within the quadrat. This was determined by taking a calculated visual estimation, which is deemed to be an accurate and repeatable method to measure percentage cover. This used available Seagrass-Watch percentage cover guides [34,35]. Canopy height measurements of the seagrass were taken by measuring five seagrass shoots within each quadrat from the base of the shoot to the tip of the leaf (ignoring the tallest shoots), with an average height being calculated. A description of the sediment type was then assessed and recorded following the Seagrass-Watch guide [35].
2.4. Preparation of Sample and Zinc Chloride Solution
Sediment samples were placed in a drying oven in aluminium trays at temperatures between 35–40 °C until a constant weight was reached. Once dry, the samples were passed through a sieve stack and shaken for ten minutes. Sediments passed through an Endecotts sieve stack (>1 mm, 1–2 mm, 500 μm–1 mm, 250–500 μm, 25–250 μm). Each size fraction was weighted in order to classify the sediment composition at each site. Categorization of grain size was determined from the Wentworth scale and values for Fineness Modulus were calculated for each sample [36]. The site fractions were then added together again into one sample and stored in sealed containers until microplastic extraction.
The density of commonly produced plastics ranges from 800 to 1400 kg m−3, hence a zinc chloride solution of 1.5 g cm−3 was deemed appropriate for a density separation [37,38]. To achieve the chosen density, 0.972 kg of the salt were dissolved in 1 litre of ultrapure water. The solution was stirred for approximately an hour before being vacuum filtered through a 10-µm membrane to remove external contaminants prior to use. Zinc chloride was favoured over other salt solutions, such as sodium chloride and sodium iodide, as it is able to achieve a higher density whilst remaining relatively cost effective [39], and resulting in a more effective removal of plastics.
2.5. Sediment Microplastic Isolation (SMI) Unit—Assembly and Purging
In order to perform a rapid and efficient density separation of microplastics from each of the samples, a Sediment Microplastic Isolation (SMI) unit was constructed [39]. The unit was constructed by securing two 12-cm lengths of PVC pipe to a 63-mm ball valve, with a 63-mm pipe cap base. Prior to assembly, the individual components were thoroughly washed with clean freshwater to avoid contamination. The unit’s design ensured that all internal surfaces were smooth, allowing free movement of any plastic particles present.
Following assembly, 800 mL of filtered zinc chloride solution was added to the SMI unit. It was key to purge the ball valve by opening and closing the valve several times, ensuring the internal cavity was submerged to avoid air bubbles escaping upon valve opening when processing samples. The unit was topped back up and left for five minutes to allow any external contaminants to float to the surface, with the zinc chloride solution then being filtered into a clean flask for continued use (using Whatman™ 30-µm mesh filters, Whatman, Cytiva, UK).
2.6. Extraction of Microplastic from Samples
For each extraction, a subsample of dry sediment, a magnetic stir bar and 800 mL of zinc chloride solution was added to the prepared SMI unit. A magnetic stir plate was placed under the SMI unit and left to stir the sample for five minutes. The sample was then left to settle for 4 h until the supernatant was clear of sediment, allowing any microplastics present to surface. The valve was then carefully closed in order to separate any microplastics from the sediments. The headspace was filtered onto a 30-µm mesh filter (Whatman) and microplastic particles separated from the zinc chloride solution, with the solution being retained for the next sample. The mesh filter containing microplastics was rinsed into a petri dish, sealed and stored until stereomicroscope analysis. This process was repeated for both seagrass and non-seagrass sediment samples at each of the eight sites sampled. Blank analyses of all containers and wrappings were also conducted and found to not contain any contamination, necessitating no level of data correction.
Petri dishes were viewed under a stereomicroscope (with a digital PC viewer) at a 4.5× magnification. Microplastics were identified, isolated and then categorised according to clear guidelines [40]. The guidelines used describe a process of identifying under the microscope key characteristics in items that can be assessed to clearly distinguish microplastics from natural fibres and other natural organic and inorganic material whilst under the microscope. Items were therefore classified as microplastic under the basis that they contained no cellular or organic structures, they were small in size (largest dimension ≤5 mm), they exhibited clear and homogeneous colour throughout, they did not break when prodded with forceps, bent under heat, and the fibres were equally thick throughout their entire length. Whilst this method does not guarantee that all items were microplastics (e.g., see Suaria et al. 2020 [41]), care was taken to rigorously follow such classification throughout and we expect that classification errors were minimal if not non-existent. Microplastics were photographed for each sample and classified by quantifying the type (fibres, fragments, films and fibre bundles), as well as the colour.
2.7. Literature Review
In order to position our findings within the published literature we conducted a literature search for “All document types” on Web of Science and Google Scholar published before June 2020. The following search terms were used in all combinations (seagrass, plastic, microplastic, eelgrass, sea grass, submerged aquatic vegetation). The literature was then checked for studies of seagrass habitats and the quantification of plastics within them. All data from those studies were then tabulated and the mean effect size (Cohen’s d) determined for each study and across the dataset (Table 1). Cohen’s d is defined as the difference between the means divided by the pooled standard deviation [42].
2.8. Data Analysis
All summary data are presented as a mean ± sample standard deviation. Statistical analysis was conducted using PRIMER v7 and R version 3.6.0 [43]. Univariate analysis of microplastic abundance data was conducted using permutational analysis of variance (PERMANOVA+) using a Euclidean resemblance matrix. All PERMANOVA tests were nested and based on 9999 unrestricted permutations under a reduced model.
Using a linear mixed-effects model [44] with maximum likelihood estimation, we investigated the influence of vegetation (seagrass, non-vegetative) and environment type (coastal, estuarine) on microplastic abundance data. The linear mixed-effects model was fitted with the lmer() function in lme4 [45] and included the random effect of site to account for any biogeographic differences in environmental conditions and anthropogenic impacts. We report the fit of model using R2GLMM, calculated using the rsquared() in piecewiseSEM [46].
3. Results
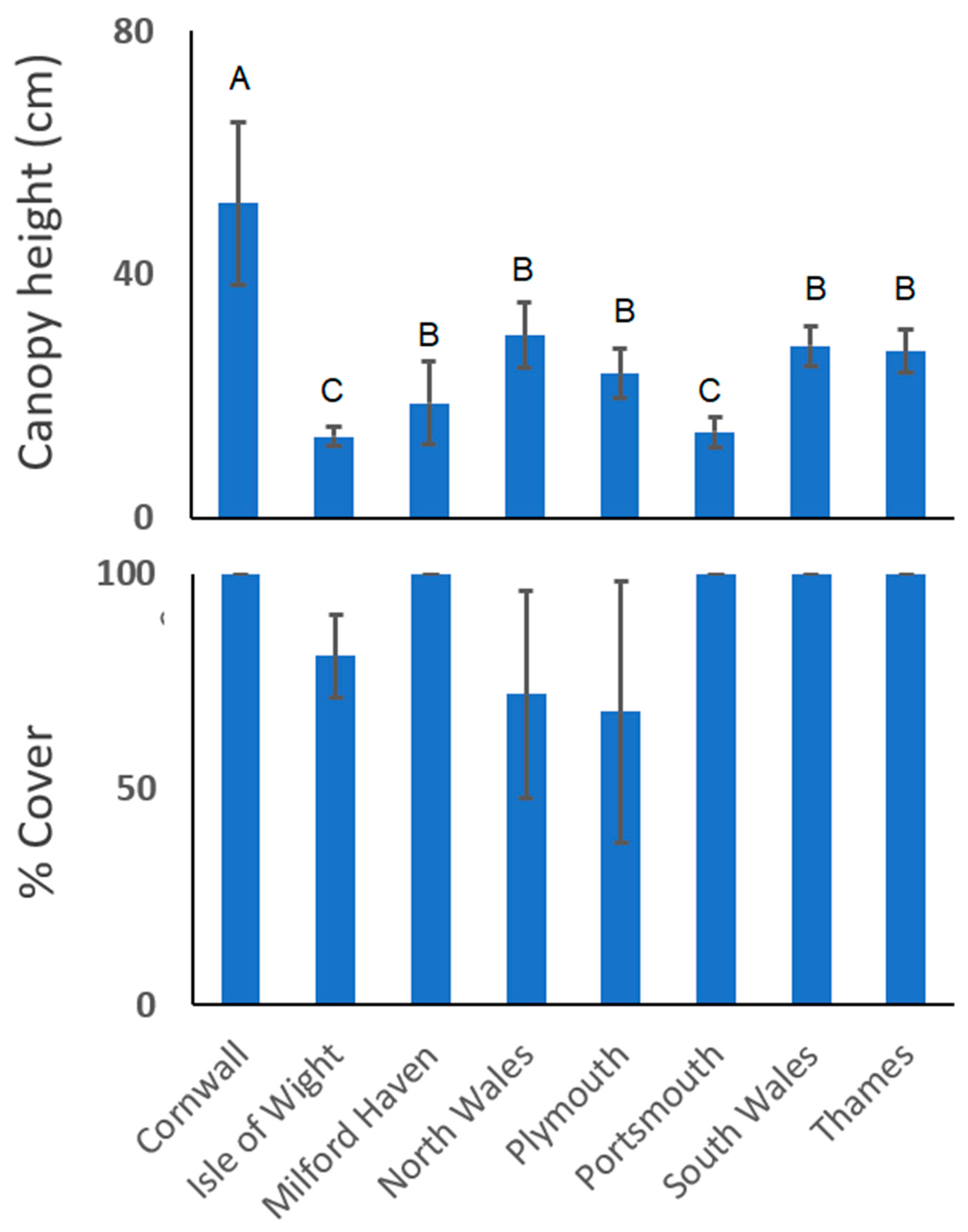
Canopy heights differed significantly between the eight sites (nested in environment) (PERMANOVA Pseudo F6,39 = 20.8, p < 0.001), with heights ranging from 13 to 51 cm (mean 26 ± 13 cm). No significant difference was found with respect to environment. Unsurprisingly, greatest canopy height was observed at a Zostera marina site, and shortest at two Zostera noltii sites (Isle of Wight and Portsmouth; Figure 2).
Seagrass cover between the sites was significantly different (PERMANOVA F6,39 = 3.16, p = 0.004) and ranged from 68% to 100% (mean 90.1% ± 18.5), with Isle of Wight (mean 81% ± 9.6), Plymouth (mean 68% ± 30.3) and North Wales (mean 72% ± 24.1) being the only sites not to display 100% seagrass cover within sampled quadrats (Figure 2).
Sediment type and composition differed both within (seagrass and non-vegetative) and between sites. Three sites (South Wales, Milford Haven and Thames) were dominated by silts/muds (<125 μm), in both the seagrass and non-vegetative areas. Two sites (Isle of Wight and Cornwall) were dominated by fine sands (125–250 μm) in both seagrass and non-seagrass areas. Fine–muddy sediments dominated the seagrass meadow in North Wales, whereas medium sized sands dominated the non-vegetative area. Mean sediment size (Fineness Modulus) did not significantly change with respect to site (Pseudo F4,54 = 3.83, p = 0.08) or habitat (nested in site) (Pseudo F5,54 = 0.48, p = 0.81).
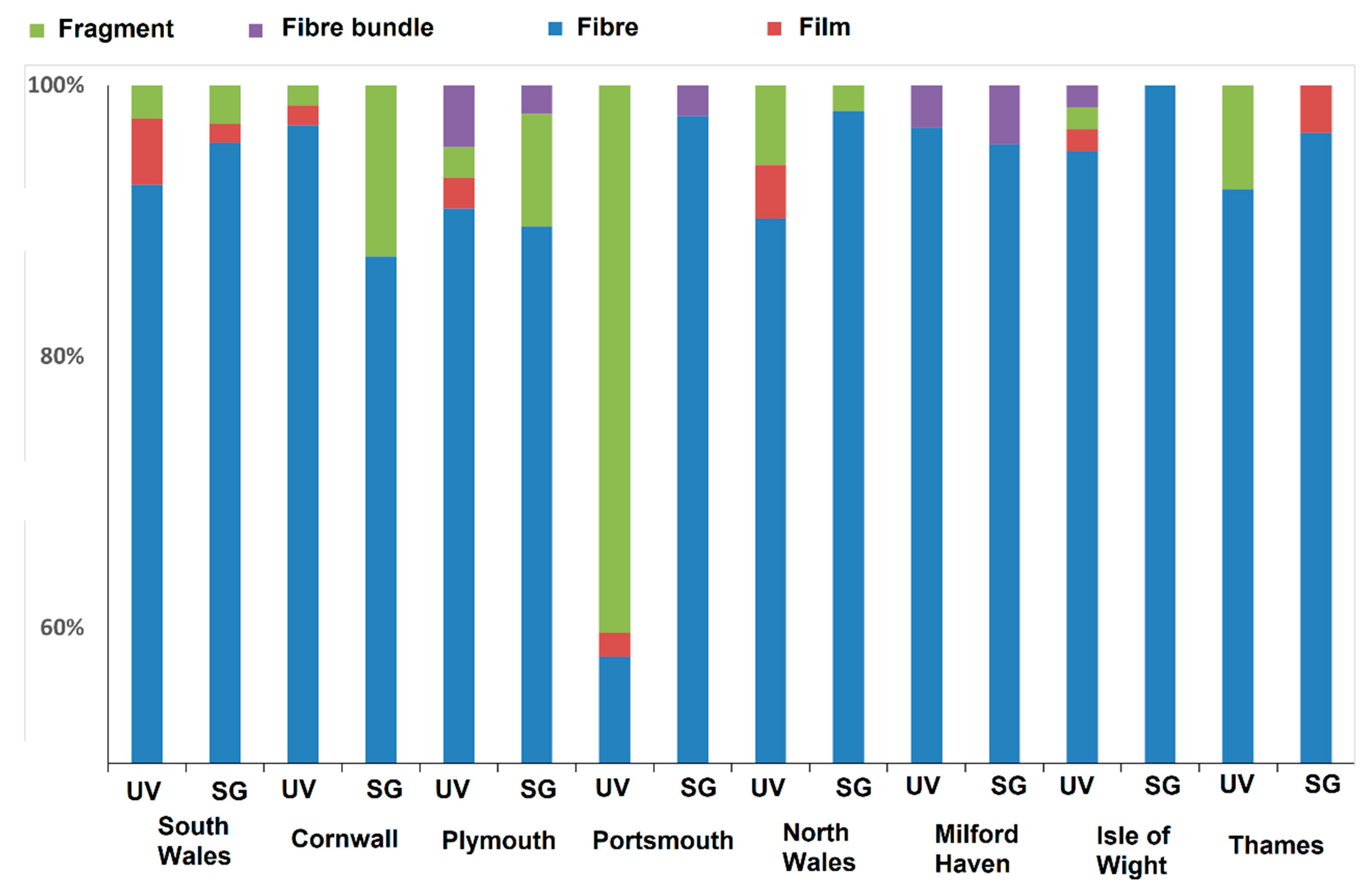
Plastics were found in 98% of the samples, with fibres making up 91.8% of all microplastics identified. Films (1%), fragments (6.6%) and bundles of fibres (0.6%) made up the remainder of identified plastics. The variation of microplastic type between seagrass and non-vegetated areas was minimal with no indication of any relationship between the two habitats (Figure 3). Microplastic abundance, colour and type ranged across the sites with blue fibres occurring most commonly in all samples (Figure 3 and Figure 4).
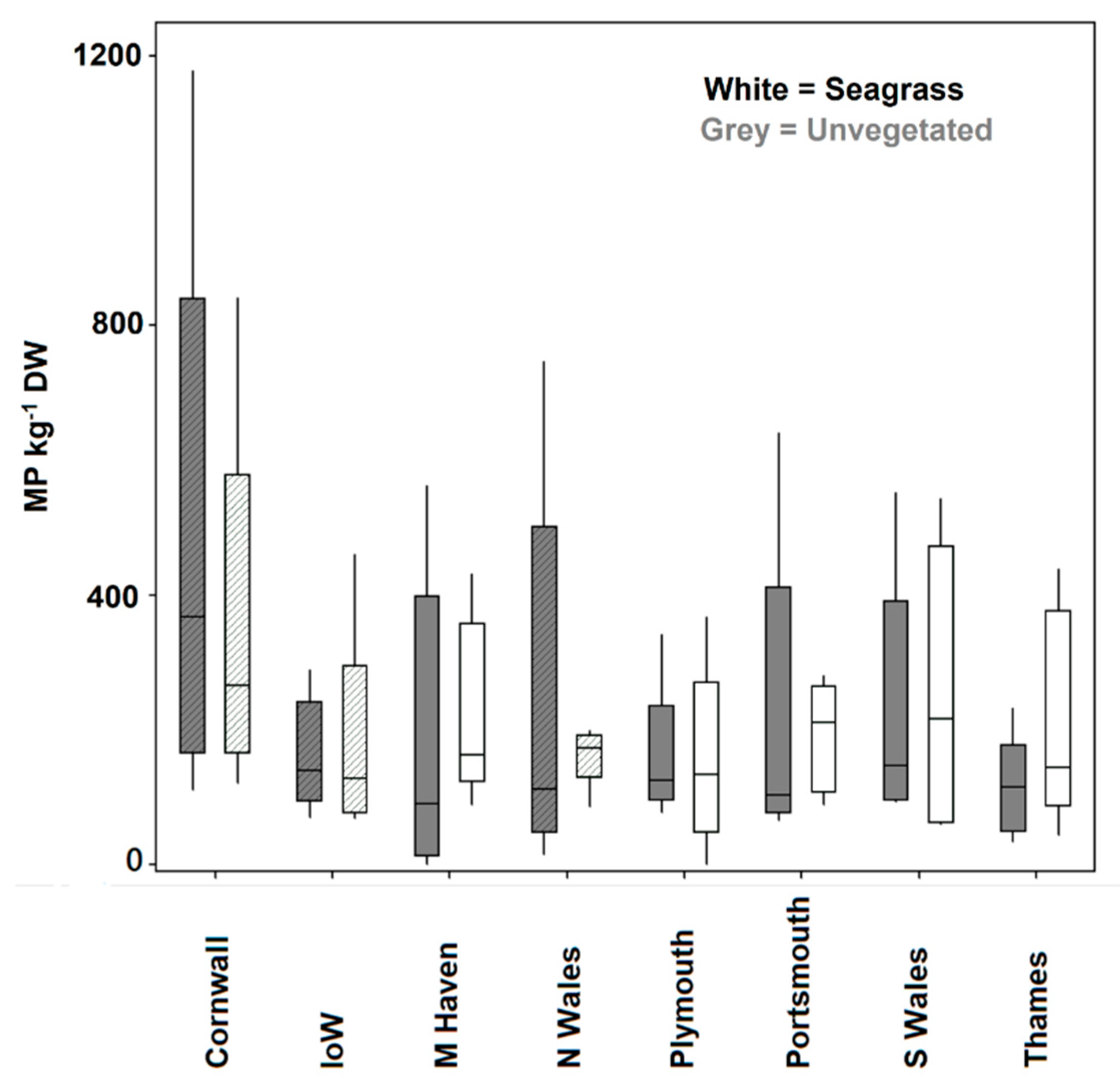
In total 1042 microplastic particles (MP) were identified across the 80 samples. On average there were 218 ± 202 MP kg−1 DW of sediment. This varied with respect to site and density. The highest number of particles was found in Cornwall (Marazion), at over double the average at 413 ± 344 MP kg−1 DW of sediment and lowest at 163 ± 129 particles MP kg−1 DW of sediment in the Thames (Southend On Sea). MP abundance was recorded to overall be 215 ± 163 MP kg−1 DW of sediment in seagrass and 221 ± 236 MP kg−1 DW of sediment in unvegetated habitats. There were no significant differences (PERMANOVA) found between the number of MP with respect to vegetation (nested in site—seagrass versus non vegetated) (Pseudo F7,78 = 0.95, p = 0.48) and site (Pseudo F6,78 = 071, p = 0.8). In addition, using a linear mixed-effects model, we found that there was no measurable effect of vegetation (including cover and canopy height) (seagrass, non-vegetated) or environment (coastal, estuarine) on MP kg−1 DW when controlling for site (R2 = 0.02). However, four of the eight seagrass sites show elevated densities of MP kg−1 DW compared to a non-vegetated seabed (Figure 5). In addition, no relationship was found between MP abundance and the sediment type (measured as fineness modulus).
Eight studies (including our study) were found in the literature that have examined microplastics within seagrass meadows; only six of these were recorded to quantify microplastics in seagrass sediments (Table 1). These six (including our study) were spread across four continents and seven countries. Only five of the studies compared seagrass sediments against unvegetated habitats. They covered microplastics in nine species of seagrass. Microplastics in seagrass sediments were extracted mostly using density separation in a salt media (ZnBr, ZnCl, NaCl, or CaCl2) but methods varied widely. All studies recorded microplastics to be present in >90% of seagrass sediments with exception of the study from Indonesia that recorded it in 27% of samples. Of 140 samples recorded in seagrass globally within the literature 110 (64%) of these were found to contain microplastics. The majority of those not containing microplastics were recorded at the Indonesian study which was on a small island offshore whereas all other studies were conducted in coastal areas. Of the six studies, most had between one and three locations that were geographically in close-proximity and had a small number of samples (with the exception of the Indonesian study and our study). Fibres (originating from clothing or netting) were the dominant form of microplastics in all samples globally.
Abundance of microplastics relative to sediment weight found average microplastic abundance to range from 18 to 780 MP kg−1 DW. Our study is within a central point of this range and is similar to the abundance recorded in Scotland [23]. Our study is spatially the most extensive study of its type to date with 8 locations and 80 samples. Although, our study recorded no statistical difference overall between microplastic abundance in seagrass and unvegetated habitats the studies in Scotland and China found a significant difference. Calculation of the mean effect size (Cohens d) found a mean effect size of 3.0 ± 3.4; the large standard deviation illustrates the high variability within the data. In addition, three studies recorded microplastics to be present on seagrass leaves and in the seagrass food web [23,24,27].
4. Discussion
Here we report evidence at local (UK) and global scales the almost ubiquitous contamination of seagrass meadows with marine microplastic. In the UK, 100% of sediments irrespective of their habitat type were contaminated with microplastics. At a global scale we find that the sediments of 17 meadows spanning 7 countries and 4 continents all contain microplastics. Most samples were contaminated with >90% consistency, with the exception of a remote island sites in Indonesia where only 27% of samples were contaminated. While some studies have highlighted the presence of microplastics at elevated concentrations within seagrass ecosystems relative to unvegetated habitats, all previous studies have been spatially limited and presented conflicting information as to whether the accumulation of plastic particles is heightened in the sediments of seagrass meadows, especially in the context of non-vegetated sea floor [25,26,27]. We conclude that at a local and global level there is limited field evidence that seagrass meadows do accumulate microplastics at higher concentrations than surrounding sediments. We hypothesise that local physical and anthropogenic factors as well as the type of type of microplastic are of greater significance in determining where microplastics end up and in some locations, this may lead to a higher concentration of microplastics in seagrass sediments.
We found microplastic items in UK seagrass sediments (215 ± 163 MP kg−1 DW) to be an order of magnitude higher than those recorded within a less spatially expansive surveys conducted in Portugal and the US, but at a similar level to those within a Scottish survey and at sites in Portugal [23]. Sites in China were found to have much higher densities of MP than any other locality but utilised potentially more efficient extraction methods that may have enhanced concentrations relative to other studies. The UK levels in seagrass across eight sites were not significantly different to those in non-vegetated benthos, indicating these elevated levels to be the result of geographic location and the anthropogenic site pressures rather than habitat; however, there was some site level variability within this assessment. Some sites had higher densities in unvegetated whilst some had higher densities with vegetated. This is a surprising result given the heightened capacity of seagrass to trap particles and bioengineering their environment [4] but is consistent with the variability recorded within the literature. Although this result does not support our hypothesis, it is in accordance with the current literature that highlights the widespread distribution of this emerging pollutant within coastal benthic habitats [7,14]. Given the lighter molecular weight of microplastic particulates (particularly the dominant fibres present in most studies) relative to many inorganic sediment particles, their slower sink rate may therefore result in no preferential trapping by seagrass.
A study by Goss et al. [25] proposed several mechanisms for how microplastics become entrapped on seagrass blades of Thalassia testudinum in the Caribbean. These mechanisms all related to the seagrass blades “trapping” microplastics in the water column—for example, microplastics sticking to blades via adhesive biofilms. Recent high profile evidence from the Mediterranean has also found that the creation of so called as ‘Seaballs or Neptune balls’ leads to the trapping of microplastics [47]. Such seaballs of fibrous seagrass material tend to be much more common in the Mediterranean than in other localities.
We proposed that a taller canopy height would increase the ability of the seagrass to trap microplastics due to a greater surface area of blades. Eventually, this would lead to heightened microplastic abundance within seagrass bed sediments as seagrass blades die and degrade with plastics remaining in a sediment sink. Our study data and the wider global meta-analysis does not support this assumption. The variable UK findings of variable trapping with respect to seagrass cover and canopy agree with those from seagrass sediments in one of the most detailed previous studies to date from Portugal [7]. In the present study, fibres, fragments, films and bundles of fibres were found in the sediments of both seagrass and non-vegetative areas. Of these identified plastic types, fibres were found in the greatest proportions (91.8% total occurrence), which is consistent with the current literature that highlights plastic fibres to be the most frequently found type of microplastic pollution in the marine environment [14,48]. Microfibers from synthetic origin are estimated to constitute more than 80% of marine microplastics and contaminate shores on a global scale with wastewater effluent systems being the primary vector of transport [14,48,49].
An experimental controlled lab study has revealed that seagrass of increasing density does potentially trap more microplastic particles, but that the nature of such an effect is dependent upon both the density of the microplastics and the water flow velocity [50]. Lighter fractions (e.g., microfibres) are less likely to be caught in seagrass relative to higher density particles. The denser particles become trapped at a far higher rate due to their faster sinking rate. Given that such a high proportion of microplastics in seagrass in the present data and globally is comprised of light weight microfibers it is potentially therefore not surprising to record seagrass to not show an enhanced microplastic sink role within basic field assessments.
The high levels of microfibers present within the sediment samples is concordant with the hypothesis of [51] which suggested that transitional systems (such as estuaries) are more prone to fibre contamination due to their close proximity to wastewater systems and human populations. Of the fibres identified across the eight study sites it is likely that many of them originated from the wastewater effluent systems or from the fragmentation of fishing gear, which is also considered to be a primary contributor of plastic pollution—with FAO estimating 640,000 tonnes of fishing gear being lost annually [14,52].
The present study found no evidence that microplastic concentrations increased with a decreasing sediment grain size. We hypothesised that where water movement was reduced, creating decreased particle size in sediments, microplastic deposition would similarly be greater given their light weight. This was contrary to other studies findings that highlight increasing concentrations of alternate pollutants (heavy metals) and organic material with respect to changing particle size [53,54]. Other studies have also found no clear relationship between a smaller sediment fraction (clays) and microplastic accumulation, with it being suggested that aggregation with organic material, are key in influencing microplastic movement and distribution [10,15,55]
The present findings indicate that numerical simulation models of the movement of microplastics in and around seagrass needs to be conducted to understand the small scale interacting influence of habitat, hydrodynamics and sediments on microplastics [56]. This would enable improved understanding of the influence of physical (e.g., hydrodynamics) and anthropogenic (e.g., proximity to point sources of microplastics) to MP distribution.
The locality of the eight UK study sites differed in terms of their remoteness and scale of surrounding human settlement, with some sites being identified as more remote (and of low human population) than others. It was expected that sites such as the Thames would see significantly higher levels of plastic pollution compared to sites such as that at the North Wales site (in an Special Area of Conservation) or Marine Conservation Zone sites such as that in Cornwall (Marazion), due principally to the high human population and related activity. Given that no difference was seen between the UK sites, we hypothesise that within the sample area human activity has no measurable impact. Previous studies have highlighted the widespread nature of microplastic particles, with them being found in both urbanized and Marine Protected Areas [55]. Regardless of the current lack of understanding surrounding microplastics and their distributions, the present study adds to a growing body of research indicating that natural or protected areas are not excluded from microplastic pollution and that combating this problem requires an integrated coastal management approach targeting the source of microplastics.
Our current knowledge on the sources, distributions and sinks of microplastics remains limited to only a handful of data points globally [7,21], however the present study adds to a growing body of evidence highlighting the potential for seagrass sediments to act as a sink of microplastic particles, but importantly not as an enhanced sink relative to other unvegetated habitats. Given that carbon remains locked in seagrass sediments for many millennia [57], it is therefore likely to be the case that microplastics within such an environment will remain intact and stored for long periods too, given that there is less sediment resuspension in seagrass meadows [29].
Although the present study finds no evidence of elevated microplastic levels in seagrass, relative to other seabed habitats, the near ubiquitous presence of microplastics in seagrass sediments is of cause for concern given the higher density of juvenile fish of economic value (e.g., Gadus morhua) present in these habitats [58,59,60] and their widespread use as a fisheries habitat [61]. An extrapolation of the present data onto a per unit area basis indicates that UK seagrass at a known extent of 8493 hectares [62] may contain approximately 92 billion particles of microplastics. Studies have identified the potential for microplastic bioaccumulation starting at lower trophic levels [25,63] and the ingestion of microplastics has been shown in seagrass-associated fauna with it being suggested that ingestion can lead to physical and chemical effects on organisms [27,52]. Microplastic contamination in seagrass sediments therefore could pose currently unidentified problems to human health and food safety.
5. Conclusions
In conclusion, we find evidence of the almost ubiquitous contamination of seagrass sediments with microplastics both in the UK and globally. However, this contamination is found to reflect a build-up of microplastics in the wider environment rather than focused on or concentrated in seagrass. Such contamination in seagrass is of cause for concern given the high dependency of many species of fish on these habitat types and the potential for plastics to move up the food chain. Additional sampling will prove to be critical in order to determine the factors that influence the build-up on microplastics in marine sediments.
Author Contributions
Conceptualization, R.K.F.U., L.C.C.-U., B.L.J.; Methodology, R.K.F.U., A.H., B.W., L.C.C.-U., B.L.J.; Lab analysis, A.H., B.W.; Data Analysis, A.H., R.K.F.U., I.I., B.L.J.; Investigation, A.H., I.I.; Data Curation, A.H.; Writing—Original Draft Preparation, A.H., R.K.F.U.; Writing—Review & Editing, R.K.F.U., B.L.J., B.W.; Funding Acquisition, L.C.C.-U., B.L.J. All authors have read and agreed to the published version of the manuscript.
Funding
Initial funding for this study was provided to BJ and LCU by the Sustainable Places Research Institute, Cardiff University.
Informed Consent Statement
Not Applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass ecosystem services and their variability across genera and geographical regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unsworth, R.K.F.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass meadows support global fisheries production. Conserv. Lett. 2019, 12, e12566. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Fisher, J.S.; Zieman, J.C. Influence of the seagrass, IZostera marina L., on current flow. Est. Coast. Shelf Sci. 1982, 15, 351–364. [Google Scholar] [CrossRef]
- Maxwell, P.S.; Eklöf, J.S.; van Katwijk, M.M.; O’Brien, K.R.; de la Torre-Castro, M.; Boström, C.; Bouma, T.J.; Krause-Jensen, D.; Unsworth, R.K.F.; van Tussenbroek, B.I.; et al. The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems—A review. Biol. Rev. 2017, 92, 1521–1538. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Randall Hughes, A.; Kendrick, G.A.; Judson Kenworthy, W.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, L.; Nicastro, K.R.; Zardi, G.I.; de los Santos, C.B. Species-specific plastic accumulation in the sediment and canopy of coastal vegetated habitats. Sci. Total Environ. 2020, 723, 138018. [Google Scholar] [CrossRef]
- Statista. Global Plastic Production. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/282732/global-production-of-plastics-since-1950/ (accessed on 1 May 2019).
- Van Cauwenberghe, L.; Devriese, L.; Galgani, F.; Robbens, J.; Janssen, C.R. Microplastics in sediments: A review of techniques, occurrence and effects. Mar. Environ. Res. 2015, 111, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.A.; Galloway, T.S.; Thompson, R.C. Spatial Patterns of Plastic Debris along Estuarine Shorelines. Environ. Sci. Technol. 2010, 44, 3404–3409. [Google Scholar] [CrossRef]
- Derraik, J.G.B. The pollution of the marine environment by plastic debris: A review. Mar. Poll. Bull. 2002, 44, 842–852. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Claessens, M.; Vandegehuchte, M.B.; Janssen, C.R. Microplastics are taken up by mussels (Mytilus edulis) and lugworms (Arenicola marina) living in natural habitats. Environ. Poll. 2015, 199, 10–17. [Google Scholar] [CrossRef]
- Kazmiruk, T.N.; Kazmiruk, V.D.; Bendell, L.I. Abundance and distribution of microplastics within surface sediments of a key shellfish growing region of Canada. PLoS ONE 2018, 13, e0196005. [Google Scholar] [CrossRef] [Green Version]
- van Raamsdonk, L.W.D.; van der Zande, M.; Koelmans, A.A.; Hoogenboom, R.L.A.P.; Peters, R.J.B.; Groot, M.J.; Peijnenburg, A.A.C.M.; Weesepoel, Y.J.A. Current Insights into Monitoring, Bioaccumulation, and Potential Health Effects of Microplastics Present in the Food Chain. Foods 2020, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in bivalves cultured for human consumption. Environ. Poll. 2014, 193, 65–70. [Google Scholar] [CrossRef]
- Gacia, E.; Duarte, C.M. Sediment Retention by a Mediterranean Posidonia oceanica Meadow: The Balance between Deposition and Resuspension. Est. Coast. Shelf Sci. 2001, 52, 505–514. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Zieman, J.C.; Thayer, G.W.; Fisher, J.S. The role of current velocity in structuring eelgrass (Zostera marina L.) meadows. Est. Coast. Shelf Sci. 1983, 17, 367–380. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, X.; Xu, C.; Perianen, Y.D.; Hu, J.; Holmer, M. Seagrass beds acting as a trap of microplastics—Emerging hotspot in the coastal region? Environ. Poll. 2020, 257, 113450. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.D.; Sinclair, M.; Levi, C.J.; Reeves, S.E.; Edgar, G.J. Ubiquity of microplastics in coastal seafloor sediments. Mar. Poll. Bull. 2017, 121, 104–110. [Google Scholar] [CrossRef]
- Jones, K.L.; Hartl, M.G.J.; Bell, M.C.; Capper, A. Microplastic accumulation in a Zostera marina L. bed at Deerness Sound, Orkney, Scotland. Mar. Poll. Bull. 2020, 152, 110883. [Google Scholar] [CrossRef]
- Seng, N.; Lai, S.; Fong, J.; Saleh, M.F.; Cheng, C.; Cheok, Z.Y.; Todd, P.A. Early evidence of microplastics on seagrass and macroalgae. Mar. Freshw. Res. 2020. [Google Scholar] [CrossRef]
- Goss, H.; Jaskiel, J.; Rotjan, R. Thalassia testudinum as a potential vector for incorporating microplastics into benthic marine food webs. Mar. Poll. Bull. 2018, 135, 1085–1089. [Google Scholar] [CrossRef]
- Pietrelli, L.; Di Gennaro, A.; Menegoni, P.; Lecce, F.; Poeta, G.; Acosta, A.T.R.; Battisti, C.; Iannilli, V. Pervasive plastisphere: First record of plastics in egagropiles (Posidonia spheroids). Environ. Poll. 2017, 229, 1032–1036. [Google Scholar] [CrossRef]
- Remy, F.; Collard, F.; Gilbert, B.; Compère, P.; Eppe, G.; Lepoint, G. When Microplastic Is Not Plastic: The Ingestion of Artificial Cellulose Fibers by Macrofauna Living in Seagrass Macrophytodetritus. Environ. Sci. Technol. 2015, 49, 11158–11166. [Google Scholar] [CrossRef] [PubMed]
- UNEP-WCMC; Short, F.T. Global Distribution of Seagrasses (Version 6.0). Sixth Update to the Data Layer Used in Green and SHORT (2003). Cambridge (UK): UN Environment World Conservation Monitoring Centre. 2018. Available online: http://data.unep-wcmc.org/datasets/7 (accessed on 1 December 2020).
- Ganthy, F.; Soissons, L.; Sauriau, P.G.; Verney, R.; Sottolichio, A. Effects of short flexible seagrass Zostera noltei on flow, erosion and deposition processes determined using flume experiments. Sedimentology 2015, 62, 997–1023. [Google Scholar] [CrossRef] [Green Version]
- Lanuru, M.; Fitri, R. Sediment deposition in South Sulawesi sea-grass bed. Mar. Res. Indones. 2008, 33, 221–224. [Google Scholar] [CrossRef]
- Martin, J.; Lusher, A.; Thompson, R.C.; Morley, A. The Deposition and Accumulation of Microplastics in Marine Sediments and Bottom Water from the Irish Continental Shelf. Sci. Rep. 2017, 7, 10772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.; Sobral, P. Plastic marine debris on the Portuguese coastline: A matter of size? Mar. Poll. Bull. 2011, 62, 2649–2653. [Google Scholar] [CrossRef]
- Löder, M.G.J.; Gerdts, G. Methodology Used for the Detection and Identification of Microplastics—A Critical Appraisal. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 201–227. [Google Scholar]
- Dethier, M.N.; Graham, E.S.; Cohen, S.; Tear, L.M. Visual versus random-point percent cover estimates: ‘objective’ is not always better. Mar. Ecol. Prog. Ser. 1993, 96, 93–100. [Google Scholar] [CrossRef]
- McKenzie, L.J.; Campbell, S.J.; Roder, C.A. Seagrass-Watch: Manual for Mapping & Monitoring Seagrass Resources by Community (Citizen) Volunteers; Queensland Fisheries Service, Department of Primary Industries: Cairns, Switzerland, 2001; p. 100. [Google Scholar]
- Wentworth, C.K. A Scale of Grade and Class Terms for Clastic Sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef] [PubMed]
- Imhof, H.K.; Schmid, J.; Niessner, R.; Ivleva, N.P.; Laforsch, C. A novel, highly efficient method for the separation and quantification of plastic particles in sediments of aquatic environments. Limnol. Oceanogr. Methods 2012, 10, 524–537. [Google Scholar] [CrossRef]
- Coppock, R.L.; Cole, M.; Lindeque, P.K.; Queirós, A.M.; Galloway, T.S. A small-scale, portable method for extracting microplastics from marine sediments. Environ. Poll. 2017, 230, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Baltic, C.C. Guide to Microplastic Identification; Marine and Environmental Research Institute: Blue Hill, ME, USA, 2017. [Google Scholar]
- Suaria, G.; Achtypi, A.; Perold, V.; Lee, J.R.; Pierucci, A.; Bornman, T.G.; Aliani, S.; Ryan, P.G. Microfibers in oceanic surface waters: A global characterization. Sci. Adv. 2020, 6, eaay8493. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Lawrence Erlbaum Associates: New Jersey, NJ, USA, 1988. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Sanchez-Vidal, A.; Canals, M.; de Haan, W.P.; Romero, J.; Veny, M. Seagrasses provide a novel ecosystem service by trapping marine plastics. Sci. Rep. 2021, 11, 254. [Google Scholar] [CrossRef]
- Hartline, N.L.; Bruce, N.J.; Karba, S.N.; Ruff, E.O.; Sonar, S.U.; Holden, P.A. Microfiber Masses Recovered from Conventional Machine Washing of New or Aged Garments. Environ. Sci. Technol. 2016, 50, 11532–11538. [Google Scholar] [CrossRef] [Green Version]
- Bessa, F.; Barría, P.; Neto, J.M.; Frias, J.P.G.L.; Otero, V.; Sobral, P.; Marques, J.C. Occurrence of microplastics in commercial fish from a natural estuarine environment. Mar. Poll. Bull. 2018, 128, 575–584. [Google Scholar] [CrossRef]
- de los Santos, C.B.; Krång, A.-S.; Infantes, E. Microplastic retention by marine vegetated canopies: Simulations with seagrass meadows in a hydraulic flume. Environ. Poll. 2021, 269, 116050. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, K.; Su, L.; Li, J.; Yang, D.; Tong, C.; Mu, J.; Shi, H. Microplastics and mesoplastics in fish from coastal and fresh waters of China. Environ. Poll. 2017, 221, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Lusher, A.L.; McHugh, M.; Thompson, R.C. Occurrence of microplastics in the gastrointestinal tract of pelagic and demersal fish from the English Channel. Mar. Poll. Bull. 2013, 67, 94–99. [Google Scholar] [CrossRef]
- Chakraborty, P.; Sarkar, A.; Vudamala, K.; Naik, R.; Nath, B.N. Organic matter—A key factor in controlling mercury distribution in estuarine sediment. Mar. Chem. 2015, 173, 302–309. [Google Scholar] [CrossRef]
- Schorer, M. Pollutant and organic matter content in sediment particle size fractions. Int. Assoc. Hydrol. Sci. 1997, 243, 59–68. [Google Scholar]
- Alomar, C.; Estarellas, F.; Deudero, S. Microplastics in the Mediterranean Sea: Deposition in coastal shallow sediments, spatial variation and preferential grain size. Mar. Environ. Res. 2016, 115, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hardesty, B.D.; Harari, J.; Isobe, A.; Lebreton, L.; Maximenko, N.; Potemra, J.; van Sebille, E.; Vethaak, A.D.; Wilcox, C. Using Numerical Model Simulations to Improve the Understanding of Micro-plastic Distribution and Pathways in the Marine Environment. Front. Mar. Sci. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Röhr, M.E.; Holmer, M.; Baum, J.K.; Björk, M.; Chin, D.; Chalifour, L.; Cimon, S.; Cusson, M.; Dahl, M.; Deyanova, D.; et al. Blue Carbon Storage Capacity of Temperate Eelgrass (Zostera marina) Meadows. Glob. Biogeochem. Cycles 2018, 32, 1457–1475. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.M.; Unsworth, R.K.F. Protecting the hand that feeds us: Seagrass (Zostera marina) serves as commercial juvenile fish habitat. Mar. Poll. Bull. 2014, 83, 425–429. [Google Scholar] [CrossRef]
- Lilley, R.; Unsworth, R.K.F. Atlantic Cod (Gadus morhua) benefits from the availability of seagrass (Zostera marina) nursery habitat. Glob. Ecol. Conserv. 2014, 2, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Furness, E.; Unsworth, R.K.F. Demersal Fish Assemblages in NE Atlantic Seagrass and Kelp. Diversity 2020, 12, 366. [Google Scholar] [CrossRef]
- Nordlund, L.; Unsworth, R.; Gullström, M.; Cullen-Unsworth, L. Global significance of seagrass fishery activity. Fish Fish. 2018, 19, 399–412. [Google Scholar] [CrossRef]
- Green, A.E.; Unsworth, R.K.F.; Chadwick, M.A.; Jones, P.J. Historical analysis exposes catastrophic seagrass loss for the United Kingdom. Front. Plant Sci. 2021. In Press. [Google Scholar]
- Deudero, S.; Alomar, C. Mediterranean marine biodiversity under threat: Reviewing influence of marine litter on species. Mar. Poll. Bull. 2015, 98, 58–68. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Eight seagrass sampling locations around the United Kingdom. Sites at Cornwall and North Wales were Zostera marina dominated. The remaining six sites were Zostera noltii-dominated. Sites are labelled as coastal (c) or estuarine (e).
Figure 1.
Eight seagrass sampling locations around the United Kingdom. Sites at Cornwall and North Wales were Zostera marina dominated. The remaining six sites were Zostera noltii-dominated. Sites are labelled as coastal (c) or estuarine (e).

Figure 2.
Mean (±SD) canopy heights (cm) and seagrass cover (%) for each of the eight sampled sites. All sites are dominated by Zostera noltii except for North Wales and Cornwall that were Zostera marina-dominated. Pairwise comparisons from PERMANOVA are shown as letter groupings.
Figure 2.
Mean (±SD) canopy heights (cm) and seagrass cover (%) for each of the eight sampled sites. All sites are dominated by Zostera noltii except for North Wales and Cornwall that were Zostera marina-dominated. Pairwise comparisons from PERMANOVA are shown as letter groupings.

Figure 3.
Percentage breakdown of the composition of the marine microplastics found within sediments of seagrass (SG) and unvegetated habitats (UV) at eight sites around the UK.
Figure 3.
Percentage breakdown of the composition of the marine microplastics found within sediments of seagrass (SG) and unvegetated habitats (UV) at eight sites around the UK.

Figure 4.
Examples of all identified microplastic types within sediments of both seagrass and non-vegetative areas. Images A and D show fragments. B and E shows fibres. C shows films and F shows a fibre bundle.
Figure 4.
Examples of all identified microplastic types within sediments of both seagrass and non-vegetative areas. Images A and D show fragments. B and E shows fibres. C shows films and F shows a fibre bundle.

Figure 5.
Boxplots of data of marine microplastics (marine plastic per kilogram dryweight of sediment) recorded within sediments of seagrass and unvegetated habitats at eight intertidal sites around the UK. Cross hatched bars indicate sites where seagrass was of the species Zostera marina whereas non hatched bars are of the seagrass Zostera noltii.
Figure 5.
Boxplots of data of marine microplastics (marine plastic per kilogram dryweight of sediment) recorded within sediments of seagrass and unvegetated habitats at eight intertidal sites around the UK. Cross hatched bars indicate sites where seagrass was of the species Zostera marina whereas non hatched bars are of the seagrass Zostera noltii.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary table of marine microplastics data from all known academic research studies known to have quantified microplastics in seagrass sediments.
Table 1.
Summary table of marine microplastics data from all known academic research studies known to have quantified microplastics in seagrass sediments.
| Seagrass | Unvegetated | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | Media | Location | No. of Locations | Location Type | Dominant Seagrass Species | Sub- (S) or Inter-tidal (I) | n | MP Abundance (MP kg−1 DW) | % of Samples Contain MP | Dominant MP Form | MP (MP kg−1 DW) | % of Samples Contain MP | Dominant MP Form | Effect Size (Cohens d) |
| Tahir et al. 2019 | ZnBr | Indonesia | 3 | Remote island | Halophila, Cymodocea, Enhalus, Thalassia, Syringodium, Halodule | S | 81 | 131 ± 100 | 27 | Fibres Filaments | NA | NA | NA | 1.31 |
| Huang et al. 2020 | ZnCl2 | China | 2 | Coastal | Enhalus acodoides | I | 12 (3 each) | 196.7 ± 16.1 | 100 | Fibres Filaments | 93.3 ± 15.3 | 100 | Fibres Filaments | 6.5 |
| 780.2 ± 147.0 | 267.1 ± 60.5 | 4.9 | ||||||||||||
| Jones et al. 2020 | NaCl | Scotland | 1 | Island | Zostera marina | S | 25 | 300 ± 30 (20) | 100 | Fibres | 110 ± 20 (5) | 100 | Fibres | 7.6 |
| Cozzolino et al. 2020 | NaCl | Portugal | 1 | Lagoon | Z. marina | I | 40 (10 each) | 18.2 ± 15.5 (0–11) | 100 | Fibres | 29.8 ± 14.9 (3–15) | 100 | Fibres | −0.76 |
| Cymodocea nodosa, Z. marina | S | 35.2 ± 26.7 (1–21) | 20.4 ± 14.5 (1–10) | 0.71 | ||||||||||
| Plee and Pomory 2020 | NaCl | USA | 1 | Lagoon | NA | S | 6 | 24 ± 6 | 100 | Fibres Fragments | 15 ± 3 | 100 | Fibres | 2 |
| Our study | ZnCl | England and Wales | 8 | Coastal and Estuarine | Z.marina Z. noltii | I | 80 (40 each) | 215 ± 163 | 98 | Fibres Filaments | 221 ± 236 | 98 | Fibres Filaments | −0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Unsworth, R.K.F.; Higgs, A.; Walter, B.; Cullen-Unsworth, L.C.; Inman, I.; Jones, B.L. Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics? Oceans 2021, 2, 162-178. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010010
AMA Style
Unsworth RKF, Higgs A, Walter B, Cullen-Unsworth LC, Inman I, Jones BL. Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics? Oceans. 2021; 2(1):162-178. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010010
Chicago/Turabian StyleUnsworth, Richard K. F., Alex Higgs, Bettina Walter, Leanne C. Cullen-Unsworth, Isabella Inman, and Benjamin L. Jones. 2021. "Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics?" Oceans 2, no. 1: 162-178. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010010