
Ocean Circulation Drives the Variability of the Carbon System in the Eastern Tropical Atlantic

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
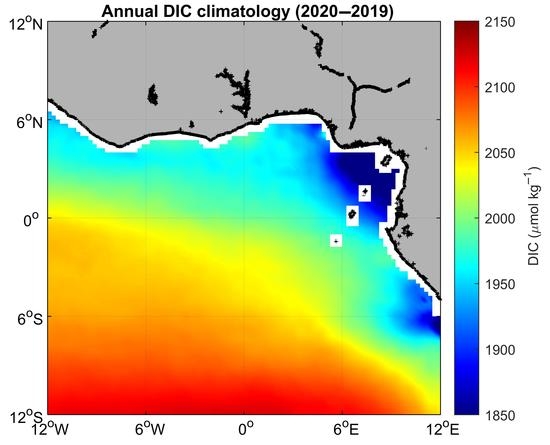
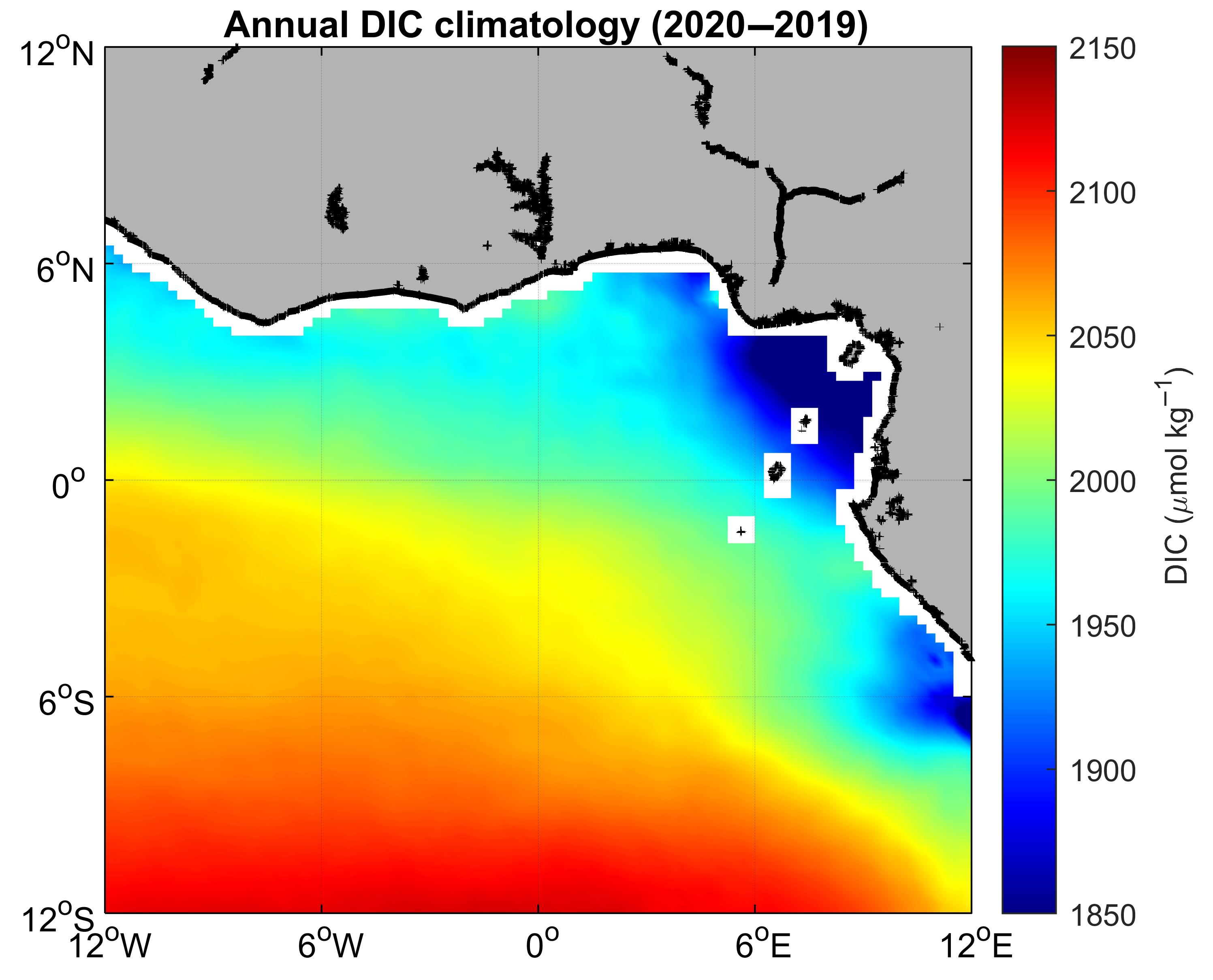
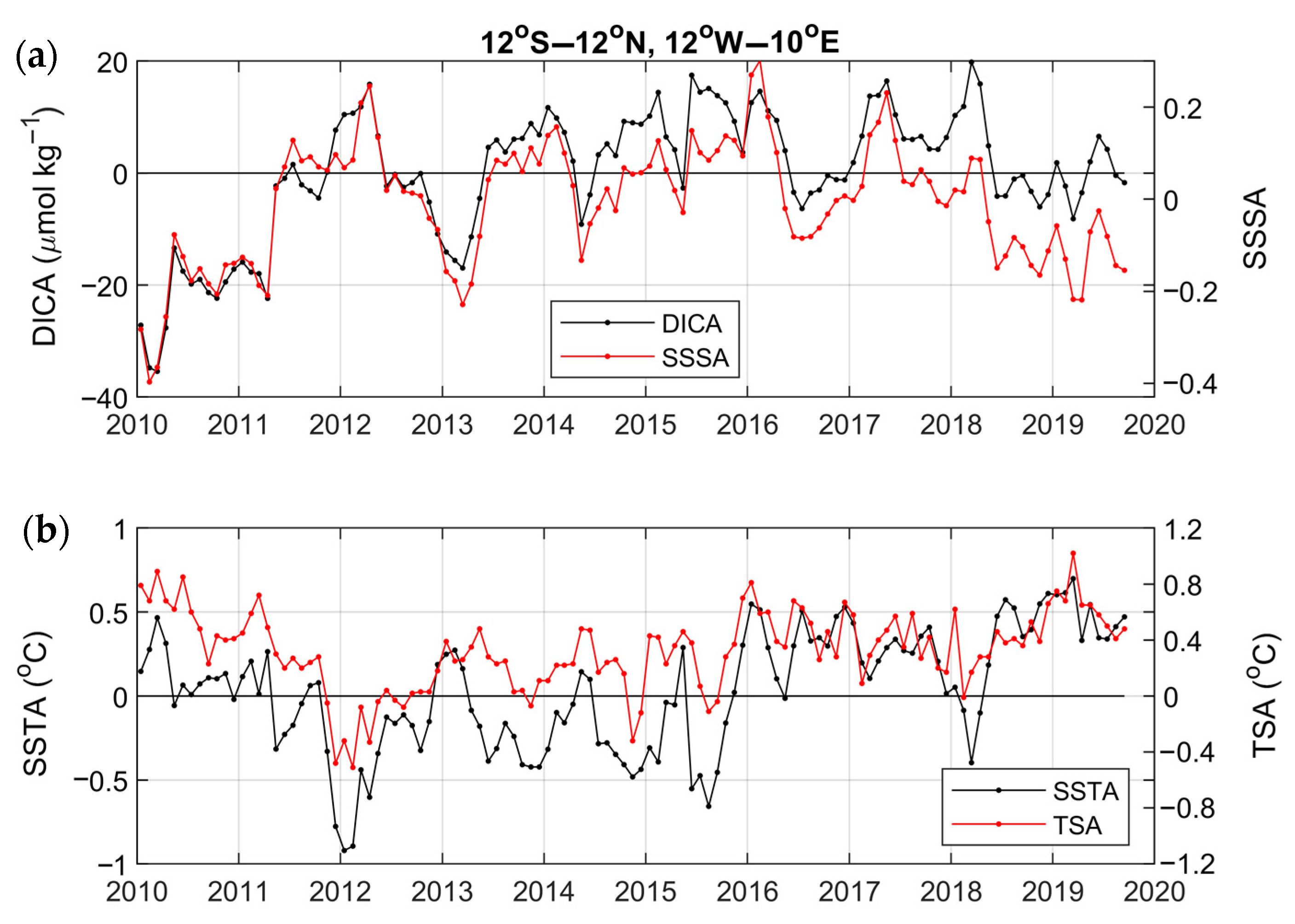
3. Results
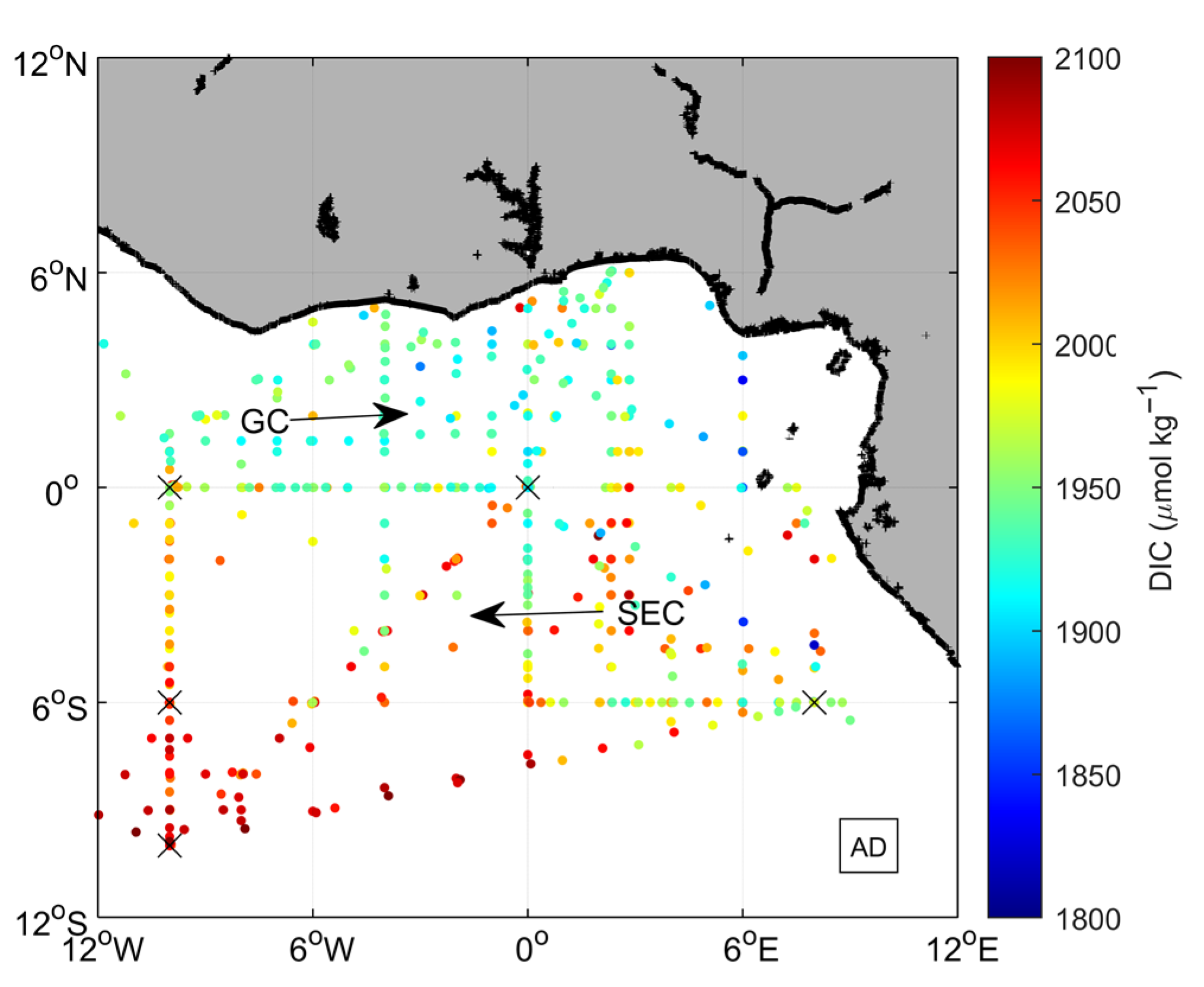
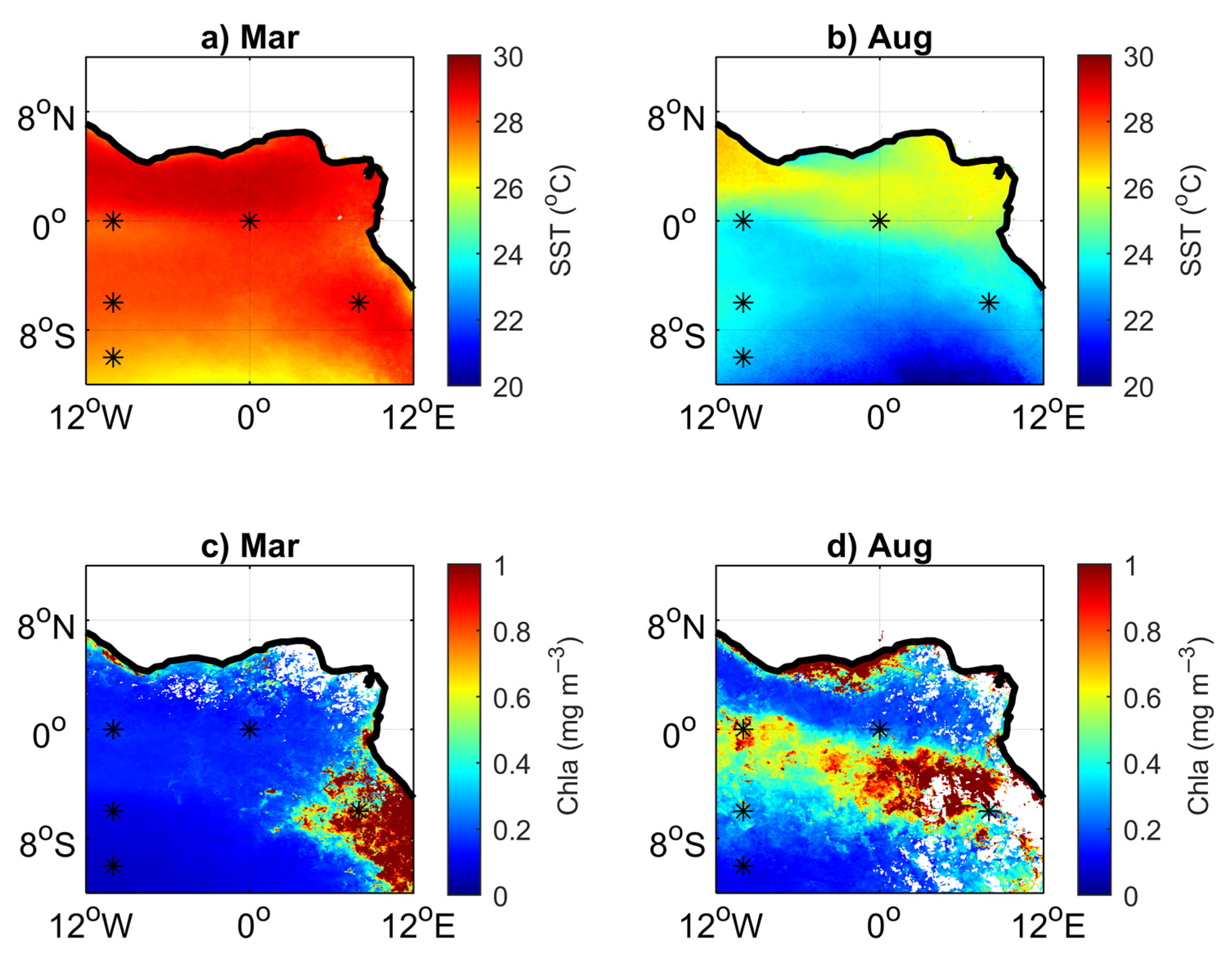
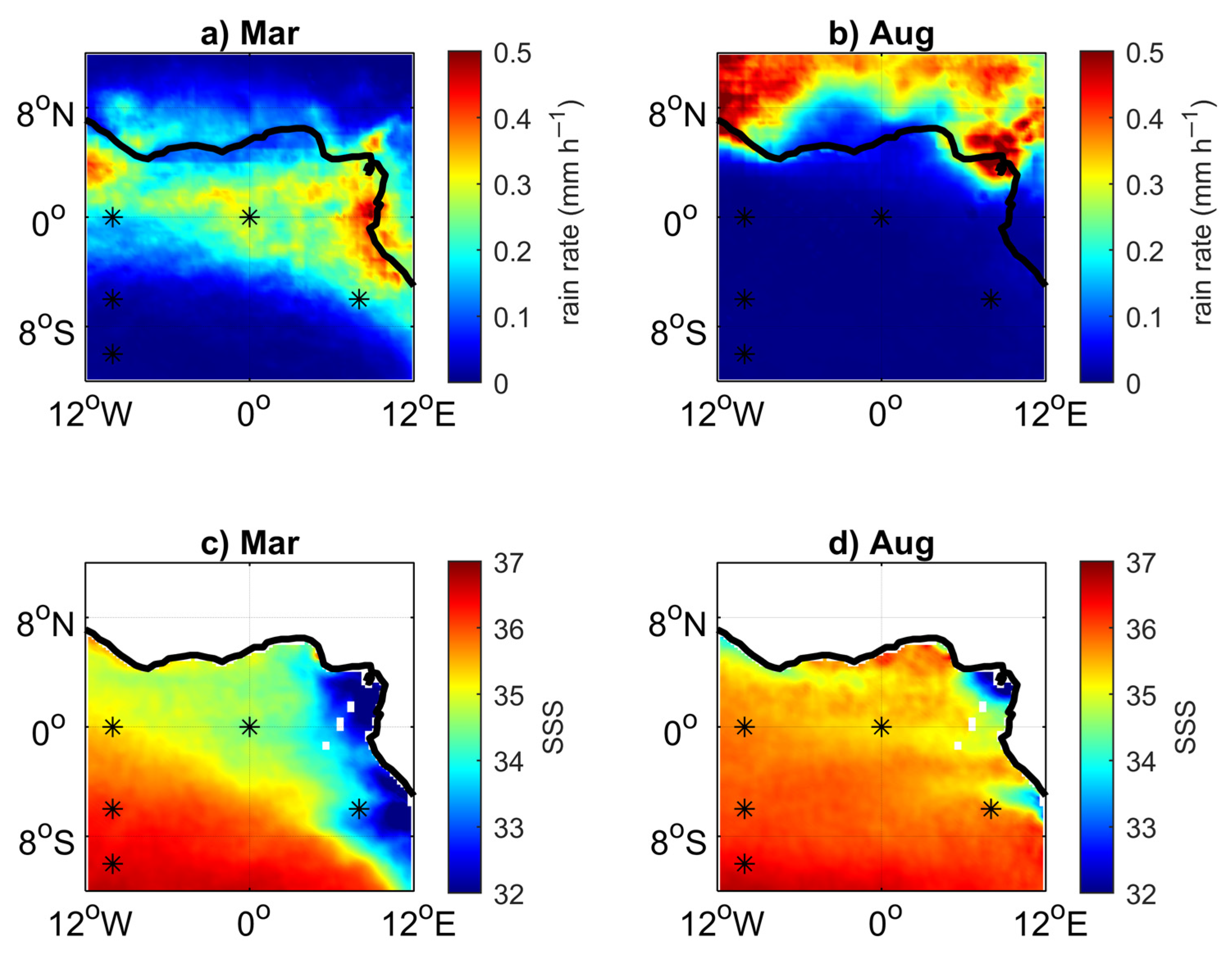
3.1. Environmental Setting
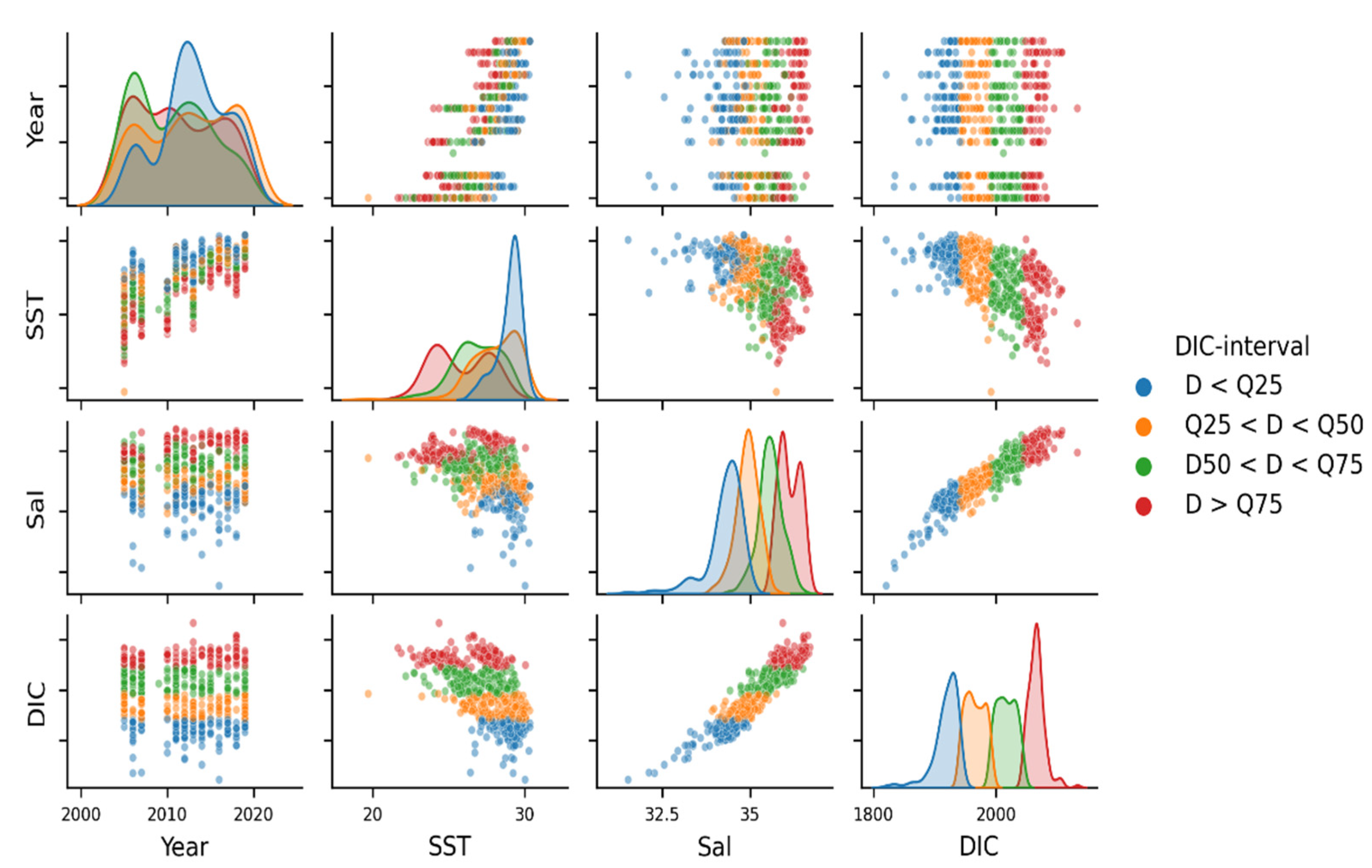
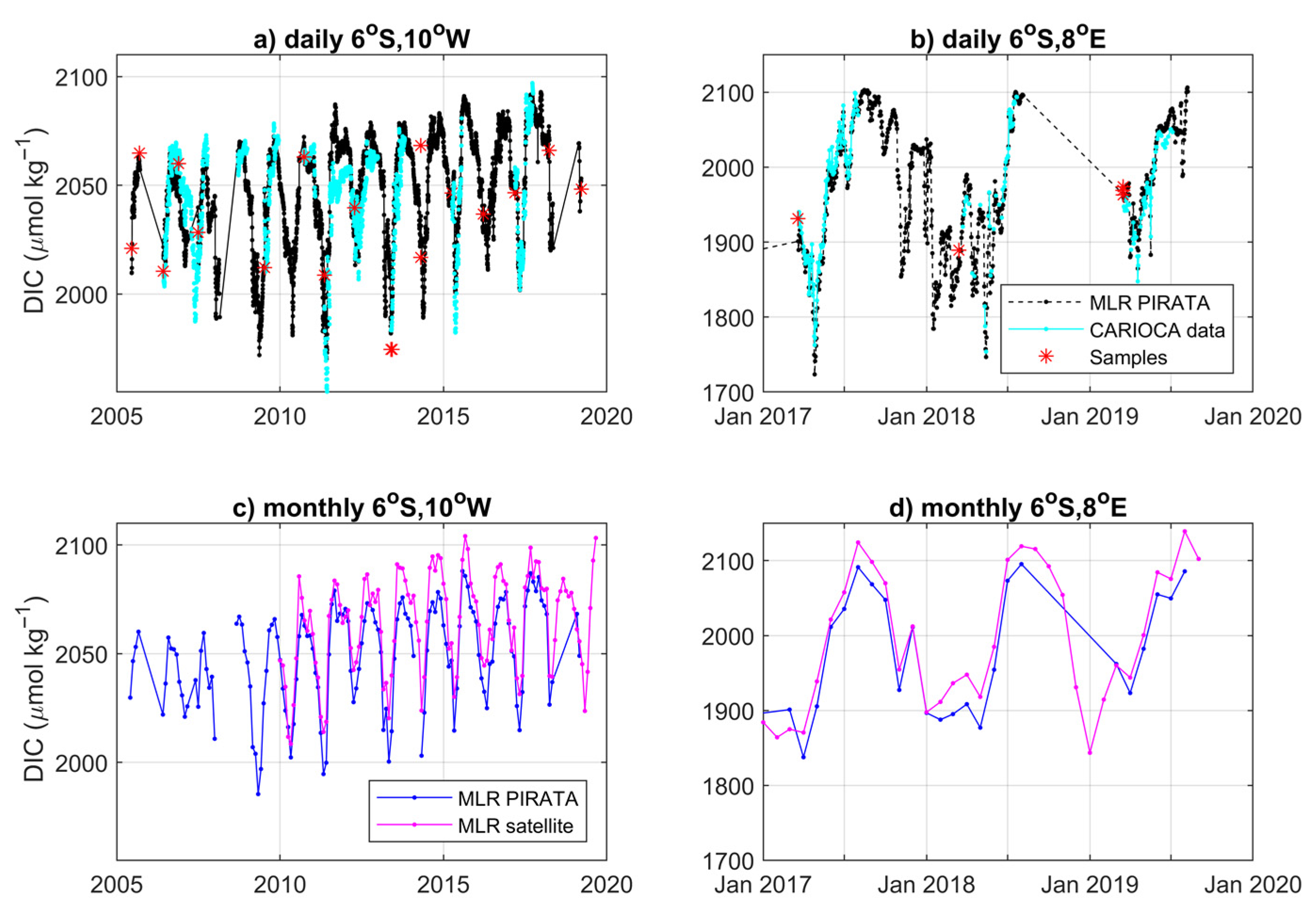
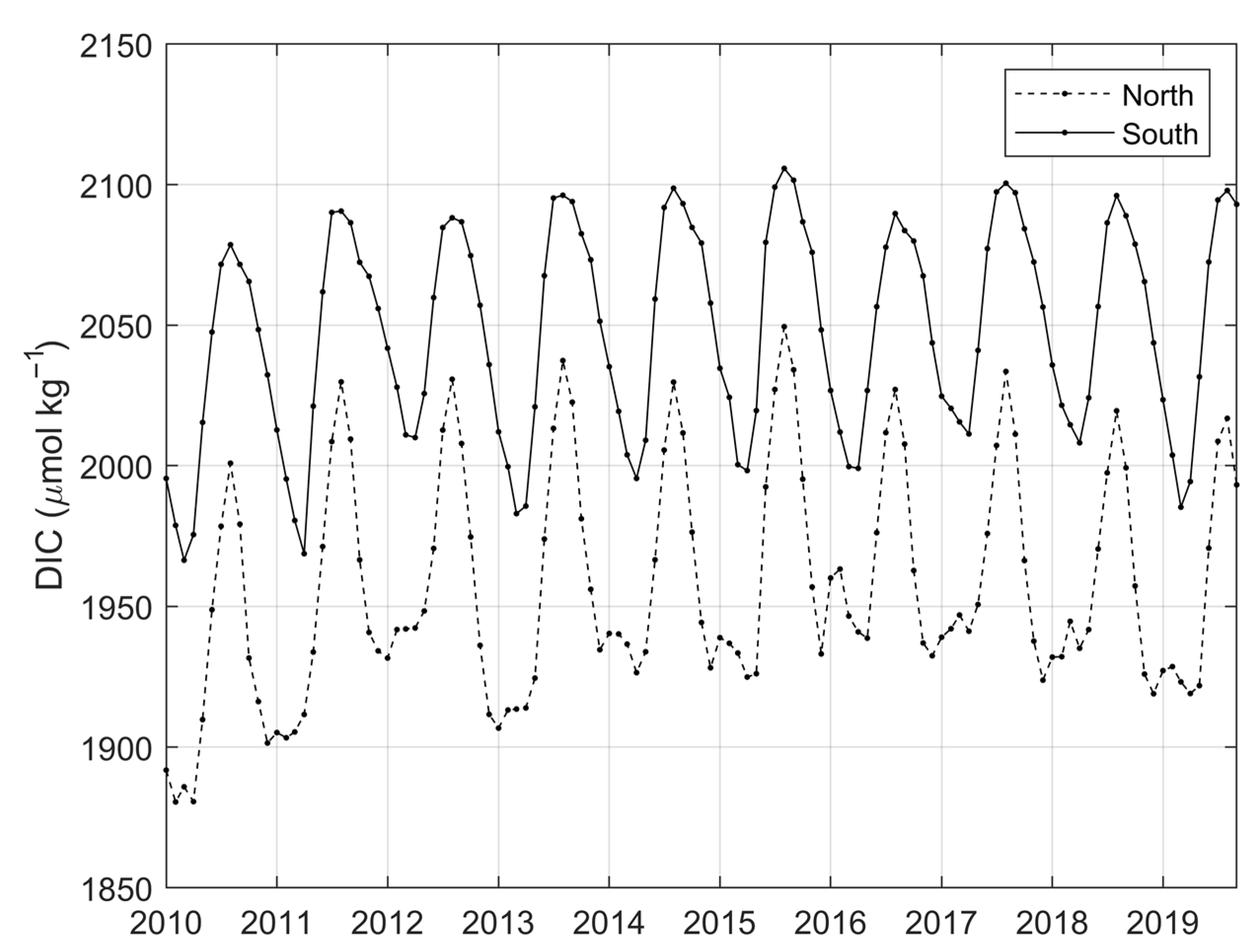
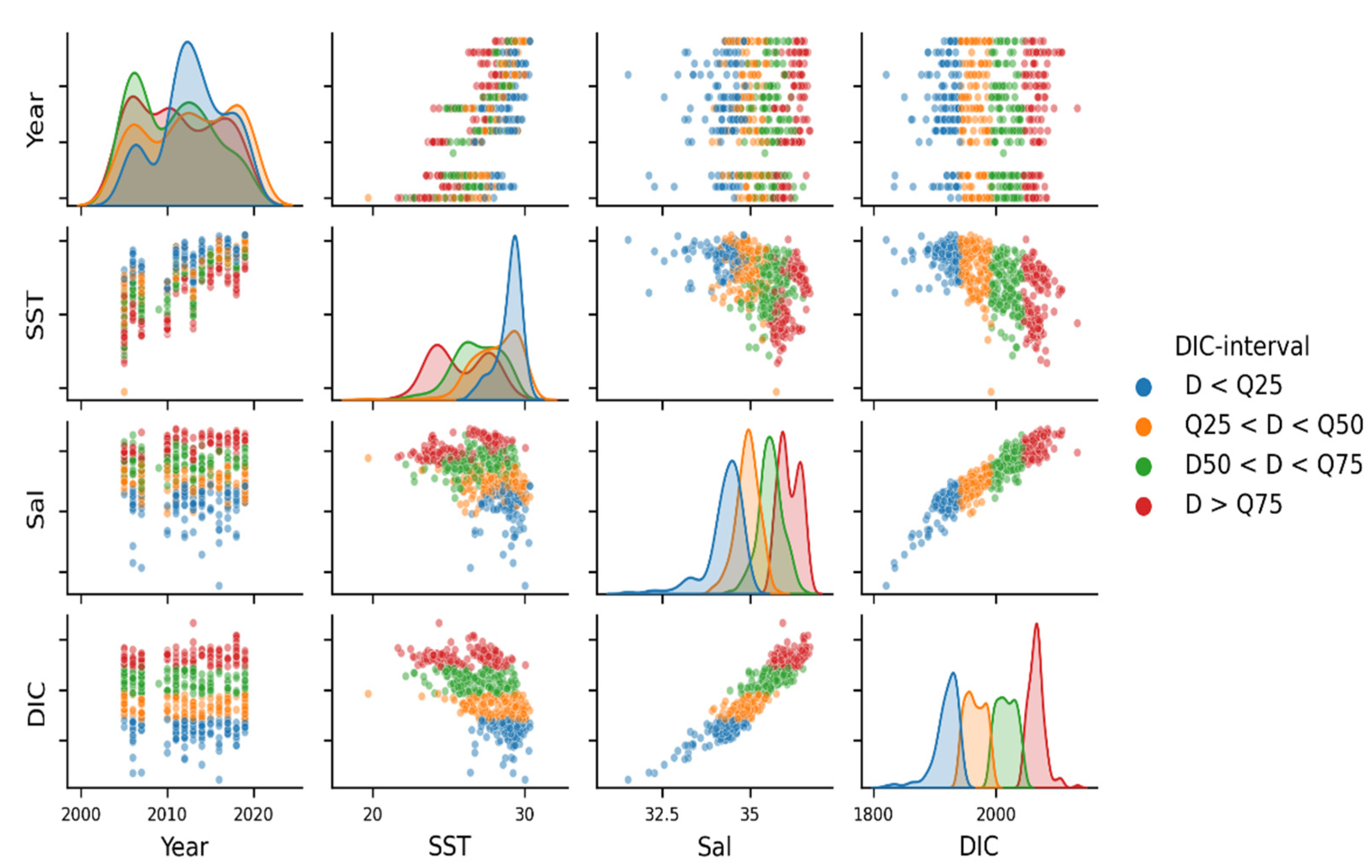
3.2. Variability and Drivers of the Carbon System
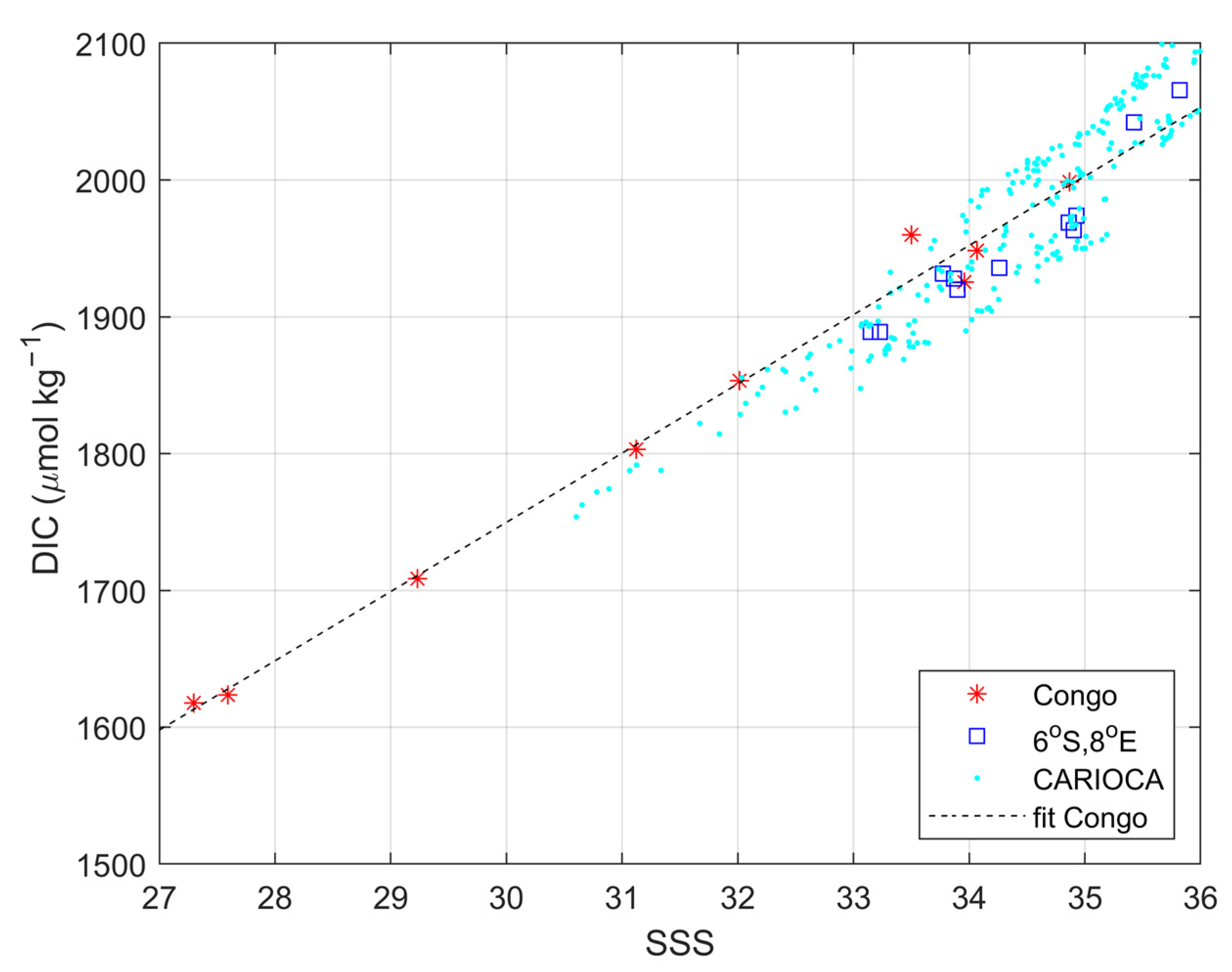
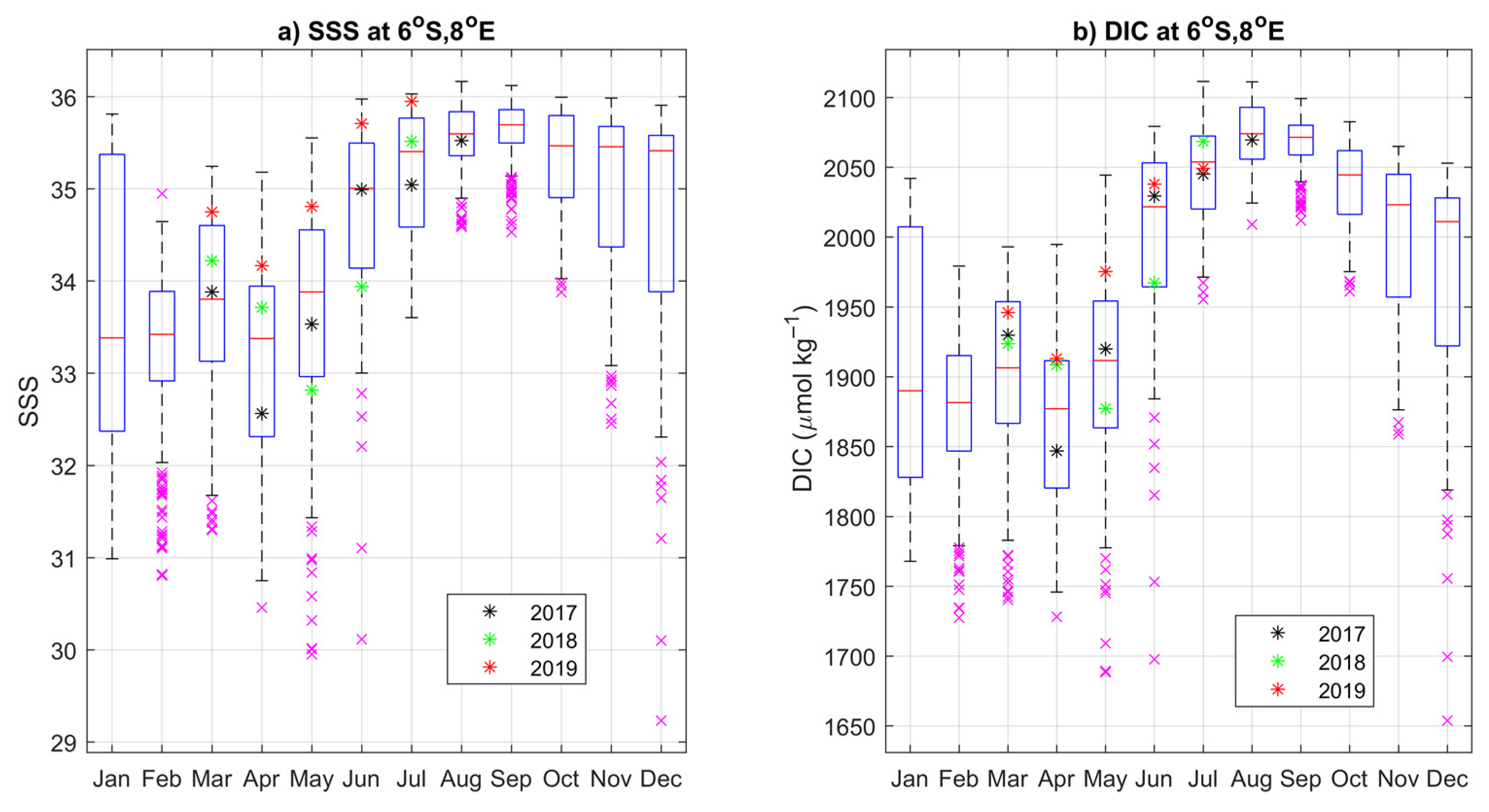
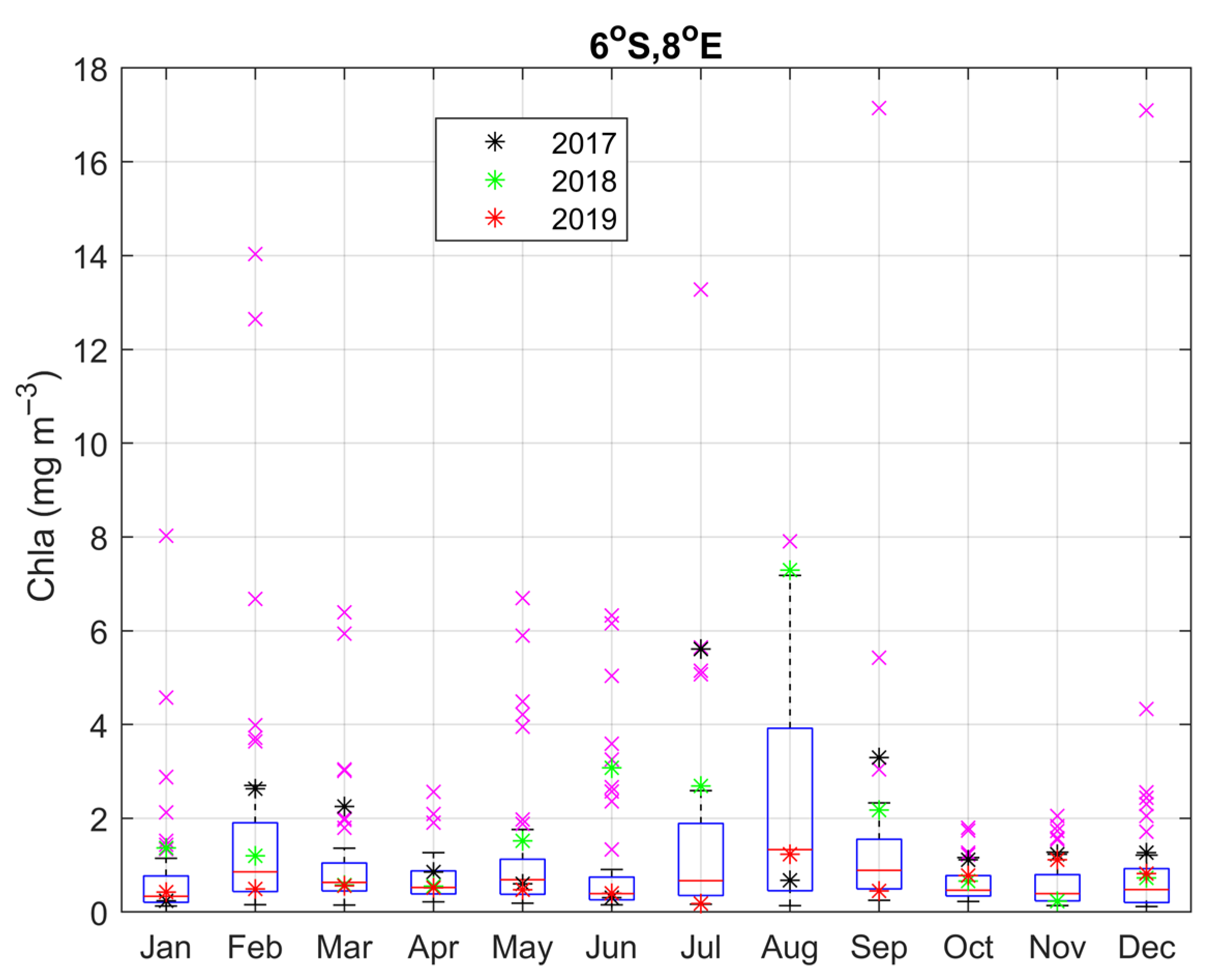
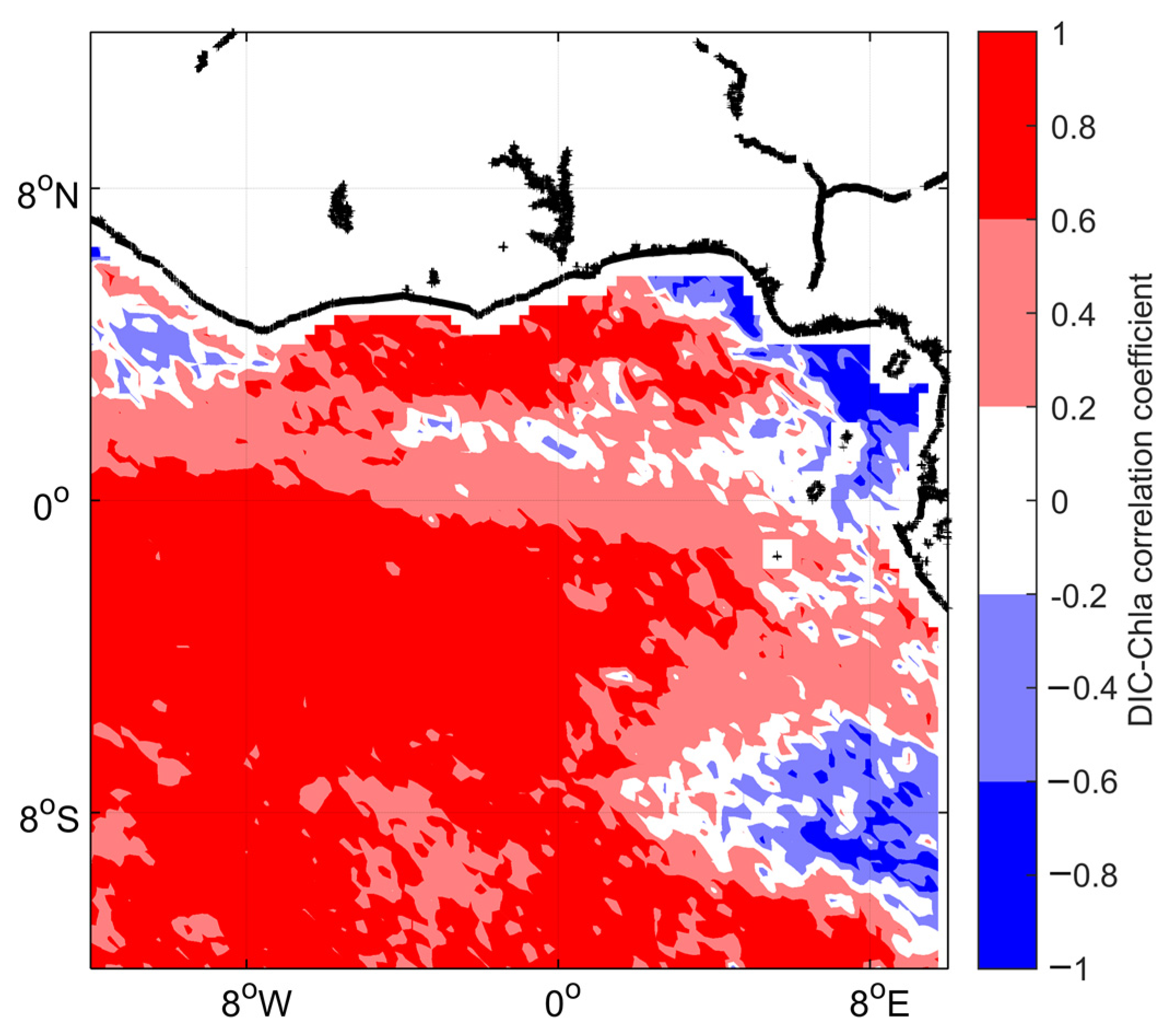
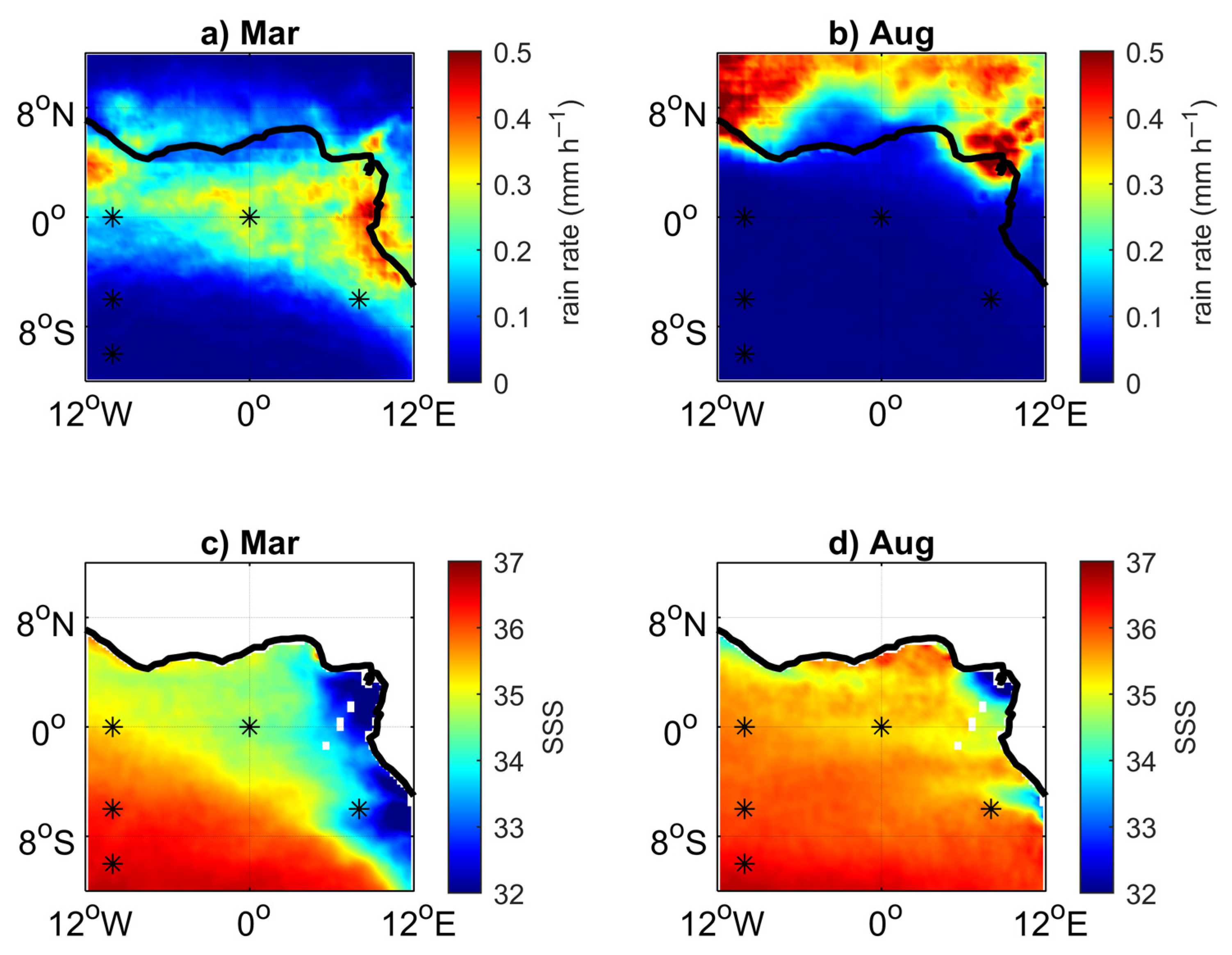
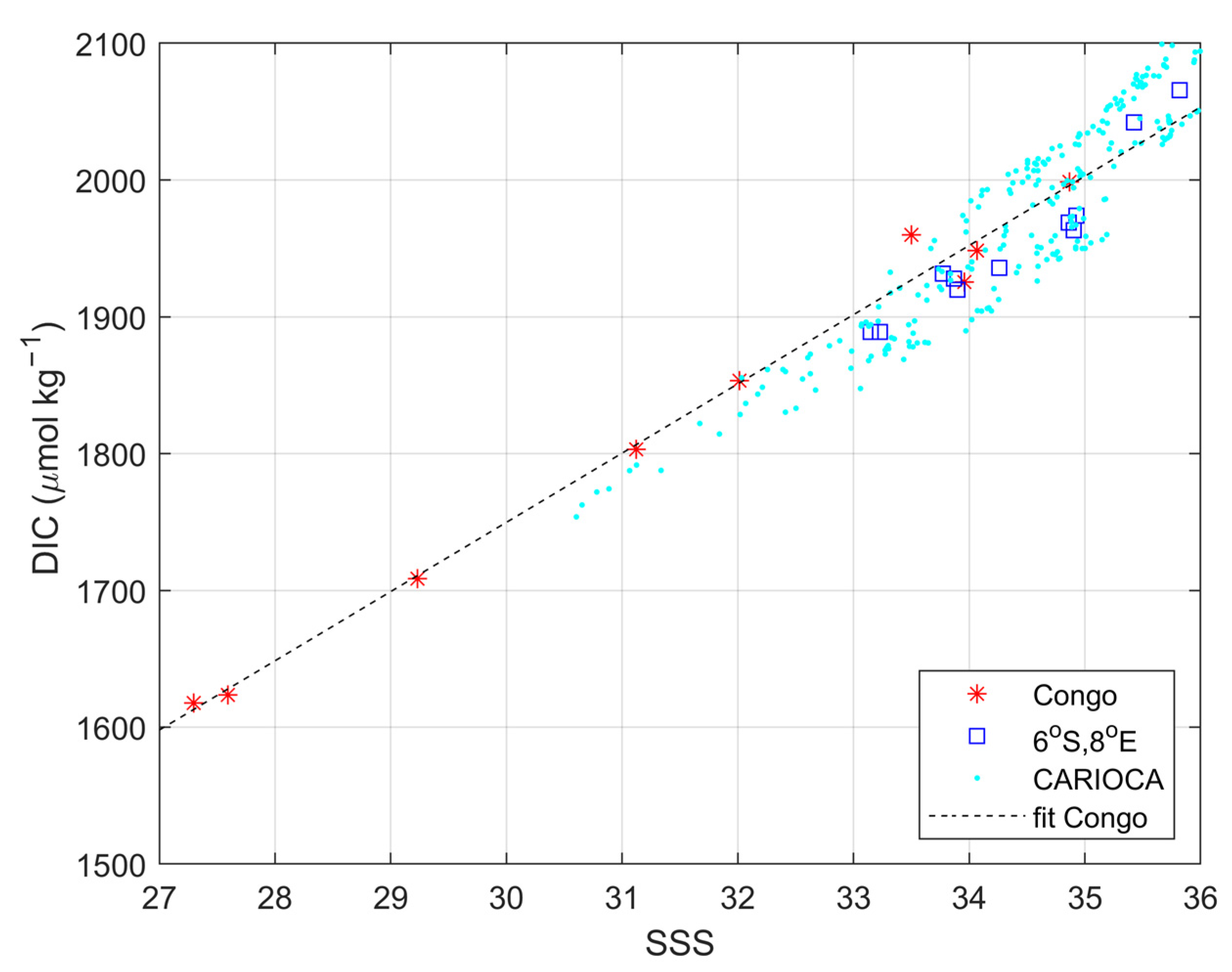
3.3. Impact of the Congo Plume
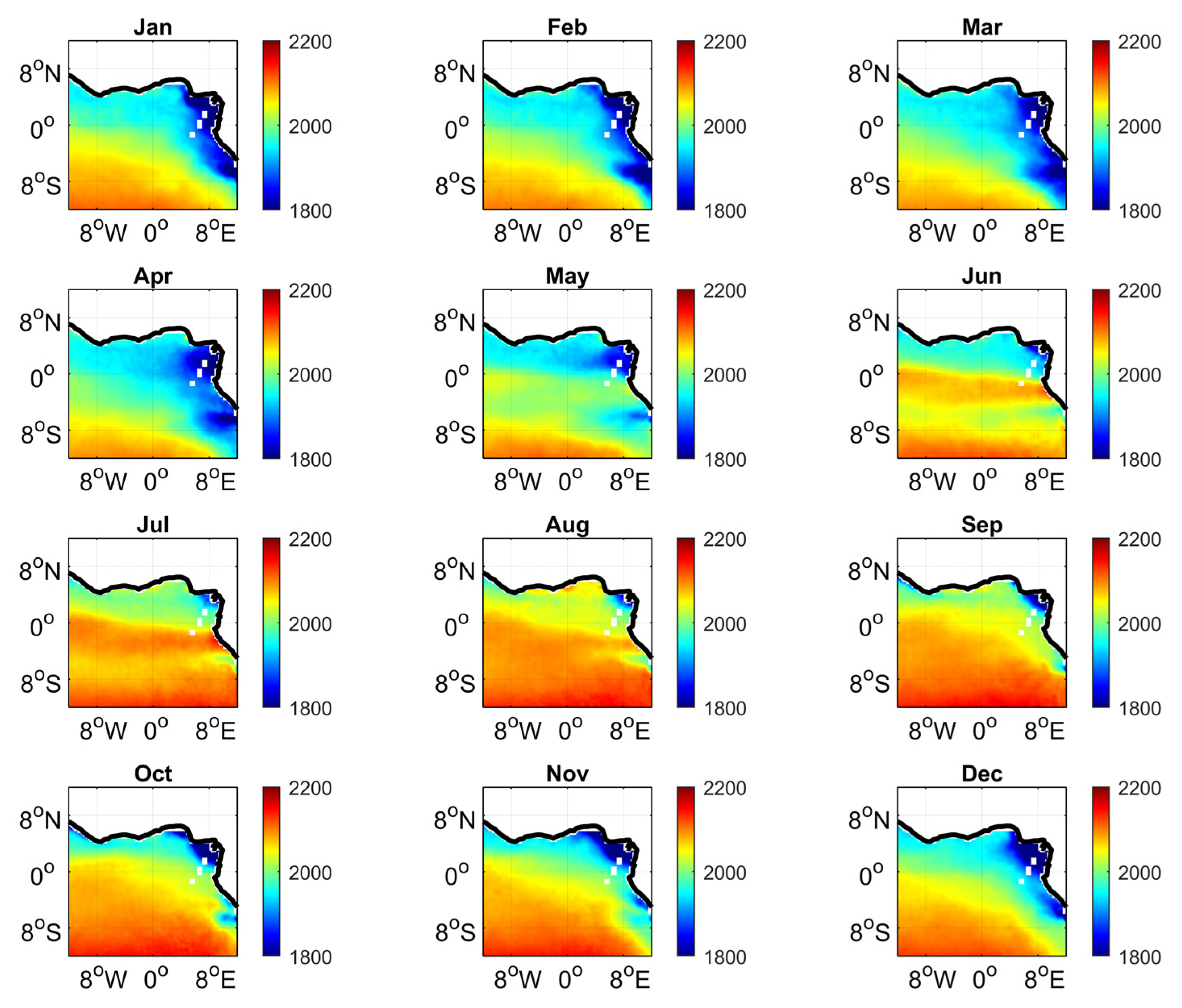
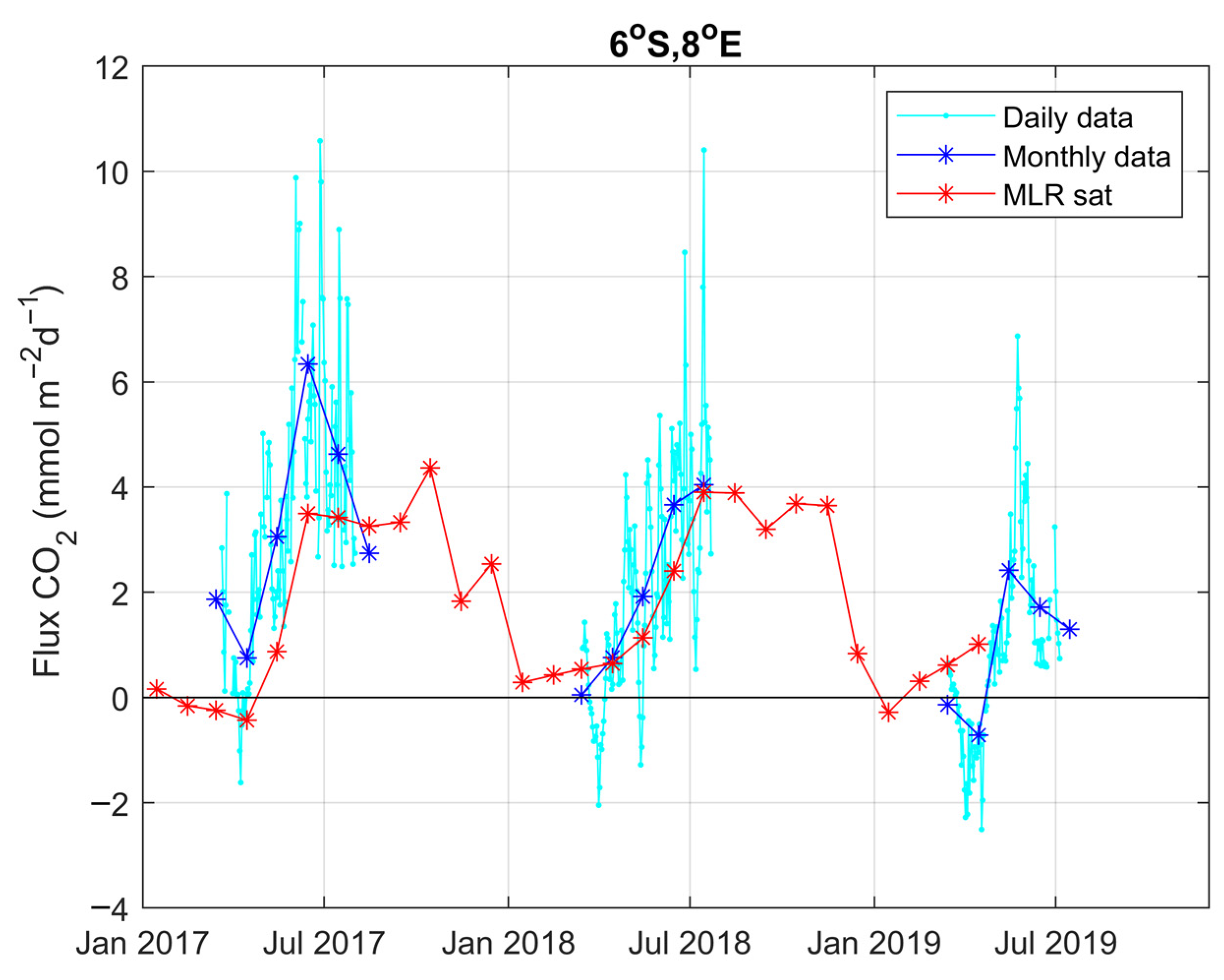
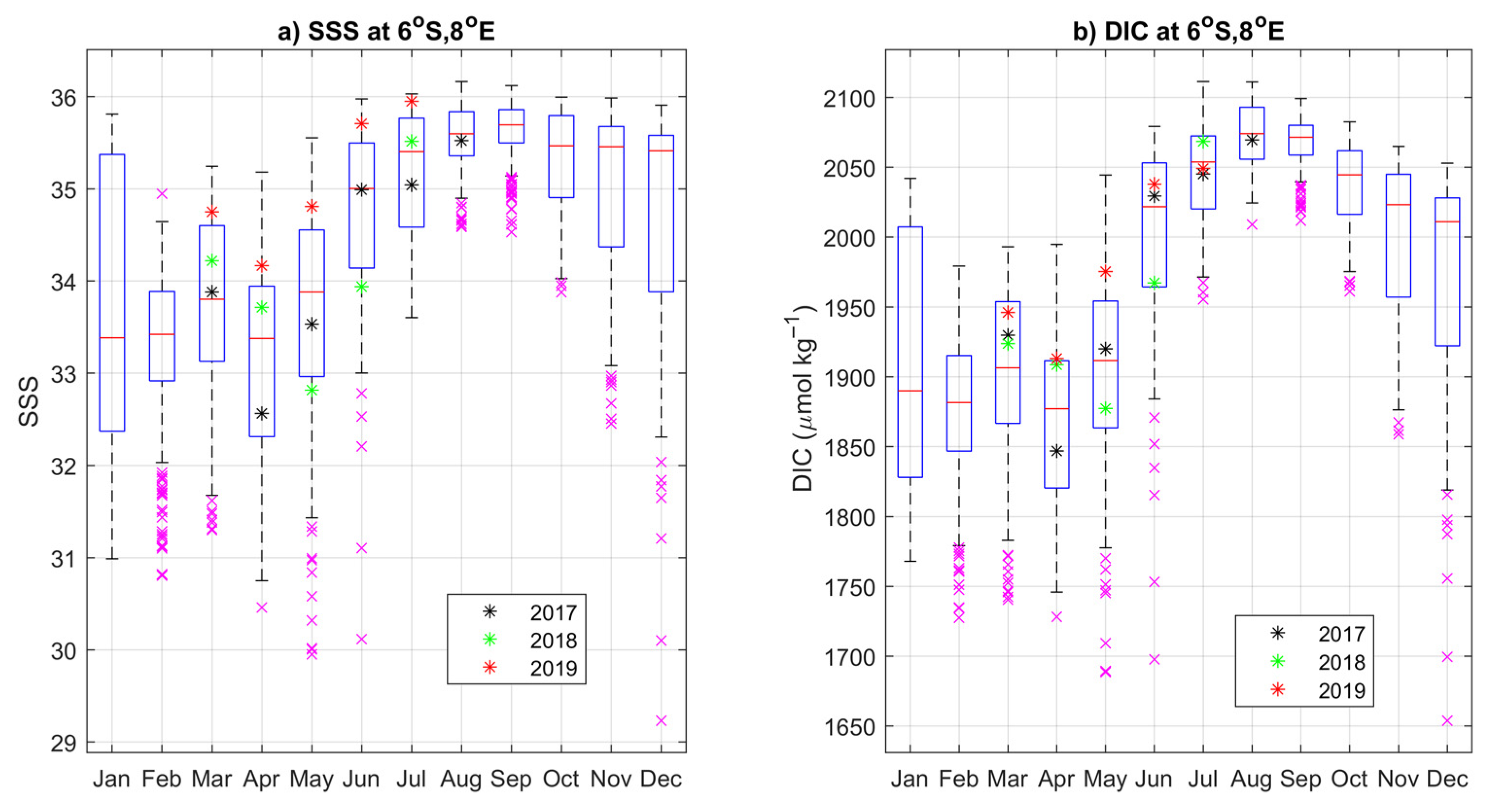
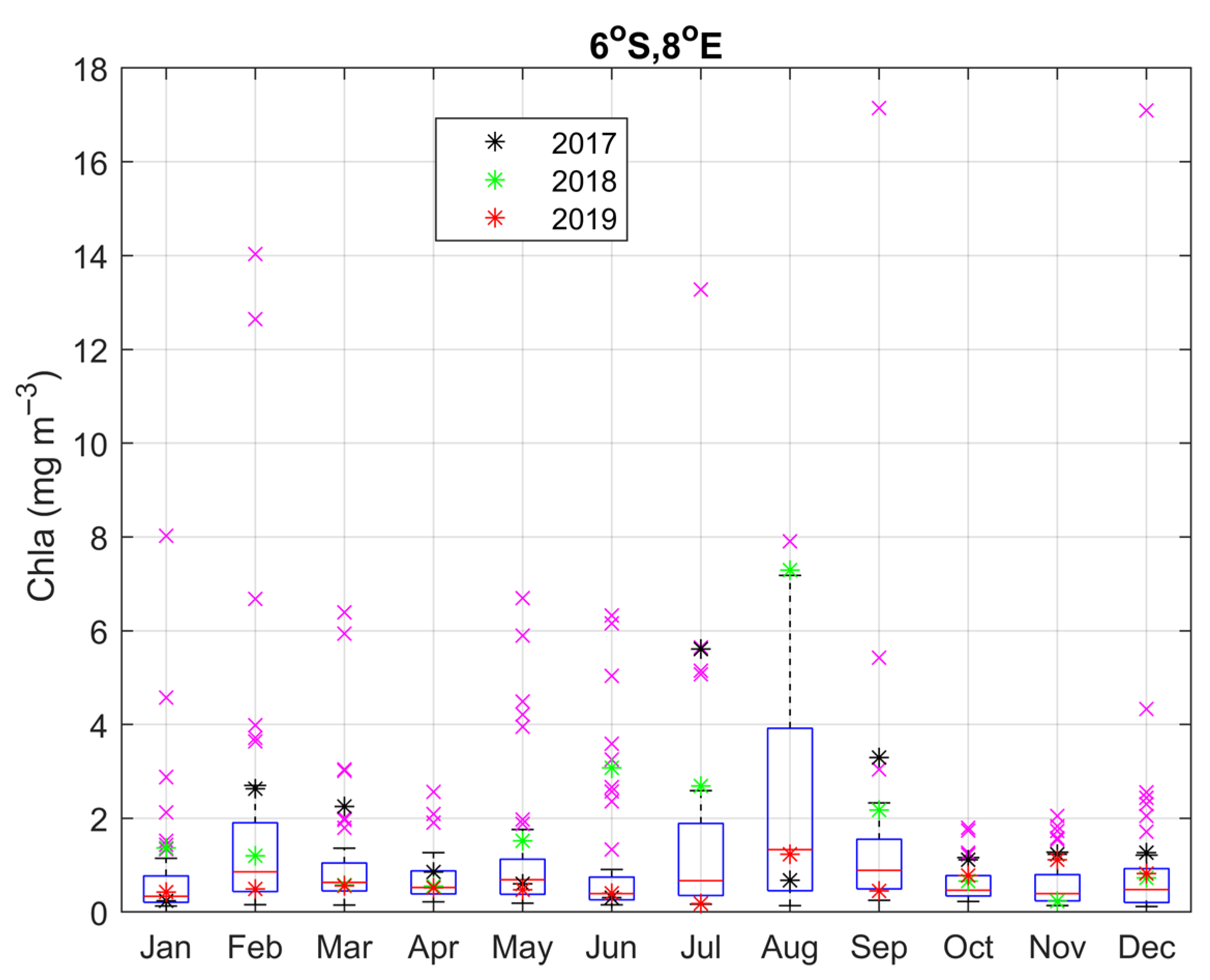
3.4. Year-to-Year Variability of the Carbon Parameters
4. Discussion
4.1. Main Features of the Carbon Parameters
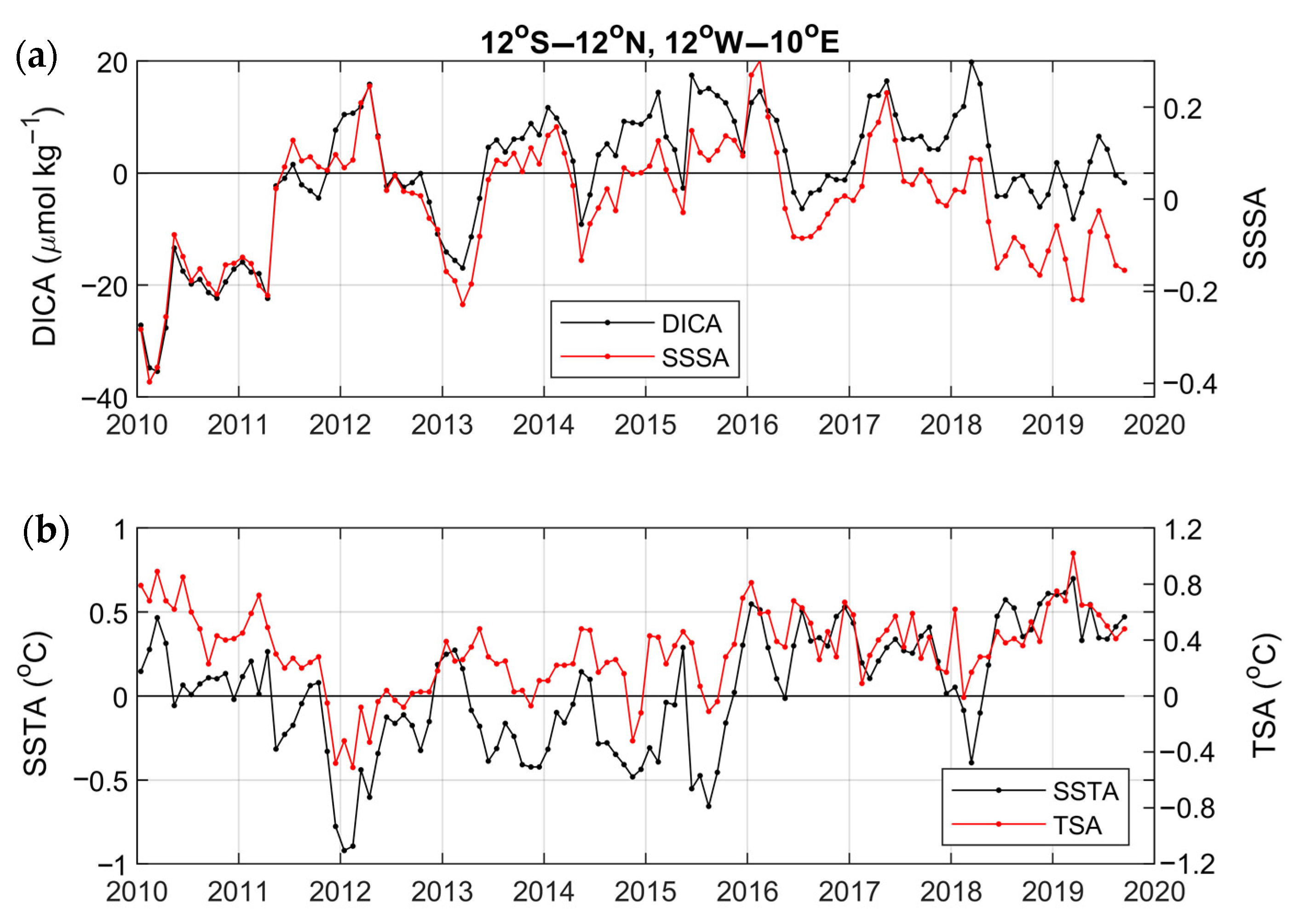
4.2. Year-to-Year Variability
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Regression Methods and Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meta-Parameters | Specifications |
|---|---|
| Input variables | Year, SST, SSS, SinDoY, CosDoY |
| Output variables | DIC |
| Train/Val | DIC 4 quantiles (1/6 for Val from each quant.) |
| Normalization | Center-reduction (both Input and Output) |
| Architecture | Layers size: Input: 5, Hidden: 15, Output: 1 |
| Condition | N | NTrain | NVal | |
|---|---|---|---|---|
| DIC < Q25 | DIC < 1941.1 | 159 | 132 | 27 |
| Q25 < DIC < Q50 | 1941.1 ≤ DIC < 1992.2 | 159 | 132 | 27 |
| Q50 < DIC < Q75 | 1992.2 ≤ DIC < 2044.6 | 158 | 132 | 26 |
| DIC > Q75 | DIC ≥ 2044.6 | 161 | 135 | 27 |
| Mooring or Cruise | Regression Method | DIC < Q25 | Q25 < DIC < Q50 | Q50 < DIC < Q75 | DIC > Q75 |
|---|---|---|---|---|---|
| RMSE [µmol kg−1] | RMSE [µmol kg−1] | RMSE [µmol kg−1] | RMSE [µmol kg−1] | ||
| 6° S, 10° W | MLR | NaN | 13.8 | 7.2 | 7.8 |
| NN | NaN | 13.4 | 7.3 | 7.0 | |
| 6° S, 8° E | MLR | 18.0 | 15.0 | 12.7 | 9.4 |
| NN | 14.0 | 15.0 | 11.5 | 8.9 | |
| EGEE 3 | MLR | 7.1 | 8.3 | 12.5 | 18.1 |
| NN | 4.8 | 7.5 | 9.9 | 14.1 | |
| PIRATA-FR-29 | MLR | 11.2 | 8.1 | 4.8 | 6.9 |
| NN | 8.1 | 11.1 | 5.3 | 9.2 |
| Mooring or Cruise | Regression Method | RMSE (mol kg−1) | r | N | Time Period |
|---|---|---|---|---|---|
| 6° S, 10° W | MLR | 7.7 | 0.96 | 6611 | 2006–2017 |
| DT | 14.1 | 0.87 | |||
| RF | 9.8 | 0.94 | |||
| NN | 7.3 | 0.97 | |||
| 6° S, 8° E | MLR | 14.8 | 0.98 | 239 | 2017–2019 |
| DT | 25 | 0.95 | |||
| RF | 24 | 0.95 | |||
| NN | 12.8 | 0.99 | |||
| EGEE 3 | MLR | 11.8 | 0.97 | 6895 | 2006 |
| DT | 14 | 0.96 | |||
| RF | 10 | 0.98 | |||
| NN | 9.3 | 0.98 | |||
| PIRATA FR-29 | MLR | 8.1 | 0.99 | 4462 | 2019 |
| DT | 11 | 0.98 | |||
| RF | 9 | 0.98 | |||
| NN | 9.4 | 0.99 |
References
- Le Quéré, C.; Orr, J.C.; Monfray, P.; Aumont, O.; Madec, G. Interannual variability of the oceanic sink of CO2 from 1979 through 1997. Glob. Biogeochem. Cycles 2000, 14, 1247–1266. [Google Scholar] [CrossRef]
- Wang, X.; Murtugudde, R.; Hackert, E.; Wang, J.; Beauchamp, J. Seasonal to decadal variations of sea surface pCO2 and sea-air CO2 flux in the equatorial oceans over 1984–2013: A basin-scale comparison of the Pacific and Atlantic Oceans. Glob. Biogeochem. Cycles 2015, 29, 597–609. [Google Scholar] [CrossRef]
- Cooley, S.R.; Coles, V.J.; Subramaniam, A.; Yager, P.L. Seasonal variations in the Amazon plume-related atmospheric carbon sink. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef] [Green Version]
- Cooley, S.R.; Yager, P.L. Physical and biological contributions to the western tropical North Atlantic Ocean carbon sink formed by the Amazon River plume. J. Geophys. Res. 2006, 111. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, A.; Yager, P.L.; Carpenter, E.J.; Mahaffey, C.; Björkman, K.; Cooley, S.; Kustka, A.B.; Montoya, J.P.; Sañudo-Wilhelmy, S.A.; Shipe, R.; et al. Amazon River enhances diazotrophy and carbon sequestration in the tropical North Atlantic Ocean. Proc. Natl. Acad. Sci. USA 2008, 105, 10460–10465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ternon, J.F.; Oudot, C.; Dessier, A.; Diverrès, D. A seasonal tropical sink for atmospheric CO2 in the Atlantic Ocean: The role of the Amazon River discharge. Mar. Chem. 2000, 68, 183–201. [Google Scholar] [CrossRef]
- Körtzinger, A. A significant sink of CO2 in the tropical Atlantic Ocean associated with the Amazon River plume. Geophys. Res. Lett. 2003, 30, 2287. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, N.; Flores, M.M.; Gaspar, F.L.; Rocha, C.; Jiang, S.; Araujo, M.; Ibánhez, J.S.P. Net heterotrophy in the Amazon continental shelf changes rapidly to a sink of CO2 in the outer Amazon plume. Front. Mar. Sci. 2017, 4, 278. [Google Scholar] [CrossRef] [Green Version]
- Ibánhez, J.S.P.; Araujo, M.; Lefèvre, N. The overlooked tropical oceanic CO2 sink. Geophys. Res. Lett. 2016, 43, 3804–3812. [Google Scholar] [CrossRef] [Green Version]
- Ibánhez, J.S.P.; Diverrès, D.; Araujo, M.; Lefèvre, N. Seasonal and interannual variability of sea-air CO2 fluxes in the tropical Atlantic affected by the Amazon River plume. Glob. Biogeochem. Cycles 2015, 29, 1640–1655. [Google Scholar] [CrossRef]
- Chao, Y.; Farrara, J.D.; Schumann, G.; Andreadis, K.M.; Moller, D. Sea surface salinity variability in response to the Congo river discharge. Cont. Shelf Res. 2015, 99, 35–45. [Google Scholar] [CrossRef]
- Signorini, S.R.; Murtugudde, R.G.; McClain, C.R.; Christian, J.R.; Picaut, J.; Busalacchi, A.J. Biological and physical signatures in the tropical and subtropical Atlantic. J. Geophys. Res. Ocean. 1999, 104, 18367–18382. [Google Scholar] [CrossRef] [Green Version]
- Bakker, D.C.E.; De Baar, H.J.W.; De Jong, E. Dissolved carbon dioxide in tropical East Atlantic surface waters. Phys. Chem. Earth 1999, 24, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, N. Low CO2 concentrations in the Gulf of Guinea during the upwelling season in 2006. Mar. Chem. 2009, 113, 93–101. [Google Scholar] [CrossRef]
- Caniaux, G.; Giordani, H.; Redelsperger, J.-L.; Guichard, F.; Key, E.; Wade, M. Coupling between the Atlantic cold tongue and the West African monsoon in boreal spring and summer. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef]
- Hisard, P. Variations saisonnières à l’équateur dans le golfe de Guinée. Cah. ORSTOM Sér. Océanograph. 1973, 11, 349–358. [Google Scholar]
- Hardman-Mountford, N.J.; McGlade, J.M. Seasonal and interannual variability of oceanographic processes in the Gulf of Guinea: An investigation using AVHRR sea surface temperature data. Int. J. Remote Sens. 2003, 24, 3247–3268. [Google Scholar] [CrossRef]
- Herbland, A.; Le Borgne, R.; Le Bouteiller, A.; Voituriez, B. Structure hydrologique et production primaire dans l’Atlantique tropical oriental (Hydrological structure and primary production in the eastern tropical Atlantic Ocean). Océanograph. Trop. 1983, 18, 249–293. [Google Scholar]
- Djagoua, É.V.; Affian, K.; Larouche, P.; Saley, B. Variabilité saisonnière et interannuelle de la chlorophylle en surface de la mer sur le plateau continental de la Côte d’Ivoire à l’aide des images de SeaWiFS de 1997 à 2004. Télédétection 2006, 6, 143–151. [Google Scholar]
- Nieto, K.; Mélin, F. Variability of chlorophyll-a concentration in the Gulf of Guinea and its relation to physical oceanographic variables. Prog. Oceanogr. 2017, 151, 97–115. [Google Scholar] [CrossRef]
- Abe, J.; Brown, B.; Ajao, E.A.; Donkor, S. Local to regional polycentric levels of governance of the Guinea Current Large Marine Ecosystem. Environ. Dev. 2016, 17, 287–295. [Google Scholar] [CrossRef]
- Radenac, M.H.; Jouanno, J.; Tchamabi, C.C.; Awo, M.; Bourlès, B.; Arnault, S.; Aumont, O. Physical drivers of the nitrate seasonal variability in the Atlantic cold tongue. Biogeosciences 2020, 17, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Jouanno, J.; Marin, F.; Du Penhoat, Y.; Molines, J.M.; Sheinbaum, J. Seasonal Modes of Surface Cooling in the Gulf of Guinea. J. Phys. Oceanogr. 2011, 41, 1408–1416. [Google Scholar] [CrossRef] [Green Version]
- Voituriez, B.; Herbland, A. Etude de la production pélagique de la zone équatoriale de l’Atlantique à 4oW: I—Relations entre la structure hydrologique et la production primaire. Cah. ORSTOM. Sér. Océanograph. 1977, 15, 313–331. [Google Scholar]
- Christian, J.R.; Murtugudde, R. Tropical Atlantic variability in a coupled physical–biogeochemical ocean model. Deep Sea Res. Part II Top. Stud. Oceanogr. 2003, 50, 2947–2969. [Google Scholar] [CrossRef]
- Andrié, C.; Oudot, C.; Genthon, C.; Merlivat, L. CO2 fluxes in the tropical Atlantic during FOCAL cruises. J. Geophys. Res. 1986, 91, 11741–11755. [Google Scholar] [CrossRef] [Green Version]
- Oudot, C.; Ternon, J.F.; Lecomte, J. Measurements of atmospheric and oceanic CO2 in the tropical Atlantic: 10 years after the 1982–1984 FOCAL cruises. Tellus 1995, 47, 70–85. [Google Scholar] [CrossRef] [Green Version]
- Parard, G.; Lefèvre, N.; Boutin, J. Sea water fugacity of CO2 at the PIRATA mooring at 6oS, 10oW. Tellus B 2010, 62, 636–648. [Google Scholar] [CrossRef]
- Lefèvre, N.; Veleda, D.; Araujo, M.; Caniaux, G. Variability and trends of carbon parameters at a time series in the eastern tropical Atlantic. Tellus B 2016, 68, 1147. [Google Scholar] [CrossRef] [Green Version]
- Bourlès, B.; Araujo, M.; McPhaden, M.J.; Brandt, P.; Foltz, G.R.; Lumpkin, R.; Giordani, H.; Hernandez, F.; Lefèvre, N.; Nobre, P.; et al. PIRATA: A Sustained Observing System for Tropical Atlantic Climate Research and Forecasting. Earth Space Sci. 2019, 6, 577–616. [Google Scholar] [CrossRef]
- Lefèvre, N.; Guillot, A.; Beaumont, L.; Danguy, T. Variability of fCO2 in the Eastern Tropical Atlantic from a moored buoy. J. Geophys. Res. 2008, 113, C01015. [Google Scholar] [CrossRef]
- Edmond, J.M. High precision determination of titration alkalinity and total carbon dioxide content of seawater by potentiometric titration. Deep Sea Res. 1970, 17, 737–750. [Google Scholar]
- Stramma, L.; Schott, F. The mean flow field of the tropical Atlantic Ocean. Deep Sea Res. Part II 1999, 46, 279–303. [Google Scholar] [CrossRef]
- Van Heuven, S.; Pierrot, D.; Rae, J.W.B.; Lewis, E.; Wallace, D.W.R. MATLAB Program Developed for CO2 System Calculations; Carbon Dioxide Information Analysis Center: Oak Ridge, TN, USA, 2011. [Google Scholar]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicz, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Res. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- McLaughlin, K.; Weisberg, S.B.; Dickson, A.G.; Hofmann, G.E.; Newton, J.A.; Aseltine-Neilson, D.; Barton, A.; Cudd, S.; Feely, R.A.; Jefferds, I.W.; et al. Core Principles of the California Current Acidification Network: Linking Chemistry, Physics, and Ecological Effects. Oceanography 2015, 28, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Millero, F.J.; Byrne, R.H.; Wanninkhof, R.; Feely, R.A.; Clayton, T.; Murphy, P.; Lamb, M.F. The internal consistency of CO2 measurements in the equatorial Pacific. Mar. Chem. 1993, 44, 269–280. [Google Scholar] [CrossRef]
- Sweeney, C.; Gloor, E.; Jacobson, A.R.; Key, R.M.; McKinley, G.; Sarmiento, J.L.; Wanninkhof, R. Constraining global air-sea gas exchange for CO2 with recent bomb14C measurements. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Weiss, R.F. CO2 in water and seawater: The solubility of a non-ideal gas. Mar. Chem. 1974, 2, 203–215. [Google Scholar] [CrossRef]
- Wentz, F.J.J.; Scott, R.; Hoffman, M.; Leidner, R.; Atlas, J.A. Remote Sensing Systems Cross-Calibrated Multi-Platform (CCMP) 6-Hourly Ocean Vector Wind Analysis Product on 0.25 deg Grid; Version 2.0.; Remote Sensing Systems: Santa Rosa, CA, USA, 2015; Available online: www.remss.com/measurements/ccmp (accessed on 30 September 2020).
- Koffi, U.; Lefèvre, N.; Kouadio, G.; Boutin, J. Surface CO2 parameters and air-sea CO2 flux distribution in the eastern equatorial Atlantic Ocean. J. Mar. Syst. 2010, 82, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Millero, F.J.; Lee, K.; Roche, M. Distribution of alkalinity in the surface waters of the major oceans. Mar. Chem. 1998, 60, 111–130. [Google Scholar] [CrossRef]
- Enfield, D.B.; Mestas, A.M.; Mayer, D.A.; Cid-Serrano, L. How ubiquitous is the dipole relationship in tropical Atlantic sea surface temperatures? J. Geophys. Res. 1999, 104, 7841–7848. [Google Scholar] [CrossRef]
- Gallardo, Y.; Dandonneau, Y.; Voituriez, B. Variabilité, Circulation et Chlorophylle Dans la Région du Dôme d’Angola en Février-Mars 1971; Documents Scientifiques; Centre de Recherches Océanographiques: Abidjan, Cote d’Ivoire, 1974; Volume 5, pp. 1–51. [Google Scholar]
- Wang, Z.A.; Bienvenu, D.J.; Mann, P.J.; Hoering, K.A.; Poulsen, J.R.; Spencer, R.G.M.; Holmes, R.M. Inorganic carbon speciation and fluxes in the Congo River. Geophys. Res. Lett. 2013, 40, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, J.; Lucas, M.; Dufau, C.; Sutton, M.; Stum, J.; Lauret, O.; Channelliere, C. Detection and variability of the Congo River plume from satellite derived sea surface temperature, salinity, ocean colour and sea level. Remote Sens. Environ. 2013, 139, 365–385. [Google Scholar] [CrossRef]
- Materia, S.; Gualdi, S.; Navarra, A.; Terray, L. The effect of Congo River freshwater discharge on Eastern Equatorial Atlantic climate variability. Clim. Dyn. 2012, 39, 2109–2125. [Google Scholar] [CrossRef]
- Bonou, F.K.; Noriega, C.; Lefèvre, N.; Araujo, M. Distribution of CO2 parameters in the Western Tropical Atlantic Ocean. Dyn. Atmos. Ocean. 2016, 73, 47–60. [Google Scholar] [CrossRef]
- Vangriesheim, A.; Pierre, C.; Aminot, A.; Metzl, N.; Baurand, F.; Caprais, J.-C. The influence of Congo River discharges in the surface and deep layers of the Gulf of Guinea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 2183–2196. [Google Scholar] [CrossRef] [Green Version]
- Neto, A.V.N.; Giordani, H.; Caniaux, G.; Araujo, M. Seasonal and Interannual Mixed-Layer Heat Budget Variability in the Western Tropical Atlantic From Argo Floats (2007–2012). J. Geophys. Res. Ocean. 2018, 123, 5298–5322. [Google Scholar] [CrossRef]
- Ibánhez, J.S.P.; Flores, M.; Lefèvre, N. Collapse of the tropical and subtropical North Atlantic CO2 sink in boreal spring of 2010. Sci. Rep. 2017, 7, 41694. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Marchine Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Bishop, C.M. Networks for Pattern Recognition; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Chollet, F. Keras. 2015. Available online: https://github.com/keras-team/keras (accessed on 31 October 2020).
- Dean, J.; Monga, R. TensorFlow: Large-Scale Machine Learning on Heterogeneous Systems. 2015. Available online: https://www.tensorflow.org/ (accessed on 31 October 2020).







| Pair | Calculated Parameter | RMSE | r |
|---|---|---|---|
| TA–DIC | fCO2 | 11.7 atm | 0.90 |
| DIC–fCO2 | TA | 7.4 mol kg−1 | 0.99 |
| fCO2–TA | DIC | 5.9 mol kg−1 | 0.99 |
| Mooring or Cruise | RMSE * | r | N | Time Period |
|---|---|---|---|---|
| 6° S, 10° W | 9.7 mol kg−1 | 0.96 | 6611 | 2006–2017 |
| 6° S, 8° E | 14.4 mol kg−1 | 0.99 | 239 | 2017–2019 |
| EGEE 3 | 9.8 mol kg−1 | 0.98 | 6895 | 2006 |
| PIRATA FR-29 | 10.0 mol kg−1 | 0.99 | 4462 | 2019 |
| Mixing Equation | Salinity Range | Reference |
|---|---|---|
| DIC = 50.6 (±2.0) ∗ SSS + 231.7 (±62.2) | S > 27 | This work |
| DIC = 54.0 ∗ S + 109 | S > 33 | [13] |
| DIC = 46.5 (±1) ∗ SSS + 355 (±48) | S > 22 | [49] |
| Site | CO2 Flux | ΔfCO2 | DIC | SSS | SST | Wind |
|---|---|---|---|---|---|---|
| (mmol m−2d−1) | (atm) | (mol kg−1) | (°C) | (m s−1) | ||
| 6° S, 8° E | 3.06 ± 1.74 | 59 ± 28 | 1973 ± 87 | 34.3 ± 1.1 | 26.6 ± 2.8 | 5.0 ± 0.6 |
| 6° S, 10° W | 5.61 ± 1.49 | 55 ± 16 | 2047 ± 27 | 35.9 ± 0.2 | 26.9 ± 1.7 | 7.1 ± 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefèvre, N.; Mejia, C.; Khvorostyanov, D.; Beaumont, L.; Koffi, U. Ocean Circulation Drives the Variability of the Carbon System in the Eastern Tropical Atlantic. Oceans 2021, 2, 126-148. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010008
Lefèvre N, Mejia C, Khvorostyanov D, Beaumont L, Koffi U. Ocean Circulation Drives the Variability of the Carbon System in the Eastern Tropical Atlantic. Oceans. 2021; 2(1):126-148. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010008
Chicago/Turabian StyleLefèvre, Nathalie, Carlos Mejia, Dmitry Khvorostyanov, Laurence Beaumont, and Urbain Koffi. 2021. "Ocean Circulation Drives the Variability of the Carbon System in the Eastern Tropical Atlantic" Oceans 2, no. 1: 126-148. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2010008