
Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Morphological Analyses and Specimen Collection
2.3. DNA Extraction, PCR and ITS 2
2.4. Phylogenetic Analysis
3. Results
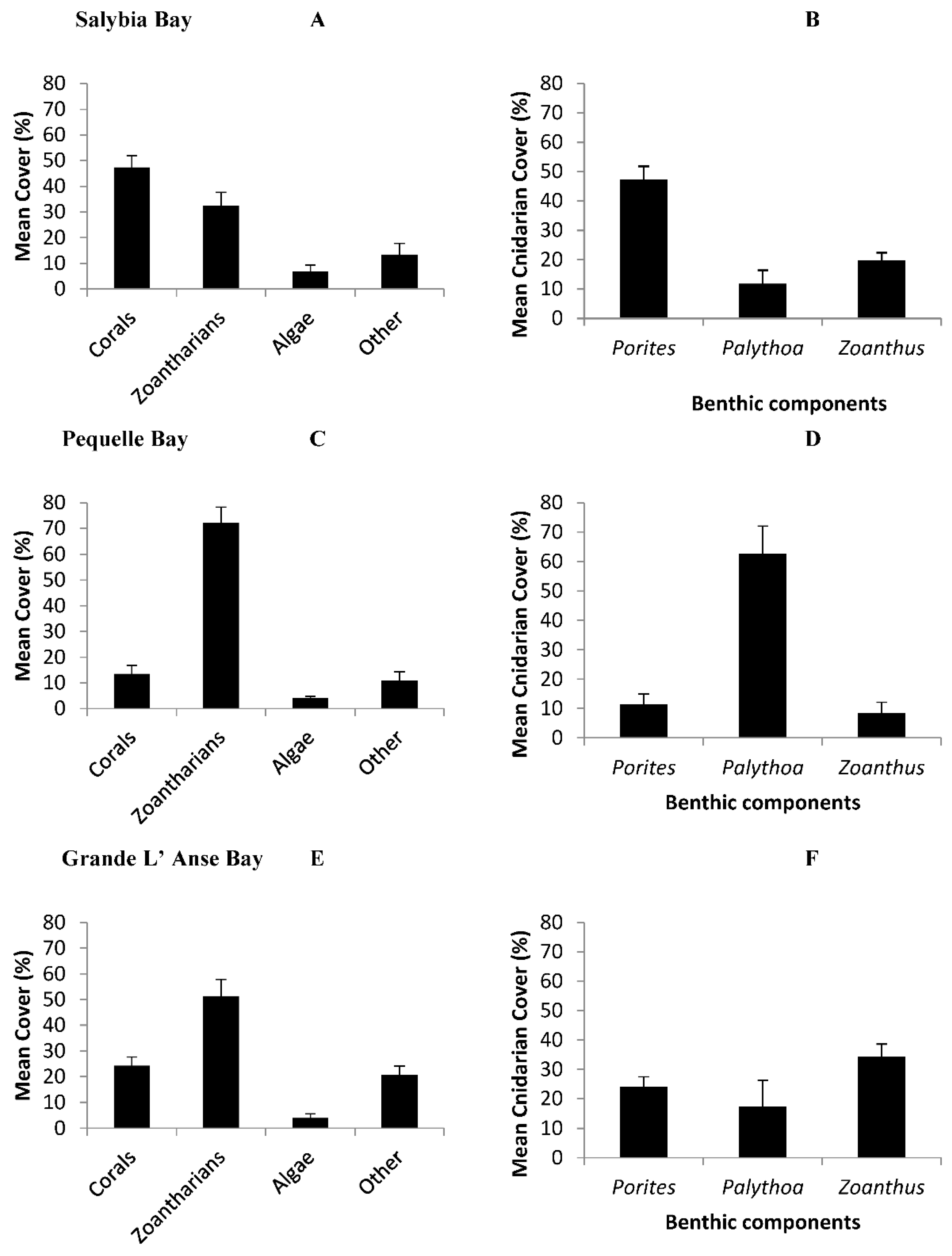
3.1. Zoantharian Distribution
3.2. Specimen Morphological and Molecular Analyses
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karlson, R.H. Alternate competitive strategies in a periodically disturbed habitat. Bull. Mar. Sci. 1980, 30, 894–900. [Google Scholar]
- Irei, Y.; Nozawa, Y.; Reimer, J.D. Distribution patterns of five zoanthid species in Okinawa Island, Japan. Zool. Stud. 2011, 50, 426–433. [Google Scholar]
- Irei, Y.; Sinniger, F.; Reimer, J.D. Description of two azooxanthellate Palythoa species (Subclass Hexacorallia, Order Zoantharia) from Ryukyu Archipelago, southern Japan. Zookeys 2015, 478, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimer, J.D.; Shusuke, O.; Takishita, K.; Tsukahara, J.; Maruyama, T. Molecular evidence suggesting species in the zoanthid genera Palythoa and Protopalythoa (Anthozoa: Hexacorallia) are congeneric. Zool. Sci. 2006, 23, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinniger, F.; Montoya-Burgos, J.I.; Chevaldonné, P.; Pawlowski, J. Phylogeny of the order Zoantharia (Anthozoa, Hexacorallia) based on the mitochondrial ribosomal genes. Mar. Biol. 2005, 147, 1121–1128. [Google Scholar] [CrossRef]
- Karlson, R.H. Reproduction patterns on Zoanthus spp. from Discovery Bay, Jamaica. Proceedings 4th International Coral Reef Symposium. Manilla 1981, 2, 699–704. [Google Scholar]
- Sebens, P.S. Intertidal distribution of zoanthids on the Caribbean coast of Panama: Effects of predation and desiccation. Bull. Mar. Sci. 1982, 32, 316–335. [Google Scholar]
- Reimer, J.D.; Ono, S.; Fujiwara, Y.; Takishita, K.; Tsukahara, J. Reconsidering Zoanthus spp. diversity: Molecular evidence of conspecificity within four previously presumed species. Zool. Sci. 2004, 21, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Belford, S.G.; Phillip, D.A.T. Rapid assessment of a coral reef community in a marginal habitat in the southern Caribbean: S simple way to know what’s out there. Asian J. Biol. Sci. 2011, 4, 520–531. [Google Scholar] [CrossRef] [Green Version]
- Belford, S.G.; Phillip, D.A.T. Intertidal distribution patterns of zoanthids compared to their scleractinian counterparts in the southern Caribbean. Int. J. Oceanogr. Mar. Ecol. Syst. 2012, 3, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Reimer, J.D.; Wee, H.B.; García-Hernández, J.E.; Hoeksema, B.W. Zoantharia (Anthozoa: Hexacorallia) abundance and associations with Porifera and Hydrozoa across a depth gradient on the west coast of Curaçao. Syst. Biodivers. 2018, 16, 820–830. [Google Scholar] [CrossRef]
- Belford, S.G.; Phillip, D.A.T.; Rutherford, M.G.; Schmidt, R.S.; Duncan, E.J. Biodiversity of coral reef communities in marginal environments along the north-eastern coast of Trinidad, southern Caribbean. Prog. Aqu. Farm. Mar. Biol. 2019, 2, 180017. [Google Scholar]
- Goreau, T.F. The ecology of Jamaican coral reefs I. Species composition and zonation. Ecology 1959, 40, 67–90. [Google Scholar] [CrossRef]
- Burnett, W.J.; Benzie, J.A.H.; Beardmore, J.A.; Ryland, J.S. Zoanthids (Anthozoa, Hexacorallia) from the Great Barrier Reef and Torres Straights, Australia: Systematics, evolution and a key to species. Coral Reefs 1997, 16, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Ryland, J.S.; Lancaster, J.E. Revision of methods for separating species of Protopalythoa (Hexacorallia: Zoanthidea) in the tropical West Pacific. Invert. Syst. 2003, 17, 407–428. [Google Scholar] [CrossRef] [Green Version]
- López, C.; Reimer, J.D.; Brito, A.; Simón, D.; Clemente, S.; Hernández, M. Diversity of zoantharian species and their symbionts from Macaronesian and Cape Verde ecoregions demonstrates their widespread distribution in the Atlantic Ocean. Coral Reefs 2019, 38, 269–283. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.S. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbiont. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, W.J. Longitudinal variation in algal symbionts (zooxanthellae) from the Indian Ocean zoanthid Palythoa caesia. Mar. Ecol. Prog. Ser. 2002, 234, 105–109. [Google Scholar] [CrossRef]
- Reimer, J.D.; Shusuke, O.; Yasuo, F.; Junzo, T. Seasonal changes in morphological condition of symbiotic dinoflagellates (Symbiodinium spp.) in Zoanthus sansibaricus (Anthozoa: Hexacorallia) in Southern Japan. South Pac. Stud. 2007, 27, 2. [Google Scholar]
- Reimer, J.D.; Foord, C.; Irei, Y. Species diversity of shallow water zoanthids (Cnidaria: Anthozoa: Hexacorallia) in Florida. Hindawi Publ. Corp. J. Mar. Biol. 2012, 2012, 856079. [Google Scholar] [CrossRef] [Green Version]
- Pax, F. Studien an westindischen Actinien. In Ergebnisse einer Zoologischen nach Westindien von Prof. W. Kukenthal und Dr. R. Hartmeyer im Jahre, 1907; Spengel, J.W., Ed.; Zoologische Jahrbucher Supplement: Ann Arbor, MI, USA, 1910; Volume 11, pp. 157–330. [Google Scholar]
- Sinniger, F.; Reimer, J.; Pawlowski, J. The Parazoanthidae (Hexacorallia: Zoantharia) DNA taxonomy: Description of two new genera. Mar. Biodivers. 2010, 40, 57–70. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Trench, R. Biogeography of two species of Symbiodinium (Freudenthal) inhabiting the intertidal sea anemone Anthopleura elegantissima (Brandt). Biol. Bull. 2000, 199, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaJeunesse, T.C. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs. Mar. Biol. 2002, 141, 387–400. [Google Scholar]
- LaJeunesse, T.C.; Thornhill, D.J. Improved resolution of reef-coral endosymbiotic dinoflagellate (Symbiodinium) species diversity, ecology, and evolution through psbA non-coding region genotyping. PLoS ONE 2011, 6, e29013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Strecher, G.; Li, M.; Knyaz, C.; Tamura, K. Molecular evolutionary genetic analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, E.F.; Soares, M.D.; Bezerra, L.E.; Matthews-Cascon, H. Distribution patterns of zoanthids (Cnidaria: Zoantharia) on a tropical reef. Mar. Biol. Res. 2015, 11, 584–592. [Google Scholar] [CrossRef]
- Belford, S.G. Spatial abundance and colour morphotype densities of the rock boring sea urchin (Echinometra lucunter) at two different habitats. Thalassas 2020, 36, 157–164. [Google Scholar] [CrossRef]
- López, C.; Freitas, R.; Magileviciute, E.; Ratão, S.S.; Brehmer, P.; Reimer, J.D. Report of a Zoanthus zone from the Cabo Verde islands (Central eastern Atlantic). Thalassas 2018, 34, 409–413. [Google Scholar] [CrossRef]
- Ong, C.W.; Reimer, J.D.; Todd, P.A. Morphological plastic responses to shading in the zoanthids Zoanthus sansibaricus and Palythoa tuberculosa. Mar. Biol. 2013, 160, 1053–1064. [Google Scholar] [CrossRef]
- Sinniger, F.; Reimer, J.D.; Pawlowski, J. Potential of DNA sequences to identify zoanthids (Cnidaria: Zoantharia). Zool. Sci. 2008, 25, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Loh, W.; Van Woesik, R.; Hoegh-Guldberg, O.; Schmidt, G.; Fitt, W. Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Am. Soc. Limnol. Oceanogr. 2003, 48, 2046–2054. [Google Scholar] [CrossRef] [Green Version]
- Kumara, S.; Zacharia, P.U.; Sreenath, K.R.; Kripa, V.; George, G. GIS based mapping of zoanthids along Saurashtra coast, Gujarat, India. J. Mar. Biol. Assoc. India 2017, 59, 19–25. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate change human impacts and the resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample # | Collection Site α | Disk Color | Tentacle Color | Tentacle Count (3 Polyps Per Colony) | Disk Diameter (mm) | COI I.D. (GenBank Accession #) |
|---|---|---|---|---|---|---|
| P1-br-GA | GA | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ150796) |
| P2-br-GA | GA | Brown | Brown | 36 | 10.8 | P. caribaeorum (MZ150797) |
| P3-br-SB | SB | Brown | Brown | 37 | 10.5 | P. caribaeorum (MZ150798) |
| P4-gr-GA | GA | Green | Brown | 46 | 13.8 | P. grandiflora (MZ150799) |
| P5-gr-GA | GA | Green | Brown | 45 | 12.6 | P. grandiflora (MZ150800) |
| P6-br-GA | GA | Brown | Brown | 33 | 10.7 | P. caribaeorum (MZ150801) |
| P7-br-WSB | WSB | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ147090) |
| P8-br-WSB | WSB | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ147091) |
| Z1-gr-StB | StB | Green/Blue | Green | 41 | 10.2 | Z. sociatus (MZ147096) |
| Z2-gr-StB | StB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ147097) |
| Z3-gr-WSB | WSB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ150806) |
| Z4-gr-WSB | WSB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ150802) |
| Z5-gr-SB | SB | Green | Green | 42 | 7.5 | Z. pulchellus (MZ150803) |
| Z6-gr-SB | SB | Green | Green | 42 | 6.9 | Z. pulchellus (MZ156026) |
| Z7-br-gr-SB | SB | Green/Blue | Green | 42 | 7.6 | Z. sociatus (MZ150807) |
| Z8-org-SB | SB | Green | Green | 46 | 10.2 | Z. pulchellus (MZ150805) |
| Z9-org-GA | GA | Orange | Brown | 46 | 10.3 | Z. pulchellus (MZ150804) |
| Z10-gr-TLH | TLH | Green | Green | 42 | 10.3 | Z. pulchellus (MZ147092) |
| Z13-grey-GA | SB | Grey | Green | 44 | 5.2 | Z. pulchellus (MZ147093) |
| Z14-org-GA | SB | Orange | Brown | 45 | 10.2 | Z. pulchellus (MZ147094) |
| Z15-blu-TLH | TLH | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ147095) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belford, S. Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean. Oceans 2021, 2, 477-488. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030027
Belford S. Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean. Oceans. 2021; 2(3):477-488. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030027
Chicago/Turabian StyleBelford, Stanton. 2021. "Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean" Oceans 2, no. 3: 477-488. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030027