The Role of Taste Receptors in Airway Innate Immune Defense


1
Department of Otorhinolaryngology, Head and Neck Surgery, University of Pennsylvania, Philadelphia, PA 19104, USA
2
Monell Smell and Taste Center, Philadelphia, PA 19104, USA
3
Philadelphia Veterans Affairs Medical Center, Philadelphia, PA 19104, USA
*
Author to whom correspondence should be addressed.
Sinusitis 2018, 3(2), 6; https://0-doi-org.brum.beds.ac.uk/10.3390/sinusitis3020006
Submission received: 14 May 2018
/
Revised: 3 June 2018
/
Accepted: 5 June 2018
/
Published: 7 June 2018
Abstract
:Bitter (T2R) and sweet (T1R) taste receptors are expressed in the upper airway, where they play key roles in antimicrobial innate immune defense. Bitter bacterial products are detected by taste receptors on ciliated cells and solitary chemosensory cells, resulting in downstream nitric oxide and antimicrobial peptide release, respectively. Genetic polymorphisms in taste receptors contribute to variations in T1R and T2R functionality, and phenotypic differences correlate with disease status and disease severity in chronic rhinosinusitis (CRS). Correspondingly, there are also subjective bitter and sweet taste differences between patients with CRS and individuals without CRS across a number of compounds. The ability to capture these differences with a simple and inexpensive taste test provides a potentially useful diagnostic tool, while bitter compounds themselves could potentially serve as therapeutic agents. The present review examines the physiology of airway taste receptors and the recent literature elucidating the role taste receptors play in rhinologic disease.
1. Introduction
Taste receptors are typically associated with oral sensory perception as an adaptive mechanism for detecting energy rich foods as well as poisons and other unpalatable compounds. Bitter taste receptors are a specific subset of taste receptors that classically respond to toxins, chemicals, and other aversive products that can be detrimental to organismal health. However, recent research has identified taste receptors in many other anatomic compartments of the body with a variety of functions extending far beyond the canonical sensory capacity of the tongue [1,2,3,4,5,6]. Taste receptors have been found in the brain, pancreas, testicles, bladder, and gastrointestinal tract [1,2,3,4,7]. This review will examine the role of bitter and sweet taste receptors that are expressed in the airway, and the important roles that these taste receptors play in innate immune defense [8,9].
2. Taste Receptor Physiology
Bitter and sweet taste receptors, unlike the ion-sensitive salt and sour taste receptors, are G-protein coupled receptors (GPCRs) [10,11]. The sweet taste receptor (T1R) family responds to sugars, including sucrose, glucose, and fructose, and T1Rs are classified as a part of taste receptor family 1 subtype 2 and 3 (TAS1R2/TAS1R3) [5,12,13]. A wider variety of bitter taste receptors exist in taste receptor family 2 (T2Rs), and these diverse receptors respond to an assortment of bitter compounds [14], including sesquiterpene lactones, strychnine, and denatonium [15]. Each bitter taste receptor can respond to a multitude of chemically similar compounds, and each compound can stimulate more than one taste receptor. Humans have at least 25 T2R subtypes, reflecting a broad perceptual range [12,16]. There is also a high degree of genetic diversity in T2Rs. On a phenotypic level, this results in differing sensitivity to specific bitter compounds among individuals, both on the tongue and in the airway. This diversity partially explains variation in taste preferences between groups and within groups [17,18]. For example, certain individuals find some bitter foods, such as coffee, to be aversive, while others are less sensitive.
The mechanisms involved in taste receptor activation are relatively conserved and follow similar pathways in the tongue and airway. However, while the expression of taste receptors in the sinonasal epithelium is ubiquitous on disparate cell types, including ciliated cells and solitary chemosensory cells, in the tongue taste receptor expression is confined to type II cells within the taste buds. Furthermore, while some bitter taste receptors in the airway are upstream of a nervous signaling cascade, others act in a cell-autonomous fashion only [8,19,20]. When a ligand binds to a taste GPCR, there is activation of phospholipase C isoform β2 (PLCB2), which triggers downstream inositol 1,4,5-trisphosphate (IP3) production. The IP3 receptor on the endoplasmic reticulum releases calcium in response to this increase in IP3 [21]. Simultaneously, there is also an activation of phosphodiesterases (PDEs) that attenuate cyclic adenosine monophosphate (cAMP) levels and protein kinase A (PKA) activity. As PKA is an inhibitor of an IP3 receptor isoform, removal of this inhibition causes further release of calcium from the endoplasmic reticulum [22]. Calcium ultimately activates the non-selective cation channel, TRPM5, that causes cellular depolarization, activates voltage-gated sodium (Na+) channels, and ultimately results in an action potential that causes ATP release through CALHM1, a large pore channel [5,22,23,24,25]. In the tongue, this ATP activates receptors on taste cells and sensory fibers that transmit sensations to the central nervous system [5,25,26].
3. Bitter Taste Receptors in the Airway
Many different bitter taste receptors are expressed in the rodent and human airways [9,27,28,29,30] and in these locations, they respond to bitter bacterial products that are produced. One example of this is the lactone class of bitter compounds, which includes acyl-homoserine lactones (AHLs) that are produced by many gram-negative bacteria [31,32]. These lactones serve as biofilm “quorum-sensing molecules”; bacteria will initiate biofilm formation when a high enough concentration of AHLs is reached in a localized area. Biofilms can provide protection for bacteria from host innate immune defenses as well as antibiotics [33]. It is hypothesized that bitter taste receptors attempt to “spy” on these bacterial communications, effectively detecting AHLs before a sufficient concentration is reached for biofilm formation [8]. The bitter taste receptors themselves elicit innate immune responses that can eradicate bacteria before pathogenic levels are achieved.
This highlights a critical component of upper airway immunity: recognition of foreign bacteria, viruses, fungi, or toxins, followed by prompt reduction in pathogenic biomass. Toll-like receptors (TLRs) respond to pathogen-associated molecular patterns (PAMPs), which include foreign cellular components. However, TLR signaling is gradual, taking up to 12 h to exert an immune response through changes in expression of genes that play a role in innate immunity [34]. Conversely, bitter taste receptors can detect bacterial products, such as AHLs, and elicit downstream increases in immune defenses in a much more expedient fashion (seconds to minutes).
3.1. Bitter Taste Receptors on Ciliated Cells
Bitter taste receptors on ciliated cells respond to bacterial compounds and elicit a potent downstream response of nitric oxide (NO) production [35,36] (Figure 1). Nitric oxide diffuses quickly into bacteria, where it participates in destruction of cellular components [9,37]. Some bacteria, such as Pseudomonas aeruginosa, are highly sensitive to NO, while others are more resistant [38]. In addition to this antimicrobial activity, NO also activates protein kinase G (PKG) and guanylyl cyclase to directly speed up ciliary beat frequency (CBF), increasing mucociliary clearance [39]. Rapid ciliary beating can clear bacteria and mucus to the nasopharynx or oropharynx, where they can be eliminated by swallowing. Additionally, released innate immune products are spread out across the airway surface by ciliary beating [40]. These compounds—including lactoferrin, lysozyme, and defensins—act in concert with NO and other reactive oxygen species to create a potent antimicrobial response [41].
T2R38 is a bitter taste receptor located on ciliated cells in humans, and it responds to at least three AHLs produced by P. aeruginosa: N-butyryl-l-homoserine lactone, N-hexanoyl-l-homoserine lactone and N-3-oxo-dodecanoyl-l-homoserine lactone [9]. In addition to its response to bacterial compounds, T2R38 reacts in a similar fashion to the compounds phenylthiocarbamide (PTC) and propylthiouracil (PROP) [42]. In response to PTC stimulation, sinonasal epithelial cells expressing a functional T2R38 receptor demonstrate a substantial increase in NO production. Importantly, the TRPM5 channel and PLCβ2 are necessary for this NO response, and these are two canonical components of taste signaling. Interestingly, the taste G-protein gustducin does not appear to be involved [9]. The resultant NO production following PTC stimulation is sufficient for a highly bactericidal response.
Just as the genetic variation in T2Rs can cause differences in taste preferences on the tongue, receptor variation in the airway also appears to play a key role in the ability to mount a respiratory defense in response to bitter compounds. The genetic locus for T2R38, TAS2R38, has common polymorphisms that can render the receptor non-functional. Individuals with a proline-alanine-valine (PAV) amino acid sequence at a key portion of the taste receptor are able to respond to T2R38 agonists, while individuals with an alanine-valine-isoleucine (AVI) sequence at this same locus possess a non-functional receptor variant [18]. Cells isolated from individuals with an AVI/AVI genotype show highly attenuated NO production in response to AHLs, PTC, or PROP stimulation, compared to cells isolated from individuals with a PAV/PAV genotype. Downstream reductions in mucociliary clearance and bacterial killing are correspondingly observed [9]. As would be expected, AVI/AVI individuals also do not taste PTC or PROP when presented with an oral taste test challenge [43].
This reduction in responsiveness observed in AVI-expressing individuals has clinical consequences. Several studies in the past five years have highlighted a potential relevance of T2R38 in chronic rhinosinusitis (CRS). Individuals who express the fully functional, PAV/PAV genotype are less likely to require surgical intervention for CRS symptoms than patients with an AVI/AVI genotype [43,44]. Additionally, levels of gram-negative infection are lower in PAV/PAV patients [43,44,45,46], confirming that the NO-dependent response of T2R38 acts as a critical defense for this class of bacteria. A hallmark of CRS is mucociliary stasis, in which bacteria are inadequately cleared. At pathogenic levels of proliferation, bacterial toxins can be destructive to cells and cilia, perpetuating the process of impaired mucociliary function [47]. It is known that sinonasal explants from patients with CRS have an attenuated response to a variety of compounds (bitter and non-bitter) that stimulate ciliary beating in control tissue [48]. Other studies, while part of an inconclusive set of literature, have shown differences in NO levels in patients with airway diseases [49]. Without the action of NO to kill bacteria and increase ciliary beating in response to AHLs, it appears that the non-functional T2R38 polymorphism has a phenotypic effect on upper airway disease [9].
Other bitter taste receptors on ciliated cells, such as T2R4 and T2R14 [50], respond to different bitter agonists, such as quinine hydrochloride. Quinine is an alkaloid derivative that is isolated from the cinchona tree, and is found in several medicinal and commercial products [51]. Recent work shows that quinine stimulates a rapid T2R-dependent NO response from ciliated cells in the airway [52]. While quinine is a more promiscuous bitter taste receptor agonist than PTC or PROP, there are common genetic variants in bitter taste receptor genes on chromosome 12 that strongly contribute to the perception of quinine taste intensity [53]. Quinine taste sensitivity has also been selected independently in some world populations, especially for low concentrations of quinine [54]. Concentrations of bitter microbial products in the airway are also at low concentrations [9], and these differences in taste perception of dilute quinine solutions may be reflective of varying responses of these bitter taste receptors in both the airway and on the tongue. Allele expression studies have shown that patients with CRS differ from control patients at several genetic loci for taste receptors, including TAS2R14 and TAS2R49 [45].
3.2. Taste Receptors on Solitary Chemosensory Cells
Solitary chemosensory cells (SCCs) are a non-ciliated airway cell type that is relatively rare, representing approximately 1% of the total upper airway epithelial cell population [55]. These cells are immunoreactive with α-gustducin, a taste signalling component, and they share many similarities with taste bud cells [28]. Because of their rarity, they are difficult to isolate experimentally [19]. Solitary chemosensory cells express both sweet and bitter taste receptors that are capable of responding to a variety of compounds [8,20,27,56,57]. In response to bitter stimulation, these cells do not activate NO production, but instead mediate a separate cohort of responses. In mouse SCCs, the calcium response resulting from bitter taste receptor stimulation causes acetylcholine (ACh) release that has breath holding effects and also results in downstream inflammatory mediator release [8,19,20]. Both of these responses are at least partially immunomodulatory in nature: breath holding limits toxin or organism aspiration, while inflammatory mediators often participate in a larger immune signaling cascade. In the human upper airway, SCC stimulation results in the calcium-mediated release of antimicrobial peptides from adjacent ciliated cells, including β-defensin 1 (DEFB1) and β-defensin 2 (DEFB2) [29,58] (Figure 1). These defensins are potently antimicrobial and have effects on both gram-positive and gram-negative bacteria, including methicillin-resistant Staphylococcus aureus and P. aeruginosa [59]. Unlike the antimicrobial peptide release observed with TLR stimulation, which occurs over several hours as a result of changes in messenger RNA [34], bitter taste receptor stimulation causes release of pre-formed stores of antimicrobial peptides. Denatonium is a specific bitter compound that has agonist properties for bitter taste receptors on SCCs, and application of denatonium to airway epithelial cells from mouse and human cultures stimulates calcium responses that spread to adjacent cells via gap junctions [29]. Similar to the cascades observed in bitter taste receptor stimulation in ciliated cells, the calcium responses from SCC stimulation also require canonical components of taste signaling, such as TRPM5, PLCβ2, and gustducin [29].
In addition to expressing bitter taste receptors, SCCs also express sweet taste receptors, the T1Rs [27,29,60]. These receptors are sensitive to sweet compounds, such as glucose, in concentrations far lower than those detected orally [61]. Typically, airway surface liquid (ASL) glucose levels are maintained at a homeostatic level of approximately 0.5 mM; there is a physiologic leak and continuous reuptake of glucose from the adjacent basolateral serum that maintains this concentration [29]. At this physiologic level of glucose, T1R2 and T1R3 receptors are tonically activated. The activation of sweet taste receptors on SCCs appears to antagonize the action of bitter taste receptor cascades through activation of cAMP and phosphodiesterase which subsequently block activation of the IP3 receptor [29]. During bacterial infection, there is a reduction in ASL glucose due to increased bacterial consumption. It is hypothesized that it is this reduction in glucose that causes a reduction in sweet taste receptor activation, resulting in a corresponding increase in bitter taste receptor activity and responsivity to microbial bitter products [29]. Thus, the balance tips in favor of T2R responses and mobilization of innate immune defenses, theoretically restoring the balance towards airway microbial homeostasis and normalized glucose concentrations.
Several experiments have been conducted to support this hypothesis of antagonistic actions of bitter and sweet taste receptors. When glucose or sucrose is added to airway surface liquid of in vitro mouse cultures, calcium responses to denatonium are greatly diminished. Mice that were genetically modified to not express sweet taste receptors showed a normal SCC response to denatonium under the same conditions [29,62]. Additional experiments have shown that T1Rs can also be activated by d-amino acids produced by bacteria. Lee et al. demonstrated that at least two T1R-activating d-amino acids produced by S. aureus suppress SCC calcium responses, with corresponding decreases in antimicrobial peptide secretion [58]. These d-amino acids may be produced by the bacteria for protection from host innate immune responses and may allow for increased colonization and potential opportunistic infection. Just as observed with T2Rs, there is TAS1R genetic variation that contributes to preferences in oral sweet taste perception [63]. Several allelic variations in TAS1R genes demonstrate frequency differences of greater than 10% when comparing CRS patients and control individuals [45]. Just as is the case with bitter receptors, there is genetic variation in TAS1R genes that manifests as individual preference in sweet taste [63]. While no single locus has yet been identified, there are allele variations among the TAS1R genes that show frequency differences of >10% in 16 loci between patients with CRS and controls [45].
3.3. Bitter and Sweet Taste Testing
Based on the existing evidence that genetic variation in bitter and sweet taste receptors is correlated with disease status and disease severity, phenotypic oral taste tests may be clinically useful to assess taste receptor variation. On a broad level, individuals with insensitive bitter taste receptors or hypersensitive sweet taste receptors would be expected to be overrepresented in a cohort of individuals with CRS. Two recent papers report on taste testing results in hundreds of patients with CRS as well as propensity matched control individuals without rhinologic disease [52,64]. Patients with CRS without nasal polyps perceived denatonium, an SCC T2R agonist, as having lower subjective intensity, while patients with CRS with nasal polyps perceived quinine, a ciliated cell T2R agonists, as having lower subjective intensity as well (Table 1). All CRS patients rated sucrose, a T1R agonist, as having higher subjective intensity. There was no difference in taste sensitivity to a neutral compound, sodium chloride, between CRS and control patients [52,64]. When bitter and sweet taste ratings were aggregated into an overall “score” that took into account the opposing physiologic effects of bitter and sweet taste receptor stimulation in SCCs, there were even more highly significant differences between CRS and control subjects. Some of these subjective taste differences also appear to be reflected at the physiologic level; experiments have shown an inverse association between in vitro biofilm formation and PTC taste intensity ratings [65]. The implications for these differences are broad. Physiologically, this may reflect a less active SCC response to bitter microbial products in the airway of CRS patients, with an additional compounding effect of an increased sensitivity to glucose. This increased glucose sensitivity would perhaps inhibit SCC T2R immunomodulatory function even with intact T2R responses, due to the relative T1R affinity. These phenotypic differences can also help explain why CRS tends to run in families, suggesting a critical genetic influence in the disease [66]. Furthermore, the stratification of patient sensitivity in ciliated and SCCs T2Rs based on CRS polyp status is additionally interesting, as this may demonstrate unique T2R contributions to different types of CRS.
3.4. Diagnostics and Therapeutics
Oral taste tests are inexpensive to produce and administer, and the ability to assess variations in airway taste receptor functionality could help predict impaired innate immunity or predisposition to respiratory disease. Bitter taste testing with specific agonists, such as PTC, could potentially be used to stratify surgical candidates or identify individuals who should receive more aggressive management. With optimized taste testing compound concentrations that reflect respiratory tract affinity levels, improved patient stratification for CRS and control patients could be achieved. Additionally, the utilization of multiple bitter and sweet compounds for taste testing could improve performance parameters with an overall “taste score”. Beyond the diagnostic realm, bitter taste receptor agonists may have therapeutic potential as topical agents in harnessing potent innate immune defenses as an alternative to more conventional treatments, such as antibiotics. T1R antagonists, such as lactisole and amiloride, could also release the inhibition of T1Rs on antimicrobial responses [29,67].
4. Conclusions
Bitter and sweet taste receptors play important roles in the regulation of innate immune defenses in the upper respiratory tract. Bitter taste receptors mediate rapid antimicrobial nitric oxide or β-defensin responses in the presence of bacterial compounds, while sweet taste receptors attenuate these responses at higher levels of glucose. There is a high degree of genetic variation in airway taste receptors, and genetic polymorphisms can predispose people to recalcitrant CRS and create an increased susceptibility to infection. Phenotypic oral taste tests can capture some of this taste receptor variation, correlating with disease status in CRS. Further, bitter taste receptor agonists and sweet taste receptor antagonists can potentially serve as alternative therapies for respiratory disease that harness endogenous immune defenses.
Author Contributions
A.D.W., N.N.P., R.M.C., and E.C.K. drafted the manuscript, N.A.C. conceived and designed many experiments described and critically reviewed the manuscript.
Funding
This work was supported by a grant from the RLG Foundation, In, and USPHS grant R01DC013588.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral and extraoral tissues. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.A.; Liggett, S.B.; Munger, S.D. Extraoral bitter taste receptors as mediators of off-target drug effects. FASEB J. 2012, 26, 4827–4831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depoortere, I. Taste receptors of the gut: Emerging roles in health and disease. Gut 2014, 63, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Meyerhof, W. Oral and extraoral bitter taste receptors. Results Probl. Cell Differ. 2010, 52, 87–99. [Google Scholar] [PubMed]
- Kinnamon, S.C. Taste receptor signalling—From tongues to lungs. Acta Physiol. 2012, 204, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Sternini, C.; Anselmi, L.; Rozengurt, E. Enteroendocrine cells: A site of ‘taste’ in gastrointestinal chemosensing. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Malki, A.; Fiedler, J.; Fricke, K.; Ballweg, I.; Pfaffl, M.W.; Krautwurst, D. Class I odorant receptors, TAS1R and TAS2R taste receptors, are markers for subpopulations of circulating leukocytes. J. Leukoc. Biol. 2015, 97, 533–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizzano, M.; Gulbransen, B.D.; Vandenbeuch, A.; Clapp, T.R.; Herman, J.P.; Sibhatu, H.M.; Churchill, M.E.; Silver, W.L.; Kinnamon, S.C.; Finger, T.E. Nasal chemosensory cells use bitter taste signaling to detect irritants and bacterial signals. Proc. Natl. Acad. Sci. USA 2010, 107, 3210–3215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Invest. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Mueller, K.L.; Cook, B.; Wu, D.; Zuker, C.S.; Ryba, N.J. Coding of sweet, bitter, and umami tastes: Different receptor cells sharing similar signaling pathways. Cell 2003, 112, 293–301. [Google Scholar] [CrossRef]
- Iwata, S.; Yoshida, R.; Ninomiya, Y. Taste transductions in taste receptor cells: Basic tastes and moreover. Curr. Pharm. Des. 2014, 20, 2684–2692. [Google Scholar] [CrossRef] [PubMed]
- Margolskee, R.F. Molecular mechanisms of bitter and sweet taste transduction. J. Biol. Chem. 2002, 277, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Treesukosol, Y.; Smith, K.R.; Spector, A.C. The functional role of the T1R family of receptors in sweet taste and feeding. Physiol. Behav. 2011, 105, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, E.; Hoon, M.A.; Mueller, K.L.; Chandrashekar, J.; Ryba, N.J.; Zuker, C.S. A novel family of mammalian taste receptors. Cell 2000, 100, 693–702. [Google Scholar] [CrossRef]
- Brockhoff, A.; Behrens, M.; Massarotti, A.; Appendino, G.; Meyerhof, W. Broad tuning of the human bitter taste receptor hTAS2R46 to various sesquiterpene lactones, clerodane and labdane diterpenoids, strychnine, and denatonium. J. Agric. Food Chem. 2007, 55, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; Zuker, C.S.; Ryba, N.J. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef]
- Hayes, J.E.; Wallace, M.R.; Knopik, V.S.; Herbstman, D.M.; Bartoshuk, L.M.; Duffy, V.B. Allelic variation in TAS2R bitter receptor genes associates with variation in sensations from and ingestive behaviors toward common bitter beverages in adults. Chem. Senses 2011, 36, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.J.; Christensen, M.; Finger, T.E.; Tizzano, M. Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation. Proc. Natl. Acad. Sci. USA 2014, 111, 6075–6080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulbransen, B.; Silver, W.; Finger, T.E. Solitary chemoreceptor cell survival is independent of intact trigeminal innervation. J. Comp. Neurol. 2008, 508, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannucci, D.R.; Groblewski, G.E.; Sneyd, J.; Yule, D.I. Targeted phosphorylation of inositol 1,4,5-trisphosphate receptors selectively inhibits localized Ca2+ release and shapes oscillatory Ca2+ signals. J. Biol. Chem. 2000, 275, 33704–33711. [Google Scholar] [CrossRef] [PubMed]
- Taruno, A.; Matsumoto, I.; Ma, Z.; Marambaud, P.; Foskett, J.K. How do taste cells lacking synapses mediate neurotransmission? CALHM1, a voltage-gated ATP channel. Bioessays 2013, 35, 1111–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhao, Z.; Margolskee, R.; Liman, E. The transduction channel TRPM5 is gated by intracellular calcium in taste cells. J. Neurosci. 2007, 27, 5777–5786. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, M.A.; Abe, K.; Emori, Y. IP(3) receptor type 3 and PLCβ2 are co-expressed with taste receptors T1R and T2R in rat taste bud cells. Chem. Senses 2001, 26, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Taruno, A.; Vingtdeux, V.; Ohmoto, M.; Ma, Z.; Dvoryanchikov, G.; Li, A.; Adrien, L.; Zhao, H.; Leung, S.; Abernethy, M.; et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes. Nature 2013, 495, 223–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, M.; Born, S.; Redel, U.; Voigt, N.; Schuh, V.; Raguse, J.D.; Meyerhof, W. Immunohistochemical detection of TAS2R38 protein in human taste cells. PLoS ONE 2012, 7, e40304. [Google Scholar] [CrossRef] [PubMed]
- Barham, H.P.; Cooper, S.E.; Anderson, C.B.; Tizzano, M.; Kingdom, T.T.; Finger, T.E.; Kinnamon, S.C.; Ramakrishnan, V.R. Solitary chemosensory cells and bitter taste receptor signaling in human sinonasal mucosa. Int. Forum Allergy Rhinol. 2013, 3, 450–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizzano, M.; Cristofoletti, M.; Sbarbati, A.; Finger, T.E. Expression of taste receptors in solitary chemosensory cells of rodent airways. BMC Pulm. Med. 2011, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile cilia of human airway epithelia are chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.N.; Koch, G.; Thompson, J.A.; Xavier, K.B.; Cool, R.H.; Quax, W.J. The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2012, 76, 46–65. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Nair, S.K. Quorum sensing: How bacteria can coordinate activity and synchronize their response to external signals? Protein Sci. 2012, 21, 1403–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsek, M.R.; Greenberg, E.P. Acyl-homoserine lactone quorum sensing in gram-negative bacteria: A signaling mechanism involved in associations with higher organisms. Proc. Natl. Acad. Sci. USA 2000, 97, 8789–8793. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.A.; Underhill, D.M.; Sweet, M.J.; Ozinsky, A.O.; Liew, F.Y.; Aderem, A. Macrophages exposed continuously to lipopolysaccharide and other agonists that act via toll-like receptors exhibit a sustained and additive activation state. BMC Immunol. 2001, 2, 11. [Google Scholar] [CrossRef]
- Carey, R.M.; Workman, A.D.; Chen, B.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Lee, R.J.; Cohen, N.A. Staphylococcus aureus triggers nitric oxide production in human upper airway epithelium. Int. Forum Allergy Rhinol. 2015, 5, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Workman, A.D.; Yan, C.H.; Chen, B.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Lee, R.J.; Cohen, N.A. Sinonasal T2R-mediated nitric oxide production in response to Bacillus cereus. Am. J. Rhinol. Allergy 2017, 31, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Barraud, N.; Hassett, D.J.; Hwang, S.H.; Rice, S.A.; Kjelleberg, S.; Webb, J.S. Involvement of nitric oxide in biofilm dispersal of Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 7344–7353. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Carey, R.M.; Kohanski, M.A.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Cohen, N.A. Relative susceptibility of airway organisms to antimicrobial effects of nitric oxide. Int. Forum Allergy Rhinol. 2017, 7, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Salathe, M. Regulation of mammalian ciliary beating. Annu. Rev. Physiol. 2007, 69, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Sleigh, M.A.; Blake, J.R.; Liron, N. The propulsion of mucus by cilia. Am. Rev. Respir. Dis. 1988, 137, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.; Prince, A. Innate immunity in the respiratory epithelium. Am. J. Respir. Cell Mol. Biol. 2011, 45, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.K.; Drayna, D. Genetics of individual differences in bitter taste perception: Lessons from the PTC gene. Clin. Genet. 2005, 67, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Zhang, Z.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Scott, T.; Zhao, N.W.; Owens, D.; et al. The bitter taste receptor T2R38 is an independent risk factor for chronic rhinosinusitis requiring sinus surgery. Int. Forum Allergy Rhinol. 2014, 4, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Howland, T.J.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Lee, R.J.; Cohen, N.A. Genetics of the taste receptor T2R38 correlates with chronic rhinosinusitis necessitating surgical intervention. Int. Forum Allergy Rhinol. 2013, 3, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Mfuna Endam, L.; Filali-Mouhim, A.; Boisvert, P.; Boulet, L.P.; Bosse, Y.; Desrosiers, M. Genetic variations in taste receptors are associated with chronic rhinosinusitis: A replication study. Int. Forum Allergy Rhinol. 2014, 4, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Workman, A.D.; Hadjiliadis, D.; Dorgan, D.J.; Frame, D.; Brooks, S.; Doghramji, L.; Palmer, J.N.; Mansfield, C.; Reed, D.R.; et al. T2R38 genotype is correlated with sinonasal quality of life in homozygous ΔF508 cystic fibrosis patients. Int. Forum Allergy Rhinol. 2016, 6, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.G.; Oh, S.J.; Won, T.B.; Kim, Y.M.; Shim, W.S.; Rhee, C.S.; Min, J.Y.; Dhong, H.J. Effects of staphylococcal enterotoxin on ciliary activity and histology of the sinus mucosa. Acta Otolaryngol. 2006, 126, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Shaari, J.; Claire, S.E.; Palmer, J.N.; Chiu, A.G.; Kennedy, D.W.; Cohen, N.A. Altered sinonasal ciliary dynamics in chronic rhinosinusitis. Am. J. Rhinol. 2006, 20, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Naraghi, M.; Deroee, A.F.; Ebrahimkhani, M.; Kiani, S.; Dehpour, A. Nitric oxide: A new concept in chronic sinusitis pathogenesis. Am. J. Otolaryngol. 2007, 28, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.H.; Hahn, S.; McMahon, D.; Bonislawski, D.; Kennedy, D.W.; Adappa, N.D.; Palmer, J.N.; Jiang, P.; Lee, R.J.; Cohen, N.A. Nitric oxide production is stimulated by bitter taste receptors ubiquitously expressed in the sinonasal cavity. Am. J. Rhinol. Allergy 2017, 31, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, J.D.; Chakraborty, R.; Shaik, F.A.; Jaggupilli, A.; Bhullar, R.P.; Chelikani, P. The Pharmacochaperone Activity of Quinine on Bitter Taste Receptors. PLoS ONE 2016, 11, e0156347. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Maina, I.W.; Brooks, S.G.; Kohanski, M.A.; Cowart, B.J.; Mansfield, C.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Reed, D.R.; et al. The Role of Quinine-Responsive Taste Receptor Family 2 in Airway Immune Defense and Chronic Rhinosinusitis. Front. Immunol. 2018, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.R.; Zhu, G.; Breslin, P.A.; Duke, F.F.; Henders, A.K.; Campbell, M.J.; Montgomery, G.W.; Medland, S.E.; Martin, N.G.; Wright, M.J. The perception of quinine taste intensity is associated with common genetic variants in a bitter receptor cluster on chromosome 12. Hum. Mol. Genet. 2010, 19, 4278–4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledda, M.; Kutalik, Z.; Souza Destito, M.C.; Souza, M.M.; Cirillo, C.A.; Zamboni, A.; Martin, N.; Morya, E.; Sameshima, K.; Beckmann, J.S.; et al. GWAS of human bitter taste perception identifies new loci and reveals additional complexity of bitter taste genetics. Hum. Mol. Genet. 2014, 23, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Zancanaro, C.; Caretta, C.M.; Merigo, F.; Cavaggioni, A.; Osculati, F. α-Gustducin expression in the vomeronasal organ of the mouse. Eur. J. Neurosci. 1999, 11, 4473–4475. [Google Scholar] [CrossRef] [PubMed]
- Osculati, F.; Bentivoglio, M.; Castellucci, M.; Cinti, S.; Zancanaro, C.; Sbarbati, A. The solitary chemosensory cells and the diffuse chemosensory system of the airway. Eur. J. Histochem. 2007, 51, 65–72. [Google Scholar] [PubMed]
- Lin, W.; Ezekwe, E.A.; Zhao, Z., Jr.; Liman, E.R.; Restrepo, D. TRPM5-expressing microvillous cells in the main olfactory epithelium. BMC Neurosci. 2008, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Hariri, B.M.; McMahon, D.B.; Chen, B.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Jiang, P.; Margolskee, R.F.; et al. Bacterial d-amino acids suppress sinonasal innate immunity through sweet taste receptors in solitary chemosensory cells. Sci. Signal. 2017, 10, eaam7703. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Tang, Y.Q.; Morris, W.L.; McGuire, P.A.; Novotny, M.J.; Smith, W.; Henschen, A.H.; Cullor, J.S. Purification, primary structures, and antibacterial activities of β-defensins, a new family of antimicrobial peptides from bovine neutrophils. J. Biol. Chem. 1993, 268, 6641–6648. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Mack, B.; Kramer, M.F. Solitary chemosensory cells in the respiratory and vomeronasal epithelium of the human nose: A pilot study. Rhinology 2011, 49, 507–512. [Google Scholar] [PubMed]
- Kalsi, K.K.; Baker, E.H.; Fraser, O.; Chung, Y.L.; Mace, O.J.; Tarelli, E.; Philips, B.J.; Baines, D.L. Glucose homeostasis across human airway epithelial cell monolayers: Role of diffusion, transport and metabolism. Pflugers Arch. 2009, 457, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Lemon, C.H.; Margolskee, R.F. Contribution of the T1R3 taste receptor to the response properties of central gustatory neurons. J. Neurophysiol. 2009, 101, 2459–2471. [Google Scholar] [CrossRef] [PubMed]
- Bachmanov, A.A.; Bosak, N.P.; Lin, C.; Matsumoto, I.; Ohmoto, M.; Reed, D.R.; Nelson, T.M. Genetics of taste receptors. Curr. Pharm. Des. 2014, 20, 2669–2683. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Brooks, S.G.; Kohanski, M.A.; Blasetti, M.T.; Cowart, B.J.; Mansfield, C.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Reed, D.R.; et al. Bitter and sweet taste tests are reflective of disease status in chronic rhinosinusitis. J. Allergy Clin. Immunol. Pract. 2017, 6, 1078–1080. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Truesdale, C.M.; Workman, A.D.; Doghramji, L.; Mansfield, C.; Kennedy, D.W.; Palmer, J.N.; Cowart, B.J.; Cohen, N.A. Correlation of T2R38 taste phenotype and in vitro biofilm formation from nonpolypoid chronic rhinosinusitis patients. Int. Forum Allergy Rhinol. 2016, 6, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, N.A.; Widelitz, J.S.; Chiu, A.G.; Palmer, J.N.; Kennedy, D.W. Familial aggregation of sinonasal polyps correlates with severity of disease. Otolaryngol. Head Neck Surg. 2006, 134, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Cui, M.; Zhao, B.; Liu, Z.; Snyder, L.A.; Benard, L.M.; Osman, R.; Margolskee, R.F.; Max, M. Lactisole interacts with the transmembrane domains of human T1R3 to inhibit sweet taste. J. Biol. Chem. 2005, 280, 15238–15246. [Google Scholar] [CrossRef] [PubMed]
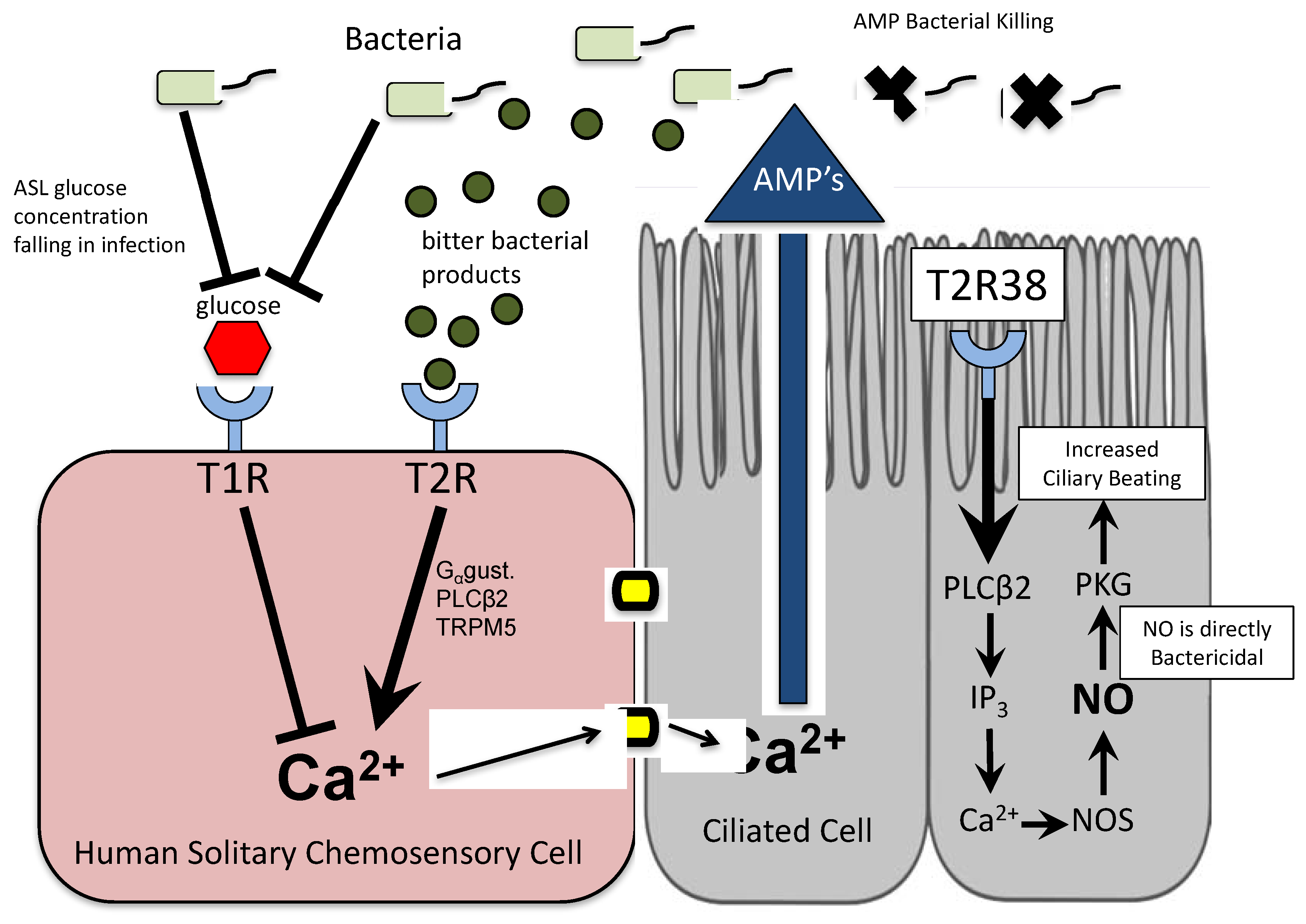
Figure 1.
Bitter and sweet taste receptor-induced innate immune defenses in ciliated and solitary chemosensory cells in the human upper airway. ASL: airway surface liquid; AMP: anti-microbial peptide; NOS: nitric oxide synthase; NO: nitric oxide; PKG: protein kinase G.
Figure 1.
Bitter and sweet taste receptor-induced innate immune defenses in ciliated and solitary chemosensory cells in the human upper airway. ASL: airway surface liquid; AMP: anti-microbial peptide; NOS: nitric oxide synthase; NO: nitric oxide; PKG: protein kinase G.


{kind=link}
Table 1.
Subjective taste intensity ratings in patients with chronic rhinosinusitis (CRS) without nasal polyps (CRSsNP) and with nasal polyps (CRSwNP), relative to control patient taste intensity ratings.
Table 1.
Subjective taste intensity ratings in patients with chronic rhinosinusitis (CRS) without nasal polyps (CRSsNP) and with nasal polyps (CRSwNP), relative to control patient taste intensity ratings.
| Bitter Perception | Sweet Perception | Salt Perception | ||
|---|---|---|---|---|
| Quinine | Denatonium | Sucrose | NaCl | |
| CRSsNP | No difference | Decreased | Increased | No difference |
| CRSwNP | Decreased | No difference | Increased | No difference |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Workman, A.D.; Patel, N.N.; Carey, R.M.; Kuan, E.C.; Cohen, N.A. The Role of Taste Receptors in Airway Innate Immune Defense. Sinusitis 2018, 3, 6. https://0-doi-org.brum.beds.ac.uk/10.3390/sinusitis3020006
AMA Style
Workman AD, Patel NN, Carey RM, Kuan EC, Cohen NA. The Role of Taste Receptors in Airway Innate Immune Defense. Sinusitis. 2018; 3(2):6. https://0-doi-org.brum.beds.ac.uk/10.3390/sinusitis3020006
Chicago/Turabian StyleWorkman, Alan D., Neil N. Patel, Ryan M. Carey, Edward C. Kuan, and Noam A. Cohen. 2018. "The Role of Taste Receptors in Airway Innate Immune Defense" Sinusitis 3, no. 2: 6. https://0-doi-org.brum.beds.ac.uk/10.3390/sinusitis3020006