Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation


Pathology, New York Presbyterian - Brooklyn Methodist Hospital, 506 6th St., Brooklyn, NY 11215, USA
Reprod. Med. 2020, 1(1), 17-31; https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed1010003
Submission received: 16 March 2020
/
Revised: 24 April 2020
/
Accepted: 29 April 2020
/
Published: 30 April 2020
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Background: Wilm’s tumor 1 gene (WT1) is a transcription factor with versatile cellular functions in embryonic development, the maintenance of adult tissue functions, and regeneration. WT1 is known to be regulated by progesterone and it is abundantly expressed in endometrium, but its function is unclear. Design: in this observational and descriptive study, WT1 expression was detected by immunohistochemical staining in endometrium of various physiological and pathological conditions. Result: WT1 was detected in endometrial stromal cells and vascular smooth muscle cells, in both proliferative and secretory phases of menstrual cycles. WT1 appeared increased in vascular smooth muscle cells in spiral artery in early pregnancy and it was also detected in regenerative endothelial cells and smooth muscle cells in decidual vasculopathy at term. WT1 expression appeared decreased in endometrial stromal cells in adenomyosis (endometriosis). Conclusion: WT1 potentially links the hormonal effects on endometrial decidualization and may play a role in gestational vascular transformation during pregnancy and restoration after pregnancy.
1. Introduction
Endometrial decidualization is characterized by the morphological transformation of epithelium, endometrial stromal tissue and terminal segments of the spiral artery, through the concerted action of steroid hormones (estrogen and progesterone), secreted by the ovary and subsequently corpus luteum after ovulation [1,2]. Embryonic implantation occurs in a narrow window of time after fertilization and the fertilized embryo attaches and invades into endometrial tissue, through complex interactions between the developing trophectoderm and the decidualized endometrium [2]. Extravillous trophoblasts derived from the early trophectoderm invade the decidualized endometrial stromal tissue and the spiral artery, leading to trophoblasts-dependent spiral artery remodeling, by forming the endovascular trophoblastic “plugs” with concurrent replacement of smooth muscle wall and endothelium [3]. Uterine natural killer cells (NK) play critical roles in trophoblasts dependent spiral artery remodeling and the formation of endovascular trophoblastic “plugs” is associated with phenotypic switch of the endovascular trophoblasts to express CD56, a defining molecular marker for NK cell lineage [4,5,6,7,8]. Meanwhile, endometrial spiral artery is undergoing trophoblasts independent remodeling morphologically characterized by arterial wall hypertrophy/hyperplasia (mural hypertrophy/hyperplasia) and endothelial vacuolization, and this morphological transformation occurs at a distance from the extravillous trophoblasts, without direct trophoblastic interaction [9]. The molecular mechanism of trophoblasts independent remodeling of spiral artery (mural hypertrophy/hyperplasia) is likely through endocrine (hormonal) actions, but the precise hormonal action on the smooth muscle cells of the vascular wall is poorly understood. During endometrial decidualization and subsequent embryonic/blastocyst implantation, estrogen, progesterone and human chorionic gonadotrophin (hCG) play critical roles in decidual, trophoblastic and fetal development, and the maintenance of pregnancy in the first trimester. However, the role of these steroid and peptide hormones on spiral artery remodeling is yet to be defined [2].
Wilm’s tumor 1 gene (WT1) is a tumor suppressor gene discovered in patients of Wilm’s tumor, a high grade pediatric malignancy with versatile molecular functions [10,11,12,13,14,15]. WT1 is a transcription factor regulating a large number of target gene expressions, mediating a variety of cellular functions in organ development, homeostasis and diseases [16,17]. Mutations of WT1 gene were shown to be associated with Denys–Drash syndrome and Frasier syndrome, and deletion of the short arm of the chromosome 11 containing WT1 gene is associated with WAGR syndrome (Wilm’s tumor, anirida, genitourinary anomalies, and mental retardation) [15,18,19]. WT1 is interacting with many different cellular partner proteins important for various cellular activities [10,15]. Normal WT1 gene function is critical for the development of urogenital system, mesothelial cell function and vascular endothelial cell functions [15]. Clinically, WT1 gene is also important for adult kidney function and cardiovascular functions [15,20]. In pathology practice, immunohistochemical staining for WT1 serves as a marker for ovarian carcinoma, mesothelial cell and endothelial cell lineages [21,22]. There is abundant WT1 gene expression in the endometrium by RNA microarray studies (human protein atlas https://www.proteinatlas.org/), but the function of WT1 in endometrium is yet to be defined, especially when the endometrium undergoes significant changes during the menstrual cycle and pregnancy. As initial steps, expression of WT1 protein in endometrium by immunohistochemical staining was attempted to locate the WT1 protein expression during menstrual cycles and pregnancy. These results will provide the global view of WT1 gene expression in endometrium during the changes of menstrual cycle and pregnancy, with particular emphasis on maternal vascular remodeling during pregnancy and vascular restoration after pregnancy.
2. Material and Methods
This study is exempt from Institutional Review Board (IRB) approval according to section 46.101(b) of 45CFR 46, which states that research involving the study of existing pathological and diagnostic specimens in such a manner that subjects cannot be identified is exempt from the Department of Health and Human Services Protection of Human Research Subjects. Endometrial and placental tissues submitted for pathology examination for a variety of clinical indications were included in the study using the routine hematoxylin and eosin (H+E) stain. Proliferative endometrium, secretory endometrium, decidua and implantation sites were obtained from the submitted pathology specimens during clinical practice. Second trimester placentas were obtained from inevitable abortion specimens due to chorioamnionitis. Paraffin-embedded tissues from routine surgical pathology specimens and routine H+E stained pathology slides were examined by light microscopy using the Amsterdam criteria for placental examination [23]. Immunohistochemical staining for WT1 was used to highlight the specific cell types of the muscular vessels, endometrium and myometrium, fetal villous stem villous arteries and umbilical cord vessels. CD34 was also used to highlight the endothelium of the endometrial or decidual vessels in comparison. Immunohistochemical staining procedures were performed in the routine clinical pathologic application on paraffin-embedded tissues, using the Leica Biosystems Bond III automated immunostaining system, following the manufacturer’s instructions. No special procedures were employed for the study. WT1, CD34 and CD56 monoclonal antibodies were purchased for clinical applications from Dako Agilent, under the catalogue number M3561, M7165 and M730401-2. Monoclonal antibody against smooth muscle myosin heavy chain (SMYOHC) was from Cell Marque (Sigma-Aldrich, St. Louis, MO, USA, under the catalogue number 298M-16.
In total, 3 cases of proliferative endometrium, 3 cases of secretory endometrium, 2 cases of therapeutic abortion specimens at 10 weeks’ gestation, 2 case of inevitable abortion at 17 weeks, 8 cases of term placentas and umbilical cords at 39–40 weeks, with and without decidual vasculopathy, were examined by light microscopy and H+E stain, as well as immunostaining for WT1, CD34, CD56 and SMYOHC. The immunostaining results were presented as qualitative measures of WT1 gene expression, and the methods were not calibrated for the direct quantitative measurement of WT1 protein expression.
3. Results
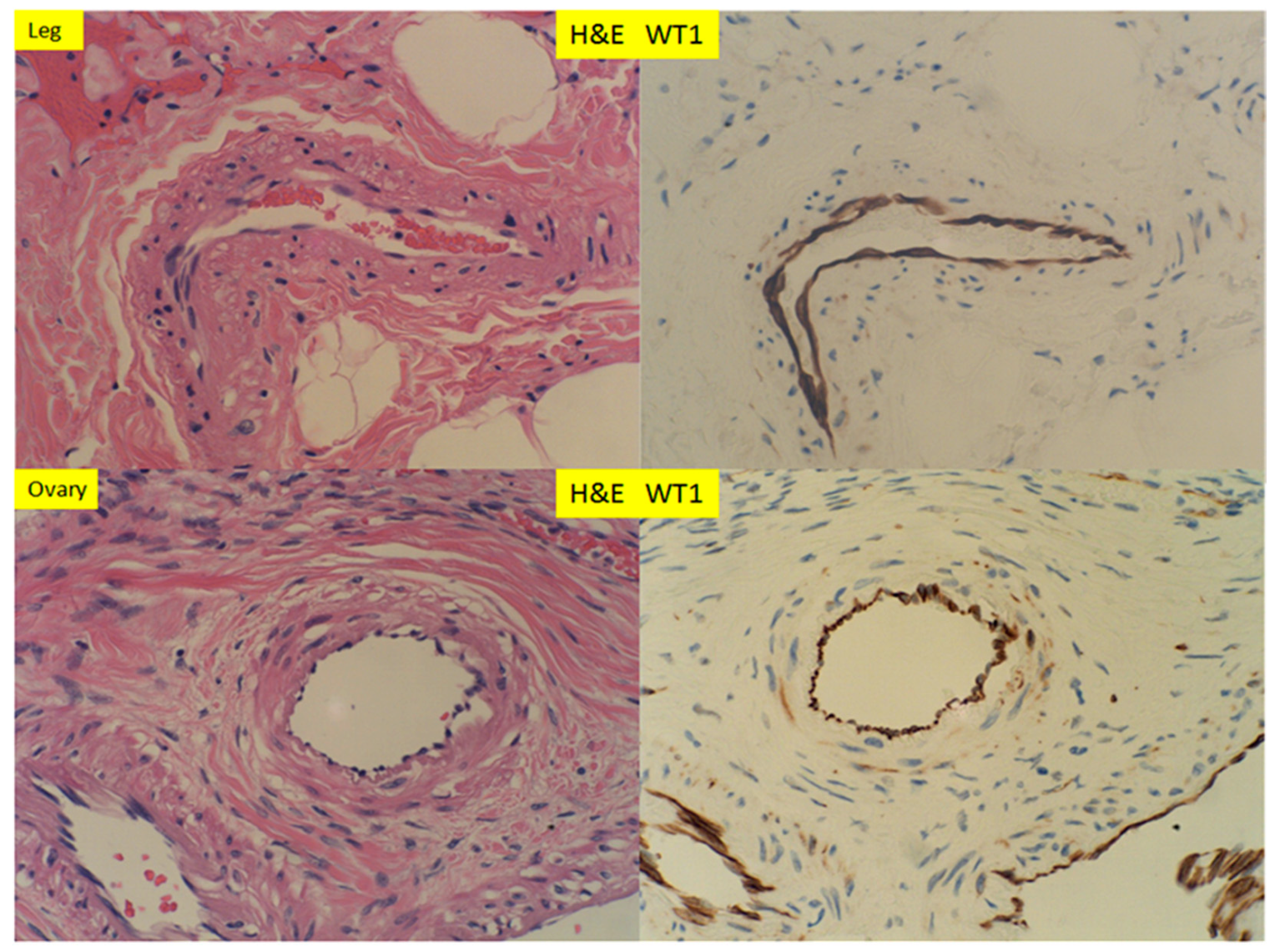
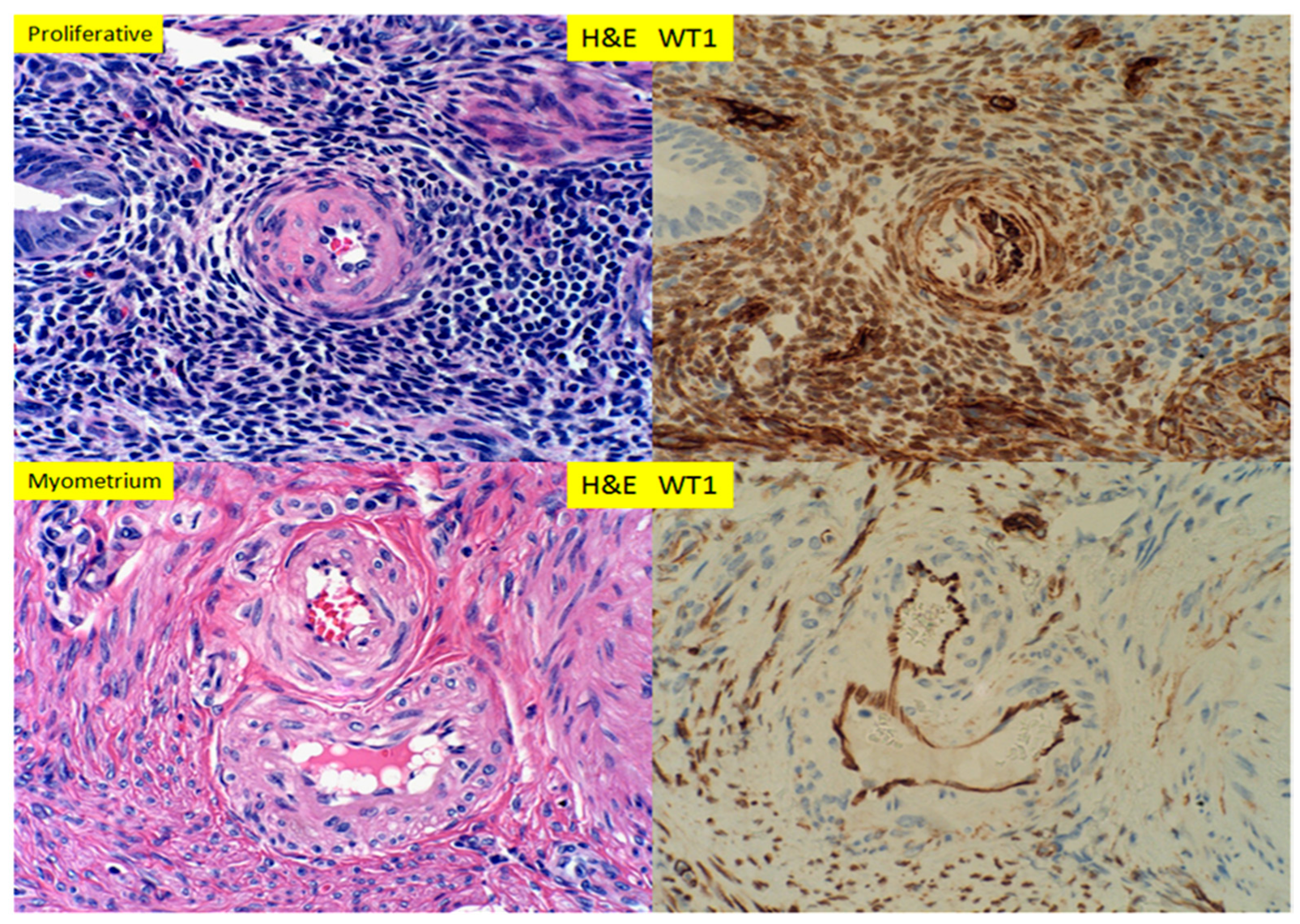
In small muscular arteries or arterioles of the lower extremities and the ovaries, WT1 was detected by immunohistochemical staining, within the endothelial cells only. The staining signals appeared to be both cytoplasmic and nuclear. The smooth muscle layers of the vascular walls showed no or minimal immunostaining signals (Figure 1). The adventitia layer of the artery was evident on H+E staining slides, but no WT1 signals were present within the adventitia. In proliferative endometrium, WT1 expression was present in the nuclei of stromal cells and the endothelial cells, as well as the smooth muscle cells of the spiral artery (Figure 2, top panel). The endometrial glands, epithelium and lymphocytes showed no staining signals. In myometrium, the endothelial cells of the artery showed WT1 staining signals, as well as the entire myometrium, but no smooth muscle cells of muscular artery were staining for WT1 (Figure 2 bottom panel). The adventitia layer was not evident in the endometrial arteries, and no convincing staining signals for WT1 were noted for the adventitial layers of myometrial arteries. In contrast to those of the proliferative endometrium, the endometrial stromal cells in adenomyosis (endometriosis) showed no nuclear staining signals for WT1 (Figure 3, bottom panel). In secretory endometrium, WT1 expression was identified in the stromal cells, endothelial cells and smooth muscle cells of the spiral artery walls (Figure 4, top panel), similar to those of proliferative endometrium (Figure 2). There appeared to be stronger staining signals for WT1 in smooth muscle layers of spiral artery, but the immunohistochemical staining method was not calibrated for the quantitative measurement of WT1 expression. WT1 was seen in the endothelial cells of the myometrial segment of the spiral artery, and no significant WT1 signals were present in the smooth muscle layers or adventitial layers (Figure 4, bottom panel).
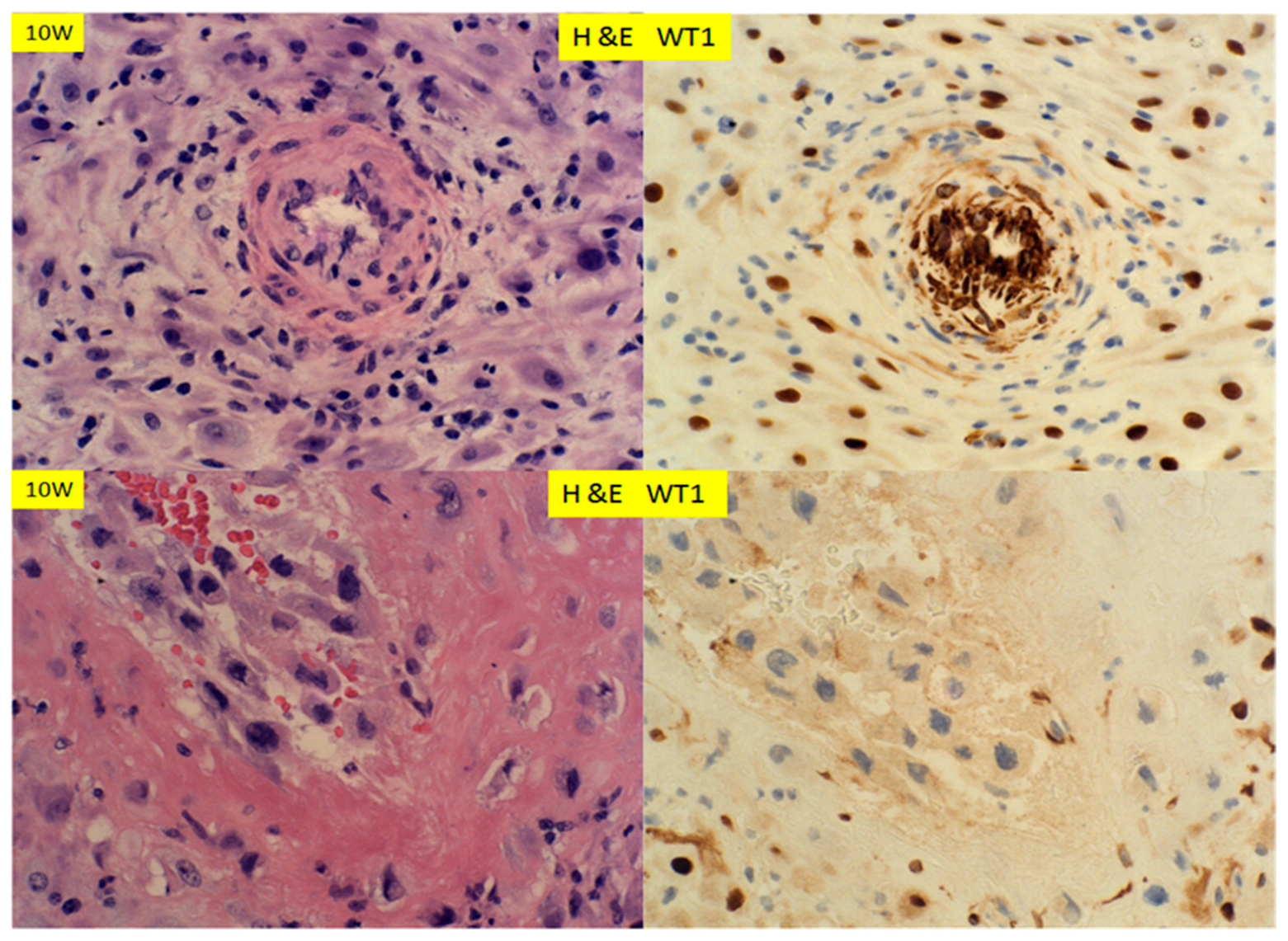
In early pregnancy, at 10 weeks, when the spiral arteries were undergoing remodeling in both the trophoblasts dependent and independent manners, the decidual cells expressed WT1 strongly within the nuclei (Figure 5). There were strong nuclear signals for WT1 in smooth muscle walls of the spiral artery, which showed mural hypertrophy/hyperplasia (Figure 5, top panel). There appeared to be cytoplasmic WT1 signals within the endothelial cells in the arterial lumen. In contrast, the spiral arteries with trophoblastic “plugs” and the fibrinoid replacement of muscular walls were negative for WT1 expression (Figure 5, bottom panel). At 17 weeks’ gestation (Figure 6), the expression patterns of WT1 in decidua, spiral artery with hypertrophy/hyperplasia and those with trophoblastic plugs were similar to those at 10 weeks’ gestation (Figure 5). There was no difference in WT1 expression pattern and intensity between those of 10 weeks’ and 17 weeks’ gestation. No WT1 staining signals were noted in the adventitia layers of vessels, at either 10 weeks’ or 17 weeks’ gestation.
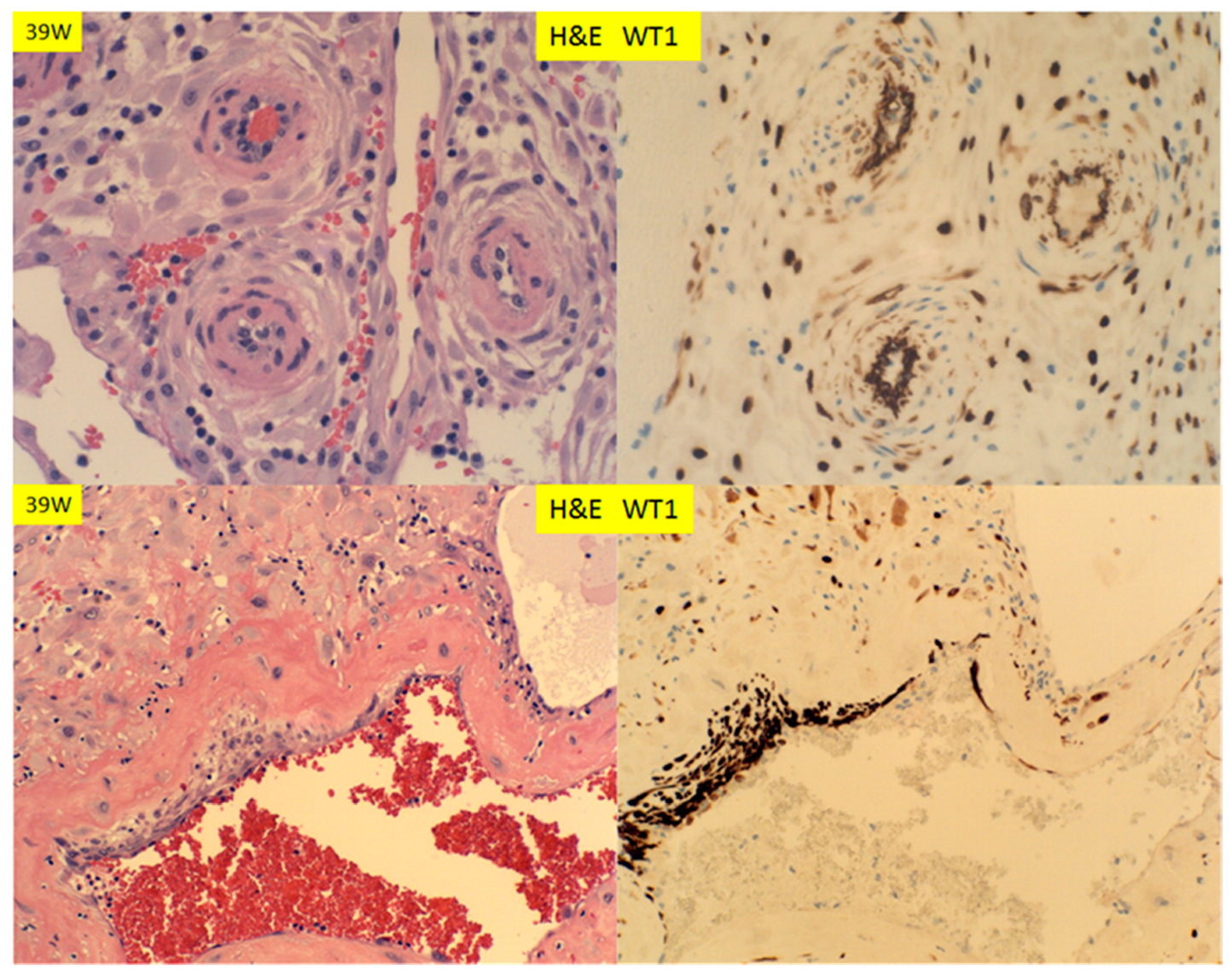
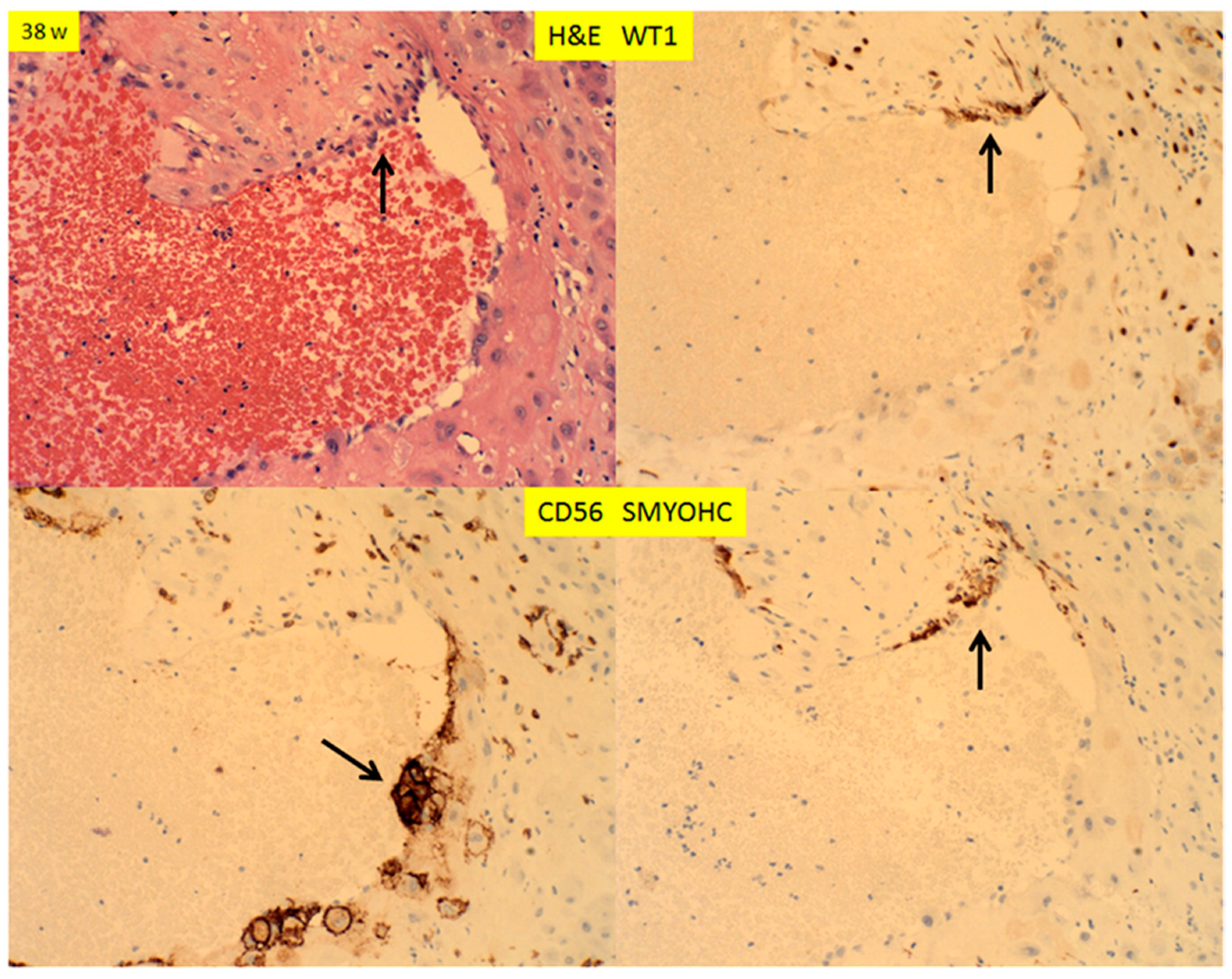
There are two types of decidual vasculopathy; mural arterial hypertrophy/hyperplasia and classic type, including atherosis and fibrinoid medial necrosis. Mural hypertrophy/hyperplasia in the membrane roll (decidual vera) showed WT1 expression in endothelial cells and smooth muscle cells (Figure 7 top panel), and it appeared that the expression in the smooth muscle cells was somewhat decreased in comparison to those in early pregnancy (Figure 5; Figure 6). There was no WT1 expression in classic decidual vasculopathy, including atherosis and fibrinoid medial necrosis. However, WT1 was detected in the regenerating endothelial cells and smooth muscle cells of the arterial walls (Figure 7, bottom panel). The fibrinoid medial necrosis was partially present in the same vessel, with regenerating endothelium and smooth muscle wall, and the fibrinoid medial necrosis was negative for WT1 expression (Figure 7, bottom panel). To illustrate this point of regeneration further, the endovascular trophoblasts can be identified by immunostaining for CD56, and the regenerating smooth muscle cells by smooth muscle myosin heavy chain (SMYOHC), as well as WT1 expression (Figure 8).
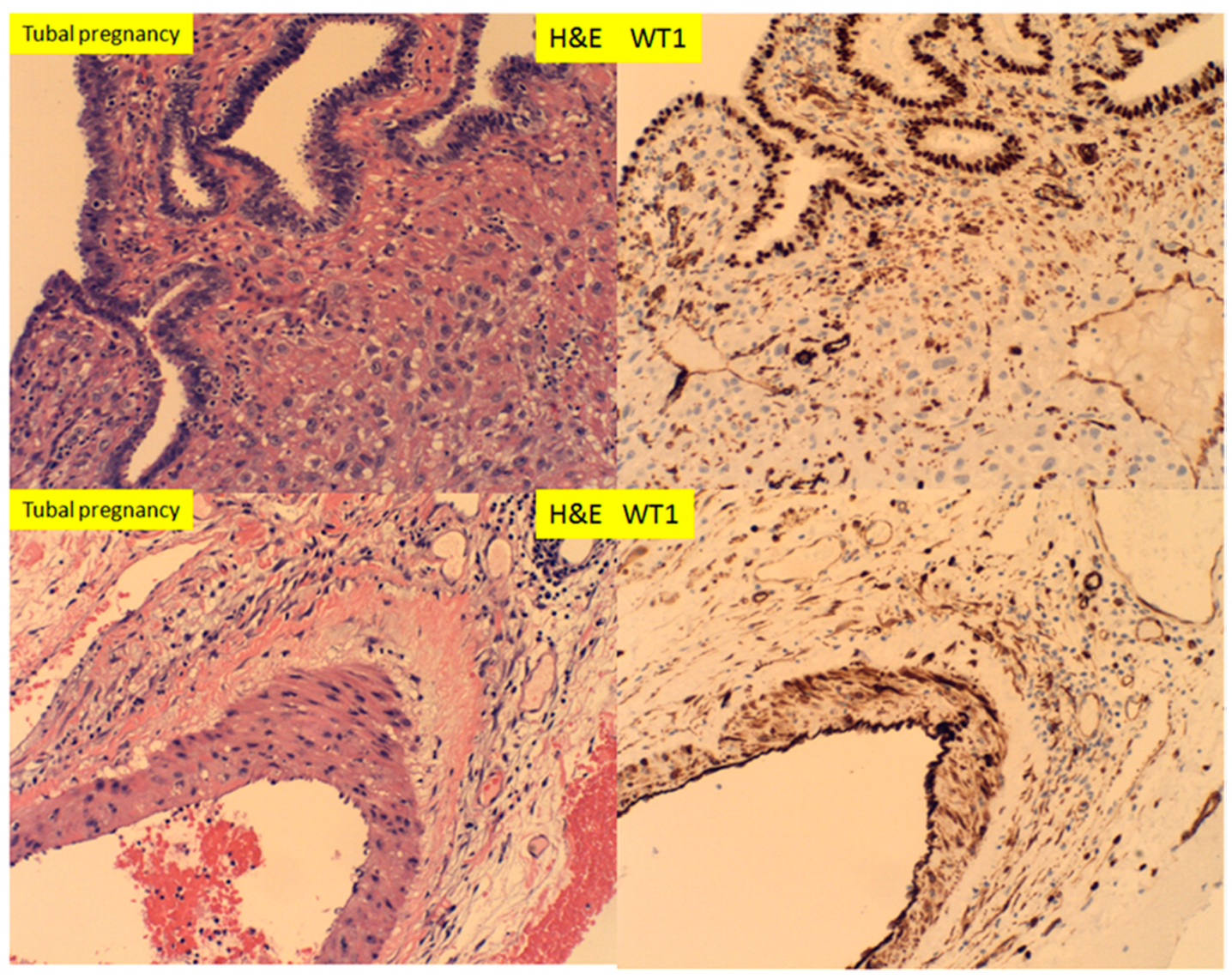
In tubal pregnancy, WT1 expression was noted to be abundant in the tubal epithelium, smooth muscle wall and endothelium of the vessels, as well as small capillary endothelial cells (Figure 9). This is in contrast to the endometrium, where WT1 was only expressed in the endometrial stromal cells, not in the endometrial glandular epithelium.
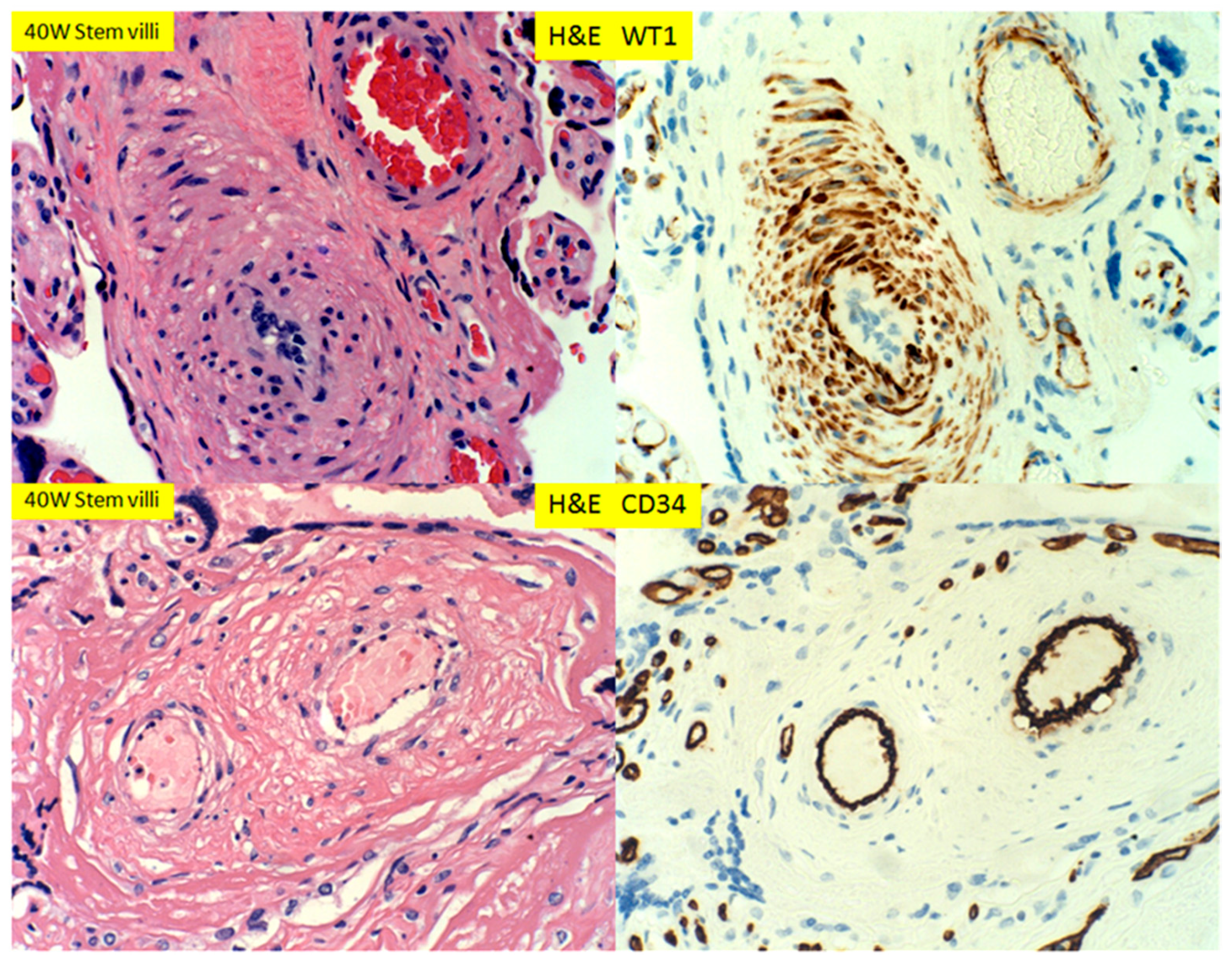
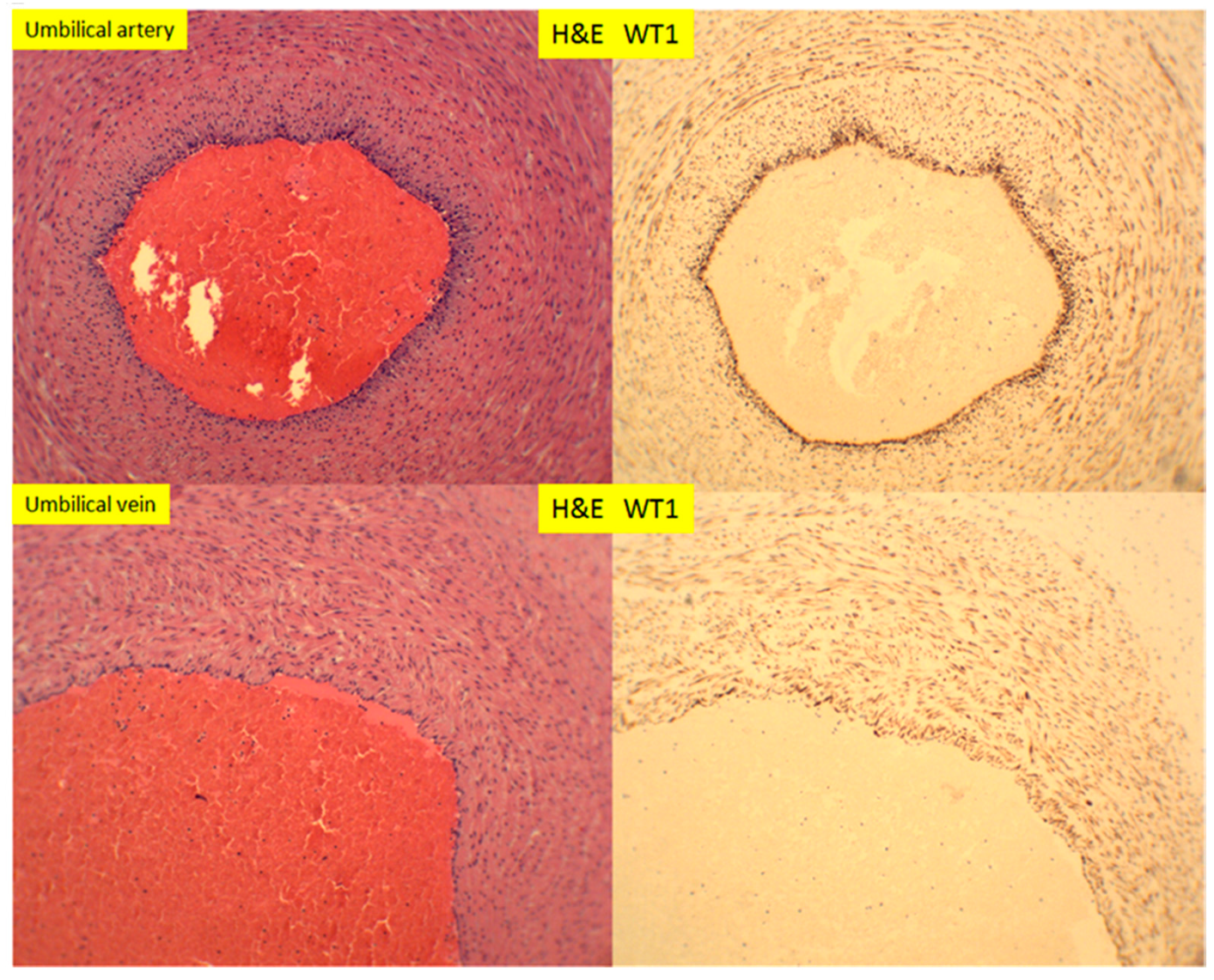
In placental stem villi at term, the muscular artery showed intense WT1 staining signals, but in the luminal endothelial cells, there appeared to be no staining (Figure 10, top panel). This WT1 staining pattern in stem villous artery was in direct contrast to CD34 staining signals, in which only the luminal endothelial cells were positive (Figure 10, top and bottom panels). The WT1 expression patterns in stem villous artery were similar to those in the umbilical artery and veins in umbilical cord at term (Figure 11). There were no adventitia layers of these vessels, as these vessels are anatomically different from those in the systemic circulation [24].
4. Discussion
WT1 gene expression has been studied extensively in a variety of tissues and organs and many aspects of WT1 gene functions are known (Figure 12). WT1 is important for organogenesis of urogenital system, endothelial cell system and mesothelial cell system during embryogenesis, and its versatile cellular function is likely to be conferred through not only regulating multiple target gene expression, but also receiving signals from multiple factors in growth and development [10,15]. Our interest in WT1 gene expression is centered on the role of WT1 gene in decidual vasculopathy of pregnancy, in relation to preeclampsia and other pregnancy related complications. The WT1 function in endometrium appeared at least in part to be through regulation by the steroid hormones (estrogen and progesterone) and insulin-like growth factors (IGF), which are known to be critical for decidualization, implantation and fetal development [25,26,27]. WT1 has been shown to be up-regulated by progesterone in in vitro models, and it can also be self-regulated [25,28]. Our current WT1 expression data in endometrium is consistent with the known function of progesterone in both menstrual cycles and pregnancy. However, the basal expression of WT1 in the stromal cells of proliferative endometrium and the epithelium of the fallopian tubes appears to be unrelated to the progesterone effect, and there is no estrogen or progesterone receptor expression in the tubal epithelium (data not shown). The potential link of WT1 expression in stromal cells to the estrogen function could be through amphiregulin, which was shown to be transcriptionally activated by WT1 [29], although the amphiregulain and estrogen effects was demonstrated in mammary tissue [30]. In the current study, the WT1 expression is abundant in the stromal cells of the secretory endometrium and the decidua in first and second trimesters, as well as the smooth muscle wall of the spiral arteries. The spiral artery transformation in secretory phase of endometrium and remodeling in early pregnancy correlates with the expression of WT1 in the smooth muscle wall, as well as in the stromal cells (decidua cells). There is no WT1 expression in villous trophoblasts or extravillous trophoblasts, including the endovascular trophoblasts. There appears no WT1 expression in uterine NK cells or lymphocytes (data not shown), although WT1 functions are known to be important for myeloid cells and myeloid leukemia [31].
Spiral artery remodeling in early pregnancy is critical for normal pregnancy, and two separate molecular mechanisms are proposed: trophoblasts-dependent or trophoblasts-independent [3,32]. Trophoblasts-dependent remodeling is associated with the direct invasion of extravillous trophoblasts into the spiral artery, with the replacement of the smooth muscle wall and endovascular trophoblasts plugs in the lumen [7]. WT1 gene function is not known to play any role in the process of trophoblastic invasion into the vessels, as there is no WT1 expression in trophoblasts or NK cells. It is unclear if these remodeled arteries contain the adventitia layer, but no WT1 expression was observed in the entire transformed arterial wall. However, the smooth muscle hypertrophy/hyperplasia of trophoblasts-independent spiral artery remodeling appears to correlate with WT1 gene expression in the smooth muscle cells in early pregnancy. Both of the mechanisms result in the narrowing of the vascular lumen, thus limiting the maternal blood flow into the intervillous spaces and embryo, leading to the hypoxic environment which was shown to be critical for fetal development [33]. Furthermore, in the third trimester, after the trophoblasts-dependent smooth muscle remodeling, the spiral artery restores the endothelial layers and the smooth muscle layers within decidua and superficial (inner) segments of myometrium, and the restoration of endothelium and smooth muscle cells appears to be associated with increased WT1 expression (Figure 7 and Figure 8). The presence of endovascular trophoblasts, fibrinoid medial necrosis of spiral artery wall, regenerating endothelial cells and smooth muscle cells within the same vessel, captured the dynamic changes and interactions among these cells at the specific time of gestation. WT1 and WT1 associated protein (WTAP) have been shown to regulate vascular smooth muscle proliferation [34,35]. The restoration of the spiral artery also appears associated with the regression (involution, de-plugging) of endovascular trophoblasts.
The adventitia layer of muscular arteries is equivalent to the serosal layer of visceral organs, and the serosal surface is covered by mesothelium which expresses WT1 [36]. In mice model, WT1 gene deletion resulted in fetal death, due to the abnormal development of vasculature [10,20,36]. In young adult mice, WT1 deletion can lead to multiple organ failures including renal failure, diminished splenic erythropoiesis, bone and fat development as well as abnormal circulating IGF1 and cytokines levels [10,20]. The serosal mesothelial cells served as progenitor cells, not only to the visceral fat tissue, but also to the smooth muscle cells of the vessels such as coronary arteries, and WT1 is critical for maintenance of mesothelial progenitor cells [36,37,38,39]. Based on the current data, the endothelial cells appear important to restoration of smooth muscle wall of the spiral artery after trophoblasts-dependent remodeling, and no adventitia layer or serosal mesothelial layers were observed for the spiral artery of endometrium, placental stem villous arteries or umbilical vessels. WT1 functions in spiral artery remodeling during pregnancy, and restoration after pregnancy requires more in vitro and animal model studies, and this process is likely to be important for further risk of cardiovascular disease after pregnancy.
It is interesting to note that WT1 expression was reduced in adenomyosis (endometriosis). It is known that defective decidualization of the endometrium is related to endometriosis and IGF family members appear important for decidualization [40,41,42]. WT1 regulates the expression of multiple IGFs, IGF binding proteins and IGF receptors, and the WT1 function appears more complex in physiological condition during menstrual cycles and pregnancy [20,43]. It is also interesting to note that WT1 expression is abundant in the tubal epithelium but not in tubal stromal cells, and the fallopian tubes do not undergo decidualization during pregnancy. The fact that tubal pregnancy occurs occasionally raises interesting questions whether decidualization of endometrium is absolutely required for embryonic implantation, and how embryonic implantation occurs in tubal pregnancy in the absence of decidualization. Finally, WT1 is a gene with multiple functions and the upstream factors influencing the WT1 gene function/expression in the regeneration of endothelial cells and smooth muscle cells as well as in endometriosis will likely yield important information, not only about the decidual vasculopathy and preeclampsia, but also the pathogenesis of endometriosis. Searching for these important upstream factors of WT1 gene expression requires establishment of in vitro cellular and animal models, as well as quantitative biochemical studies.
Funding
This research received no external funding.
Acknowledgments
The author is thankful to Missver Jocson for technical assistance.
Conflicts of Interest
The author declares no conflict of interest.
References
- Benirschke, K.; Burton, G.J.; Baergen, R.N. Pathology of the Human Placenta, 6th ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Cunningham, F.G. Williams Obstetrics, 25th ed.; McGraw-Hill: New York, NY, USA, 2018; Volume xvi, p. 1328. [Google Scholar]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Smith, S.D.; Keogh, R.J.; Jones, R.; Baker, P.; Knöfler, M.; Cartwright, J.E.; Whitley, G.S.; Aplin, J. Trophoblast- and vascular smooth muscle cell-derived MMP-12 mediates elastolysis during uterine spiral artery remodeling. Am. J. Pathol. 2010, 177, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Benagiano, M.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Placental bed research: II. Functional and immunological investigations of the placental bed. Am. J. Obstet. Gynecol. 2019, 221, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P. Decidual vasculopathy and spiral artery remodeling revisited II: Relations to trophoblastic dependent and independent vascular transformation. J. Matern. Neonatal Med. 2020, 1–7. [Google Scholar] [CrossRef]
- Zhang, P. Phenotypic Switch of Endovascular Trophoblasts in Decidual Vasculopathy with Implication for Preeclampsia and Other Pregnancy Complications. Fetal Pediatr. Pathol. 2020, 1–20. [Google Scholar] [CrossRef]
- Zhang, P. Decidual Vasculopathy in Preeclampsia and Spiral Artery Remodeling Revisited: Shallow Invasion versus Failure of Involution. Am. J. Perinatol. Rep. 2018, 8, e241–e246. [Google Scholar] [CrossRef] [Green Version]
- Craven, C.M.; Morgan, T.; Ward, K. Decidual spiral artery remodelling begins before cellular interaction with cytotrophoblasts. Placenta 1998, 19, 241–252. [Google Scholar] [CrossRef]
- Hastie, N.D. Wilms’ tumour 1 (WT1) in development, homeostasis and disease. Development 2017, 144, 2862–2872. [Google Scholar] [CrossRef] [Green Version]
- Call, K.M.; Glaser, T.; Ito, C.Y.; Buckler, A.J.; Pelletier, J.; Haber, D.A.; Rose, E.A.; Kral, A.; Yeger, H.; Lewis, W.H.; et al. Isolation and characterization of a zinc finger polypeptide gene at the human chromosome 11 Wilms’ tumor locus. Cell 1990, 60, 509–520. [Google Scholar] [CrossRef]
- Haber, D.A.; Buckler, A.J.; Glaser, T.; Call, K.M.; Pelletier, J.; Sohn, R.L.; Douglass, E.C.; Housman, D.E. An internal deletion within an 11p13 zinc finger gene contributes to the development of Wilms’ tumor. Cell 1990, 61, 1257–1269. [Google Scholar]
- Haber, D.A.; Housman, D.E. The genetics of Wilms’ tumor. Adv. Cancer Res. 1992, 59, 41–68. [Google Scholar]
- Haber, D.A.; Buckler, A.J. WT1: A novel tumor suppressor gene inactivated in Wilms’ tumor. New Biol. 1992, 4, 97–106. [Google Scholar]
- Van den Heuvel-Eibrink, M.M. Wilms Tumor. 2016. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK373360/?term=Wilms%20tumor (accessed on 30 April 2020).
- Buckler, A.J.; Pelletier, J.; Haber, D.A.; Glaser, T.; Housman, D.E. Isolation, characterization, and expression of the murine Wilms’ tumor gene (WT1) during kidney development. Mol. Cell. Boil. 1991, 11, 1707–1712. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.; Schalling, M.; Buckler, A.J.; Rogers, A.; Haber, D.A.; Housman, D. Expression of the Wilms’ tumor gene WT1 in the murine urogenital system. Genes Dev. 1991, 5, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Patek, C.E.; Little, M.H.; Fleming, S.; Miles, C.; Charlieu, J.-P.; Clarke, A.; Miyagawa, K.; Christie, S.; Doig, J.; Harrison, D.J.; et al. A zinc finger truncation of murine WT1 results in the characteristic urogenital abnormalities of Denys-Drash syndrome. Proc. Natl. Acad. Sci. USA 1999, 96, 2931–2936. [Google Scholar] [CrossRef] [Green Version]
- Francke, U.; Holmes, L.; Atkins, L.; Riccardi, V. Aniridia-Wilms’ tumor association: Evidence for specific deletion of 11p13. Cytogenet. Genome Res. 1979, 24, 185–192. [Google Scholar] [CrossRef]
- Chau, Y.-Y.; Brownstein, D.; Mjoseng, H.K.; Lee, W.-C.; Buza-Vidas, N.; Nerlov, C.; Jacobsen, S.E.W.; Perry, P.; Berry, R.; Thornburn, A.; et al. Acute multiple organ failure in adult mice deleted for the developmental regulator WT1. PLoS Genet. 2011, 7, e1002404. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, H.G.; Crum, C.P. The utility of immunohistochemistry in the differential diagnosis of gynecologic disorders. Arch. Pathol. Lab. Med. 2015, 139, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Husain, A.N.; Colby, T.V.; Ordóñez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Khong, T.Y.; Mooney, E.E.; Ariel, I.; Balmus, N.C.M.; Boyd, T.K.; Brundler, M.-A.; Derricott, H.; Evans, M.J.; Faye-Petersen, O.M.; Gillan, J.E.; et al. Sampling and Definitions of Placental Lesions: Amsterdam Placental Workshop Group Consensus Statement. Arch. Pathol. Lab. Med. 2016, 140, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Gebrane-Younes, J.; Hoang, N.M.; Orcel, L. Ultrastructure of human umbilical vessels: A possible role in amniotic fluid formation? Placenta 1986, 7, 173–185. [Google Scholar] [CrossRef]
- Anthony, F.W.; Mukhtar, D.D.; Pickett, M.A.; Cameron, I.T. Progesterone up-regulates WT1 mRNA and protein, and alters the relative expression of WT1 transcripts in cultured endometrial stromal cells. J. Soc. Gynecol. Investig. 2003, 10, 509–516. [Google Scholar] [CrossRef]
- Werner, H.; Rauscher, F.J.; Sukhatme, V.P.; A Drummond, I.; Roberts, C.T.; Leroith, D. Transcriptional repression of the insulin-like growth factor I receptor (IGF-I-R) gene by the tumor suppressor WT1 involves binding to sequences both upstream and downstream of the IGF-I-R gene transcription start site. J. Boil. Chem. 1994, 269, 12577–12582. [Google Scholar]
- Roberts, C.T. Control of insulin-like growth factor (IGF) action by regulation of IGF-I receptor expression. Endocr. J. 1996, 43, S49–S55. [Google Scholar] [CrossRef] [Green Version]
- Moffett, P.; Bruening, W.; Nakagama, H.; Bardeesy, N.A.; Housman, D.; Housman, D.E.; Pelletier, J. Antagonism of WT1 activity by protein self-association. Proc. Natl. Acad. Sci. USA 1995, 92, 11105–11109. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Huang, K.; Palmer, R.; Truong, V.B.; Herzlinger, D.; A Kolquist, K.; Wong, J.; Paulding, C.; Yoon, S.K.; Gerald, W.; et al. The Wilms tumor suppressor WT1 encodes a transcriptional activator of amphiregulin. Cell 1999, 98, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Ciarloni, L.; Mallepell, S.; Brisken, C. Amphiregulin is an essential mediator of estrogen receptor alpha function in mammary gland development. Proc. Natl. Acad. Sci. USA 2007, 104, 5455–5460. [Google Scholar] [CrossRef] [Green Version]
- Pronier, E.; Bowman, R.L.; Ahn, J.; Glass, J.; Kandoth, C.; Merlinsky, T.R.; Whitfield, J.T.; Durham, B.H.; Gruet, A.; Somasundara, A.V.H.; et al. Genetic and epigenetic evolution as a contributor to WT1-mutant leukemogenesis. Blood 2018, 132, 1265–1278. [Google Scholar] [CrossRef]
- Moser, G.; Windsperger, K.; Pollheimer, J.; Lopes, S.C.D.S.; Huppertz, B. Human trophoblast invasion: New and unexpected routes and functions. Histochem. Cell Boil. 2018, 150, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.; Kirschner, K.M. Oxygen-Dependent Gene Expression in Development and Cancer: Lessons Learned from the Wilms’ Tumor Gene, WT1. Front. Mol. Neurosci 2011, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Small, T.W.; Bolender, Z.; Bueno, C.; O’Neil, C.; Nong, Z.; Rushlow, W.; Rajakumar, N.; Kandel, C.; Strong, J.; Madrenas, J.; et al. Wilms’ tumor 1-associating protein regulates the proliferation of vascular smooth muscle cells. Circ. Res. 2006, 99, 1338–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, T.W.; Penalva, L.O.; Pickering, J.G. Vascular biology and the sex of flies: Regulation of vascular smooth muscle cell proliferation by wilms’ tumor 1-associating protein. Trends Cardiovasc. Med. 2007, 17, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Estrada, O.M.M.; Lettice, L.A.; Essafi, A.; Guadix, J.A.; Slight, J.; Velecela, V.; Hall, E.; Reichmann, J.; Devenney, P.S.; Hohenstein, P.; et al. WT1 is required for cardiovascular progenitor cell formation through transcriptional control of Snail and E-cadherin. Nat. Genet. 2010, 42, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, Y.Y.; Hastie, N.D. The role of WT1 in regulating mesenchyme in cancer, development, and tissue homeostasis. Trends Genet. 2012, 28, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Chau, Y.-Y.; Bandiera, R.; Serrels, A.; Estrada, O.M.M.; Qing, W.; Lee, M.; Slight, J.; Thornburn, A.; Berry, R.; McHaffie, S.; et al. Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nat. Cell Biol. 2014, 16, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, Y.Y.; Hastie, N. WT1, the mesothelium and the origins and heterogeneity of visceral fat progenitors. Adipocyte 2015, 4, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, S.; Canis, M.; Darcha, C.; Déchelotte, P.J.; Pouly, J.-L.; Mage, G. Expression of WT1 is down-regulated in eutopic endometrium obtained during the midsecretory phase from patients with endometriosis. Fertil. Steril. 2006, 86, 554–558. [Google Scholar] [CrossRef]
- Sbracia, M.; Scarpellini, F.; Zupi, E.; Manna, C.; Marconi, D.; Romanini, C.; Alo, P.; Tondo, U.; Grasso, J. Differential expression of IGF-I and IGF-II in eutopic and ectopic endometria of women with endometriosis and in women without endometriosis. Am. J. Reprod. Immunol. 1997, 37, 326–329. [Google Scholar] [CrossRef]
- Gurgan, T.; Bukulmez, O.; Yarali, H.; Tanir, M.; Akyildiz, S. Serum and peritoneal fluid levels of IGF I and II and insulinlike growth binding protein-3 in endometriosis. J. Reprod. Med. 1999, 44, 450–454. [Google Scholar]
- Reeve, A.E.; Eccles, M.R.; Wilkins, R.J.; Bell, G.I.; Millow, L.J. Expression of insulin-like growth factor-II transcripts in Wilms’ tumour. Nature 1985, 317, 258–260. [Google Scholar] [CrossRef]
Figure 1.
Hematoxylin and eosin (H+E) staining and immunostaining for Wilm’s tumor 1 gene (WT1) expression of muscular arteries in the soft tissue of the lower leg, after traumatic amputation and the ovary tissue of peri-menopausal women (400× magnification).
Figure 1.
Hematoxylin and eosin (H+E) staining and immunostaining for Wilm’s tumor 1 gene (WT1) expression of muscular arteries in the soft tissue of the lower leg, after traumatic amputation and the ovary tissue of peri-menopausal women (400× magnification).

Figure 2.
H+E staining and immunostaining for WT1 expression of proliferative endometrium, with glandular, stromal cells and spiral artery and myometrium with spiral artery (400× magnification).
Figure 2.
H+E staining and immunostaining for WT1 expression of proliferative endometrium, with glandular, stromal cells and spiral artery and myometrium with spiral artery (400× magnification).

Figure 3.
H+E staining and immunostaining for WT1 expression of proliferative endometrium and adenomyosis in hysterectomy specimen. Noted a significant reduction of WT1 signals within the stromal cells (400× magnification).
Figure 3.
H+E staining and immunostaining for WT1 expression of proliferative endometrium and adenomyosis in hysterectomy specimen. Noted a significant reduction of WT1 signals within the stromal cells (400× magnification).

Figure 4.
H+E staining and immunostaining for WT1 expression of secretory endometrium and myometrium (400× magnification).
Figure 4.
H+E staining and immunostaining for WT1 expression of secretory endometrium and myometrium (400× magnification).

Figure 5.
H+E staining and immunostaining for WT1 expression of decidua at 10 weeks’ gestation of therapeutic abortion. Spiral artery with mural hypertrophy/hyperplasia (top panel) and endovascular trophoblastic plug (bottom panel) (400× magnification).
Figure 5.
H+E staining and immunostaining for WT1 expression of decidua at 10 weeks’ gestation of therapeutic abortion. Spiral artery with mural hypertrophy/hyperplasia (top panel) and endovascular trophoblastic plug (bottom panel) (400× magnification).

Figure 6.
H+E staining and immunostaining for WT1 expression of decidua at 17 weeks gestations, with inevitable abortion and chorioamnionitis. Spiral artery with mural hypertrophy/hyperplasia (top panel) and endovascular trophoblastic plug (bottom panel) (400× magnification).
Figure 6.
H+E staining and immunostaining for WT1 expression of decidua at 17 weeks gestations, with inevitable abortion and chorioamnionitis. Spiral artery with mural hypertrophy/hyperplasia (top panel) and endovascular trophoblastic plug (bottom panel) (400× magnification).

Figure 7.
H+E staining and immunostaining for WT1 expression of decidual vessels in the membrane roll (vera) and decidua basalis at 39 weeks (decidual vasculopathy). Spiral artery at decidua vera (top panel) and partially restoring spiral artery after vasculopathy (bottom panel) (400× magnification).
Figure 7.
H+E staining and immunostaining for WT1 expression of decidual vessels in the membrane roll (vera) and decidua basalis at 39 weeks (decidual vasculopathy). Spiral artery at decidua vera (top panel) and partially restoring spiral artery after vasculopathy (bottom panel) (400× magnification).

Figure 8.
H+E staining and immunostaining for WT1, CD56 and smooth muscle myosin heavy chain (SMYOHC) expression of decidual vessels in the membrane roll (vera) and decidua basalis at term (decidual vasculopathy), at 38 weeks. Arrows indicate the focus of regenerating endothelium and smooth muscle cells with the remaining endovascular trophoblasts (200× magnification).
Figure 8.
H+E staining and immunostaining for WT1, CD56 and smooth muscle myosin heavy chain (SMYOHC) expression of decidual vessels in the membrane roll (vera) and decidua basalis at term (decidual vasculopathy), at 38 weeks. Arrows indicate the focus of regenerating endothelium and smooth muscle cells with the remaining endovascular trophoblasts (200× magnification).

Figure 9.
H+E staining and immunostaining for WT1 expressions of tubal pregnancy, at 11 weeks after salpingectomy. Noted the tubal epithelium and tubal vessels with WT1 expression and no WT1 expression in the extravillous trophoblasts (400× magnification).
Figure 9.
H+E staining and immunostaining for WT1 expressions of tubal pregnancy, at 11 weeks after salpingectomy. Noted the tubal epithelium and tubal vessels with WT1 expression and no WT1 expression in the extravillous trophoblasts (400× magnification).

Figure 10.
H+E staining and immunostaining for WT1 and CD34 expression of muscular arteries of stem villi of 39 week placentas. WT1 was noted in smooth muscle wall but not in endothelium, whereas CD34 was only seen in endothelium (400× magnification).
Figure 10.
H+E staining and immunostaining for WT1 and CD34 expression of muscular arteries of stem villi of 39 week placentas. WT1 was noted in smooth muscle wall but not in endothelium, whereas CD34 was only seen in endothelium (400× magnification).

Figure 11.
H+E staining and immunostaining for WT1 expression of umbilical artery and vein of 39-week placenta (100× magnification).
Figure 11.
H+E staining and immunostaining for WT1 expression of umbilical artery and vein of 39-week placenta (100× magnification).

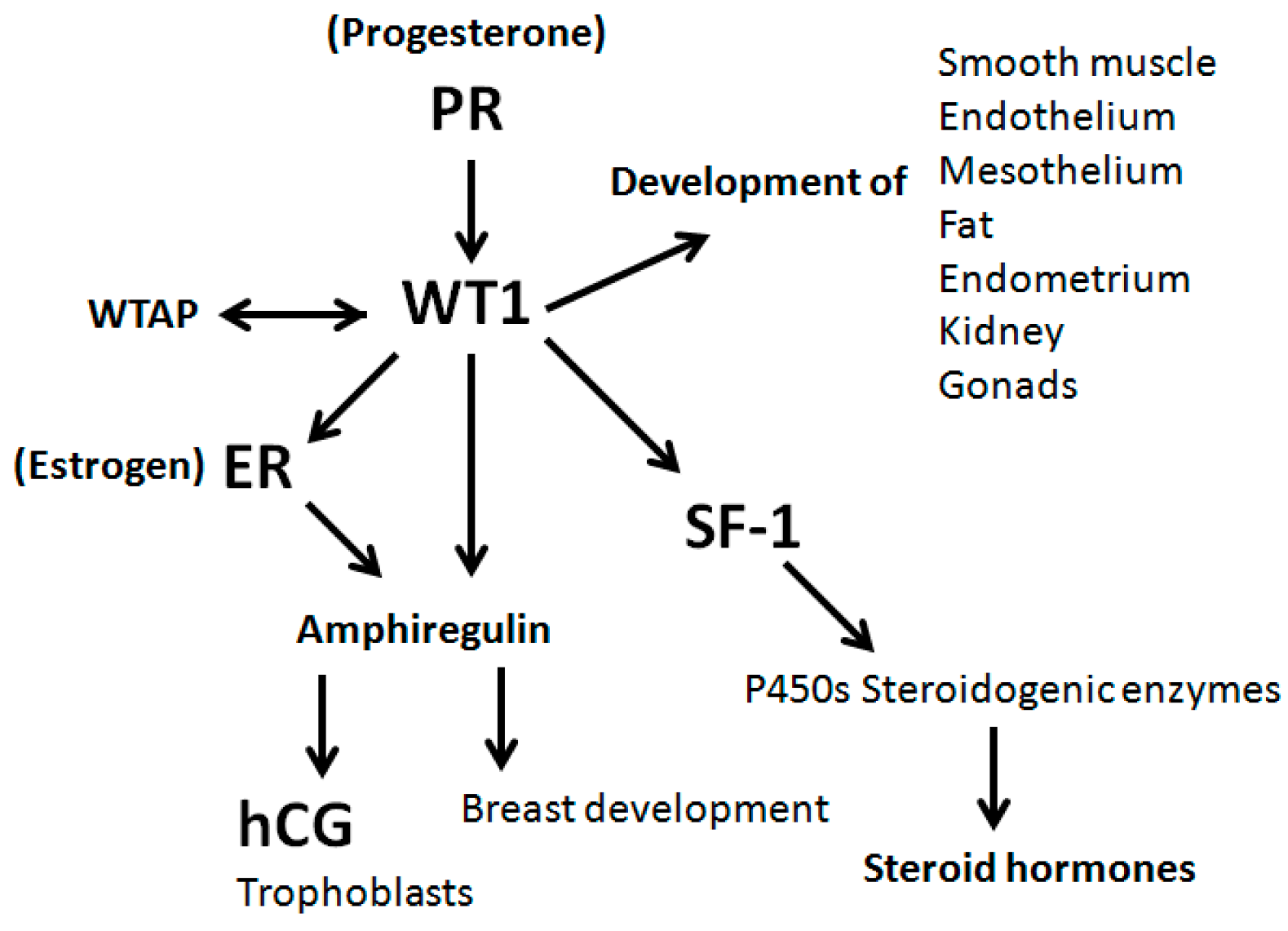
Figure 12.
Schematic representation of WT1 gene expression, in relation to other steroid and peptide hormones and receptors in regulation of organ development, homeostasis and disease development.
Figure 12.
Schematic representation of WT1 gene expression, in relation to other steroid and peptide hormones and receptors in regulation of organ development, homeostasis and disease development.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, P. Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation. Reprod. Med. 2020, 1, 17-31. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed1010003
AMA Style
Zhang P. Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation. Reproductive Medicine. 2020; 1(1):17-31. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed1010003
Chicago/Turabian StyleZhang, Peilin. 2020. "Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation" Reproductive Medicine 1, no. 1: 17-31. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed1010003