Recent Progress in Stem Cell Research of the Pituitary Gland and Pituitary Adenoma


Department of Neurosurgery, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan
*
Author to whom correspondence should be addressed.
Endocrines 2020, 1(1), 49-57; https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines1010006
Submission received: 5 June 2020
/
Revised: 16 July 2020
/
Accepted: 23 July 2020
/
Published: 27 July 2020
{kind=link}
{kind=link}
Abstract
:Regenerative medicine and anti-tumoral therapy have been developed through understanding tissue stem cells and cancer stem cells (CSCs). The concept of tissue stem cells has been applied to the pituitary gland (PG). Recently, PG stem cells (PGSCs) were successfully differentiated from human embryonic stem cells and induced pluripotent stem cells, showing an in vivo therapeutic effect in a hypopituitary model. Pituitary adenomas (PAs) are common intracranial neoplasms that are generally benign, but treatment resistance remains a major concern. The concept of CSCs applies to PA stem cells (PASCs). Genetic alterations in human PGSCs result in PASC development, leading to treatment-resistant PAs. To determine an efficient treatment against refractory PAs, it is of paramount importance to understand the relationship between PGSCs, PASCs and PAs. The goal of this review is to discuss several new findings about PGSCs and the roles of PASCs in PA tumorigenesis.
1. Introduction
1.1. Pituitary Gland
In early development, the oral ectoderm is closely juxtaposed to the neural ectoderm. In response to signals from the neural ectoderm, the oral ectoderm invaginates to produce Rathke’s pouch (oropharyngeal diverticulum), a cavity of ectodermal cells of the oropharynx. Rathke’s pouch grows between 1 and 2 weeks of gestation and develops into the anterior pituitary gland (PG). At 2–3 weeks of gestation, anterior PG cells begin to differentiate. Functional development of the anterior PG involves the regulation of several transcription factors expressed in pituitary gland stem cells (PGSCs). This process involves highly coordinated sequential steps in the cellular commitment and positioning of pituitary cell lineages. Corticotrope, thyrotrope, somatotrope, lactotrope and gonadotrope cells develop in association with respective gene expression. At 20 days, the hypophyseal portal system develops. It is organized with blood vessels connecting the hypothalamus to the anterior pituitary, which exchange hormones between them [1,2].
PG morphogenesis depends on neural ectodermal signals such as bone morphogenetic protein 4 (BMP4). Gradients of BMP4, fibroblast growth factor (FGF) 8 and Sonic hedgehog (Shh) are important for ventral side developmental patterning and the expression of transcription factors. Wnt4, Wnt5 and Notch are also essential for PG cell proliferation [3,4].
1.2. Pituitary Adenoma
Pituitary adenomas (PAs) are common intracranial neoplasms. These benign epithelial tumors originate from intrinsic cells of the adenohypophysis. PAs have a prevalence from 1 in 865 adults to 1 in 2688 adults [5]. Most cases of PAs are histopathologically benign, and pituitary carcinomas are extremely rare [6].
PAs can be traditionally classified as either (1) hormone-secreting adenomas, as defined by the presence of one or more hyperplastic pituitary endocrine populations (lactotrophs, gonadotrophs, somatotrophs, corticotrophs and thyrotrophs) or (2) non-functional adenomas that are thought to be mainly derived from gonadotrophs [7]. Some PAs exhibit a more aggressive course and cause significant morbidity. Atypical morphological features, such as cavernous sinus invasion, have been demonstrated as a sign of aggressive behavior, which are emphasized in the revised 2017 World Health Organization classification [8].
1.3. Tissue and Cancer Stem Cells
Tissue stem cells (TSCs) were first identified in the 1950s. TSCs are undifferentiated cells capable of proliferation and self-renewal, which also produce differentiated functional progeny and are involved in repair and regeneration [9]. The concept of cancer stem cells (CSCs) was established following the discovery of TSCs in acute myeloid leukemia in 1994 [10].
CSCs have two basic properties: self-renewal and differentiation into multiple lineages. Tumor malignancy correlates to the number of CSCs that drive tumor growth and recurrence [11]. Recently, these concepts were applied to the PG and PAs.
2. Pituitary Gland Stem Cells
2.1. Historical Background
The anterior PG contains hormone-producing cells and non-hormone-producing cells such as folliculostellate cells and vascular cells. Because anterior pituitary hormonal cells are highly differentiated, and the turnover rate, which considerably declines from infancy to adulthood, is as low as 1.58% per day [12,13], the existence of PGSCs has been controversial [14]. However, recent studies have demonstrated that a subpopulation of undifferentiated stem/progenitor cells, with self-renewal and multipotent capacities, is maintained in the adult anterior PG [15]. It has been reported that gonadotrophs and thyrotrophs increase after gonadectomy and thyroidectomy, suggesting the existence of PGSCs [16,17]. PGSCs are considered to supply hormone-producing cells in response to stress, damage and physiological demand [16].
2.2. Markers
In 2008, the transcription factor sex-determining region Y-box 2 (SOX2) was detected in non-hormone-producing cells of the adult PG [18]. In vitro assessment showed that SOX2-positive cells had a sphere-forming capacity and could differentiate into multiple lineages, such as S100β-positive folliculostellate cells and hormone-producing cells [18]. Most SOX2-positive cells are in a quiescent state in the adult PG. They are activated as the origin of hormone-producing cells in response to injury and capable of regeneration [19].
2.3. Stem Cell Niche
TSCs are maintained within the local microenvironment “niche”. Niches regulate multipotency, self-renewal, asymmetric cell division, and migration from niches for differentiation via signaling by soluble factors, cell surface proteins, and extracellular matrices. In general, CSCs reside in their niches that sustain the self-renewal of CSCs and trigger tumorigenesis [22]. Two types of niches—the marginal cell layer and SOX2 cluster—have been proposed in the adult PG. A recent study isolated and successfully cultured SOX2 cluster niches [13]. To date, the relationship between niches and pituitary tumorigenesis has remained unclear. Therefore, further studies are warranted on this topic.
2.4. Differentiation
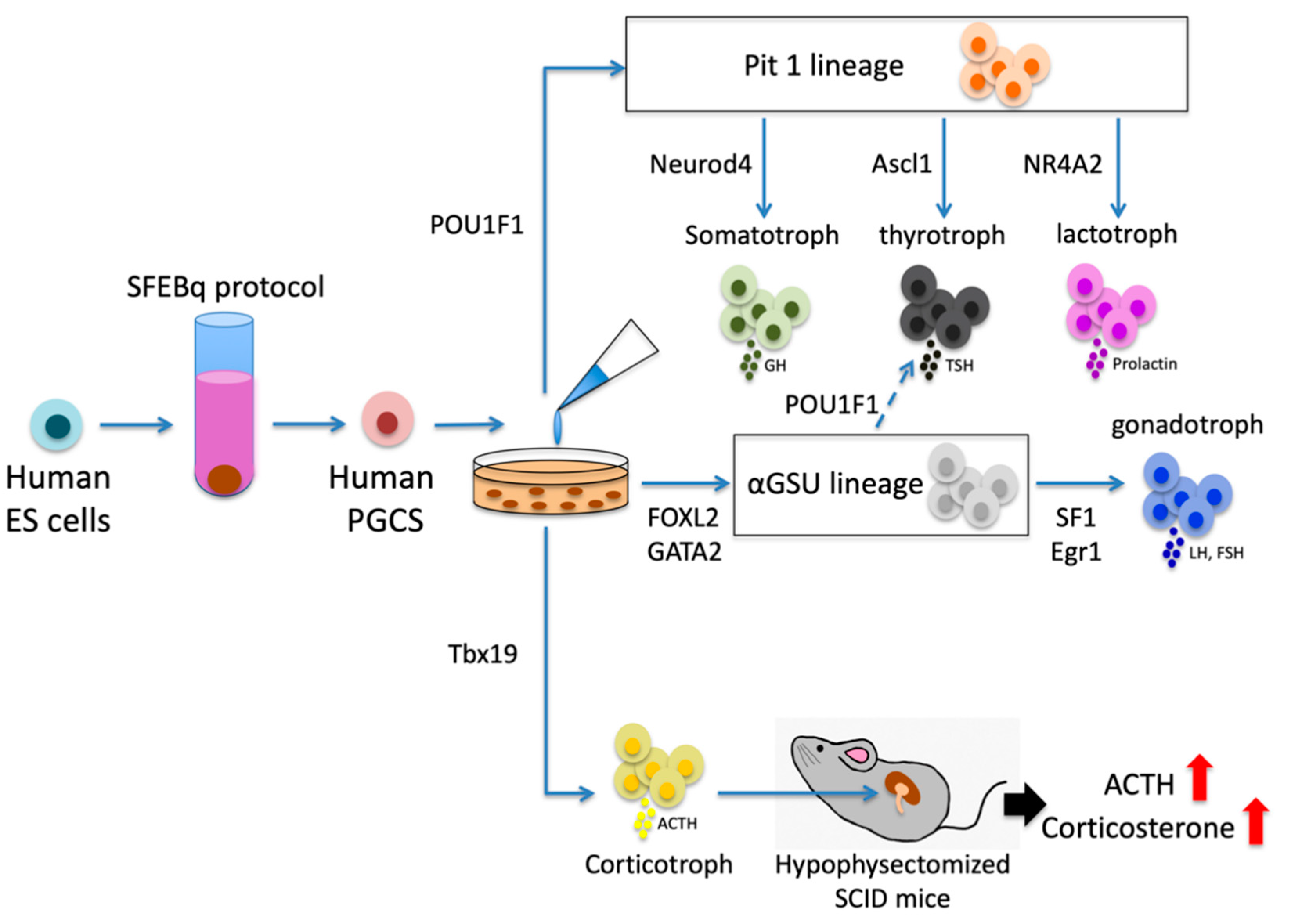
The pituitary placode is known to differentiate into three lineages—the Pit1 lineage (somatotroph-lactotroph-thyrotroph), αGSU lineage (gonadotroph-thyrotroph) and corticotroph lineage—under the control of various transcription factors. Generation of the Pit1 lineage depends on POU1F1, which is positively regulated by Prop1 and negatively regulated by Hesx1 [23]. This lineage terminally differentiates into somatotrophs under the control of Neurod4 and thyrotrophs under the control of Ascl1 [3]. NR4A2, which binds adjacently to POU1F1 at the Prl promoter, synergizes with POU1F1 to enhance Prl gene expression, leading to the Pit1 lineage to differentiate into lactotrophs [24]. FOXL2 and GATA2 stimulate the expression of αGSU. Cells expressing αGSU differentiate into gonadotrophs under the control of SF1 and Egr1 and thyrotrophs under the control of POU1F1 [25,26]. Tbx19 is transcription factor of the corticotroph lineage and Neurod1 is required for early corticotroph differentiation, but not lineage commitment [27] (Figure 1).
2.5. Treatment Strategies Using PGSCs
Generating a human anterior PG that retains regulatory hormonal control in vivo was necessary to develop cell-based therapies for pituitary deficiencies. In 2016, Ozone et al. used human ES cells to differentiate PG cells by employing serum-free floating culture of embryoid body-like aggregates prepared by the quick reaggregation (SFEBq) protocol [28,29,30]. In vitro self-forming of the anterior PG was followed by co-induction of hypothalamic and oral ectoderms in medium with BMP, Shh, and FGF. The juxtaposition of these tissues promoted the formation of a Rathke’s pouch-like pituitary placode that differentiated into hormone-producing cells. These cells clearly responded to hormonal feedback signals [28]. Portal vessels from the hypothalamus irrigate defined areas within the pituitary to finely control the distribution of hypothalamic hormones. Additionally, the vascular microarchitecture and perivascular space are important for hormone-producing cells to secrete produced hormones properly [31]. Therefore, from the viewpoint of actual clinical application, for stimulation by upstream hormones and secreting hormones appropriately, establishing a connection between blood flow and transplanted hormone-producing cells is needed. There have been few reports on the relationship between transplanted cells and blood flow; further research is expected.
Some studies have reported the ability of PGSCs in hypopituitary model mice. The number of somatotrophs recovered over a 5-month period in adult GH/Cre-inducible diphtheria toxin receptor (iDTR) mice was affected by a rapid increase in SOX2-positive cells [32]. Furthermore, after transplanting corticotrophs generated from human ES cells under the renal capsule, plasma ACTH and corticosterone levels were elevated, leading to rescue of physical activity at 10 d after transplantation and improvement of survival in hypopituitary model mice [28]. This report showed a therapeutic effect in an in vivo hypopituitary model using a stem cell-based therapy (Figure 1). Further studies are warranted to confirm the importance of PGSCs in disease models.
In recent years, induced pluripotent stem (iPS) cells have been established from both mice and humans [33,34]. iPS cells overcome the ethical issues of ES cells. In 2016, Zimmer et al. demonstrated that human iPS cells can differentiate into hormone-producing cells, which holds promise for the future [35].
3. Pituitary Adenoma Stem Cells
3.1. Tumor-Initiating Capacity
In general, CSCs are derived from normal TSCs or re-differentiate and/or de-differentiate from progenitor/differentiated cells. Some studies have discussed the relationship between genetic alterations in human PGSCs and tumors formed by pituitary adenoma stem cells (PASCs) [36].
The relationship between stem cells and PAs has been demonstrated in PA model mice. Gleiberman et al. crossed Rb (+/−) mice, which develop PA in almost 100% of cases, with nestin-GFP mice, and the crossed mice developed PA nodules surrounded by nestin-GFP-positive cells [37]. Dopamine receptor D2 knockout [Drd2 (−/−)] mice, prolactinoma model mice, contain more SOX2-positive cells in their anterior pituitary than control mice [38].
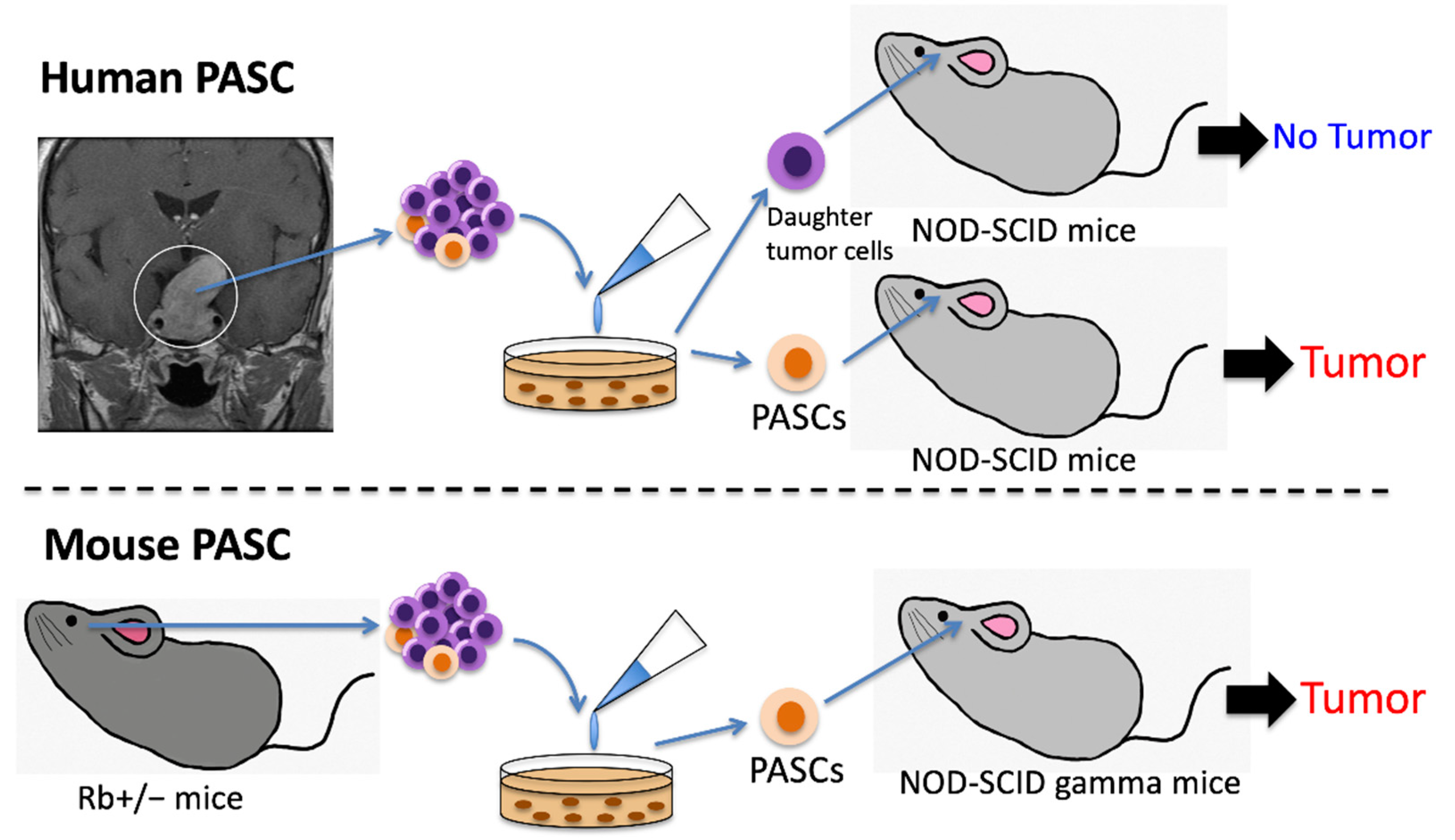
Human PASCs were first isolated from somatotropinomas and non-functioning adenomas using the sphere-forming system in 2009, which initiated hormone-producing PAs as xenografts. The initiating PAs produced GH, PRL, FSH, LH and TSH, thereby responding to hypothalamic stimulation [39] (Figure 2).
Since then, some studies have demonstrated that stem cell side populations can be isolated from human PAs (mainly somatotropinomas and non-functional PAs) using several stem cell markers, such as SOX2, CD133 and CD15 and successful tumor cell proliferation as xenografts has been shown [40,41]. In particular, the relationship between CXCR4 expression and the tumor-initiating capacity of PASCs has been discussed [42]. Tunici et al. clearly revealed the multilineage potential of PASCs isolated from human PAs. In their report, PASCs treated with epidermal growth factor and bFGF expressed S-100 (molecular marker of folliculostellate cells) and synaptophysin in vitro [14].
In contrast, some studies have demonstrated the difficulty of tumorigenesis as xenografts even if stem cell side populations can be isolated successfully [38,43]. Wurth et al. demonstrated that the CD133-positive side population isolated from human PAs proliferates for only 2 months [43]. The cause of the discrepancy in the status of tumorigenesis among these studies remains unclear. It may be attributed to the heterogeneity of different tumor types, including somatotropinomas and non-functioning adenomas and the difference in culture methodologies.
Recently, some factors promoting the tumorigenesis of PASCs have been identified. Mertens et al. demonstrated that upregulated stem cell markers (CD44, CXCR4, KIT, KLF4, Nestin and SOX2) and mesenchymal markers (VIM and fibronectin 1), as well as downregulated epithelial markers, including E-cadherin (CDH1) and claudin-1 (CLDN1), were associated with the tumorigenesis of human PASCs as xenografts [38]. This suggests that epithelial–mesenchymal transition (EMT) is an important process for PAs. EMT is involved in morphological changes in tumors, from tightly packed columnar-type cells to more loosely distributed cells [44]. In fact, a recent study of human PA specimens demonstrated that SNAIL1, an EMT-related transcriptional factor, is associated with the suprasellar expansion of human PAs [45]. Other signaling pathways, such as CXCL12 CXCR4 and the activation of mutated β-catenin, Wnt, Notch and MAPK/ERK, have also been reported to promote the proliferation of PASCs [36,38,46,47]. It is known that angiogenesis promotes PA progression [48]. Interestingly, PASCs themselves promote angiogenesis in PAs by upregulated expression of PECAM1 and VCAM1 [38].
Overactivation of the WNT pathway in SOX2-positive cells is involved in the tumor initiation of both pituitary adenoma and craniopharyngioma in mice and humans [19,49]. In craniopharyngioma, the proliferation of SOX2-mutated cells results in forming β-catenin-accumulating cell clusters that secrete tumorigenic factors such as SHH, BMP4, WNT, interleukins, chemokines and growth factors. In contrast to the classical paradigm of CSCs, which cell-autonomously generate tumors, the cell clusters act as paracrine tumorigenesis paradigm which promote tumorigenesis either directly or indirectly through cell–cell signaling interactions or microenvironmental changes [19,47]. This paracrine tumorigenesis paradigm may be applicable to PAs.
Donangelo et al. first isolated PASCs from murine PAs in 2014. SCA1-positive tumor cells expressing SOX2 and Nestin were isolated from Rb+/− mice with PAs, which exhibited a tumor-forming capacity when re-transplanted into the mouse brain [50] (Figure 2). Further studies of these genetically engineered mouse models with PAs may provide information on the detailed characteristics of PASCs.
3.2. Treatment Strategies for PASCs
Recently, the relationship between upregulated PASC markers and high proliferative activity was evaluated using human PA specimens [40,41]. Therefore, PASC-targeted therapy has attracted attention. Zubeldia-Brenner et al. demonstrated that a γ-secretase inhibitor downregulates stemness by suppressing Notch signaling, resulting in the reduction of prolactin-producing xenografted tumors [51]. Mertens et al. reported that the injection of AMD3100, which is a CXCR4 antagonist, suppresses the tumor growth of PASCs isolated from murine corticotropinoma in vivo [38]. Inhibitors that target TGF-β receptor I kinase [52] and Wnt/β-catenin [53] might be useful to treat progressive PAs.
Treatment strategies for PASCs might be applied to multiple endocrine neoplasia type 1 (MEN1) that is characterized by a combination of tumors of the anterior pituitary, parathyroid, gastrointestinal tract and pancreas [54]. MEN1-related PAs are often invasive and resistant to treatment [55]. Menin deficiency is the consequence of a MEN1 mutation. It is associated with Notch signaling, which maintains PASCs [56] and TGFβ signaling that promotes EMT of PASCs [57]. Thus, PASC-targeted therapy may be a novel treatment strategy for aggressive MEN-related PAs.
Specific markers are needed to selectively target PASCs, which have not been established to date. Although SOX2 is frequently used as a representative PASC marker, it is also expressed on neural stem cells [58]. Recently, Horiguchi et al. demonstrated that CD9 is coexpressed in most S100β/SOX2-positive cells in PAs, which might be a potent new marker of PASCs [59].
4. Conclusions and Future Perspectives
Currently, there is no curative therapy for pituitary deficiencies. PGSCs supply hormone-producing cells in response to stress, damage and physiological demand. PGSCs differentiate into hormone-producing cells that clearly respond to hormonal feedback signals and demonstrate a therapeutic effect in an in vivo hypopituitary model. ES and iPS cells are sources of PGSCs, leading to a novel type of cell-based therapy by transplanting hormone-producing cells generated from PGSCs for pituitary deficiencies.
The concept of PASCs has attracted attention. Recent studies suggest that PASCs promote tumorigenesis via several pathways, including the paracrine tumorigenesis paradigm, and targeting one of these pathways results in the reduction in xenografted PA. This implies that PASC-targeted therapy could be a novel therapy for unresectable or recurrent PAs. Although the tumor-initiating capacity of PASCs remains controversial, the establishment of PASC-derived PA animal models may elucidate mechanisms of tumorigenesis, treatment resistance and novel treatment strategies of PA.
Author Contributions
Conceptualization, M.T. (Masataro Toda); writing—original draft preparation, M.T. (Masataro Toda); writing—review and editing, R.T. and M.T. (Masahiro Toda); funding acquisition, M.T. (Masahiro Toda). All authors have read and agreed to the published version of the manuscript.
Funding
This communication was funded by grants from the Japan Society for the Promotion of Science (JSPS) (17H04306 and 18K19622 to T. Masahiro).
Acknowledgments
We thank Mitchell Arico from Edanz Group (https://en-author-services.edanzgroup.com/) for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bancalari, R.E.; Gregory, L.C.; McCabe, M.J.; Dattani, M.T. Pituitary gland development: An update. Endocr. Dev. 2012, 23, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.D.; Stone, B.M.; Raetzman, L.T.; Camper, S.A. Cell proliferation and vascularization in mouse models of pituitary hormone deficiency. Mol. Endocrinol. 2006, 20, 1378–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhang, J.; Tollkuhn, J.; Ohsawa, R.; Bresnick, E.H.; Guillemot, F.; Kageyama, R.; Rosenfeld, M.G. Sustained Notch signaling in progenitors is required for sequential emergence of distinct cell lineages during organogenesis. Genes Dev. 2006, 20, 2739–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potok, M.A.; Cha, K.B.; Hunt, A.; Brinkmeier, M.L.; Leitges, M.; Kispert, A.; Camper, S.A. WNT signaling affects gene expression in the ventral diencephalon and pituitary gland growth. Dev. Dyn. 2008, 237, 1006–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molitch, M.E. Diagnosis and Treatment of Pituitary Adenomas: A Review. Jamanetwork 2017, 317, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Melmed, S. Pathogenesis of pituitary tumors. Nat. Rev. Endocrinol. 2011, 7, 257–266. [Google Scholar] [CrossRef]
- Melmed, S. Pituitary tumors. Endocrinol. Metab. Clin. N. Am. 2015, 44, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mete, O.; Lopes, M.B. Overview of the 2017 WHO Classification of Pituitary Tumors. Endocr. Pathol. 2017, 28, 228–243. [Google Scholar] [CrossRef]
- Loeffler, M.; Roeder, I. Tissue stem cells: Definition, plasticity, heterogeneity, self-organization and models—A conceptual approach. Cells Tissues Organs 2002, 171, 8–26. [Google Scholar] [CrossRef]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Levy, A. Physiological Implications of Pituitary Trophic Activity. J. Endocrinol. 2002, 174, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Nishimura, N.; Ueharu, H.; Kanno, N.; Higuchi, M.; Horiguchi, K.; Kato, T.; Kato, Y. Isolation of adult pituitary stem/progenitor cell clusters located in the parenchyma of the rat anterior lobe. Stem Cell Res. 2016, 17, 318–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunici, P.; Yu, J.S. Pituitary Adenoma Stem Cells. Methods Mol. Biol. 2009, 568, 195–201. [Google Scholar] [CrossRef]
- Chen, J.; Gremeaux, L.; Fu, Q.; Liekens, D.; Van Laere, S.; Vankelecom, H. Pituitary progenitor cells tracked down by side population dissection. Stem Cells 2009, 27, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Rizzoti, K.; Akiyama, H.; Lovell-Badge, R. Mobilized adult pituitary stem cells contribute to endocrine regeneration in response to physiological demand. Cell Stem Cell 2013, 13, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, F.; Ishikawa, H. Identification of the thyrotrophs with the gonadotrophs in the anterior pituitaries of thyroidectomized rats. Endocrinol. Jpn. 1969, 16, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Fauquier, T.; Rizzoti, K.; Dattani, M.; Lovell-Badge, R.; Robinson, I.C. SOX2-expressing progenitor cells generate all of the major cell types in the adult mouse pituitary gland. Proc. Natl. Acad. Sci. USA 2008, 105, 2907–2912. [Google Scholar] [CrossRef] [Green Version]
- Andoniadou, C.L.; Matsushima, D.; Mousavy Gharavy, S.N.; Signore, M.; Mackintosh, A.I.; Schaeffer, M.; Gaston-Massuet, C.; Mollard, P.; Jacques, T.S.; Le Tissier, P.; et al. Sox2(+) stem/progenitor cells in the adult mouse pituitary support organ homeostasis and have tumor-inducing potential. Cell Stem Cell 2013, 13, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Kato, T.; Higuchi, M.; Yako, H.; Chen, M.; Kanno, N.; Ueharu, H.; Kato, Y. Rapid transition of NESTIN-expressing dividing cells from PROP1-positive to PIT1-positive advances prenatal pituitary development. J. Neuroendocrinol. 2013, 25, 779–791. [Google Scholar] [CrossRef]
- Mantovani, G.; Giardino, E.; Treppiedi, D.; Catalano, R.; Mangili, F.; Spada, A.; Arosio, M.; Peverelli, E. Stem Cells in Pituitary Tumors: Experimental Evidence Supporting Their Existence and Their Role in Tumor Clinical Behavior. Front. Endocrinol. 2019, 10, 745. [Google Scholar] [CrossRef] [Green Version]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Olson, L.E.; Tollkuhn, J.; Scafoglio, C.; Krones, A.; Zhang, J.; Ohgi, K.A.; Wu, W.; Taketo, M.M.; Kemler, R.; Grosschedl, R.; et al. Homeodomain-mediated beta-catenin-dependent switching events dictate cell-lineage determination. Cell 2006, 125, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peel, M.T.; Ho, Y.; Liebhaber, S.A. Transcriptome Analyses of Female Somatotropes and Lactotropes Reveal Novel Regulators of Cell Identity in the Pituitary. Endocrinology 2018, 159, 3965–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellsworth, B.S.; Egashira, N.; Haller, J.L.; Butts, D.L.; Cocquet, J.; Clay, C.M.; Osamura, R.Y.; Camper, S.A. FOXL2 in the pituitary: Molecular, genetic, and developmental analysis. Mol. Endocrinol. 2006, 20, 2796–2805. [Google Scholar] [CrossRef] [PubMed]
- Egashira, N.; Takekoshi, S.; Takei, M.; Teramoto, A.; Osamura, R.Y. Expression of FOXL2 in human normal pituitaries and pituitary adenomas. Mod. Pathol. 2011, 24, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Lamolet, B.; Poulin, G.; Chu, K.; Guillemot, F.; Tsai, M.J.; Drouin, J. Tpit-independent function of NeuroD1(BETA2) in pituitary corticotroph differentiation. Mol. Endocrinol. 2004, 18, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Ozone, C.; Suga, H.; Eiraku, M.; Kadoshima, T.; Yonemura, S.; Takata, N.; Oiso, Y.; Tsuji, T.; Sasai, Y. Functional anterior pituitary generated in self-organizing culture of human embryonic stem cells. Nat. Commun. 2016, 7, 10351. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Kamiya, D.; Nishiyama, A.; Katayama, T.; Nozaki, S.; Kawasaki, H.; Watanabe, Y.; Mizuseki, K.; Sasai, Y. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat. Neurosci. 2005, 8, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Eiraku, M.; Watanabe, K.; Matsuo-Takasaki, M.; Kawada, M.; Yonemura, S.; Matsumura, M.; Wataya, T.; Nishiyama, A.; Muguruma, K.; Sasai, Y. Self-organized formation of polarized cortical tissues from ESCs and its active manipulation by extrinsic signals. Cell Stem Cell 2008, 3, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Lafont, C.; Desarménien, M.G.; Cassou, M.; Molino, F.; Lecoq, J.; Hodson, D.; Lacampagne, A.; Mennessier, G.; El Yandouzi, T.; Carmignac, D.; et al. Cellular in vivo imaging reveals coordinated regulation of pituitary microcirculation and GH cell network function. Proc. Natl. Acad. Sci. USA 2010, 107, 4465–4470. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Gremeaux, L.; Luque, R.M.; Liekens, D.; Chen, J.; Buch, T.; Waisman, A.; Kineman, R.; Vankelecom, H. The adult pituitary shows stem/progenitor cell activation in response to injury and is capable of regeneration. Endocrinology 2012, 153, 3224–3235. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, B.; Piao, J.; Ramnarine, K.; Tomishima, M.J.; Tabar, V.; Studer, L. Derivation of Diverse Hormone-Releasing Pituitary Cells from Human Pluripotent Stem Cells. Stem Cell Rep. 2016, 6, 858–872. [Google Scholar] [CrossRef] [Green Version]
- Florio, T. Adult pituitary stem cells: From pituitary plasticity to adenoma development. Neuroendocrinology 2011, 94, 265–277. [Google Scholar] [CrossRef]
- Gleiberman, A.S.; Michurina, T.; Encinas, J.M.; Roig, J.L.; Krasnov, P.; Balordi, F.; Fishell, G.; Rosenfeld, M.G.; Enikolopov, G. Genetic approaches identify adult pituitary stem cells. Proc. Natl. Acad. Sci. USA 2008, 105, 6332–6337. [Google Scholar] [CrossRef] [Green Version]
- Mertens, F.; Gremeaux, L.; Chen, J.; Fu, Q.; Willems, C.; Roose, H.; Govaere, O.; Roskams, T.; Cristina, C.; Becu-Villalobos, D.; et al. Pituitary tumors contain a side population with tumor stem cell-associated characteristics. Endocr. Relat. Cancer 2015, 22, 481–504. [Google Scholar] [CrossRef]
- Xu, Q.; Yuan, X.; Tunici, P.; Liu, G.; Fan, X.; Xu, M.; Hu, J.; Hwang, J.Y.; Farkas, D.L.; Black, K.L.; et al. Isolation of tumour stem-like cells from benign tumours. Br. J. Cancer 2009, 101, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ye, H.; Wang, X.; Tang, X.; Mao, Y.; Zhao, Y.; Wu, Z.; Mao, X.O.; Xie, L.; Jin, K.; et al. Evidence of brain tumor stem progenitor-like cells with low proliferative capacity in human benign pituitary adenoma. Cancer Lett. 2014, 349, 61–66. [Google Scholar] [CrossRef]
- Manoranjan, B.; Mahendram, S.; Almenawer, S.A.; Venugopal, C.; McFarlane, N.; Hallett, R.; Vijayakumar, T.; Algird, A.; Murty, N.K.; Sommer, D.D.; et al. The identification of human pituitary adenoma-initiating cells. Acta Neuropathol. Commun. 2016, 4, 125. [Google Scholar] [CrossRef] [Green Version]
- Wurth, R.; Thellung, S.; Corsaro, A.; Barbieri, F.; Florio, T. Experimental Evidence and Clinical Implications of Pituitary Adenoma Stem Cells. Front. Endocrinol. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurth, R.; Barbieri, F.; Pattarozzi, A.; Gaudenzi, G.; Gatto, F.; Fiaschi, P.; Ravetti, J.L.; Zona, G.; Daga, A.; Persani, L.; et al. Phenotypical and Pharmacological Characterization of Stem-Like Cells in Human Pituitary Adenomas. Mol. Neurobiol. 2017, 54, 4879–4895. [Google Scholar] [CrossRef] [PubMed]
- Perez-Moreno, M.A.; Locascio, A.; Rodrigo, I.; Dhondt, G.; Portillo, F.; Nieto, M.A.; Cano, A. A new role for E12/E47 in the repression of E-cadherin expression and epithelial-mesenchymal transitions. J. Biol. Chem. 2001, 276, 27424–27431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, R.; Ohara, K.; Morimoto, Y.; Kosugi, K.; Oishi, Y.; Sato, M.; Yoshida, K.; Toda, M. PITX2 Expression in Non-functional Pituitary Neuroendocrine Tumor with Cavernous Sinus Invasion. Endocr. Pathol. 2019, 30, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Haston, S.; Pozzi, S.; Carreno, G.; Manshaei, S.; Panousopoulos, L.; Gonzalez-Meljem, J.M.; Apps, J.R.; Virasami, A.; Thavaraj, S.; Gutteridge, A.; et al. MAPK pathway control of stem cell proliferation and differentiation in the embryonic pituitary provides insights into the pathogenesis of papillary craniopharyngioma. Development 2017, 144, 2141–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreno, G.; Gonzalez-Meljem, J.M.; Haston, S.; Martinez-Barbera, J.P. Stem cells and their role in pituitary tumorigenesis. Mol. Cell. Endocrinol. 2017, 445, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Lasorella, A.; Rothschild, G.; Yokota, Y.; Russell, R.G.; Iavarone, A. Id2 mediates tumor initiation, proliferation, and angiogenesis in Rb mutant mice. Mol. Cell. Biol. 2005, 25, 3563–3574. [Google Scholar] [CrossRef] [Green Version]
- Elston, M.S.; Gill, A.J.; Conaglen, J.V.; Clarkson, A.; Shaw, J.M.; Law, A.J.; Cook, R.J.; Little, N.S.; Clifton-Bligh, R.J.; Robinson, B.G.; et al. Wnt pathway inhibitors are strongly down-regulated in pituitary tumors. Endocrinology 2008, 149, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Donangelo, I.; Ren, S.G.; Eigler, T.; Svendsen, C.; Melmed, S. Sca1(+) murine pituitary adenoma cells show tumor-growth advantage. Endocr. Relat. Cancer 2014, 21, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Zubeldia-Brenner, L.; De Winne, C.; Perrone, S.; Rodriguez-Segui, S.A.; Willems, C.; Ornstein, A.M.; Lacau-Mengido, I.; Vankelecom, H.; Cristina, C.; Becu-Villalobos, D. Inhibition of Notch signaling attenuates pituitary adenoma growth in Nude mice. Endocr. Relat. Cancer 2019, 26, 13–29. [Google Scholar] [CrossRef]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFβ) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS ONE 2014, 9, e90353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, B.; Zhou, L.; Yu, S.; Su, Z.; Song, J.; Sun, Q.; Sha, O.; Wang, X.; Jiang, W.; et al. Prodigiosin inhibits Wnt/beta-catenin signaling and exerts anticancer activity in breast cancer cells. Proc. Natl. Acad. Sci. USA 2016, 113, 13150–13155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakker, R.V.; Newey, P.J.; Walls, G.V.; Bilezikian, J.; Dralle, H.; Ebeling, P.R.; Melmed, S.; Sakurai, A.; Tonelli, F.; Brandi, M.L. Clinical practice guidelines for multiple endocrine neoplasia type 1 (MEN1). J. Clin. Endocrinol. Metab. 2012, 97, 2990–3011. [Google Scholar] [CrossRef]
- Verges, B.; Boureille, F.; Goudet, P.; Murat, A.; Beckers, A.; Sassolas, G.; Cougard, P.; Chambe, B.; Montvernay, C.; Calender, A. Pituitary disease in MEN type 1 (MEN1): Data from the France-Belgium MEN1 multicenter study. J. Clin. Endocrinol. Metab. 2002, 87, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Stalberg, P.; Santesson, M.; Ekeblad, S.; Lejonklou, M.H.; Skogseid, B. Recognizing genes differentially regulated in vitro by the multiple endocrine neoplasia type 1 (MEN1) gene, using RNA interference and oligonucleotide microarrays. Surgery 2006, 140, 921–929; discussion 929–931. [Google Scholar] [CrossRef]
- Kaji, H.; Canaff, L.; Lebrun, J.-J.; Goltzman, D.; Hendy, G.N. Inactivation of menin, a Smad3−interacting protein, blocks transforming growth factor type b signaling. Proc. Natl. Acad. Sci. USA 2001, 98, 3837–3842. [Google Scholar] [CrossRef] [Green Version]
- Ellis, P.; Fagan, B.M.; Magness, S.T.; Hutton, S.; Taranova, O.; Hayashi, S.; McMahon, A.; Rao, M.; Pevny, L. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev. Neurosci. 2004, 26, 148–165. [Google Scholar] [CrossRef]
- Horiguchi, K.; Fujiwara, K.; Yoshida, S.; Nakakura, T.; Arae, K.; Tsukada, T.; Hasegawa, R.; Takigami, S.; Ohsako, S.; Yashiro, T.; et al. Isolation and characterisation of CD9-positive pituitary adult stem/progenitor cells in rats. Sci. Rep. 2018, 8, 5533. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Differentiation of hormone-producing cells from human ES cells. Human ES cells can be differentiated into PGSCs by the SFEBq protocol. In vitro, PGSCs form the pituitary placode that subsequently differentiates into three lineages, namely the Pit1 lineage (somatotroph-lactotroph-thyrotroph), αGSU lineage (gonadotroph-thyrotroph) and corticotroph lineage. Generation of the Pit1 lineage depends on POU1F1 and this lineage terminally differentiates into somatotrophs under the control of Neurod4, thyrotrophs under the control of Ascl1 and lactotrophs under the control of NR4A2 synergizing with POU1F1. PGSCs differentiate into the corticotroph lineage under the control of Tbx19 and the αGSU lineage under the control of FOXL2 and GATA2. The αGSU lineage differentiates into gonadotrophs under the control of SF1 and Egr1. After transplantation of generated corticotrophs into hypophysectomized SCID mice, plasma ACTH and corticosterone levels elevate, leading to rescue of physical activity and the improvement of survival in hypopituitary model mice. ES, embryonic stem; PGSC, pituitary gland stem cell; SCID, severe combined immunodeficiency; SFEBq, serum-free floating culture of embryoid body-like aggregates with quick reaggregation.
Figure 1.
Differentiation of hormone-producing cells from human ES cells. Human ES cells can be differentiated into PGSCs by the SFEBq protocol. In vitro, PGSCs form the pituitary placode that subsequently differentiates into three lineages, namely the Pit1 lineage (somatotroph-lactotroph-thyrotroph), αGSU lineage (gonadotroph-thyrotroph) and corticotroph lineage. Generation of the Pit1 lineage depends on POU1F1 and this lineage terminally differentiates into somatotrophs under the control of Neurod4, thyrotrophs under the control of Ascl1 and lactotrophs under the control of NR4A2 synergizing with POU1F1. PGSCs differentiate into the corticotroph lineage under the control of Tbx19 and the αGSU lineage under the control of FOXL2 and GATA2. The αGSU lineage differentiates into gonadotrophs under the control of SF1 and Egr1. After transplantation of generated corticotrophs into hypophysectomized SCID mice, plasma ACTH and corticosterone levels elevate, leading to rescue of physical activity and the improvement of survival in hypopituitary model mice. ES, embryonic stem; PGSC, pituitary gland stem cell; SCID, severe combined immunodeficiency; SFEBq, serum-free floating culture of embryoid body-like aggregates with quick reaggregation.

Figure 2.
Tumor formation capacities of human and mouse PASCs. Human PASCs are isolated from PAs, such as somatotropinomas and non-functioning adenomas, using CD133 and nestin as markers. These human PASCs with a high sphere-forming capacity initiated tumor formation as xenografts. In contrast, daughter tumor cells isolated from the same human PAs did not initiate tumor formation. Mouse PASCs, SCA1-positive tumor cells with expression of Sox2 and Nestin, were isolated from Rb+/− mice with PAs and exhibited a high tumor-forming capacity when retransplanted into SCID gamma mice. PA, pituitary adenoma; PASC, pituitary adenoma stem cell; SCID, severe combined immunodeficiency.
Figure 2.
Tumor formation capacities of human and mouse PASCs. Human PASCs are isolated from PAs, such as somatotropinomas and non-functioning adenomas, using CD133 and nestin as markers. These human PASCs with a high sphere-forming capacity initiated tumor formation as xenografts. In contrast, daughter tumor cells isolated from the same human PAs did not initiate tumor formation. Mouse PASCs, SCA1-positive tumor cells with expression of Sox2 and Nestin, were isolated from Rb+/− mice with PAs and exhibited a high tumor-forming capacity when retransplanted into SCID gamma mice. PA, pituitary adenoma; PASC, pituitary adenoma stem cell; SCID, severe combined immunodeficiency.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Toda, M.; Tamura, R.; Toda, M. Recent Progress in Stem Cell Research of the Pituitary Gland and Pituitary Adenoma. Endocrines 2020, 1, 49-57. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines1010006
AMA Style
Toda M, Tamura R, Toda M. Recent Progress in Stem Cell Research of the Pituitary Gland and Pituitary Adenoma. Endocrines. 2020; 1(1):49-57. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines1010006
Chicago/Turabian StyleToda, Masataro, Ryota Tamura, and Masahiro Toda. 2020. "Recent Progress in Stem Cell Research of the Pituitary Gland and Pituitary Adenoma" Endocrines 1, no. 1: 49-57. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines1010006