
The Effect of High-Intensity Exercise on Changes in Salivary and Serum Cortisol Proportion Dynamics


Department of Kinesiology, School of Health and Human Sciences, University of North Carolina at Greensboro, Greensboro, NC 27402, USA
*
Author to whom correspondence should be addressed.
Endocrines 2021, 2(1), 44-53; https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2010005
Submission received: 7 January 2021
/
Revised: 16 February 2021
/
Accepted: 19 February 2021
/
Published: 23 February 2021
(This article belongs to the Special Issue Exercise Endocrinology)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Typically, salivary cortisol is reported as 5–10% of total cortisol, but the stability of this proportion and the effect of exercise on the 24-h profile is unclear. Therefore, this study investigated the circadian rhythm of the proportion of serum cortisol represented by salivary cortisol, and the impact of acute high-intensity exercise. Recreationally trained males (n = 8, age = 25.7 ± 2.4 years, height = 174.7 ± 7.8 cm, mass = 69.8 ± 12.1 kg) completed two 24-h profiles (rest and exercise conditions) for serum (Q60) and salivary (Q120) cortisol. Exercise consisted of 5 × 30 s sprinting intervals on the cycle ergometer. Cortisol was assessed using commercially available assays. The proportion (Cprop) of serum cortisol (Cser) represented by salivary cortisol (Csal) was calculated as [Cprop = Csal/ Cser × 100]. Multilevel growth models tested for trends across the 24-h profile. The highest relation between Cser and Csal was observed at 08:00 AM (r = 0.90). The average Cprop was 5.95% and demonstrated a circadian profile characterized by a cubic model. Acute exercise did not alter Cser, Csal, or Cprop. Thus, the proportion of Cser represented by Csal changes across a 24-h period and should be accounted for if using salivary cortisol to reflect circadian output of cortisol.
1. Introduction
Cortisol is the predominate glucocorticoid in humans. Almost all human cells contain glucocorticoid receptors [1], and thus cortisol is considered a pleiotropic hormone, activating or inhibiting a range of genes dependent on numerous factors such as cell type, total exposure, length of exposure, and presence of cofactors. Cortisol demonstrates a robust circadian and diurnal profile in humans with peak concentrations observed in the morning soon after waking and the hormonal nadir occurring in the early morning period during nocturnal sleep [2,3].
The diurnal profile is controlled via self-oscillating transcriptional loops in the suprachiasmatic nucleus of the hypothalamus. These transcriptional/translation loops consist of positive arm proteins—CLOCK and BMAL1—that form a heterodimer and promote the synthesis of the negative arm proteins—Per 1/2/3 and Cry 1/2—which inhibit the binding of the positive arm proteins [4,5]. Auxiliary loop proteins, such as REV-Erb and ROR, also provide a further level of control and stability to this loop to regulate the 24-h rhythmicity [4,6]. The suprachiasmatic nucleus becomes entrained to the external environment primarily via light signals transmitted through the optic nerves. The primary function of the circadian variation in cortisol concentration has been theorized to be a neuroendocrine synchronization of peripheral clock mechanisms [5,7]. Disruption of this hypothalamic–pituitary–adrenal axis circadian rhythm has therefore been associated with a range of physical and psychological health disorders [5,8], and is of interest to both clinicians and researchers.
Cortisol is derived from cholesterol molecules and, thus, the hormone is lipophilic and circulates in the blood primarily bound to binding proteins. Approximately 80% of circulating cortisol is bound to the high-affinity, low-capacity protein corticosteroid binding globulin (CBG) [9]. Albumin, by contrast, has a high-capacity but low-affinity for cortisol [10], and binds approximately 10% of cortisol in circulation. The remaining approximately 10% of cortisol is considered to be “free” and according to the free-hormone hypothesis, is able to be taken up by target cells and exert intracellular effects [11]. Physiological stressors such as exercise can result in elevations in circulating cortisol concentrations [12], which can also be observed in the saliva [13]. For example, Hough and colleagues demonstrated an increase in salivary cortisol following a 30-min high-intensity cycling protocol [14]. There is also evidence to suggest that an afternoon exercise protocol can impact the nocturnal serum cortisol profile [15].
The use of saliva to measure cortisol has gained popularity due to its non-invasive nature and relative ease of collection. Salivary cortisol is significantly correlated with cortisol in circulation under resting conditions [16,17] and following stimulation of the hypothalamic–pituitary–adrenal axis via physiological challenges such as exercise [18,19,20,21,22,23]. The portion of cortisol that is considered free in the serum is able to readily pass into the salivary glands [24] and, thus, salivary cortisol reflects the free portion of the hormone [23].
In order to quantify the free portion of cortisol in the blood, first the free and bound portions must be separated through techniques such as equilibrium dialysis or ultrafiltration [25]. The fluid with only free cortisol remaining can then be analyzed using analytical techniques such as liquid chromatography and mass spectroscopy [26,27]. These techniques are time consuming, costly, and unavailable to many researchers or medical practitioners, and can also be impacted by analytic choices such as incubation temperature [28]. Salivary immunoassays therefore provide a useful alternative for assessing the so-called free, biologically active portion of the hormone.
As representative of the free portion, salivary cortisol is commonly reported in the literature as being 5–10% of circulating cortisol, although the range has been reported to be as wide as 1–9% [29]. This variance is due to a number of factors including the total concentration of both the hormone and binding proteins, as well as binding affinities of the binding proteins that can be modulated by other factors [30]. Salivary cortisol has been demonstrated to approximate the circadian profile of serum cortisol [31]. However, CBG also demonstrates a circadian rhythm [32], and thus it is likely that the unbound portion of cortisol also follows a circadian rhythm and disproportionality between CBG and unbound cortisol will impact the salivary portion of the hormone.
Although the range of 5–10% of total cortisol that is represented by salivary cortisol appears to be a small absolute range lending validity to its use as a non-invasive marker of total cortisol, this actually represents up to a two-fold difference in the total free cortisol concentration. For example, a salivary cortisol concentration of 5 nmol.l-1 may be associated with a circulating free cortisol concentration of 50–100 nmol.l-1. Within a short time window under stable conditions, this is perhaps only a small concern, but over longer periods of time or during changes in subject allostasis, such a discrepancy may lead to erroneous conclusions regarding circulating free cortisol concentrations.
To date, no study has investigated the circadian change in the proportion of total cortisol represented by salivary cortisol, or whether high-intensity exercise may alter such a circadian rhythm. Therefore, this study aimed to investigate the relation between salivary and serum cortisol across a 24-h period, whether there was a circadian rhythm to the proportion of the two biological fluids, and the impact of high-intensity exercise on the profile. We hypothesized that (1) the proportion of total cortisol represented by salivary cortisol would be stable across the 24-h period; (2) short duration high-intensity exercise would not impact this proportion across the 24-h period; and (3) salivary and serum cortisol would be significantly correlated across the entire 24-h period, with and without mid-morning short duration high-intensity exercise.
2. Materials and Methods
2.1. Participants
Adult males (n = 8, age = 25.7 ± 2.4 years, height = 174.7 ± 7.8 cm, mass = 69.8 ± 12.1 kg) were recruited to participate in this study. Participants were required to be 18–35 years old, self-report participation in regular moderate–vigorous exercise, have a body fat percent <18%, and be free of any known metabolic, cardiovascular, or pulmonary disease. All participants gave their written informed consent for inclusion before they participated in the study. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Institutional Review Board at the University of North Carolina at Greensboro (IRB #17-0287).
2.2. Procedure
Participants completed two separate sets of visits, separated by at least 8 weeks to permit full recovery and adequate time for the replenishment of blood volume (range 8–10 weeks). Within each set of visits, participants completed a screening visit and a profile visit. At the screening visit participants completed a battery of physiological measurements including body composition via air displacement plethysmography and a maximal cycle ergometer test where maximal oxygen uptake (VO2max) was determined by indirect calorimetry (TrueOne 2400, ParvoMedics, Sandy, UT, USA).
Within a week of these screening visits, participants reported to the laboratory again for the 24-h profile. Participants arrived at the laboratory at 05:30 AM and an intravenous catheter was placed in the forearm. As part of a larger study, starting at 06:00 AM blood samples were taken at 10 min intervals (Q10) for the next 24 h (144 samples) via syringe using stopcock valves in the i.v. tubing. A saline (0.9% NaCl) drip (60 mL/h) was used to maintain line and canula patency. Saliva samples were obtained via passive drool at 06:00 AM and every 2 h thereon for the entire 24-h period. Participants retired to the sleep room at 10:30 PM where no electronic devices or other light sources were permitted and were encouraged to sleep.
Participants completed both an exercise and a control profile visit, in a randomized order. Prior to each profile visit, participants were instructed to refrain from any exercise in the 24 h prior to the visit. Participants recorded food and fluid intake during the week between the screening and profile visit, and were asked to replicate this, to the best of their ability, at the second set of visits. At the control visit, participants were permitted to complete any sedentary activities during the waking hours as desired, including reading, watching movies, or talking. Participants were required to eat breakfast, lunch, and dinner at 07:30 AM, 01:00 PM, and 08:00 PM, respectively, and were restricted to water intake only from 08:00 AM to 10:00 AM to standardize macronutrient intake prior to exercise, due to the impact of carbohydrate availability on the cortisol response to exercise [33,34].
In the exercise condition, participants completed a high-intensity sprint protocol on an electromagnetically braked cycle ergometer (Lode Excalibur Sport; Lode BV, Groningen, the Netherlands), beginning at exactly 10:00 AM (immediately following the 10:00 AM blood and saliva samples). Participants warmed-up on the cycle ergometer by cycling at a self-selected pace for 5 min and completing 2–3 high-cadence 5–10 s bouts. Participants then completed 5 sets of 30 s sprints, separated by 2.5 min of active recovery. For each sprint, participants were required to build to a maximal cadence prior to the researcher applying a resistance to the ergometer flywheel equal to 7.5% of participant body mass. Participants were then required to maintain as high a cadence as possible for the 30 s period, while receiving verbal encouragement from the research staff. This cycling protocol elicited a supramaximal power output (mean power of all repetitions = 490.0 ± 93.8 W, 6.9 ± 0.8 W/kg). This protocol was chosen to stimulate a significant physiological response across the hypothalamic–pituitary axis.
Blood (Q60) and saliva (Q120) samples were assayed for cortisol using commercially available competitive binding ELISA assays (R&D Systems). For consistency, all samples for a single individual were assayed on a single plate. The inter-assay and intra-assay CV were 20.3% and 10.1%, respectively, which were both within reported manufacturer ranges.
2.3. Statistical Analytic Plan
Mixed-effect growth models with random intercepts were first used to characterize the profile of serum cortisol (Cser) and salivary cortisol (Csal) cortisol separately. Profiles were first modeled for stationary, linear, quadratic, and cubic trends and model fits were determined by Bayesian Information Criterion (BIC) and change in -2LL. Models were then further assessed for improvements in fit using a continuous autoregressive correlation structure. Following the determination of the best fitting profile model, profile condition (rest or exercise) was included as a fixed effect and improvement in model fit was determined. Next, a simple Pearson correlation between Cser and Csal was computed for each time point throughout the profile.
The proportion (Cprop) of serum cortisol (Cser) represented by salivary cortisol (Csal) was calculated as [Cprop = Csal/Cser × 100]. Cprop was assessed for normality and then log-transformed. The profile of Cprop was then assessed in the same manner as Cser and Csal. Following determination of the best fitting profile model, condition (rest or exercise) was included as a fixed effect. Cser, Csal, and Cprop were all centered at 10:00 AM (start of exercise) and alpha level was set at p < 0.05 for all statistical inference tests.
3. Results
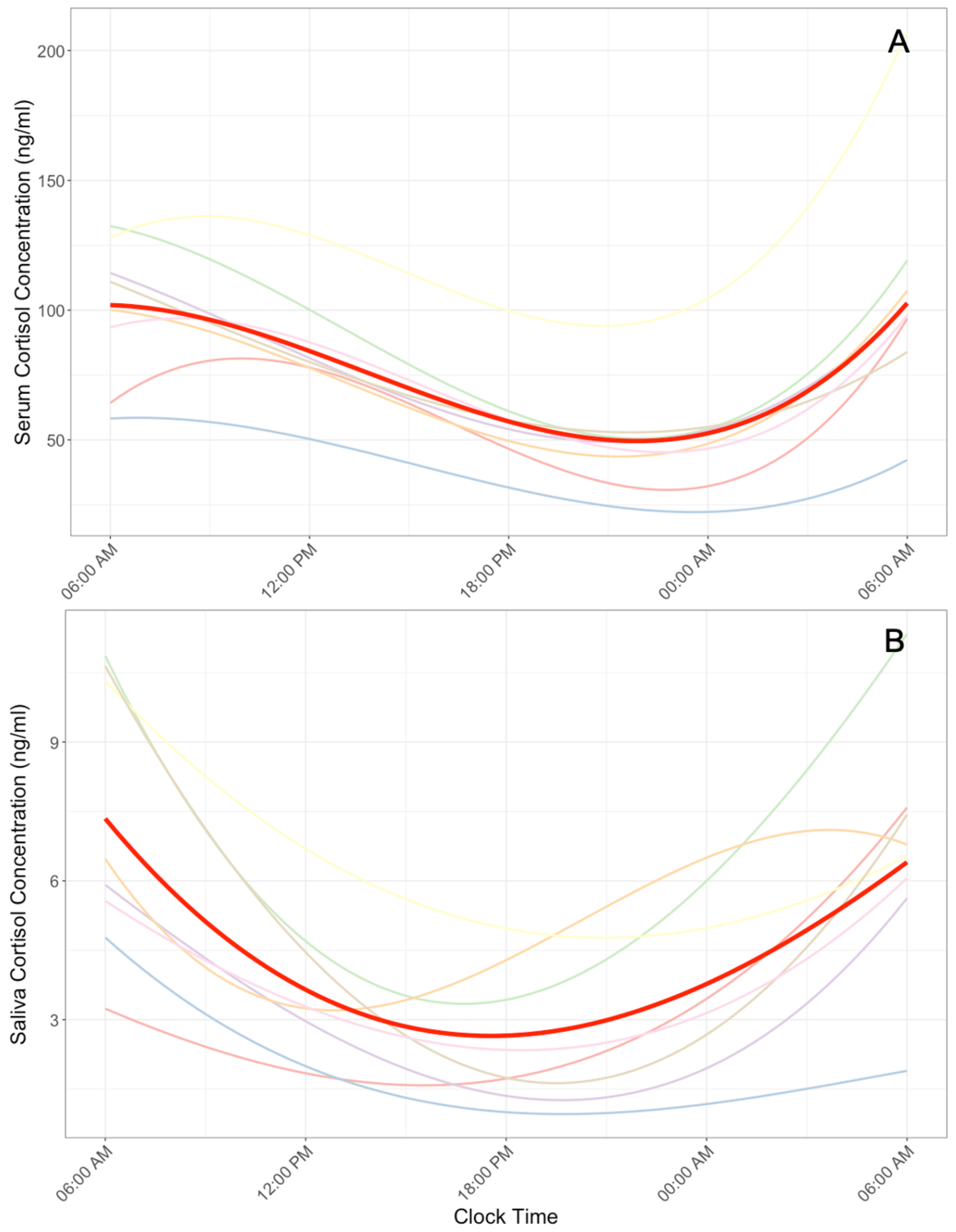
Cser profiles demonstrated significantly improved model fit with a cubic model with continuous autoregressive correlation structure (R2 = 0.30, Figure 1A). Because serum (Q60) and saliva (Q120) were sampled at different frequencies, in the post-hoc analysis serum was also modeled using only samples from the equivalent Q120 time points. These additional models also demonstrated the best fitting model was a cubic model with an autoregression correlation structure. Controlling for exercise condition did not significantly improve model fit (p = 0.26). Post-hoc analysis demonstrated that exercise condition also did not significantly interact with the linear (β = 7.2 × 10−3, p = 0.99), quadratic (β = 6.0 × 10−3, p = 0.91), or cubic (β = 3.5 × 10−4, p = 0.89) component of the Cser profile model.
The Csal profile demonstrated improved model fit only with the quadratic continuous autoregressive correlation structure (R2 = 0.28, Figure 1B). The addition of a cubic term did not significantly improve model fit (p = 0.71). Model fit did not significantly improve after controlling for exercise condition (p = 0.37). Post-hoc analysis also demonstrated that exercise condition did not significantly interact with either the linear (β = −3.2 × 10−2, p = 0.63) or quadratic (β = −3.4 × 10−2, p = 0.34) component of the Csal profile model.
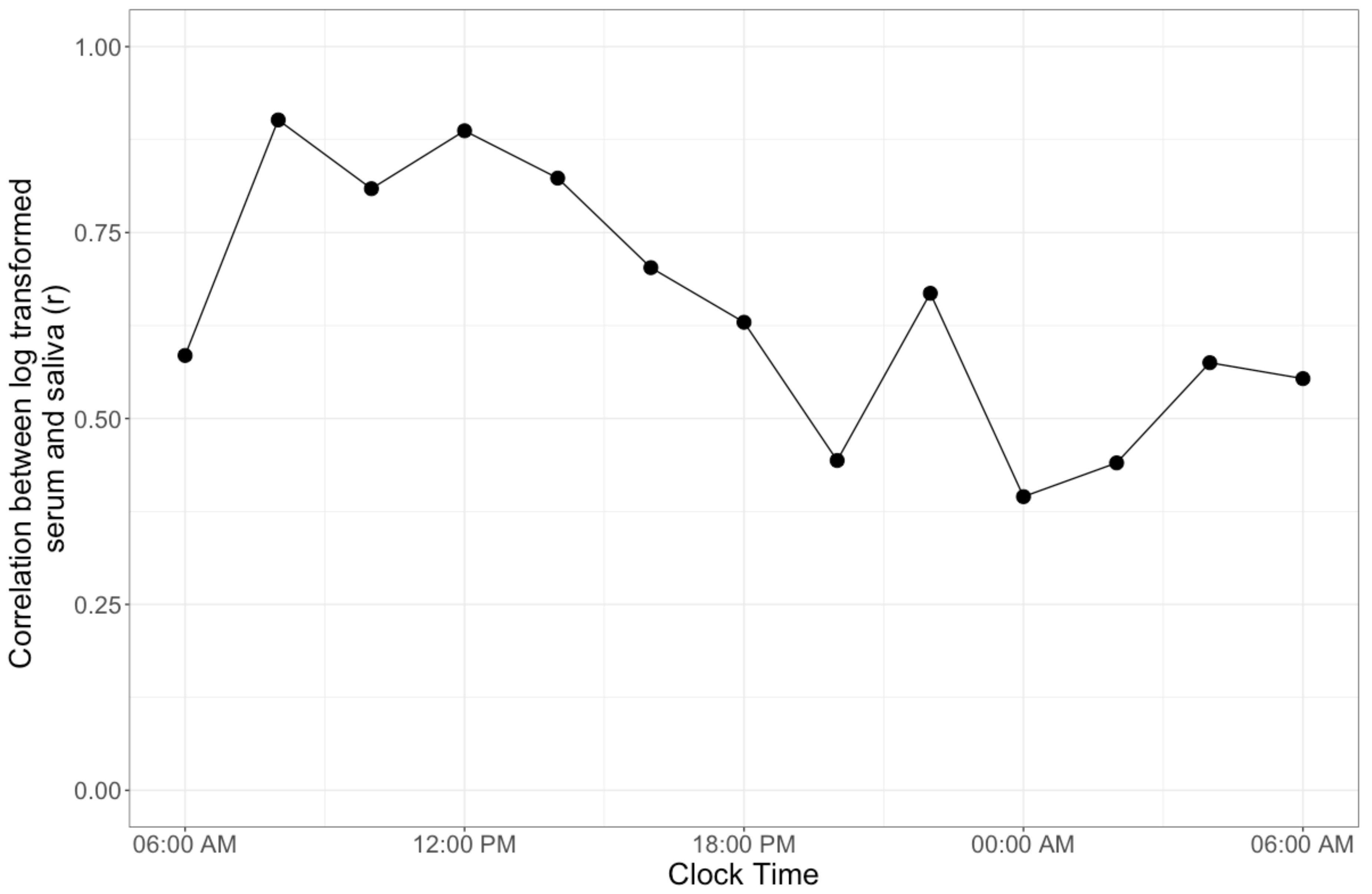
The mean (standard deviation) correlation between log transformed Cser and Csal was r = 0.65 (0.17), and was greatest at 08:00 AM (r = 0.90, Figure 2). All correlation coefficients except at 08:00 PM (r = 0.44, p = 0.09), 12:00 AM (r = 0.39, p = 0.15), and 02:00 AM (r = 0.44, p = 0.09) were significant (all others: p < 0.05).
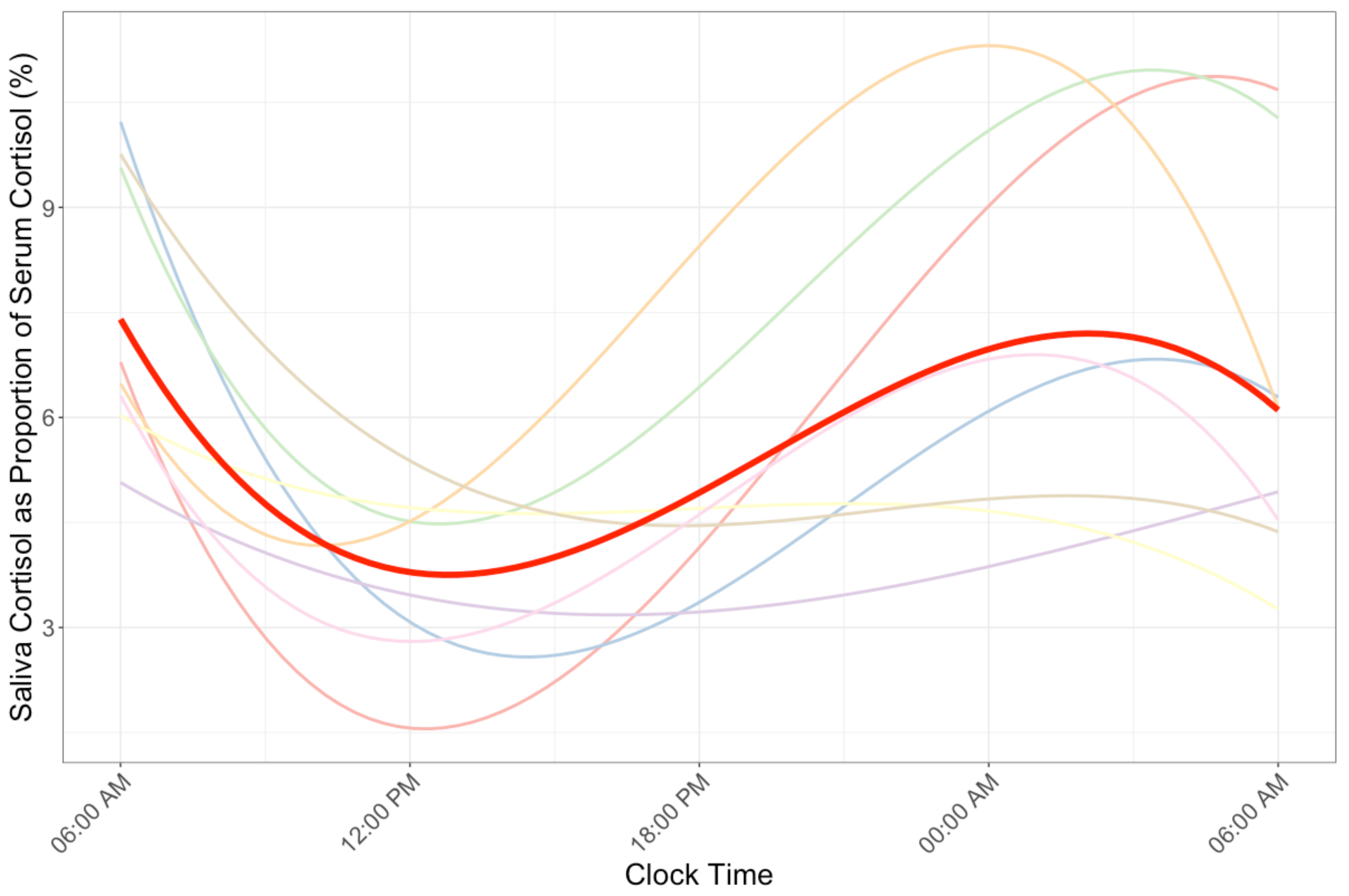
The grand mean of Cprop was 5.95% (group mean range: 3.93–8.93%). Cprop profiles demonstrated improved model fit with a cubic model with continuous autoregressive correlation structure (Conditional R2 = 0.17, Figure 3). Controlling for exercise condition did not significantly improve model fit (p = 0.91). Post-hoc analysis demonstrated that exercise condition did not significantly interact with the linear (β = −4.9 × 10−3, p = 0.65), quadratic (β = −2.2 × 10−4, p = 0.73), or cubic (β = −1.4 × 10−5, p = 0.67) component of the Cprop profile model.
4. Discussion
The purpose of this study was to assess the nature of any circadian rhythm to the proportion (Cprop) of serum total cortisol (Cser) as represented by salivary cortisol (Csal), and to investigate whether any such rhythm was altered by mid-morning high-intensity exercise. The present results demonstrate a circadian rhythm in Cprop that was best characterized by a cubic growth model and appears to follow the general trend of the circadian and diurnal profile of cortisol itself. Moreover, it appears that the variability in this proportion is significantly greater during the nocturnal period, which is also characterized by a reduction in the correlation between cortisol concentrations from these two biological fluids. Lastly, the results do not support the hypothesis that high-intensity exercise in the mid-morning period affects the circadian rhythm of Cser, Csal, or Cprop.
The free portion of cortisol is able to pass into the salivary gland and then the saliva, and thus it is believed that Csal represents the free portion of the total Cser. The circadian rhythm observed in Csal in the present study may therefore reasonably represent a circadian rhythm in the amount of free cortisol in circulation. A majority of cortisol is bound by CBG, which has a half-life of approximately 10 h [35]. Given this relatively static concentration of the binding protein and thus cortisol binding capacity, changes in the total cortisol concentration would lead to alterations in the free cortisol concentration. This may explain the reduction in the Cprop as total cortisol decreased across the diurnal period, and increases in Cprop when total cortisol concentrations in Cser are expected to be greatest. Of particular note, one participant had higher cortisol levels compared to other participants, but cortisol concentrations were higher in both serum and saliva, thus not affecting the analysis of Cprop for this individual.
It is also conceivable, however, that other factors that affect the free portion of cortisol beyond the total cortisol concentration contribute to the observed Cprop profile. There is some evidence to suggest that while concentrations of CBG do not vary substantially, there is a circadian rhythm to CBG concentrations [32], and thus the binding capacity fluctuates throughout a 24-h period. Further, CBG affinity for cortisol is known to be highly temperature sensitive [36]. Core body temperature also demonstrates a robust circadian rhythm that is integral to sleep onset [37]; given the tight thermocouple of cortisol and CBG, decreases in core body temperature during the nocturnal period may impact free cortisol and thus the Cprop. This in vitro temperature dependence of CBG should be investigated further both for understanding HPA-axis circadian rhythms and regulation, as well as potentially being an elegant local targeting mechanism [30], particularly during exercise. The apparent mismatch between serum total and salivary free cortisol that generates the cubic model across time for Cprop suggests that, for at least some portion(s) of the circadian profile, the cross-correlation between these measures should be greatest at a lag >1 unit. In fact, researchers recently reported a maximal cross-correlation between serum cortisol and subcutaneous free cortisol of five units (50 min) [38]. We therefore completed post-hoc analyses in which we computed the cross-correlation of the differenced log transformed serum and saliva values across the 24-h period. This analysis demonstrated a maximal correlation at a 4-h lag, possibly contributing to the profile in Cprop. However, this should be investigated further in saliva using more frequent sampling procedures (e.g., Q30) or via the promising microanalysis technique for ambulatory free cortisol monitoring [39].
Salivary glands may also exhibit circadian properties that modify the Cprop value over time. For example, salivary glands have a high concentration of 11β-HSD2, an enzyme that readily converts cortisol to cortisone [40]. The assay used in the present study reports a cross-reactivity with cortisone of only 0.2%, and thus it is possible that the large variance in Cprop observed in the early morning period was due to differences in the conversion of cortisol to cortisone. Indeed, in one of only several studies of circadian relations between serum and saliva steroids, a higher correlation was reported to salivary cortisone than salivary cortisol [39]. Although cortisol does not appear to be flow dependent and cross-correlations have demonstrated a similar lag (e.g., ~10 min) between serum adrenocorticotropin hormone and Cser and Csal [41], saliva production is markedly reduced during the evening period [42], providing another potential confounding factor. In sum, due to these considerations, Csal may not always accurately represent free cortisol in circulation across the circadian profile.
The lack of effect of this high-intensity exercise protocol may be a result of several factors. First, and most importantly, the exercise session was likely not of sufficient duration to elicit an acute cortisol response, and thus has minimal effect on the post-exercise period. Although the intensity of the activity was supramaximal relative to the power output achieved during the VO2max test, the participants only engaged in this activity for a total of 2.5 min, with an additional 18 min of low-intensity active recovery between these efforts. Cortisol has a known intensity [43] and duration [44] threshold for acute responses. Despite the intensity of the exercise meeting this threshold, the total work of the exercise session was not likely great enough to cause sufficient metabolic and thermoregulatory disturbances to dramatically increase cortisol output. Secondly, the circadian rhythm of the HPA-axis and cortisol specifically is robust and plays a role in the circadian regulation of a variety of other cells and systems [45]. It is possible that such a robust circadian profile requires a greater perturbation than the exercise session employed here. Lastly, the effect of the exercise employed in this specific study on the cortisol dynamics may be small and/or variable, and was therefore unable to be statistically detected with a relatively small sample size.
Although this study provides novel insights into the utility of salivary cortisol and its interpretation, there are several limitations to acknowledge. As discussed, the sample size was small (n = 8), and thus the present analyses may be underpowered. Notably however, since this is a repeated measures, randomized control study, it is important to highlight that each participant contributed 48 blood samples and 24 saliva samples across two 24 h periods to these analyses, underscoring the logistical and financial challenge of conducting research of this kind. Larger studies with more frequent sampling should be conducted in the future while understanding these challenges. Secondly, participants were awoken to provide the saliva samples during the night sample collection (the blood draws were completed with care to not wake the participant). Disrupting the sleep cycle in this manner was a methodological choice that may have perturbed the regular HPA-axis activity. There is a need therefore to devise saliva collection devices that can be used without disrupting regular sleep. Lastly, the exercise protocol employed, whilst of a maximal intensity in the work interval, likely did not elicit a robust enough metabolic or thermoregulatory challenge to the participants to disrupt the underlying circadian cortisol profile. Attempts should be made to replicate these findings while employing an exercise protocol specifically structured to maximize the acute cortisol response to exercise. Future studies should also analyze salivary cortisone in addition to Cser and Csal, as an indirect measure of 11β-HSD2 activity across the circadian profile.
5. Conclusions
These results demonstrate that the collection of saliva for the quantification of cortisol is a useful, non-invasive method, but has essential considerations if being used to describe the circadian rhythm of the HPA-axis. Many researchers are aware of the need to standardize the time of day for sample collection to account for the circadian rhythm of cortisol. However, it appears that change in salivary cortisol does not linearly scale across a 24-h period with total circulating cortisol, and so practitioners and researchers should also control for the time of day when making inferences to the total cortisol concentration. Finally, saliva collection via passive drool may not be an appropriate fluid for examining cortisol during sleep periods due to its low correlation with total circulating cortisol.
Author Contributions
Conceptualization, T.A., N.T.B. and L.W.; data curation, N.T.B.; formal analysis, T.A. and N.T.B.; funding acquisition, L.W.; investigation, T.A., N.T.B. and L.W.; methodology, N.T.B. and L.W.; project administration, N.T.B.; resources, N.T.B. and L.W.; supervision, L.W.; visualization, T.A. and N.T.B.; writing—original draft, T.A.; writing—review and editing, T.A., N.T.B. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board of the University of North Carolina at Greensboro (protocol code 17-0287 date: 2017-07-05).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Kloet, E.R.; Van Acker, S.A.; Sibug, R.M.; Oitzl, M.S.; Meijer, O.C.; Rahmouni, K.; de Jong, W. Brain Mineralocorticoid Receptors and Centrally Regulated Functions. Kidney Int. 2000, 57, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Hellman, L.; Nakada, F.; Curti, J.; Weitzman, E.D.; Kream, J.; Roffwarg, H.; Ellman, S.; Fukushima, D.K.; Gallagher, T.F. Cortisol Is Secreted Episodically by Normal Man. J. Clin. Endocrinol. Metab. 1970, 30, 411–422. [Google Scholar] [CrossRef]
- Weitzman, E.D.; Fukushima, D.; Nogeire, C.; Roffwarg, H.; Gallagher, T.F.; Hellman, L. Twenty-Four Hour Pattern of the Episodic Secretion of Cortisol in Normal Subjects. J. Clin. Endocrinol. Metab. 1971, 33, 14–22. [Google Scholar] [CrossRef]
- Nader, N.; Chrousos, G.P.; Kino, T. Interactions of the Circadian CLOCK System and the HPA Axis. Trends Endocrinol. Metab. 2010, 21, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; de Kloet, E.R.; Dijk, D.-J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids. Endocr. Rev. 2016, 38, 3–45. [Google Scholar] [CrossRef]
- Andreani, T.S.; Itoh, T.Q.; Yildirim, E.; Hwangbo, D.-S.; Allada, R. Genetics of Circadian Rhythms. Sleep Med. Clin. 2015, 10, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Spencer, R.L.; Chun, L.E.; Hartsock, M.J.; Woodruff, E.R. Glucocorticoid Hormones Are Both a Major Circadian Signal and Major Stress Signal: How This Shared Signal Contributes to a Dynamic Relationship between the Circadian and Stress Systems. Front. Neuroendocr. 2018, 49, 52–71. [Google Scholar] [CrossRef]
- Adam, E.K.; Quinn, M.E.; Tavernier, R.; McQuillan, M.T.; Dahlke, K.A.; Gilbert, K.E. Diurnal Cortisol Slopes and Mental and Physical Health Outcomes: A Systematic Review and Meta-Analysis. Psychoneuroendocrinology 2017, 83, 25–41. [Google Scholar] [CrossRef]
- Dunn, J.F.; Nisula, B.C.; Rodbard, D. Transport of Steroid Hormones: Binding of 21 Endogenous Steroids to Both Testosterone-Binding Globulin and Corticosteroid-Binding Globulin in Human Plasma. J. Clin. Endocrinol. Metab. 1981, 53, 58–68. [Google Scholar] [CrossRef]
- Levine, A.; Zagoory-Sharon, O.; Feldman, R.; Lewis, J.G.; Weller, A. Measuring Cortisol in Human Psychobiological Studies. Physiol. Behav. 2007, 90, 43–53. [Google Scholar] [CrossRef]
- Mendel, C.M. The Free Hormone Hypothesis: A Physiologically Based Mathematical Model. Endocr. Rev. 1989, 10, 232–274. [Google Scholar] [CrossRef]
- Hackney, A.C. Stress and the Neuroendocrine System: The Role of Exercise as a Stressor and Modifier of Stress. Expert Rev. Endocrinol. Metab. 2006, 1, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBruggen, M.D.; Hackney, A.C.; McMurray, R.G.; Ondrak, K.S. The Relationship between Serum and Salivary Cortisol Levels in Response to Different Intensities of Exercise. Int. J. Sports Physiol. Perform. 2011, 6, 396–407. [Google Scholar] [CrossRef] [Green Version]
- Hough, J.; Corney, R.; Kouris, A.; Gleeson, M. Salivary Cortisol and Testosterone Responses to High-Intensity Cycling before and after an 11-Day Intensified Training Period. J. Sports Sci. 2013, 31, 1614–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackney, A.C.; Ness, R.J.; Schrieber, A. Effects of Endurance Exercise on Nocturnal Hormone Concentrations in Males. Chronobiol. Int. 1989, 6, 341–346. [Google Scholar] [CrossRef]
- Aardal, E.; Holm, A.-C. Cortisol in Saliva-Reference Ranges and Relation to Cortisol in Serum. Eur. J. Clin. Chem. Clin. Biochem. 1995, 33, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Reid, J.D.; Intrieri, R.C.; Susman, E.J.; Beard, J.L. The Relationship of Serum and Salivary Cortisol in a Sample of Healthy Elderly. J. Gerontol. 1992, 47, P176–P179. [Google Scholar] [CrossRef]
- Cadore, E.; Lhullier, F.; Brentano, M.; Silva, E.; Ambrosini, M.; Spinelli, R.; Silva, R.; Kruel, L. Correlations between Serum and Salivary Hormonal Concentrations in Response to Resistance Exercise. J. Sports Sci. 2008, 26, 1067–1072. [Google Scholar] [CrossRef]
- Gozansky, W.; Lynn, J.; Laudenslager, M.; Kohrt, W. Salivary Cortisol Determined by Enzyme Immunoassay Is Preferable to Serum Total Cortisol for Assessment of Dynamic Hypothalamic–Pituitary–Adrenal Axis Activity. Clin. Endocrinol. 2005, 63, 336–341. [Google Scholar] [CrossRef]
- O’Connor, P.J.; Corrigan, D.L. Influence of Short-Term Cycling on Salivary Cortisol Levels. Med. Sci. Sports Exerc. 1987, 19, 224–228. [Google Scholar]
- Paccotti, P.; Minetto, M.; Terzolo, M.; Ventura, M.; Ganzit, G.; Borrione, P.; Termine, A.; Angeli, A. Effects of High-Intensity Isokinetic Exercise on Salivary Cortisol in Athletes with Different Training Schedules: Relationships to Serum Cortisol and Lactate. Int. J. Sports Med. 2005, 26, 747–755. [Google Scholar] [CrossRef]
- Stupnicki, R.; Obminski, Z. Glucocorticoid Response to Exercise as Measured by Serum and Salivary Cortisol. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 65, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Vining, R.F.; McGinley, R.A.; Maksvytis, J.J.; Ho, K.Y. Salivary Cortisol: A Better Measure of Adrenal Cortical Function than Serum Cortisol. Ann. Clin. Biochem. 1983, 20, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Umeda, T.; Hiramatsu, R.; Iwaoka, T.; Shimada, T.; Miura, F.; Sato, T. Use of Saliva for Monitoring Unbound Free Cortisol Levels in Serum. Clin. Chim. Acta 1981, 110, 245–253. [Google Scholar] [PubMed]
- Vogeser, M.; Möhnle, P.; Briegel, J. Free Serum Cortisol: Quantification Applying Equilibrium Dialysis or Ultrafiltration and an Automated Immunoassay System. Clin. Chem. Lab. Med. 2007, 45, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, C.J.; Galligan, J.P.; McWhinney, B.C.; Briscoe, S.E.; Ungerer, J.P. Free Cortisol Method Comparison: Ultrafiltation, Equilibrium Dialysis, Tracer Dilution, Tandem Mass Spectrometry and Calculated Free Cortisol. Clin. Chim. Acta 2011, 412, 1043–1047. [Google Scholar] [CrossRef]
- Kirchhoff, F.; Briegel, J.; Vogeser, M. Quantification of Free Serum Cortisol Based on Equilibrium Dialysis and Isotope Dilution-Liquid Chromatography–Tandem Mass Spectrometry. Clin. Biochem. 2011, 44, 894–899. [Google Scholar] [CrossRef]
- Vogeser, M.; Briegel, J. Effect of Temperature on Protein Binding of Cortisol. Clin. Biochem. 2007, 40, 724–727. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Hellhammer, D.H. Salivary Cortisol in Psychoneuroendocrine Research: Recent Developments and Applications. Psychoneuroendocrinology 1994, 19, 313–333. [Google Scholar] [CrossRef]
- Henley, D.; Lightman, S. New Insights into Corticosteroid-Binding Globulin and Glucocorticoid Delivery. Neuroscience 2011, 180, 1–8. [Google Scholar] [CrossRef]
- Dorn, L.D.; Lucke, J.F.; Loucks, T.L.; Berga, S.L. Salivary Cortisol Reflects Serum Cortisol: Analysis of Circadian Profiles. Ann. Clin. Biochem. 2007, 44, 281–284. [Google Scholar] [CrossRef]
- Lewis, J.; Möpert, B.; Shand, B.; Doogue, M.; Soule, S.; Frampton, C.; Elder, P. Plasma Variation of Corticosteroid-Binding Globulin and Sex Hormone-Binding Globulin. Horm. Metab. Res. 2006, 38, 241–245. [Google Scholar] [CrossRef]
- Bishop, N.C.; Gleeson, M.; Nicholas, C.W.; Ali, A. Influence of Carbohydrate Supplementation on Plasma Cytokine and Neutrophil Degranulation Responses to High Intensity Intermittent Exercise. Int. J. Sport Nutr. Exerc. Metab. 2002, 12, 145–156. [Google Scholar] [CrossRef]
- Galbo, H.; Holst, J.; Christensen, N. The Effect of Different Diets and of Insulin on the Hormonal Response to Prolonged Exercise. Acta Physiol. Scand. 1979, 107, 19–32. [Google Scholar] [CrossRef]
- Lewis, J.G.; Saunders, K.; Dyer, A.; Elder, P.A. The Half-Lives of Intact and Elastase Cleaved Human Corticosteroid-Binding Globulin (CBG) Are Identical in the Rabbit. J. Steroid Biochem. Mol. Biol. 2015, 149, 53–57. [Google Scholar] [CrossRef]
- Cameron, A.; Henley, D.; Carrell, R.; Zhou, A.; Clarke, A.; Lightman, S. Temperature-Responsive Release of Cortisol from Its Binding Globulin: A Protein Thermocouple. J. Clin. Endocrinol. Metab. 2010, 95, 4689–4695. [Google Scholar] [CrossRef]
- Kräuchi, K.; Wirz-Justice, A. Circadian Clues to Sleep Onset Mechanisms. Neuropsychopharmacology 2001, 25, S92–S96. [Google Scholar] [CrossRef] [Green Version]
- Bhake, R.; Kluckner, V.; Stassen, H.; Russell, G.; Leendertz, J.; Stevens, K.; Linthorst, A.; Lightman, S. Continuous Free Cortisol Profiles—Circadian Rhythms in Healthy Men. J. Clin. Endocrinol. Metab. 2019, 104, 5935–5947. [Google Scholar] [CrossRef]
- Bhake, R.; Russell, G.M.; Kershaw, Y.; Stevens, K.; Zaccardi, F.; Warburton, V.E.; Linthorst, A.C.; Lightman, S.L. Continuous Free Cortisol Profiles in Healthy Men: Validation of Microdialysis Method. J. Clin. Endocrinol. Metab. 2020, 105, e1749–e1761. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.; Holmes, M.; Seckl, J. 11β-Hydroxysteroid Dehydrogenases: Intracellular Gate-Keepers of Tissue Glucocorticoid Action. Physiol. Rev. 2013, 93, 1139–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlotz, W.; Kumsta, R.; Layes, I.; Entringer, S.; Jones, A.; Wüst, S. Covariance between Psychological and Endocrine Responses to Pharmacological Challenge and Psychosocial Stress: A Question of Timing. Psychosom. Med. 2008, 70, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Dawes, C. Circadian Rhythms in Human Salivary Flow Rate and Composition. J. Physiol. 1972, 220, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.; Zack, E.; Battaglini, C.; Viru, M.; Viru, A.; Hackney, A. Exercise and Circulating Cortisol Levels: The Intensity Threshold Effect. J. Endocrinol. Investig. 2008, 31, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Duclos, M.; Corcuff, J.; Rashedi, M.; Fougere, V.; Manier, G. Trained versus Untrained: Different Hypothalamo-Pituitary Adrenal Axis Responses to Exercise Recovery. Eur. J. Appl. Physiol. 1997, 75, 343–350. [Google Scholar] [CrossRef]
- Nicolaides, N.C.; Charmandari, E.; Kino, T.; Chrousos, G.P. Stress-Related and Circadian Secretion and Target Tissue Actions of Glucocorticoids: Impact on Health. Front. Endocrinol. 2017, 8, 70. [Google Scholar] [CrossRef]
Figure 1.
Serum (A) and saliva (B) cortisol profiles across 24-h. The serum and saliva profiles were best characterized via a cubic and quadratic model, respectively. Mean profiles are depicted in red.
Figure 1.
Serum (A) and saliva (B) cortisol profiles across 24-h. The serum and saliva profiles were best characterized via a cubic and quadratic model, respectively. Mean profiles are depicted in red.

Figure 2.
The correlation of log transformed saliva and serum cortisol across the 24-h profile. The correlation was greatest at 0800 (r = 0.90) but demonstrated a general decrease across the remainder of the profile.
Figure 2.
The correlation of log transformed saliva and serum cortisol across the 24-h profile. The correlation was greatest at 0800 (r = 0.90) but demonstrated a general decrease across the remainder of the profile.

Figure 3.
Saliva cortisol as a proportion of serum cortisol across the 24-h profile. The best fitting model demonstrated a cubic circadian rhythm. The mean profile is depicted in red.
Figure 3.
Saliva cortisol as a proportion of serum cortisol across the 24-h profile. The best fitting model demonstrated a cubic circadian rhythm. The mean profile is depicted in red.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Anderson, T.; Berry, N.T.; Wideman, L. The Effect of High-Intensity Exercise on Changes in Salivary and Serum Cortisol Proportion Dynamics. Endocrines 2021, 2, 44-53. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2010005
AMA Style
Anderson T, Berry NT, Wideman L. The Effect of High-Intensity Exercise on Changes in Salivary and Serum Cortisol Proportion Dynamics. Endocrines. 2021; 2(1):44-53. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2010005
Chicago/Turabian StyleAnderson, Travis, Nathaniel T. Berry, and Laurie Wideman. 2021. "The Effect of High-Intensity Exercise on Changes in Salivary and Serum Cortisol Proportion Dynamics" Endocrines 2, no. 1: 44-53. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2010005