Contemporary Distribution, Estimated Age, and Prehistoric Migrations of Old World Monkey Retroviruses


Laboratory of Experimental Virology, Department of Medical Microbiology, Amsterdam UMC, University of Amsterdam, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands
Epidemiologia 2021, 2(1), 46-67; https://0-doi-org.brum.beds.ac.uk/10.3390/epidemiologia2010005
Submission received: 15 December 2020
/
Revised: 18 January 2021
/
Accepted: 29 January 2021
/
Published: 3 February 2021
Abstract
:Old World monkeys (OWM), simians inhabiting Africa and Asia, are currently affected by at least four infectious retroviruses, namely, simian foamy virus (SFV), simian immunodeficiency virus (SIV), simian T-lymphotropic virus (STLV), and simian type D retrovirus (SRV). OWM also show chromosomal evidence of having been infected in the past with four more retroviral species, baboon endogenous virus (BaEV), Papio cynocephalus endogenous virus (PcEV), simian endogenous retrovirus (SERV), and Rhesus endogenous retrovirus-K (RhERV-K/SERV-K1). For some of the viruses, transmission to other primates still occurs, resulting, for instance, in the HIV pandemic. Retroviruses are intimately connected with their host as they are normally spread by close contact. In this review, an attempt to reconstruct the distribution and history of OWM retroviruses will be made. A literature overview of the species infected by any of the eight retroviruses as well as an age estimation of the pathogens will be given. In addition, primate genomes from databases have been re-analyzed for the presence of endogenous retrovirus integrations. Results suggest that some of the oldest retroviruses, SERV and PcEV, have travelled with their hosts to Asia during the Miocene, when a higher global temperature allowed simian expansions. In contrast, younger viruses, such as SIV and SRV, probably due to the lack of a primate continuum between the continents in later times, have been restricted to Africa and Asia, respectively.
Keywords:
retrovirus; endogenous; exogenous; Old World monkey; BaEV; PcEV; SERV; SERV-K1; RhERV-K; SFV; SIV; SRV; STLV1. Introduction
The primate fossil record suggests that the radiation of Old World monkeys (OWM) began in the Oligocene or early Miocene (around 25–30 million years ago (mya)), with the divergence of apes, including the ancestors of humans, and OWM (reviewed in [1], see also [2,3,4]). Subsequently, Colobinae and Cercopithecinae diverged around 16 mya, Papionini and Cercopithecini then split around 10–12 mya, and macaques and papionins separated approximately 7.6 ± 1.3 mya [1]. Over the course of evolution, retroviruses have been infecting primate species. Evidence for this can be found in primate genomes, where endogenous (i.e., germ line propagated) proviruses bear witness of past infections. The origin of the retrovirus family itself dates back to the early Paleozoic, approximately 460–550 mya [5]. Most likely, retroviruses originate from the long terminal repeat (LTR)-containing retrotransposons found in eukaryotic genomes, by the addition of an envelope (env) gene [6,7]. Indeed, one such LTR-retroelement, HERV-L, has similarity to foamy viruses in its polymerase (pol) gene [8].
At present, two retrovirus species infect humans, namely the lentivirus human immunodeficiency virus (HIV) and the deltaretrovirus human T-lymphotropic virus (HTLV). HIV has been transmitted from African non-human primates quite recently, probably around the beginning of the 20th century [9,10,11]. So far, five independent transmissions have been recorded, from chimpanzee and gorilla (HIV-1 group M, N, O, and P), and from sooty mangabey (HIV-2), respectively [12,13,14,15,16,17]. There is, however, evidence that all eight HIV-2 subtypes in fact each represent independent transmissions of simian immunodeficiency virus from sooty mangabeys (SIVsm) [18,19,20], increasing the number of SIV transmission events considerably. Strains of HTLV have also been spreading from monkeys and apes to humans in the recent past, likely between 3000 and 378,000 years ago, depending on the particular virus type [21,22,23,24,25,26,27,28]. Moreover, transmissions of the virus are ongoing [29,30]. A third retrovirus, simian foamy virus (SFV), is likewise known for its ongoing, zoonotic transmissions from non-human primates to humans (reviewed in [31,32,33]), though a true human variant has not emerged yet. A further retrovirus, simian retrovirus (SRV), initially seen as the cause of simian AIDS in primate facilities, is largely an endemic pathogen of Asian macaques [34,35]. Serological evidence for incidental SRV infection in persons occupationally exposed to non-human primates has been reported [36].
For the existence of other simian retroviruses, namely, baboon endogenous virus (BaEV), Papio cynocephalus endogenous virus (PcEV), simian endogenous retrovirus (SERV) and rhesus endogenous retrovirus (RhERV-K/SERV-K1), all evidence we have comes from the “fossil” record. These viruses have disappeared from circulation but have left their proviral genomes in OWM chromosomes. All four appear to be OWM specific, as their sequences have not been found in hominoid genomes, and neither have novel infections in humans or apes been documented. Summarizing, the eight retroviral species listed here spread among African primates, belong to different retroviral families, and have various tendencies to target the germ-cell line. The exogenous primate retroviruses can all be transmitted to humans. This review and hypothesis paper will discuss, partly on the basis of new research, the epidemiology (who, where, and when?) and putative age of primate retroviruses from the Oligocene till present, including both the extant and the extinct members of the family, to deduce probable OWM retrovirus dispersal routes.
2. Materials and Methods
2.1. Survey Methodology
The PubMed database (www.ncbi.nlm.nih.gov/pubmed) and Google Scholar (scholar.google.nl) were manually searched using the name of the virus ”simian immunodeficiency virus”, “simian foamy virus”, “simian T-lymphotropic virus”, “Papio cynocephalus endogenous virus”, “simian endogenous retrovirus”, “baboon endogenous virus”, or their abbreviation (“SIV”, “SFV”, “STLV”, “BaEV”, “PcEV”, “RhERV”, “SERV”, respectively) and a second term appropriate to the topic, such as “age”, “prevalence”, “phylogeny”, “evolution”, “dating”, “presence”, “detection”, “monkey”, etc. Abstracts were inspected for relevance; from the selected abstracts, only those of which a full-length publication could be retrieved were read and included. In addition, references in the retrieved papers were inspected and used when appropriate.
2.2. Identifying Endogenous Retrovirus Sequences in OWM Genomes
PcEV (GenBank acc. no. AF142988 [37]) and SERV 23.1 (GenBank acc. no. U85505; [38]), both full-length proviruses obtained from a Papio cynocephalus (yellow baboon) chromosomal DNA library, were used to identify endogenous PcEV and SERV nucleotide sequences in OWM genomes from GenBank (www.ncbi.nlm.nih.gov/genome/) and Ensembl (www.ensembl.org/index.html) databases by similarity search using the BLASTn/BLAT algorithm provided. In addition, BaEV, another full-length provirus (GenBank acc. no. D10032; [39]), was used to search OWM genomes for BaEV integrations. CERV1 (GenBank acc. no. AY692036) and CERV2 (GenBank acc. no. AY692037) to search for CERV-like gammaretroviral sequences; SERV-K1 (GenBank acc. no. BK009405) and HERV-K5 (GenBank acc. no. DQ112093) were used to query OWM genomes for ERV-K family members.
Default settings were used in the NCBI BLASTn search, as the aim of the research was to retrieve and describe specifically PcEV, SERV and BaEV integrations, and not more distantly related endogenous viruses, searches were optimized for “highly similar sequences” (megablast). Only when no results were retrieved with megablast, the discontiguous megablast option (“more dissimilar sequences”) was used. In Ensembl BLAST searches, search sensitivity was the default (“normal”) and filtering low complexity regions or filtering query sequences with RepeatMasker was disabled. When no full-length proviruses were retrieved, virus presence was defined by detecting fragments with >90% homology to both coding and non-coding (long terminal repeat, LTR) fragments of the specific virus. Primate genomes queried were: Cercocebus atys, Cercopithecus mona, Cercopithecus neglectus, Chlorocebus sabaeus (formerly known as Cercopithecus aethiops sabaeus), Colobus angolensis, Erythrocebus patas, Macaca fascicularis, Macaca fuscata, Macaca mulatta, Macaca nemestrina, Mandrillus leucophaeus, Mandrillus sphinx, Papio anubis, Piliocolobus tephrosceles, Rhinopithecus bieti, Rhinopithecus roxellana, Theropithecus gelada, and Trachypithecus francoisi.
2.3. Analysis of Proviral Sequences
BLAST results were downloaded from the databases and aligned using ClustalW as implemented in BioEdit (bioedit.software.informer.com). Alignments were optimized through visual inspection. Sequence distances were calculated with the Kimura-2-parameter method and evolutionary relationships were inferred using the Neighbor-joining method with bootstrapping as implemented in MEGA6 [40,41]. Gaps/missing data treatment was set to “partial deletion” with a cut-off value of 80%. No outgroup was defined. Alignments are available as Supplementary Files 1 and 2.
3. Results
3.1. Exogenous Retroviruses in OWM Species
Four retrovirus species are currently circulating in OWM, namely SFV, SIV, SRV, and STLV. SFV is also widespread in apes and New World monkeys (NWM). STLV is present in both Asian and African monkey species and in apes plus humans, while SIV is solely found in African primate species (including African apes), and recently, in humans (Table 1).
Several limitations apply when reconstructing the history of exogenous retroviruses from epidemiological data:
- Sampling is not systematic; some species remain untested because they may be difficult to reach.
- Sampling may not be optimal: it is done at the wrong age (for instance SIV is a sexually transmitted infection with juveniles normally being negative for the virus), the wrong type of sample is taken, or the viral load is below the detection level.
- The distinction between an exogenous and endogenous retrovirus is not always clear (e.g., murine leukemia virus, MuLV, feline leukemia virus, FeLV, and koala retrovirus, KoRV, have both infectious and endogenous variants). Moreover, every type of retrovirus has the capacity to enter the mammalian germ line, so Mendelian inheritance is not a distinguishing characteristic [42,43,44].
3.1.1. Simian Foamy Virus (SFV)
Spumaviruses, also known as foamy viruses due to the effect they induce in cell cultures, are a widely spread, distinct type of retrovirus, which share some similarities with hepadnaviruses with regard to their replication cycle [45]. Infectious spumaviruses are found in OWM, NWM and apes, where they induce little pathology, as replication only takes place in epithelial cells of the oral cavity [45,46,47]. The detection of endogenous FV-like sequences in a wide variety of amphibians, fish and mammals suggests a considerable history for the group and indeed, phylogenetic analysis infers FVs as the oldest retrovirus lineage, emerging >450 mya [48]. The origin of prosimian FV was calculated to the Mesozoic (~82.5 mya). The divergence between OWM and ape SFV was dated to the Oligocene (~30 mya), with the OWM node emerging ~16 to 18 mya [49]. Species-specific FV clades are more recent; for instance, orangutan genus and subspecies-specific clades have been dated to the late Pliocene (>4.7 mya), and the Pleistocene (>1.7 mya), respectively [50]. Cross-species transmissions and recombination events, however, complicate the reconstruction of spumavirus history [51,52].
An overview of the estimated contemporary distribution of OWM retroviruses and the deduced prehistoric virus migration routes, is given in Figure 1.
3.1.2. Simian Immunodeficiency Virus (SIV)
SIV infection has been detected in African, but not Asian primates or NWM, suggesting that origin and spread of the virus occurred after the separation of OWM and NWM, and also after the major migrations of African primates to Asia in the Miocene period.
Two independent integrations of an endogenous lentivirus, termed pSIV, with a phylogenetic position in between the feline lentivirus feline immunodeficiency virus (FIV) cluster and modern simian SIV strains, have been detected in the genome of Malagasy lemur species, suggesting that SIV precursors infecting prosimian primates already existed ~4.3 mya [83,84,85,86]. Since Madagascar and Africa have been separated for at least 160 mya, transitional pSIV was somehow transmitted across a substantial body of water, indicating that lentivirus dispersal does not always need a land bridge [83].
SIV is remarkably widespread, having infected over 40 African primate species and at least 9 genera (Table 1). Prevalence in populations can be high. For instance, up to 80% of wild Chlorocebus females test positive for SIVagm infection [87,88]. SIV strains exhibit relatively large nucleotide diversity, suggestive of great age [87,89]. Initial calculations, however, indicated that SIV lineages evolved in historic times, on a time scale of only tens or hundreds of years [90,91]. Subsequent analysis of SIV isolates from primate populations on the island of Bioko, which is located 32 km off the west coast of Africa and has been separated from mainland Africa for at least 10,000 years, pushed the emergence of present-day SIV back to ≥32,000 years before present [92]. Confusingly, African green monkey (AGM, genus Chlorocebus) subspecies each harbor a distinct lineage of SIVagm. This suggests a long-time presence of the virus, with an origin dating to somewhere between AGM subspeciation, estimated at 1.5–3 mya, and subsequent monkey migrations during the Plio-Pleistocene [88]. Should SIV indeed have a relatively short history in African primates, then it is understandable that the virus did not reach Asian monkey species. The Plio-Pleistocene is known for its lower global temperatures and, in the Pleistocene, repeated glaciation. Such climate conditions discouraged primate settlements in Europe and Arabia that could bridge the gap between Africa and Asia [93]. Theropithecus species did disperse to Europe in the early Pleistocene [94]. However, modern Theropithecus is not infected by SIV, nor are baboons, which currently still inhabit Saudi Arabia and Yemen. Papio hamadryas likely migrated from Africa to Arabia in the late Pleistocene and remained there ever since [95,96]. Looking at the spread of SIV, it is commonly found in arboreal primates inhabiting tropical forests rather than terrestrial, savannah or shrubland-dwelling monkeys such as macaques and baboons (Table 1). The exception to this observation are green monkeys. As a widespread species in sub-Saharan Africa, where they can be found in numerous environments, AGMs may function as an intermediary spreading SIV to savannah monkeys. It is interesting to note that at three occasions, wild baboons were found to be infected with SIVagm strains [79,80,81]. A Senegalese patas monkey (Erythrocebus patas), another ground-dwelling species living among AGMs, was likewise reported to carry a SIVagm strain [82].
Concluding, a relatively young age has been calculated for SIV based on nucleotide sequences and from geographical separation. Such a young age is supported by its absence in Asian primates, suggesting a post-Miocene origin. Probably, the virus originates from the tropical forests of sub-Saharan Africa and is now spreading to ground-dwelling primates through AGMs, which are widespread in Africa, have high SIV prevalence, and thrive in many habitats. AGMs themselves have probably obtained SIV from cross-species transmission to one of their subspecies with subsequent onwards transmission to the other AGM subspecies in the past [97].
3.1.3. Simian Type D retrovirus (SRV)
SRV, a primate betaretrovirus, which can induce considerable pathology in captive macaques, was found to have natural reservoirs in the Asian macaque species Macaca nemestrina, Macaca fascicularis, and Macaca mulatta [63,64,66,98]. In total, eight phylogenetically related SRV genotypes have been identified, all in macaques except SRV-6. This variant was detected in wild Hanuman langurs, Semnopithecus entellus, from India [67], suggesting that SRV-like viruses could have a broader species distribution. However, it has been noted that SRV is closely related to endogenous primate type D betaretrovirus SERV sequences. Especially the single SRV-7 sequence available, a partial fragment of pol, shows high homology to SERV pol from baboons (unpublished observation, and see [66,99]). Jayashree Nandi et al. therefore suggested that SRV-6 and -7 may be the result of recombination events between exogenous SRV and endogenous SERV genomes [66], which would explain their considerable genetic distance from SRV 1–5 and SRV-8. As SRV-6 also bears resemblance to an endogenous type D retrovirus (PO-1-Lu from Trachypithecus obscurus, a langur species from East Asia), it could also be a reactivation of such a provirus [100]. PO-1-Lu was shown to be very similar to Mason–Pfizer monkey virus (MPMV) isolated from a rhesus macaque, which turned out to be a variant of SRV-3 [101,102]. These difficulties show that type D endogenous and exogenous retroviral sequences are intertwined and that recombination, reactivation and/or germ line integration has probably happened several times, which greatly confuses SRV phylogeny.
Little is known about the origin and age of SRV, except for its spread in primate centers since at least the 1970s [99,103]. SRV 1–5 and SRV-8, which are genetically and serologically closely related, spread only within the genus Macaca, and have the ability to induce severe disease. Such characteristics are typical of a recently evolved virus species, making the putative recombinant SRV-types 6 and 7 detected in Indian langurs a distinct viral species. Increasing the search for both infectious and endogenous SRV-like retroviruses in Asian monkeys should help gain a better understanding of the distribution and evolutionary connections within this virus family.
3.1.4. Simian T-Lymphotropic Virus (STLV)
The deltaretrovirus STLV comprises multiple types and subtypes and has a broad distribution in OWM and apes, but is not found in NWM [104]. Phylogenetic analysis of LTR sequences suggests an African, rather than Asian origin for PTLV (primate T-lymphotropic virus, the collective name for STLV and HTLV, the Homo-specific variants that arose after cross-species transmission) [105,106]. Three STLV types have been recognized, all of which also infect humans [107]. HTLV-1 and HTLV-2 have by now been scattered around the world by human migration, while HTLV-3 is only found in Africa [107]. A fourth, distantly related type, HTLV-4, has been isolated from an individual in Cameroon, but the search for a simian counterpart has so far been unsuccessful [108].
Pathogenicity in humans and non-human primates has only convincingly been documented for PTLV-1 (the cluster to which both STLV-1 and HTLV-1 belong, reviewed in [76,109]). PTLV mostly replicates through clonal expansion, which results in a low mutation rate and a high genetic stability [110]. However, diversity is significant in this large virus family, making phylogenetic analysis of PTLV challenging. Often, studies are based on relatively limited information. For instance, transmission of STLV-1 to humans was estimated to have occurred 27,300 ± 8200 years ago based only on the analysis of LTR and env gene sequences [21]. However, further transmissions have taken place over time or are still ongoing (discussed in [76]), whereby geographic proximity is the basis of most shared evolutionary relationships [23]. In fact, each HTLV-1 subtype is probably the result of independent transmissions from either African or Asian simians [24,111]. Analysis using a relaxed molecular clock and only first and second codon positions of the tax and env genes, respectively, suggested that HTLV-4 diverged from HTLV-2/STLV-2 approximately 49,800–378,000 years ago [28]. In this study, the most recent common ancestor (MRCA) of the major PTLV clades was calculated to have existed between 214,650 (tax gene) and 385,100 (env gene) years ago [28]. Using amino acid sequences from 20 PTLV strains and a bovine deltaretrovirus as outgroup, another study suggested a much older date of origin for the PTLV MRCA, namely, >1.3 mya [26]. A third study, based on entire genomes without LTRs, suggested an age between 632,129 and 946,936 years for the whole PTLV cluster, and dated the subclades of PTLV-1, PTLV-2, and PTLV-3 to 53,000–79,684, 191,621–286,730, and 63,294–94,770 years ago, respectively [112].
Overall, it is clear that PTLV comprises a clade of ancient, genetically diverse viruses, where repeated interspecies transmissions as well as viruses migrating between Asia and Africa pose a challenge for those trying to reconstruct the history of the virus. Despite some variation in estimated divergence dates, the above studies all agree on a late Pleistocene origin for PTLV, with the three main types likely arising in the last 200,000 years.
3.2. Endogenous Retrovirus Integrations Predating OWM Speciation
Endogenous retrovirus genomes and parts of such proviruses abound in vertebrates. Most have entered the germ line long before the rise of the order of primates. The primate germ line was thus already seeded with many of such proviral remains before the diversification of extant OWM species. Examples of ancient proviruses, with their calculated time of integration, are human endogenous retrovirus-H (HERV-H, >40 mya), HERV-W (±63 mya), HERV-S (±43 mya), HERV-R (±33 mya), HERV-I (±33 mya), and HERV-E (10.7–41.3 mya) [113,114,115,116,117,118]. With the exception of HERV-S, which has some similarity to foamy viruses, all these HERVs belong to the gammaretrovirus group.
The situation for the supergroup of primate betaretroviruses, represented by HERV-K viruses in humans, and CERV-K in chimpanzees, is more complex [119,120]. Sequences homologous to HERV-K have been detected in all Old World primate species, with an estimated integration time of 28 mya [121]. However, specific members of the family show a more limited distribution and a later germ line introduction, suggestive of repeated activation and reinfection processes over time (discussed in Section 3.3.4).
3.3. Endogenous Retroviruses Specific for OWM
Four endogenous retroviruses are exclusively present in OWM genomes, namely, BaEV, PcEV, SERV, and SERV-K1/RhERV-K. Of these viruses, full-length proviral genomes can be detected in at least some OWM species [37,38,39,122,123]. In AGM (Vero) or baboon BEF-3 cell lines, virus particles containing SERV or BaEV genomes, respectively, can be produced upon stimulation [124,125]. For PcEV, expression has only been confirmed at the RNA level [126]. It is unknown whether this RNA comprises full-length genomes nor whether it is packaged. Similarly, for SERV-K1, only env mRNA expression has been verified [123]. Distribution of the four endogenous retroviruses among OWM species is shown in Table 2; corresponding geography and deduced virus migrations are depicted in Figure 1.
Several limitations apply when reconstructing the history of endogenous retroviruses from genome assemblies:
- Not all infected species may still roam the earth
- Not all infected species may contain germ line integrations
- Not all species have had their genomes sequenced
- Not all proviral sequences are likely due to bona fide viral infection and integration; they may for instance be acquired by hybridization between species
- Proviruses may have been completely or partially lost from the germ line, making identification challenging
- The quality of a genome assembly may be insufficient for provirus detection
- The distinction between endogenous and exogenous retroviruses is not always clear (see above, and the comment in [129])
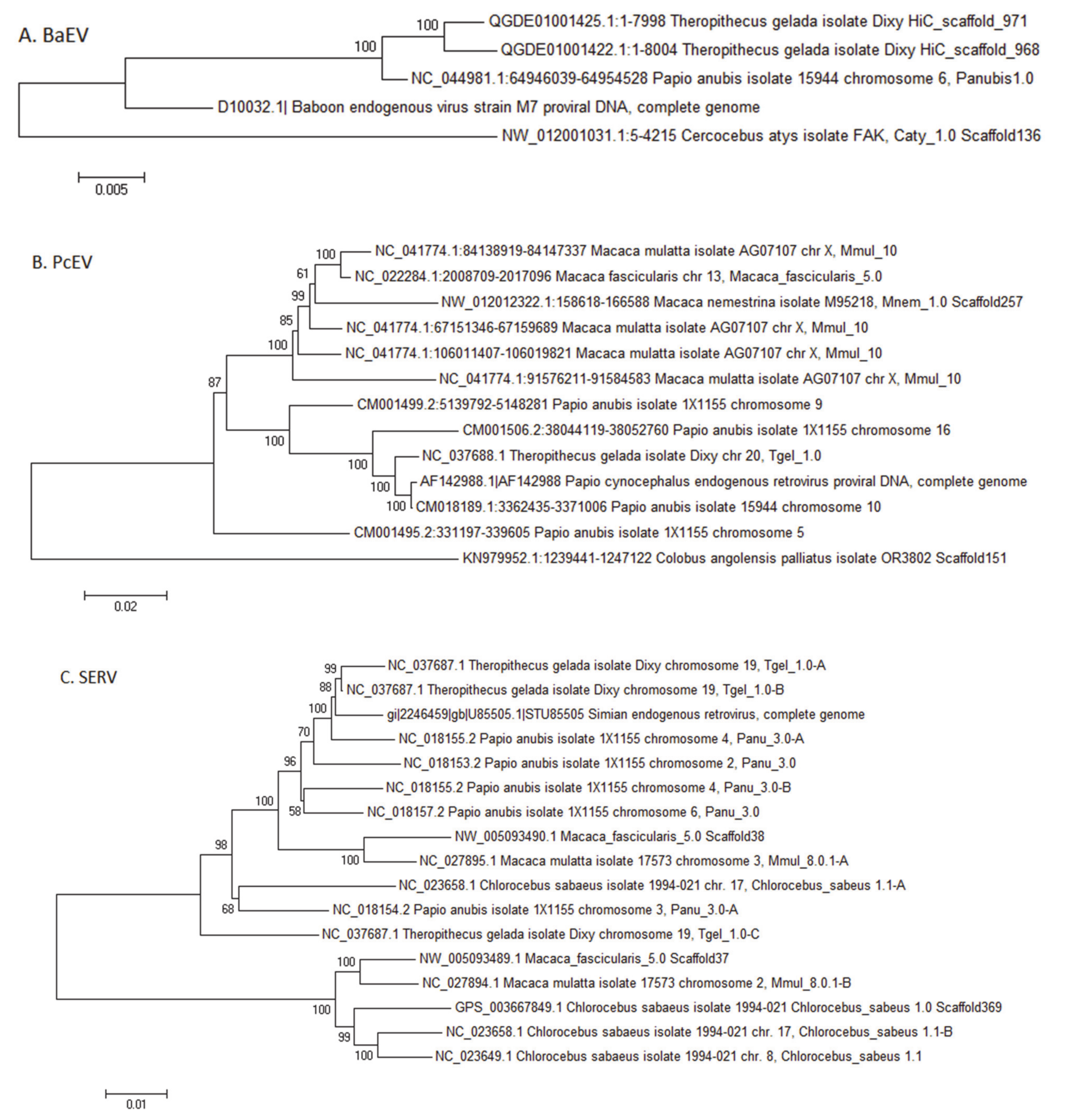
3.3.1. Baboon Endogenous Virus (BaEV)
BaEV was discovered as an integrated provirus in baboon DNA in 1978 and was completely sequenced in 1987 [39,134]. Non-infectious virus particles containing BaEV sequences can be induced from the Vero cell line, which is derived from Chlorocebus kidney epithelial tissue [124]. BaEV is a recombinant virus, originating from the now also endogenous monkey retroviruses PcEV and SERV [37]. Most BaEV proviral integrations are defective, although complete proviruses do exist in baboons and geladas (Appendix A Figure A1A) [122,125,134]. Although BaEV was at first thought to be widespread in primates, a PCR analysis of 24 African monkey species suggested that BaEV integrations are limited to few species (Table 2) [130]. In phylogenetic reconstructions, BaEV sequences did not follow the monkey phylogeny, but clustered according to habitat, implying that living together facilitates cross-species transmissions [130]. The BaEV_forest and BaEV_savannah strains were estimated to have diverged 24,000–400,000 years ago, using evolutionary rates calculated for other retroviruses [130]. Analyzing LTR divergence in full-length BaEV genomes from one baboon and two Theropithecus gelada showed a single (baboon) or no (gelada) substitutions between the 5′ and 3′ LTR, respectively (unpublished observation). As retroviral LTRs are identical at the time of integration, after which they start to diverge with the host mutation rate, little or no LTR variation is indicative of a relatively recent origin, probably at most ~300,000 years ago for proviruses with identical LTRs [135].
3.3.2. Papio Cynocephalus Endogenous Virus (PcEV)
The full-length endogenous gammaretrovirus PcEV was found by screening a baboon (Papio cynocephalus) genomic library [37]. Additional species harboring PcEV proviral sequences were identified during the analysis of OWM monkey DNA (Table 2, [131]). Interestingly, PcEV sequences (including LTRs) were found in all examined papionin species, but also in colobines of the genus Colobus: Colobus guereza and Colobus angolensis (Appendix A Figure A1B, [131]). PcEV_baboon was not found in Cercopithecini, although a variant virus is likely present there [131]. The upper limit of PcEV integration was set at 9 mya based on LTR divergence and species distribution, such as the dispersal of macaques into Asia around 5.5–7 mya [131]. Evidence for recent activity is given by proviruses with no or only a single nucleotide difference between their 5′ and 3′ LTRs, which can be identified in gelada and baboon genomic libraries, respectively (unpublished observation). In the rhesus macaque genome, PcEV was identified as one of the retroviruses with relatively recent germ line activity [136] (see erratum in [126] as PcEV was mistakenly labelled BaEV in the earlier publication). PcEV integration sites in two olive baboons (no. 15944 and no. 1X1155) differ, suggesting that the proviruses are not fixed in this subspecies (unpublished observation). In addition, baboon PcEV integration sites are empty in the rhesus macaque genome, which points to independent integrations after speciation and not inheritance from a common ancestor (unpublished observation).
So, as PcEV is present in Colobus, but not in Cercopithecini, the exogenous period of PcEV can be estimated to have started around the macaque/papionin split, 7.6 ± 1.3 mya, until about 150,000–300,000 years ago. An ancient cross-species transmission to an ancestor of present-day Colobus species could explain its presence there [131].
3.3.3. Simian Endogenous Retrovirus (SERV)
Similar to PcEV, SERV, a full-length integrated type D betaretrovirus was first described from a baboon genomic library [38]. Subsequently, a widespread distribution in OWM with a relatively large nucleotide divergence between the viral sequences suggested a relative old age for the proviruses (Table 2) [38,132]. Heterozygous integrations abound, at least in Chlorocebus sabaeus, which suggests that many are so recent that they have not had sufficient time to become fixed in the population [128]. Fixation of traits is highly dependent upon effective population size, which depends on population size and generation time. For primate populations 1–3 million years are usually sufficient to fix most markers [137], suggesting that polymorphic insertions are less than 1–3 my old. A number of OWM genomes contain several full-length SERV genomes, often with open reading frames for at least some viral proteins (unpublished observation, Appendix A Figure A1C). SERV sequences do not cluster according to host species, implying that they integrated after speciation and that ancient cross-species transmissions occurred [38]. SERV particles can be expressed from Vero cells, but those are non-infectious in cell lines known to be permissive for type D retroviruses [124].
Full-length proviruses, intact reading frames, virus expression and heterozygous insertions do suggest a young age, but SERV has been calculated to have integrated on average 6.16 ± 3.41 (range 0‒21.62) mya in Asian colobines, which, however, carry a distinct variant of SERV, and somewhat later, 3.42 ± 2.20 (range 0.27–14.09) mya, in cercopithecine species; that is, monkeys of the subfamily Cercopithecinae; thus, both Cercopithecini and Papionini [132]. It was therefore suggested that SERV originated within the last 8 million years and continued its exogenous activity until quite recently. For instance, in Asian colobines, species specific, genus specific, but also shared integrations are seen [132]. Highly divergent SERV 5′ and 3′ LTRs can be found in cercopithecines. In contrast, in Theropithecus and Chlorocebus there are also a few proviruses with only 1–2 nt substitutions between the LTRs (unpublished observation), indeed pointing to a lengthy period of germ line activity. Rhinopithecus roxellana SERV integrations with open reading frames were found to be significantly younger than the ones with frame-shift mutations [132]. Furthermore, heterozygous SERV insertions are presumed to be more recent than integrations that have been fixed in a population.
3.3.4. Simian Endogenous Retrovirus-K1 (SERV-K1)/RhERV-K
The primate non-D type betaretrovirus superfamily, to which all ERV-K viruses belong, has a complex history. Deep-rooted proviral integrations are present in all OWM and hominoids, but there is also evidence of later activity. In both humans and chimpanzees, the possession of open reading frames combined with low LTR diversity in some proviruses as well as the existence of polymorphic insertions, are illustrative of such recent activity [127]. In case of chimpanzee CERV-K proviruses, integration times postdate the Homo–Pan divergence [127]. Another study dated the average integration time of rhesus macaque ERV-K (RhERV-K) at 10.3 mya, although almost identical LTRs in three complete proviruses suggested that some integrated <4.5 mya [120]. SERV-K1, a full-length ERV-K-like provirus, has been described in the rhesus macaque [123]. Most likely, SERV-K1 is identical to one of the RhERV-K integrations, but unfortunately RhERV-K sequences are no longer available in the database.
Southern blot analysis indicated that ERV-K proviruses, HERV-K(OLD), are present in all OWM [121]. However, as more recent, lineage specific ERV-K insertions have been reported, the presence of such ERV-K proviruses in OWM was investigated by querying their genome assemblies with full-length HERV-K and SERV-K1 sequences. Although proviral fragments with substantial homology (79–96%) were indeed detected in all OWM species, phylogenetic analysis as well as their chromosomal locations suggested that these ERV-K-like sequences had been present before the radiation of OWMs, and thus represent HERV-K(OLD) insertions [121]. In contrast, full-length or near full-length proviruses were found in macaques, in the gelada, and in the olive baboon genome assemblies when using SERV-K1 as query (Table 2, Appendix B Figure A2). The coding regions of the SERV-K1-like proviruses were highly homologous (80–99% homology over substantial genomic lengths) to the HERV-K family, including its ERV-K(OLD) members. Fortunately, SERV-K1 LTRs proved to be relatively unique and suitable for virus identification. However, such acquisition or exchange of (novel) sequence stretches, which appears to be quite common for the ERV-K family, makes tracing its history through the primate germ line a challenging undertaking. It is thus not easy to identify the species infected with those specific SERV-K1 viruses, as only Macaca mulatta and Macaca fascicularis proviruses did contain both LTRs, while those in Macaca fuscata lack them. Furthermore, SERV-K1 proviruses are incomplete in Macaca nemestrina, although the particular LTR sequences are present (unpublished observation). Asian Trachypithecus francoisi and African Colobus angolensis, but not other colobine species, do contain integrations with LTR sequences that are largely homologous to SERV-K1 LTRs. Considering its age and spread, SERV-K1 may have migrated with its colobine and papionin hosts to Asia in the late Miocene. Comparing SERV-K1 LTR sequences from rhesus macaque full-length proviruses shows that the 5′ and 3′ LTRs have no, or very low, nucleotide diversity (unpublished observation, Appendix B Figure A2), suggestive of relatively recent integration events. Apparently, SERV-K1 activity started before the primate Out-of-Africa migrations and continued for quite some time after that. The occurrence of a few very similar SERV-K1 proviruses in rhesus and crab-eating macaque genomes (unpublished observation) can likely be explained by interbreeding [138]. Several SERV-K1 LTRs in papionins are almost identical in sequence and could thus predate speciation. A quick inspection of Blast hits, however, suggests that the integration sites do not match, so that the proviral integrations must largely have been independent events (result not shown).
The uneven distribution of SERV-K1 complete proviruses suggests that either the infectious virus did not always reach the OWM germ line, that the integrations were lost, or that they are somehow not in the genome assemblies. Overall, due its apparently highly infectious nature over long periods of time, and a tendency to recombine, the epidemiological trail of the ERV-K virus family is not easy to follow. The recent spread, on two continents, of two related SERV-K viruses in two very different monkey genera, papionins and Colobus/Trachypithecus, is thus an interesting subject for further study.
Infectious period estimates of the eight OWM retroviruses in relation to the geological time scale are summarized in Figure 2.
4. Discussion
The order of primates probably dates to the Paleocene (56–65 mya). Primate fossils appeared simultaneously in Europe, Asia, and Africa in the early Eocene (55–50 mya), suggesting that by then, the order was firmly established. Since primates are relatively intolerant of cooler climates, their history is largely determined by climatic variations, which in their turn are linked to tectonic and cosmic events [93]. The rapid evolution and dispersal of primate species was likely facilitated by a steep rise in global temperature at the Paleocene/Eocene boundary, which resulted in tropical climates even at higher latitudes [93]. Moreover, firm geographical connections between Asia, Europe, and North America existed at that time, giving the early primates sufficient territory to expand. Global temperatures declined during the Eocene, although warmer periods, for instance in the Miocene, would continue to occur. The much colder conditions in the Oligocene and especially in the more recent Pliocene and Pleistocene periods forced primates to retreat from previously inhabited regions. Under dryer, colder conditions, terrestrial papionins, like macaques and baboons, could from time to time establish themselves in such climates, but the arboreal monkeys of the tropical forests were unable to adapt. For instance, colobines migrated successfully from Africa to Eurasia in the late Miocene but these dispersals stopped in the Plio/Pleistocene; Papio and Theropithecus were the only species then trekking to Eurasia through Arabia [139]. Macaques migrated from Africa to Eurasia around 5.5–7 mya in the late Miocene or early Pleistocene [138]. Only a single, ancestral species, Macaca sylvanus, is now left in (northern) Africa; European macaques have gone extinct in the late Pleistocene [140]. Modern Asian macaque species show no genomic evidence of later introductions, suggesting that subsequent migrations, if any, have been unsuccessful [138].
With the above described primate evolution in mind, an attempt to retrace the history of one of their successful parasites, the retrovirus family, was made. Retroviruses are generally transmitted by close contact, for instance by the exchange of saliva or blood (biting), through sexual encounters, or from mother-to child, implying that these pathogens are closely associated with their host species. Infections by aerosols, the oral–fecal route or by insect vectors are relatively rare or are unlikely; only equine infectious anemia virus, EIAV, and possibly bovine leukemia virus, BLV, can sometimes be transmitted by flies from viremic hosts [141,142]. So, in general, the history of a retrovirus is closely related to that of its host. Of course, transmission of viruses can be hampered by cellular and genetic variation between species. For SIV, immune system activity, including the actions of restriction factors such as TRIM5alpha and APOBEC3G, as well as amino acid variation in for instance the coreceptor protein, have all been implicated in the prevention of cross-species transmission (for a review, see [143]). Such inhibitory mechanisms could lead to false conclusions as to why certain species are free of infection but would hardly play a role in conclusions drawn from infected species.
Investigating the spread and estimated age of eight OWM-infecting retroviral species suggests that SFV, the most widely spread virus of all, with a distribution over three continents, is also the oldest virus in the group. Ancestral primates most likely migrated, possibly by rafting, to South America during the Paleogene when that continent was relatively close to Africa [93]. As the nodal ancestor of OWM and NWM SFV has been dated to ~65 mya [49], the virus could have arrived in South America together with its host. A nodal ancestor by definition predates its progeny, and need not have been a primate virus, which would date the emergence of genuine SFV-NWM and SFV-OWM after 65 mya. Various approaches have estimated the node separating NWM and OWM at ~40 mya in the Eocene, which would fit with the ancestry of the virus lineages [2]. However, as cross-species transmission and recombination have been documented for foamy viruses, alternative scenarios with a later introduction in NWM could also be a possibility.
The next oldest viruses, SERV and SERV-K1 (RhERV-K), are now known only in an endogenous form. Their putative Miocene origin, together with a wide distribution in OWM species in both Africa and Asia, suggests that SERV and SERV-K1 have been travelling the land bridge(s) between the continents established after the final closure of the Tethys Seaway ~14 mya [144]. The relatively warm climate of the Miocene facilitated the dispersal of arboreal, tropical colobines as well as terrestrial macaques; the genera underwent an extensive diversifying radiation in Asia [139]. A third virus with a similar distribution, both in species and geography, is STLV. However, phylogenetic analysis indicated that the common ancestor of STLV genotypes dates to ~1 mya, so that spread of STLV cannot readily be explained by Miocene migrations. A later introduction with rapid onward transmissions and adaptation could be hypothesized, though PTLV is mainly transmitted through sexual contact, blood, or saliva exposure (biting), or vertically from mother to child, which makes spread in and between primate groups relatively slow [145,146]. Alternatively, STLV might be much older than estimated from sequence data, as we have possibly not yet sampled all PTLV variation, or the evolutionary models used to analyse PTLV may be incorrect [76,147].
Another virus that likely migrated with its host from Africa to Asia is PcEV, which is found in African papionins as well as in Asian macaques. Such a distribution suggests that PcEV is at least 5.5–7 mya old, but not much older as the virus is not found in the germ line of the Cercopithecini, implying that migration with its macaque host took place in the late Miocene or early Pliocene. Recombination of PcEV with SERV, when at least one of the two viruses was actively replicating, gave rise to BaEV, which, by definition, should be younger than its parents. Indeed, BaEV is not found in Asian macaques, suggesting its infectious period postdates the divergence of macaques from the other papionins. The putative Pleistocene origin of BaEV prohibited its spread to Asia, as by then the climate was no longer favorable for monkey expansions. BaEV may have been present in European baboons, having gone extinct there together with its host. Moreover, it would be interesting to investigate whether or not African Macaca sylvanus carry PcEV and/or BaEV integrations.
Much younger OWM retroviruses appear to be SIV and SRV; SIV has an exclusively African distribution while SRV is seemingly only found in Asian macaques. The uninhabitable nature after the Miocene period of areas in key corridors for primates between Africa and Asia, with subsequent isolation of monkey species on both continents, likely impeded virus transfer between continents for these recently emerged viruses [148,149,150,151,152]. For a relatively young virus that owes its present global spread to a recent leap to mankind, SIV has a remarkably broad distribution in African primates. Unexpectedly, the list of SIV-infected species shows only anecdotal detection in terrestrial papionins such as baboons and geladas, while infections are common in monkeys and apes inhabiting the tropical forests of sub-Saharan Africa, including the forest-dwelling papionins Mandrillus, Cercocebus, and Lophocebus. An exception here are AGMs (Chlorocebus spp.), found just about everywhere in sub-Saharan Africa, which carry a great diversity of SIV strains and have high SIV prevalence. They may have facilitated SIV cross-species transmissions in the past, similar to modern AGM transmitting SIVagm to individual baboons and patas monkeys. An example of another retrovirus shared between AGMs and papionins is BaEV.
For endogenous viruses that have gone extinct, once infected species may be missed because no germ line introductions occurred or were lost in subsequent generations. Still, the available data on endogenous viruses can be used to infer a plausible evolutionary trajectory. Retroviruses that use the neutral amino acid transporter ASCT2 as a cellular receptor, which is expressed on oocytes, spermatozoa, and pre-implantation embryos, have a tendency to enter the germ line [153,154,155]. Such viruses, here represented by the type D viruses SERV and BaEV, are thus well equipped to create a broad genomic track, and will therefore be less likely to disappear from sight. Indeed, SERV has an extensive distribution in OWM, while—putative—non-ASCT2 using PcEV has a relatively limited distribution in papionin genomes only. However, although BaEV is able to the ASCT2 receptor, it has a more limited distribution than SERV, which may either be a true representation of the species infected in the past but could also be due to other causes. For instance, BaEV can use the neutral amino acid transporter ASCT1 as an additional receptor, which other type D viruses cannot [156]. Such variation in receptor use may suggest an alternative trajectory in the host, although ASCT1 has a similar broad tissue expression as ASCT2 [157]. Alternatively, superinfection resistance (SIR), whereby for instance a homologous Env protein occupies the viral receptor so that new infections cannot take place, could play a role here (for a review, see [158]). The expression of endogenous SERV Env would make such a scenario possible. In human placenta, a HERV-F Env protein has been shown to inhibit betaretroviral Env protein mediated cell fusion through ASCT2 binding [159].
Interestingly, all four now endogenous OWM retroviruses have disappeared from the circulation, suggesting that virus endogenization ultimately means the end of the contagious variant. SIR could be such a possible mechanism to limit virus spread. Thus, opting for germ line transmission seems to be an indicator for extinction of infectious retrovirus lineages, while old age, as exemplified by the current infectivity of SFV, does not need to be.
5. Conclusions
Current distribution of primates, prehistoric primate evolution and expansion together with virus prevalence and sequence information have been used to infer the history of primate retroviruses. African and Asian monkey species are, or have been, infected by at least eight distinct retroviral species. Four of those have now become part of the germ line while four others are currently in the infectious stage, including the most ancient retrovirus, SFV. Evidence suggests that the older retroviruses—PcEV, SERV, SERV-K1—have migrated with their hosts to Asia ~5 to 10 mya, while the more recent ones—BaEV, SIV, and SRV—are limited to their respective continents of origin. For STLV, the reconstruction of its history proved more difficult, as its relatively recent age and widespread distribution does not correspond with Miocene migration.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2673-3986/2/1/5/s1, File S1: Alignment of OWM ERV sequences, File S2: Alignment of OWM SERV-K LTR sequences.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Files 1 and 2.
Conflicts of Interest
The author declares no conflict of interest.
Appendix A
Figure A1.
Evolutionary relationships of full-length or near full-length proviruses of BaEV (A), PcEV (B), and SERV (C) in OWM.
Figure A1.
Evolutionary relationships of full-length or near full-length proviruses of BaEV (A), PcEV (B), and SERV (C) in OWM.
The evolutionary history of integrated proviruses of BaEV (panel A), PcEV (panel B) and SERV (panel C) in OWM genomes with the corresponding reference virus sequence was inferred using the Neighbor-Joining method [160]. Bootstrap values of 500 replicates are shown next to the branches [161]. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Kimura 2-parameter method [40] and are given as the number of base substitutions per site. There were a total of 8492 positions in the dataset for BaEV, 8282 for PcEV, and 8382 for SERV. Evolutionary analyses were conducted in MEGA6 [41]. For some species, such as Macaca nemestrina, Mandrillus leucophaeus, and Cercocebus atys, the assembled genome sequences were of relative low quality, in other instances it was ascertained that all viral elements, gag-pol-env coding regions and LTRs, were present with high similarity (>95%), although the proviruses themselves were likely fragmented in those species and full-length or near full-length proviruses could not be retrieved. The primate species name is embedded within the sequence name. The sequence alignment file is available as Supplementary File 1.
Appendix B
Figure A2.
Evolutionary relationships of SERV-K1 LTR sequences in OWM.

The evolutionary history of LTR sequences of SERV-K1 in OWM genomes and those of the rhesus macaque reference virus were inferred using the Neighbor-Joining method [160]. For complete proviruses, both the 5′ LTR and 3′ LTR were included in the analysis. Bootstrap values of 500 replicates are shown next to the branches [161]. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Kimura 2-parameter method [40]. Evolutionary analyses were conducted in MEGA6 [41]. There were a total of 572 positions in the dataset. The primate species name is embedded within the sequence name. The sequence alignment file is available as Supplementary File 2.
References
- Elton, S. Environmental correlates of the cercopithecoid radiations. Folia Primatol. 2007, 78, 344–364. [Google Scholar] [CrossRef] [PubMed]
- Steiper, M.E.; Young, N.M. Primate molecular divergence dates. Mol. Phylogenet. Evol. 2006, 41, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Stevens, N.J.; Seiffert, E.R.; O’Connor, P.M.; Roberts, E.M.; Schmitz, M.D.; Krause, C.; Gorscak, E.; Ngasala, S.; Hieronymus, T.L.; Temu, J. Palaeontological evidence for an Oligocene divergence between Old World monkeys and apes. Nature 2013, 497, 611–614. [Google Scholar] [CrossRef]
- Raaum, R.L.; Sterner, K.N.; Noviello, C.M.; Stewart, C.-B.; Disotell, T.R. Catarrhine primate divergence dates estimated from complete mitochondrial genomes: Concordance with fossil and nuclear DNA evidence. J. Hum. Evol. 2005, 48, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A. Origin of the retroviruses: When, where, and how? Curr. Opin. Virol. 2017, 25, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Lerat, E.; Capy, P. Retrotransposons and retroviruses: Analysis of the envelope gene. Mol. Biol. Evol. 1999, 16, 1198–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Dolja, V.V. Virus world as an evolutionary network of viruses and capsidless selfish elements. Microbiol. Mol. Biol. Rev. 2014, 78, 278–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordonnier, A.; Casella, J.F.; Heidmann, T. Isolation of novel human endogenous retrovirus-like elements with foamy virus-related pol sequence. J. Virol. 1995, 69, 5890–5897. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Korber, B.T.; Nahmias, A.J.; Hooper, E.; Sharp, P.M.; Ho, D.D. An African HIV-1 sequence from 1959 and implications for the origin of the epidemic. Nature 1998, 391, 594–597. [Google Scholar] [CrossRef]
- Worobey, M.; Gemmel, M.; Teuwen, D.E.; Haselkorn, T.; Kunstman, K.; Bunce, M.; Muyembe, J.-J.; Kabongo, J.-M.M.; Kalengayi, R.M.; Van Marck, E.; et al. Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature 2008, 455, 661–664. [Google Scholar] [CrossRef] [Green Version]
- Faria, N.R.; Rambaut, A.; Suchard, M.A.; Baele, G.; Bedford, T.; Ward, M.J.; Tatem, A.J.; Sousa, J.D.; Arinaminpathy, N.; Pépin, J.; et al. HIV epidemiology. The early spread and epidemic ignition of HIV-1 in human populations. Science 2014, 346, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, V.M.; Olmsted, R.A.; Murphey-Corb, M.; Purcell, R.H.; Johnson, P.R. An African primate lentivirus (SIVsmclosely related to HIV-2. Nature 1989, 339, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Plantier, J.-C.; Leoz, M.; Dickerson, J.E.; De Oliveira, F.; Cordonnier, F.; Lemée, V.; Damond, F.; Robertson, D.L.; Simon, F. A new human immunodeficiency virus derived from gorillas. Nat. Med. 2009, 15, 871–872. [Google Scholar] [CrossRef]
- D’Arc, M.; Ayouba, A.; Esteban, A.; Learn, G.H.; Boué, V.; Liegeois, F.; Etienne, L.; Tagg, N.; Leendertz, F.H.; Boesch, C.; et al. Origin of the HIV-1 group O epidemic in western lowland gorillas. Proc. Natl. Acad. Sci. USA 2015, 112, E1343–E1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef]
- Huet, T.; Cheynier, R.; Meyerhans, A.; Roelants, G.; Wain-Hobson, S. Genetic organization of a chimpanzee lentivirus related to HIV-1. Nature 1990, 345, 356–359. [Google Scholar] [CrossRef]
- Roques, P.; Robertson, D.L.; Souquière, S.; Apetrei, C.; Nerrienet, E.; Barré-Sinoussi, F.; Müller-Trutwin, M.; Simon, F. Phylogenetic characteristics of three new HIV-1 N strains and implications for the origin of group N. AIDS 2004, 18, 1371–1381. [Google Scholar] [CrossRef]
- Chen, Z.; Telfier, P.; Gettie, A.; Reed, P.; Zhang, L.; Ho, D.D.; Marx, P.A. Genetic characterization of new West African simian immunodeficiency virus SIVsm: Geographic clustering of household-derived SIV strains with human immunodeficiency virus type 2 subtypes and genetically diverse viruses from a single feral sooty mangabey troop. J. Virol. 1996, 70, 3617–3627. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Luckay, A.; Sodora, D.L.; Telfer, P.; Reed, P.; Gettie, A.; Kanu, J.M.; Sadek, R.F.; Yee, J.; Ho, D.D.; et al. Human immunodeficiency virus type 2 (HIV-2) seroprevalence and characterization of a distinct HIV-2 genetic subtype from the natural range of simian immunodeficiency virus-infected sooty mangabeys. J. Virol. 1997, 71, 3953–3960. [Google Scholar] [CrossRef] [Green Version]
- Santiago, M.L.; Range, F.; Keele, B.F.; Li, Y.; Bailes, E.; Bibollet-Ruche, F.; Fruteau, C.; Noë, R.; Peeters, M.; Brookfield, J.F.Y.; et al. Simian immunodeficiency virus infection in free-ranging sooty mangabeys (cercocebus atys atys) from the Taï Forest, Côte d’Ivoire: Implications for the origin of epidemic human immunodeficiency virus type 2. J. Virol. 2005, 79, 12515–12527. [Google Scholar] [CrossRef] [Green Version]
- Van Dooren, S.; Salemi, M.; Vandamme, A.-M. Dating the origin of the African human T-cell lymphotropic virus type-I (HTLV-I) subtypes. Mol. Biol. Evol. 2001, 18, 661–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dooren, S.; Verschoor, E.J.; Fagrouch, Z.; Vandamme, A.-M. Phylogeny of primate T lymphotropic virus type 1 (PTLV-1) including various new Asian and African non-human primate strains. Infect. Genet. Evol. 2007, 7, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Pecon-Slattery, J.; Franchini, G.; Gessain, A. Genomic evolution, patterns of global dissemination, and interspecies transmission of human and simian T-cell leukemia/lymphotropic viruses. Genome Res. 1999, 9, 525–540. [Google Scholar]
- Reid, M.J.; Switzer, W.M.; Schillaci, M.A.; Ragonnet-Cronin, M.; Joanisse, I.; Caminiti, K.; Lowenberger, C.; Galdikas, B.M.F.; Sandstrom, P.A.; Brooks, J.I. Detailed phylogenetic analysis of primate T-lymphotropic virus type 1 (PTLV-1) sequences from orangutans (Pongo pygmaeus) reveals new insights into the evolutionary history of PTLV-1 in Asia. Infect. Genet. Evol. 2016, 43, 434–450. [Google Scholar] [CrossRef]
- Hron, T.; Elleder, D.; Gifford, R.J. Deltaretroviruses have circulated since at least the Paleogene and infected a broad range of mammalian species. Retrovirology 2019, 16, 33. [Google Scholar] [CrossRef]
- Salemi, M.; Desmyter, J.; Vandamme, A.-M. Tempo and mode of human and simian T-lymphotropic virus (HTLV/STLV) evolution revealed by analyses of full-genome sequences. Mol. Biol. Evol. 2000, 17, 374–386. [Google Scholar] [CrossRef]
- Afonso, P.V.; Cassar, O.; Gessain, A. Molecular epidemiology, genetic variability and evolution of HTLV-1 with special emphasis on African genotypes. Retrovirology 2019, 16, 39. [Google Scholar] [CrossRef] [Green Version]
- Switzer, W.M.; Salemi, M.; Qari, S.H.; Jia, H.; Gray, R.R.; Katzourakis, A.; Marriott, S.J.; Pryor, K.N.; Wolfe, N.D.; Burke, D.S.; et al. Ancient, independent evolution and distinct molecular features of the novel human T-lymphotropic virus type 4. Retrovirology 2009, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Wolfe, N.D.; Sintasath, D.M.; Tamoufe, U.; Lebreton, M.; Djoko, C.F.; Diffo, J.L.D.; Pike, B.L.; Heneine, W.; Switzer, W.M. Emergence of a novel and highly divergent HTLV-3 in a primate hunter in Cameroon. Virology 2010, 401, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Filippone, C.; Betsem, E.; Tortevoye, P.; Cassar, O.; Bassot, S.; Froment, A.; Fontanet, A.; Gessain, A. A Severe Bite from a Nonhuman Primate Is a Major Risk Factor for HTLV-1 Infection in Hunters From Central Africa. Clin. Infect. Dis. 2015, 60, 1667–1676. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Santini, D.M.; Stenbak, C.R.; Linial, M.L. Foamy virus zoonotic infections. Retrovirology 2017, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heneine, W.; Schweizer, M.; Sandstrom, P.; Folks, T. Human infection with foamy viruses. Curr. Top Microbiol. Immunol. 2003, 277, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Mouinga-Ondémé, A.; Kazanji, M. Simian foamy virus in non-human primates and cross-species transmission to humans in Gabon: An emerging zoonotic disease in Central Africa? Viruses 2013, 5, 1536–1552. [Google Scholar] [CrossRef] [Green Version]
- Power, M.D.; Marx, P.A.; Bryant, M.L.; Gardner, M.B.; Barr, P.J.; Luciw, P.A. Nucleotide sequence of SRV-1, a type D simian acquired immune deficiency syndrome retrovirus. Science 1986, 231, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.B. The history of simian AIDS. J. Med. Primatol. 1996, 25, 148–157. [Google Scholar] [CrossRef]
- Lerche, N.W.; Switzer, W.M.; Yee, J.L.; Shanmugam, V.; Rosenthal, A.N.; Chapman, L.E.; Folks, T.M.; Heneine, W. Evidence of infection with simian Type D retrovirus in persons occupationally exposed to nonhuman primates. J. Virol. 2001, 75, 1783–1789. [Google Scholar] [CrossRef] [Green Version]
- Mang, R.; Goudsmit, J.; Van Der Kuyl, A.C. Novel endogenous Type C retrovirus in baboons: Complete sequence, providing evidence for baboon endogenous virus gag-pol ancestry. J. Virol. 1999, 73, 7021–7026. [Google Scholar] [CrossRef] [Green Version]
- Van Der Kuyl, A.C.; Mang, R.; Dekker, J.T.; Goudsmit, J. Complete nucleotide sequence of simian endogenous type D retrovirus with intact genome organization: Evidence for ancestry to simian retrovirus and baboon endogenous virus. J. Virol. 1997, 71, 3666–3676. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Matsuo, K.; Nishimura, N.; Takahashi, N.; Takano, T. The entire nucleotide sequence of baboon endogenous virus DNA: A chimeric genome structure of murine type C and simian type D retroviruses. Jpn. J. Genet. 1987, 62, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzourakis, A.; Tristem, M.; Pybus, O.G.; Gifford, R.J. Discovery and analysis of the first endogenous lentivirus. Proc. Natl. Acad. Sci. USA 2007, 104, 6261–6265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkasova, H.; Hron, T.; Paces, J.; Hulva, P.; Benda, P.; Gifford, R.J.; Elleder, D. Discovery of an endogenous Deltaretrovirus in the genome of long-fingered bats (Chiroptera: Miniopteridae). Proc. Natl. Acad. Sci. USA 2017, 114, 3145–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.-Z.; Worobey, M. An endogenous foamy virus in the aye-aye (Daubentonia madagascariensis). J. Virol. 2012, 86, 7696–7698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.S.; Bodem, J.; Buseyne, F.; Gessain, A.; Johnson, W.; Kuhn, J.H.; Kuzmak, J.; Lindemann, D.; Linial, M.L.; Löchelt, M.; et al. Spumaretroviruses: Updated taxonomy and nomenclature. Virology 2018, 516, 158–164. [Google Scholar] [CrossRef]
- Falcone, V.; Leupold, J.; Clotten, J.; Urbanyi, E.; Herchenröder, O.; Spatz, W.; Volk, B.; Böhm, N.; Toniolo, A.; Neumann-Haefelin, D.; et al. Sites of simian foamy virus persistence in naturally infected African green monkeys: Latent provirus is ubiquitous, whereas viral replication is restricted to the oral mucosa. Virology 1999, 257, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Murray, S.; Picker, L.J.; Axthelm, M.K.; Hudkins, K.; Alpers, C.E.; Linial, M.L. Replication in a superficial epithelial cell niche explains the lack of pathogenicity of primate foamy virus infections. J. Virol. 2008, 82, 5981–5985. [Google Scholar] [CrossRef] [Green Version]
- Aiewsakun, P.; Katzourakis, A. Marine origin of retroviruses in the early Palaeozoic Era. Nat. Commun. 2017, 8, 13954. [Google Scholar] [CrossRef]
- Shankar, A.; Sibley, S.D.; Goldberg, T.L.; Switzer, W.M. Molecular analysis of the complete genome of a simian foamy virus infecting hylobates pileatus (pileated gibbon) reveals ancient co-evolution with lesser apes. Viruses 2019, 11, 605. [Google Scholar] [CrossRef] [Green Version]
- Reid, M.J.C.; Switzer, W.M.; Schillaci, M.A.; Klegarth, A.R.; Campbell, E.; Ragonnet-Cronin, M.; Joanisse, I.; Caminiti, K.; Lowenberger, C.; Galdikas, B.M.F.; et al. Bayesian inference reveals ancient origin of simian foamy virus in orangutans. Infect. Genet. Evol. 2017, 51, 54–66. [Google Scholar] [CrossRef]
- Ghersi, B.M.; Jia, H.; Aiewsakun, P.; Katzourakis, A.; Mendoza, P.; Bausch, D.G.; Kasper, M.R.; Montgomery, J.M.; Switzer, W.M. Wide distribution and ancient evolutionary history of simian foamy viruses in New World primates. Retrovirology 2015, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzourakis, A.; Aiewsakun, P.; Jia, H.; Wolfe, N.D.; Lebreton, M.; Yoder, A.D.; Switzer, W.M. Discovery of prosimian and afrotherian foamy viruses and potential cross species transmissions amidst stable and ancient mammalian co-evolution. Retrovirology 2014, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broussard, S.R.; Comuzzie, A.G.; Leighton, K.L.; Leland, M.; Whitehead, E.M.; Allan, J.S. Characterization of new simian foamy viruses from African nonhuman primates. Virology 1997, 237, 349–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, T.L.; Sintasath, D.M.; Chapman, C.A.; Cameron, K.M.; Karesh, W.B.; Tang, S.; Wolfe, N.D.; Rwego, I.; Ting, N.; Switzer, W.M. Coinfection of Ugandan Red Colobus (Procolobus [Piliocolobus] rufomitratus tephrosceles) with Novel, Divergent Delta-, Lenti-, and Spumaretroviruses. J. Virol. 2009, 83, 11318–11329. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.I.; Shanmugam, V.; Bhullar, V.B.; Beer, B.E.; Vallet, D.; Gautier-Hion, A.; Wolfe, N.D.; Karesh, W.B.; Kilbourn, A.M.; Tooze, Z.; et al. Screening for simian foamy virus infection by using a combined antigen Western blot assay: Evidence for a wide distribution among Old World primates and identification of four new divergent viruses. Virology 2003, 309, 248–257. [Google Scholar] [CrossRef]
- Meiering, C.D.; Linial, M.L. Historical perspective of foamy virus epidemiology and infection. Clin. Microbiol. Rev. 2001, 14, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Aghokeng, A.F.; Ayouba, A.; Mpoudi-Ngole, E.; Loul, S.; Liegeois, F.; Delaporte, E.; Peeters, M. Extensive survey on the prevalence and genetic diversity of SIVs in primate bushmeat provides insights into risks for potential new cross-species transmissions. Infect. Genet. Evol. 2010, 10, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Liegeois, F.; Courgnaud, V.; Switzer, W.M.; Murphy, H.W.; Loul, S.; Aghokeng, A.; Pourrut, X.; Mpoudi-Ngolé, E.; Delaporte, E.; Peeters, M. Molecular characterization of a novel simian immunodeficiency virus lineage (SIVtal) from northern talapoins (Miopithecus ogouensis). Virology 2006, 349, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, S.; Lafay, B.; Liegeois, F.; Ting, N.; Delaporte, E.; Peeters, M. Full molecular characterization of a simian immunodeficiency virus, SIVwrcpbt from Temminck’s red colobus (Piliocolobus badius temminckii) from Abuko Nature Reserve, The Gambia. Virology 2008, 376, 90–100. [Google Scholar] [CrossRef]
- Steve, A.-M.; Ahidjo, A.; Placide, M.-K.; Caroline, F.; Mukulumanya, M.; Simon-Pierre, N.-K.; Octavie, L.-M.; Valentin, M.-A.; Jean-Jacques, M.-T.; Eric, D.; et al. High prevalences and a wide genetic diversity of simian retroviruses in non-human primate bushmeat in rural areas of the democratic republic of congo. Ecohealth 2017, 14, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Vandewoude, S.; Apetrei, C. Going wild: Lessons from naturally occurring T-lymphotropic lentiviruses. Clin. Microbiol. Rev. 2006, 19, 728–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, T.; Ekwalanga, M.; Bikandou, B.; Ido, E.; Yamaguchi-Kabata, Y.; Ohkura, S.; Harada, H.; Takehisa, J.; Ichimura, H.; Parra, H.-J.; et al. A novel simian immunodeficiency virus from black mangabey (Lophocebus aterrimus) in the Democratic Republic of Congo. J. Gen. Virol. 2005, 86, 1967–1971. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.E.; Kerlin, R.L.; Zimmerman, T.E. Histologic lesions in cynomolgus monkeys (Macaca fascicularis) naturally infected with simian retrovirus type D: Comparison of seropositive, virus-positive, and uninfected animals. Toxicol. Pathol. 1999, 27, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moazed, T.C.; Thouless, M.E. Viral persistence of simian type D retrovirus (SRV-2/W) in naturally infected pigtailed macaques (Macaca nemestrina). J. Med. Primatol. 1993, 22, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Nandi, J.S.; Tikute, S.A.; Chhangani, A.K.; Potdar, V.A.; Tiwari-Mishra, M.; Ashtekar, R.A.; Kumari, J.; Walimbe, A.; Mohnot, S. Natural infection by simian retrovirus-6 (SRV-6) in Hanuman langurs (Semnopithecus entellus) from two different geographical regions of India. Virology 2003, 311, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Nandi, J.S.; Van Dooren, S.; Chhangani, A.K.; Mohnot, S.M. New simian β retroviruses from rhesus monkeys (Macaca mulatta) and langurs (Semnopithecus entellus) from Rajasthan, India. Virus Genes 2006, 33, 107–116. [Google Scholar] [CrossRef]
- Nandi, J.S.; Bhavalkar-Potdar, V.; Tikute, S.; Raut, C.G. A Novel Type D Simian Retrovirus Naturally Infecting the Indian Hanuman Langur (Semnopithecus entellus). Virology 2000, 277, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Ahuka-Mundeke, S.; Mbala-Kingebeni, P.; Liegeois, F.; Ayouba, A.; Lunguya-Metila, O.; Demba, D.; Bilulu, G.; Mbenzo-Abokome, V.; Inogwabini, B.-I.; Muyembe-Tamfum, J.-J.; et al. Identification and molecular characterization of new simian T cell lymphotropic viruses in nonhuman primates bushmeat from the Democratic Republic of Congo. AIDS Res. Hum. Retroviruses 2012, 28, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Ayouba, A.; Duval, L.; Liégeois, F.; Ngin, S.; Ahuka-Mundeke, S.; Switzer, W.M.; Delaporte, E.; Ariey, F.; Peeters, M.; Nerrienet, E. Nonhuman primate retroviruses from Cambodia: High simian foamy virus prevalence, identification of divergent STLV-1 strains and no evidence of SIV infection. Infect. Genet. Evol. 2013, 18, 325–334. [Google Scholar] [CrossRef]
- LeBreton, M.; Switzer, W.M.; Djoko, C.F.; Gillis, A.; Jia, H.; Sturgeon, M.M.; Shankar, A.; Zheng, H.; Nkeunen, G.; Tamoufe, U.; et al. A gorilla reservoir for human T-lymphotropic virus type 4. Emerg. Microbes Infect. 2014, 3, e7. [Google Scholar] [CrossRef]
- Leendertz, S.A.J.; Junglen, S.; Hedemann, C.; Goffe, A.; Calvignac-Spencer, S.; Boesch, C.; Leendertz, F.H. High prevalence, coinfection rate, and genetic diversity of retroviruses in wild red colobus monkeys (Piliocolobus badius badius) in Taï National Park, Côte d’Ivoire. J. Virol. 2010, 84, 7427–7436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mossoun, A.; Calvignac-Spencer, S.; Anoh, A.E.; Pauly, M.S.; Driscoll, D.; Michel, A.O.; Nazaire, L.G.; Pfister, S.; Sabwe, P.; Thiesen, U.; et al. Bushmeat hunting and zoonotic transmission of simian T-lymphotropic virus 1 in tropical west and central Africa. J. Virol. 2017, 91, e02479-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerrienet, E.; Meertens, L.; Kfutwah, A.; Foupouapouognigni, Y.; Ayouba, A.; Gessain, A. Simian T cell leukaemia virus type I subtype B in a wild-caught gorilla (Gorilla gorilla gorilla) and chimpanzee (Pan troglodytes vellerosus) from Cameroon. J. Gen. Virol. 2004, 85, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Saksena, N.K.; Hervé, V.; Durand, J.P.; Leguenno, B.; Diop, O.M.; Digoutte, J.P.; Mathiot, C.; Muller-Trutwin, M.; Love, J.L.; Dube, S.; et al. Seroepidemiologic, Molecular, and Phylogenetic Analyses of Simian T-Cell Leukemia Viruses (STLV-I) from Various Naturally infected Monkey Species from Central and Western Africa. Virology 1994, 198, 297–310. [Google Scholar] [CrossRef]
- Van Brussel, M.; Salemi, M.; Liu, H.-F.; Goubau, P.; Desmyter, J.; Vandamme, A.-M. The discovery of two new divergent STLVs has implications for the evolution and epidemiology of HTLVs. Rev. Med. Virol. 1999, 9, 155–170. [Google Scholar] [CrossRef]
- Jégado, B.; Kashanchi, F.; Dutartre, H.; Mahieux, R. STLV-1 as a model for studying HTLV-1 infection. Retrovirology 2019, 16, 41. [Google Scholar] [CrossRef]
- Courgnaud, V.; Van Dooren, S.; Liegeois, F.; Pourrut, X.; Abela, B.; Loul, S.; Mpoudi-Ngole, E.; Vandamme, A.; Delaporte, E.; Peeters, M. Simian T-Cell Leukemia Virus (STLV) Infection in Wild Primate Populations in Cameroon: Evidence for Dual STLV Type 1 and Type 3 Infection in Agile Mangabeys (Cercocebus agilis). J. Virol. 2004, 78, 4700–4709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniz, C.P.; Jia, H.; Shankar, A.; Troncoso, L.L.; Augusto, A.M.; Farias, E.; Pissinatti, A.; Fedullo, L.P.; Santos, A.F.; Soares, M.A.; et al. An expanded search for simian foamy viruses (SFV) in Brazilian New World primates identifies novel SFV lineages and host age-related infections. Retrovirology 2015, 12, 94. [Google Scholar] [CrossRef]
- Jin, M.J.; Rogers, J.; Phillips-Conroy, J.E.; Allan, J.S.; Desrosiers, R.C.; Shaw, G.M.; Sharp, P.M.; Hahn, B.H. Infection of a yellow baboon with simian immunodeficiency virus from African green monkeys: Evidence for cross-species transmission in the wild. J. Virol. 1994, 68, 8454–8460. [Google Scholar] [CrossRef] [Green Version]
- Van Rensburg, E.J.; Mwenda, J.; Robson, B.A.; Stander, T.; Laten, J.D.; Chege, G.K.; Engelbrecht, S. Simian immunodeficiency viruses (SIVs) from eastern and southern Africa: Detection of a SIVagm variant from a chacma baboon. J. Gen. Virol. 1998, 79, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Nyamota, R.; Owino, V.; Murungi, E.K.; Villinger, J.; Otiende, M.; Masiga, D.; Thuita, J.; Lekolool, I.; Jeneby, M.M. Broad diversity of simian immunodeficiency virus infecting Chlorocebus species (African green monkey) and evidence of cross-species infection in Papio anubis (olive baboon) in Kenya. J. Med. Primatol. 2020, 49, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Bibollet-Ruche, F.; Galat-Luong, A.; Cuny, G.; Sarni-Manchado, P.; Galat, G.; Durand, J.-P.; Pourrut, X.; Veas, F. Simian immunodeficiency virus infection in a patas monkey (Erythrocebus patas): Evidence for cross-species transmission from African green monkeys (Cercopithecus aethiops sabaeus) in the wild. J. Gen. Virol. 1996, 77, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Gifford, R.J.; Katzourakis, A.; Tristem, M.; Pybus, O.G.; Winters, M.; Shafer, R.W. A transitional endogenous lentivirus from the genome of a basal primate and implications for lentivirus evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 20362–20367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, C.; Maxfield, D.G.; Goodman, S.M.; Feschotte, C. Parallel germline infiltration of a lentivirus in two malagasy lemurs. PLoS Genet. 2009, 5, e1000425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.-Z.; Worobey, M. A primitive endogenous lentivirus in a colugo: Insights into the early evolution of lentiviruses. Mol. Biol. Evol. 2015, 32, 211–215. [Google Scholar] [CrossRef]
- Gifford, R.J. Viral evolution in deep time: Lentiviruses and mammals. Trends Genet. 2012, 28, 89–100. [Google Scholar] [CrossRef]
- Raehtz, K.; Pandrea, I.; Apetrei, C. The well-tempered SIV infection: Pathogenesis of SIV infection in natural hosts in the wild, with emphasis on virus transmission and early events post-infection that may contribute to protection from disease progression. Infect. Genet. Evol. 2016, 46, 308–323. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Jasinska, A.; Kristoff, J.; Grobler, J.P.; Turner, T.; Jung, Y.; Schmitt, C.; Raehtz, K.; Feyertag, F.; Sosa, N.M.; et al. SIVagm infection in wild African green monkeys from South Africa: Epidemiology, natural history, and evolutionary considerations. PLOS Pathog. 2013, 9, e1003011. [Google Scholar] [CrossRef] [Green Version]
- Apetrei, C. The history of SIVS and AIDS: Epidemiology, phylogeny and biology of isolates from naturally SIV infected non-human primates (NHP) in Africa. Front. Biosci. 2004, 9, 225–254. [Google Scholar] [CrossRef] [Green Version]
- Sharp, P.M.; Bailes, E.; Gao, F.; Beer, B.E.; Hirsch, V.M.; Hahn, B.H. Origins and evolution of AIDS viruses: Estimating the time-scale. Biochem. Soc. Trans. 2000, 28, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Wertheim, J.O.; Worobey, M. Dating the age of the SIV lineages that gave rise to HIV-1 and HIV-2. PLoS Comput. Biol. 2009, 5, e1000377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worobey, M.; Telfer, P.; Souquière, S.; Hunter, M.; Coleman, C.A.; Metzger, M.J.; Reed, P.; Makuwa, M.; Hearn, G.; Honarvar, S.; et al. Island biogeography reveals the deep history of SIV. Science 2010, 329, 1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleagle, J.G.; Gilbert, C.C. The biogeography of primate evolution: The role of plate tectonics, climate and chance. In Primate Biogeography; Lehman, S.M., Fleagle, J.G., Eds.; Springer: New York, NY, USA, 2006; pp. 375–418. [Google Scholar] [CrossRef]
- Rook, L.; Martínez-Navarro, B.; Howell, F.C. Occurrence of Theropithecus sp. in the Late Villafranchian of Southern Italy and implication for Early Pleistocene “out of Africa” dispersals. J. Hum. Evol. 2004, 47, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Winney, B.; Hammond, R.L.; Macasero, W.; Flores, B.; Boug, A.; Biquand, V.; Biquand, S.; Bruford, M.W. Crossing the Red Sea: Phylogeography of the hamadryas baboon, Papio hamadryas hamadryas. Mol. Ecol. 2004, 13, 2819–2827. [Google Scholar] [CrossRef] [PubMed]
- Kopp, G.H.; Roos, C.; Butynski, T.M.; Wildman, D.E.; Alagaili, A.N.; Groeneveld, L.F.; Zinner, D. Out of Africa, but how and when? The case of hamadryas baboons (Papio hamadryas). J. Hum. Evol. 2014, 76, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Wertheim, J.O.; Worobey, M. A Challenge to the Ancient Origin of SIVagm Based on African Green Monkey Mitochondrial Genomes. PLOS Pathog. 2007, 3, e95. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, R.; Okamoto, M.; Sakaguchi, S.; Nakagawa, S.; Miura, T.; Hirai, H.; Miyazawa, T. Simian Retrovirus 4 Induces Lethal Acute Thrombocytopenia in Japanese Macaques. J. Virol. 2015, 89, 3965–3975. [Google Scholar] [CrossRef] [Green Version]
- Zao, C.-L.; Tomanek, L.; Cooke, A.; Berger, R.; Yang, L.; Xie, C.; Chen, S.; Shi, C.; Rong, R. A novel simian retrovirus subtype discovered in cynomolgus monkeys (Macaca fascicularis). J. Gen. Virol. 2016, 97, 3017–3023. [Google Scholar] [CrossRef]
- Sommerfelt, M.A.; Harkestad, N.; Hunter, E. The endogenous langur type D retrovirus PO-1-Lu and its exogenous counterparts in macaque and langur monkeys. Virology 2003, 315, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Todaro, G.J.; Benveniste, R.E.; Sherr, C.J.; Schlom, J.; Schidlovsky, G.; Stephenson, J.R. Isolation and characterization of a new type D retrovirus from the Asian primate, Presbytis obscurus (spectacled langur). Virology 1978, 84, 189–194. [Google Scholar] [CrossRef]
- Sonigo, P.; Barker, C.; Hunter, E.; Wain-Hobson, S. Nucleotide sequence of Mason-Pfizer monkey virus: An immunosuppressive D-type retrovirus. Cell 1986, 45, 375–385. [Google Scholar] [CrossRef]
- Montiel, N.A. An updated review of simian betaretrovirus (SRV) in macaque hosts. J. Med. Primatol. 2010, 39, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Hayami, M.; Komuro, A.; Nozawa, K.; Shotake, T.; Ishikawa, K.-I.; Yamamoto, K.; Ishida, T.; Honjo, S.; Hinuma, Y. Prevalence of antibody to adult T-cell leukemia virus-associated antigens (ATLA) in Japanese monkeys and other non-human primates. Int. J. Cancer 1984, 33, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, A.-M.; Liu, H.-F.; Goubau, P.; Desmyter, J. Primate T-Lymphotropic Virus Type I LTR Sequence Variation and Its Phylogenetic Analysis: Compatibility with an African Origin of PTLV-I. Virology 1994, 202, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, A.-M.; Liu, H.-F.; Van Brussel, M.; De Meurichy, W.; Desmyter, J.; Goubau, P. The presence of a divergent T-lymphotropic virus in a wild-caught pygmy chimpanzee (Pan paniscus) supports an African origin for the human T-lymphotropic/simian T-lymphotropic group of viruses. J. Gen. Virol. 1996, 77, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Mahieux, R.; Gessain, A. HTLV-3/STLV-3 and HTLV-4 viruses: Discovery, epidemiology, serology and molecular aspects. Viruses 2011, 3, 1074–1090. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Heneine, W.; Carr, J.K.; Garcia, A.D.; Shanmugam, V.; Tamoufe, U.; Torimiro, J.N.; Prosser, A.T.; Lebreton, M.; Mpoudi-Ngole, E.; et al. Emergence of unique primate T-lymphotropic viruses among central African bushmeat hunters. Proc. Natl. Acad. Sci. USA 2005, 102, 7994–7999. [Google Scholar] [CrossRef] [Green Version]
- Eciminale, V.; Erende, F.; Ebertazzoni, U.; Romanelli, M. HTLV-1 and HTLV-2: Highly similar viruses with distinct oncogenic properties. Front. Microbiol. 2014, 5, 398. [Google Scholar] [CrossRef] [Green Version]
- Wattel, E.; Vartanian, J.P.; Pannetier, C.; Wain-Hobson, S. Clonal expansion of human T-cell leukemia virus type I-infected cells in asymptomatic and symptomatic carriers without malignancy. J. Virol. 1995, 69, 2863–2868. [Google Scholar] [CrossRef] [Green Version]
- Vandamme, A.-M. The simian origins of the pathogenic human T-cell lymphotropic virus type I. Trends Microbiol. 1998, 6, 477–483. [Google Scholar] [CrossRef]
- Switzer, W.M.; Qari, S.H.; Wolfe, N.D.; Burke, D.S.; Folks, T.M.; Heneine, W. Ancient origin and molecular features of the novel human T-lymphotropic virus type 3 revealed by complete genome analysis. J. Virol. 2006, 80, 7427–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mager, D.L.; Freeman, J. HERV-H endogenous retroviruses: Presence in the new world branch but amplification in the old world primate lineage. Virology 1995, 213, 395–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.H.-S.; Kim, D.-S.; Huh, J.-W.; Ahn, K.; Yi, J.M.; Lee, J.-R.; Hirai, H. Molecular characterization of the HERV-W env gene in humans and primates: Expression, FISH, phylogeny, and evolution. Mol. Cells 2008, 26, 53–60. [Google Scholar] [PubMed]
- Yi, J.-M.; Kim, T.-H.; Huh, J.-W.; Park, K.S.; Jang, S.B.; Kim, H.-M.; Kim, A.H.-S. Human endogenous retroviral elements belonging to the HERV-S family from human tissues, cancer cells, and primates: Expression, structure, phylogeny and evolution. Gene 2004, 342, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.H.-S.; Yi, J.-M.; Hirai, H.; Huh, J.-W.; Jeong, M.-S.; Jang, S.-B.; Kim, C.-G.; Saitou, N.; Hyun, B.-H.; Lee, W.-H. Human Endogenous Retrovirus (HERV)-R family in primates: Chromosomal location, gene expression, and evolution. Gene 2006, 370, 34–42. [Google Scholar] [CrossRef]
- Lee, J.-W.; Kim, H.-S. Endogenous retrovirus HERV-I LTR family in primates: Sequences, phylogeny, and evolution. Arch. Virol. 2006, 151, 1651–1658. [Google Scholar] [CrossRef]
- Yi, J.-M.; Kim, H.-S. Molecular evolution of the HERV-E family in primates. Arch. Virol. 2006, 151, 1107–1116. [Google Scholar] [CrossRef]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Romano, C.M.; De Melo, F.L.; Corsini, M.A.B.; Holmes, E.C.; Zanotto, P.M.D.A. Demographic histories of ERV-K in humans, chimpanzees and rhesus Monkeys. PLoS ONE 2007, 2, e1026. [Google Scholar] [CrossRef]
- Reus, K.; Mayer, J.; Sauter, M.; Zischler, H.; Müller-Lantzsch, N.; Meese, E. HERV-K(OLD): Ancestor Sequences of the Human Endogenous Retrovirus Family HERV-K(HML-2). J. Virol. 2001, 75, 8917–8926. [Google Scholar] [CrossRef] [Green Version]
- Van Der Kuyl, A.C.; Dekker, J.T.; Goudsmit, J. Full-length proviruses of baboon endogenous virus (BaEV) and dispersed BaEV reverse transcriptase retroelements in the genome of baboon species. J. Virol. 1995, 69, 5917–5924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.L.; Leon, E.J.; Wallace, L.; Nimiyongskul, F.A.; Buechler, M.B.; Newman, L.P.; Castrovinci, P.A.; Johnson, R.P.; Gifford, R.J.; Jones, R.B.; et al. Identification and spontaneous immune targeting of an endogenous retrovirus K envelope protein in the Indian rhesus macaque model of human disease. Retrovirology 2016, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Ma, Y.; Ma, W.; Williams, D.K.; Galvin, T.A.; Khan, A.S. Chemical induction of endogenous retrovirus particles from the vero cell line of African green monkeys. J. Virol. 2011, 85, 6579–6588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.; Rein, A.; Stephens, R.M.; O’Connell, C.; Gilden, R.V.; Shure, M.; Nicolson, M.O.; McAllister, R.M.; Davidson, N. Baboon endogenous virus genome: Molecular cloning and structural characterization of nondefective viral genomes from DNA of a baboon cell strain. Proc. Natl. Acad. Sci. USA 1981, 78, 5207–5211. [Google Scholar] [CrossRef] [Green Version]
- Maze, E.A.; Ham, C.; Kelly, J.; Ussher, L.; Almond, N.; Towers, G.J.; Berry, N.; Belshaw, R. Variable baseline papio cynocephalus endogenous retrovirus (PcEV) expression is upregulated in acutely SIV-infected macaques and correlated to STAT1 expression in the spleen. Front. Immunol. 2019, 10, 901. [Google Scholar] [CrossRef]
- Macfarlane, C.M.; Badge, R.M. Genome-wide amplification of proviral sequences reveals new polymorphic HERV-K(HML-2) proviruses in humans and chimpanzees that are absent from genome assemblies. Retrovirology 2015, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, C.; Sekizuka, T.; Kuroda, M.; Kasai, F.; Saito, K.; Ikeda, M.; Yamaji, T.; Osada, N.; Hanada, K. Novel endogenous simian retroviral integrations in Vero cells: Implications for quality control of a human vaccine cell substrate. Sci. Rep. 2018, 8, 644. [Google Scholar] [CrossRef]
- Vandamme, A.-M. BaEV is a relic from an ancient retrovirus that crossed species barriers. Trends Microbiol. 1996, 4, 478. [Google Scholar] [CrossRef]
- Van Der Kuyl, A.C.; Dekker, J.T.; Goudsmit, J. Distribution of baboon endogenous virus among species of African monkeys suggests multiple ancient cross-species transmissions in shared habitats. J. Virol. 1995, 69, 7877–7887. [Google Scholar] [CrossRef] [Green Version]
- Mang, R.; Maas, J.; Van Der Kuyl, A.C.; Goudsmit, J. Papio cynocephalus endogenous retrovirus among old world monkeys: Evidence for coevolution and ancient cross-species transmissions. J. Virol. 2000, 74, 1578–1586. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Satomura, K.; Sekizuka, T.; Hanada, K.; Endo, T.; Osada, N. Comprehensive phylogenomic analysis reveals a novel cluster of simian endogenous retroviral sequences in Colobinae monkeys. Am. J. Primatol. 2018, 80, e22882. [Google Scholar] [CrossRef] [PubMed]
- Van Der Kuyl, A.C.; Dekker, J.T.; Goudsmit, J. Primate genus miopithecus: Evidence for the existence of species and subspecies of dwarf guenons based on cellular and endogenous viral sequences. Mol. Phylogenetics Evol. 2000, 14, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Davidson, N.; Gilden, R.V.; McAllister, R.M.; Nicolson, M.O.; Stephens, R.M. The baboon endogenous virus genome. II. Provirus sequence variations in baboon cell DNA. Nucleic Acids Res. 1980, 8, 4423–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holloway, J.R.; Williams, Z.H.; Freeman, M.M.; Bulow, U.; Coffin, J.M. Gorillas have been infected with the HERV-K (HML-2) endogenous retrovirus much more recently than humans and chimpanzees. Proc. Natl. Acad. Sci. USA 2019, 116, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Magiorkinis, G.; Blanco-Melo, D.; Belshaw, R. The decline of human endogenous retroviruses: Extinction and survival. Retrovirology 2015, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Mailund, T.; Munch, K.; Schierup, M.H. Lineage sorting in apes. Annu. Rev. Genet. 2014, 48, 519–535. [Google Scholar] [CrossRef]
- Roos, C.; Kothe, M.; Alba, D.M.; Delson, E.; Zinner, D. The radiation of macaques out of Africa: Evidence from mitogenome divergence times and the fossil record. J. Hum. Evol. 2019, 133, 114–132. [Google Scholar] [CrossRef]
- Hughes, J.K.; Elton, S.; O’Regan, H.J. Theropithecus and ‘Out of Africa’ dispersal in the Plio-Pleistocene. J. Hum. Evol. 2008, 54, 43–77. [Google Scholar] [CrossRef]
- Elton, S.; O’Regan, H.J. Macaques at the margins: The biogeography and extinction of Macaca sylvanus in Europe. Quat. Sci. Rev. 2014, 96, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, J.A.; Adams, W.V., Jr.; Wilson, B.H.; Issel, C.J.; Roth, E.E. Transmission of equine infectious anemia virus by Tabanus fuscicostatus. J. Am. Veter-Med. Assoc. 1976, 168, 63–64. [Google Scholar]
- Kohara, J.; Takeuchi, M.; Hirano, Y.; Sakurai, Y.; Takahashi, T. Vector control efficacy of fly nets on preventing bovine leukemia virus transmission. J. Veter. Med. Sci. 2018, 80, 1524–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.C.; Burdo, T.H. HIV and SIV infection: The role of cellular restriction and immune responses in viral replication and pathogenesis. Apmis 2009, 117, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Bialik, O.M.; Frank, M.; Betzler, C.; Zammit, R.; Waldmann, N.D. Two-step closure of the Miocene Indian Ocean Gateway to the Mediterranean. Sci. Rep. 2019, 9, 8842. [Google Scholar] [CrossRef] [PubMed]
- Parrish, S.W.; Brown, A.E.; Chanbancherd, P.; Gettayacamin, M.; Parrish, J.H. Transmission of STLV in a closed colony of macaques. Am. J. Primatol. 2004, 63, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Yasunaga, J.-I.; Washizaki, A.; Seki, Y.; Kuramitsu, M.; Tan, W.K.; Hu, A.; Okuma, K.; Hamaguchi, I.; Mizukami, T.; et al. Frequent horizontal and mother-to-child transmission may contribute to high prevalence of STLV-1 infection in Japanese macaques. Retrovirology 2020, 17, 15. [Google Scholar] [CrossRef]
- Afonso, P.V.; Fagrouch, Z.; Deijs, M.; Niphuis, H.; Bogers, W.; Gessain, A.; Van Der Hoek, L.; Verschoor, E.J. Absence of accessory genes in a divergent simian T-lymphotropic virus type 1 isolated from a bonnet macaque (Macaca radiata). PLOS Neglected Trop. Dis. 2019, 13, e0007521. [Google Scholar] [CrossRef]
- Merceron, G.; Kaiser, T.M.; Kostopoulos, D.S.; Schulz-Kornas, E. Ruminant diets and the Miocene extinction of European great apes. Proc. R. Soc. B Biol. Sci. 2010, 277, 3105–3112. [Google Scholar] [CrossRef] [Green Version]
- Merceron, G.; Kostopoulos, D.S.; De Bonis, L.; Fourel, F.; Koufos, G.D.; Lécuyer, C.; Martineau, F. Stable isotope ecology of Miocene bovids from northern Greece and the ape/monkey turnover in the Balkans. J. Hum. Evol. 2013, 65, 185–198. [Google Scholar] [CrossRef]
- Nelson, S.V.; Rook, L. Isotopic reconstructions of habitat change surrounding the extinction of Oreopithecus, the last European ape. Am. J. Phys. Anthr. 2016, 160, 254–271. [Google Scholar] [CrossRef]
- Kostopoulos, D.S.; Guy, F.; Kynigopoulou, Z.; Koufos, G.D.; Valentin, X.; Merceron, G. A 2Ma old baboon-like monkey from Northern Greece and new evidence to support the Paradolichopithecus—Procynocephalus synonymy (Primates: Cercopithecidae). J. Hum. Evol. 2018, 121, 178–192. [Google Scholar] [CrossRef]
- Radović, P.; Lindal, J.; Marković, Z.; Alaburić, S.; Roksandic, M. First record of a fossil monkey (Primates, Cercopithecidae) from the Late Pliocene of Serbia. J. Hum. Evol. 2019, 137, 102681. [Google Scholar] [CrossRef]
- Sinha, A.; Johnson, W.E. Retroviruses of the RDR superinfection interference group: Ancient origins and broad host distribution of a promiscuous Env gene. Curr. Opin. Virol. 2017, 25, 105–112. [Google Scholar] [CrossRef]
- Malicorne, S.; Vernochet, C.; Cornelis, G.; Mulot, B.; Delsuc, F.; Heidmann, O.; Heidmann, T.; Dupressoir, A. Genome-Wide Screening of Retroviral Envelope Genes in the Nine-Banded Armadillo (Dasypus novemcinctus, Xenarthra) Reveals an Unfixed Chimeric Endogenous Betaretrovirus Using the ASCT2 Receptor. J. Virol. 2016, 90, 8132–8149. [Google Scholar] [CrossRef] [Green Version]
- Van Der Kuyl, A.C.; Berkhout, B. Viruses in the reproductive tract: On their way to the germ line? Virus Res. 2020, 286, 198101. [Google Scholar] [CrossRef]
- Marin, M.; Tailor, C.S.; Nouri, A.; Kabat, D. Sodium-Dependent neutral amino acid transporter type 1 is an auxiliary receptor for baboon endogenous retrovirus. J. Virol. 2000, 74, 8085–8093. [Google Scholar] [CrossRef] [Green Version]
- Shimode, S.; Nakaoka, R.; Shogen, H.; Miyazawa, T. Characterization of feline ASCT1 and ASCT2 as RD-114 virus receptor. J. Gen. Virol. 2013, 94, 1608–1612. [Google Scholar] [CrossRef]
- Nethe, M.; Berkhout, B.; Van Der Kuyl, A.C. Retroviral superinfection resistance. Retrovirology 2005, 2, 52. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, J.; Sugimoto, M.; Bernstein, H.B.; Jinno, Y.; Schust, D.J. A novel human endogenous retroviral protein inhibits cell-cell fusion. Sci. Rep. 2013, 3, srep01462. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
Figure 1.
Estimated present-day distribution by continent and proposed prehistoric migration routes of eight retroviruses that are, or have been, circulating in Old World monkeys (OWM). Eocene: ~33.9 to 56 mya; Miocene: ~5.3 to 23 mya; Pliocene: ~2.5 to 5.3 mya. The continental positions shown represent the current situation, the time periods shown are estimates. During the Eocene, South America and Africa were much closer to each other. During the Miocene, a junction existed between Africa and Asia due to the closure of the Tethys Seaway.
Figure 1.
Estimated present-day distribution by continent and proposed prehistoric migration routes of eight retroviruses that are, or have been, circulating in Old World monkeys (OWM). Eocene: ~33.9 to 56 mya; Miocene: ~5.3 to 23 mya; Pliocene: ~2.5 to 5.3 mya. The continental positions shown represent the current situation, the time periods shown are estimates. During the Eocene, South America and Africa were much closer to each other. During the Miocene, a junction existed between Africa and Asia due to the closure of the Tethys Seaway.

Figure 2.
Estimated infectious periods of eight retroviruses that are, or have been, circulating in OWM. Putative speciation events in OWM are indicated. The geological time line is not drawn to scale.
Figure 2.
Estimated infectious periods of eight retroviruses that are, or have been, circulating in OWM. Putative speciation events in OWM are indicated. The geological time line is not drawn to scale.
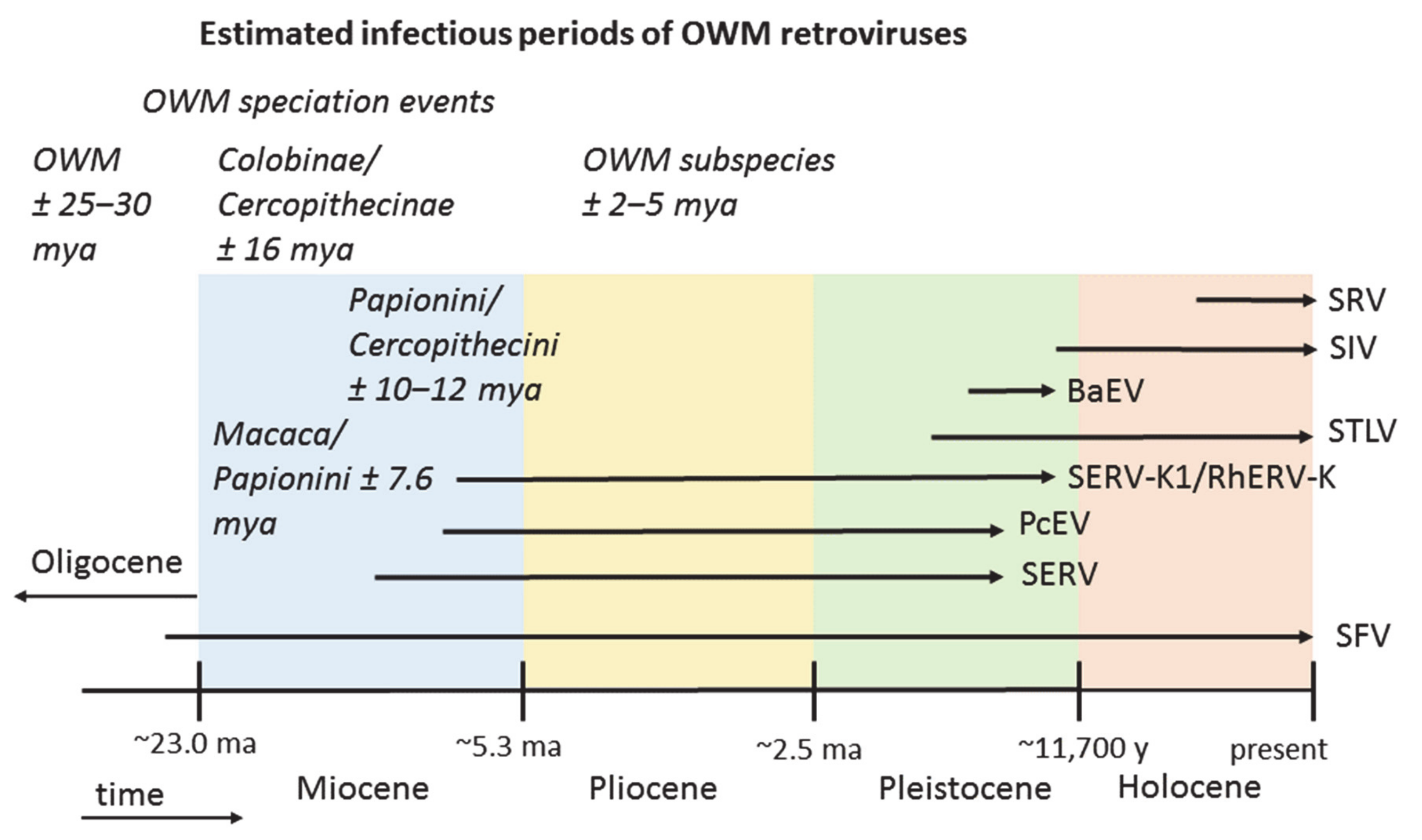

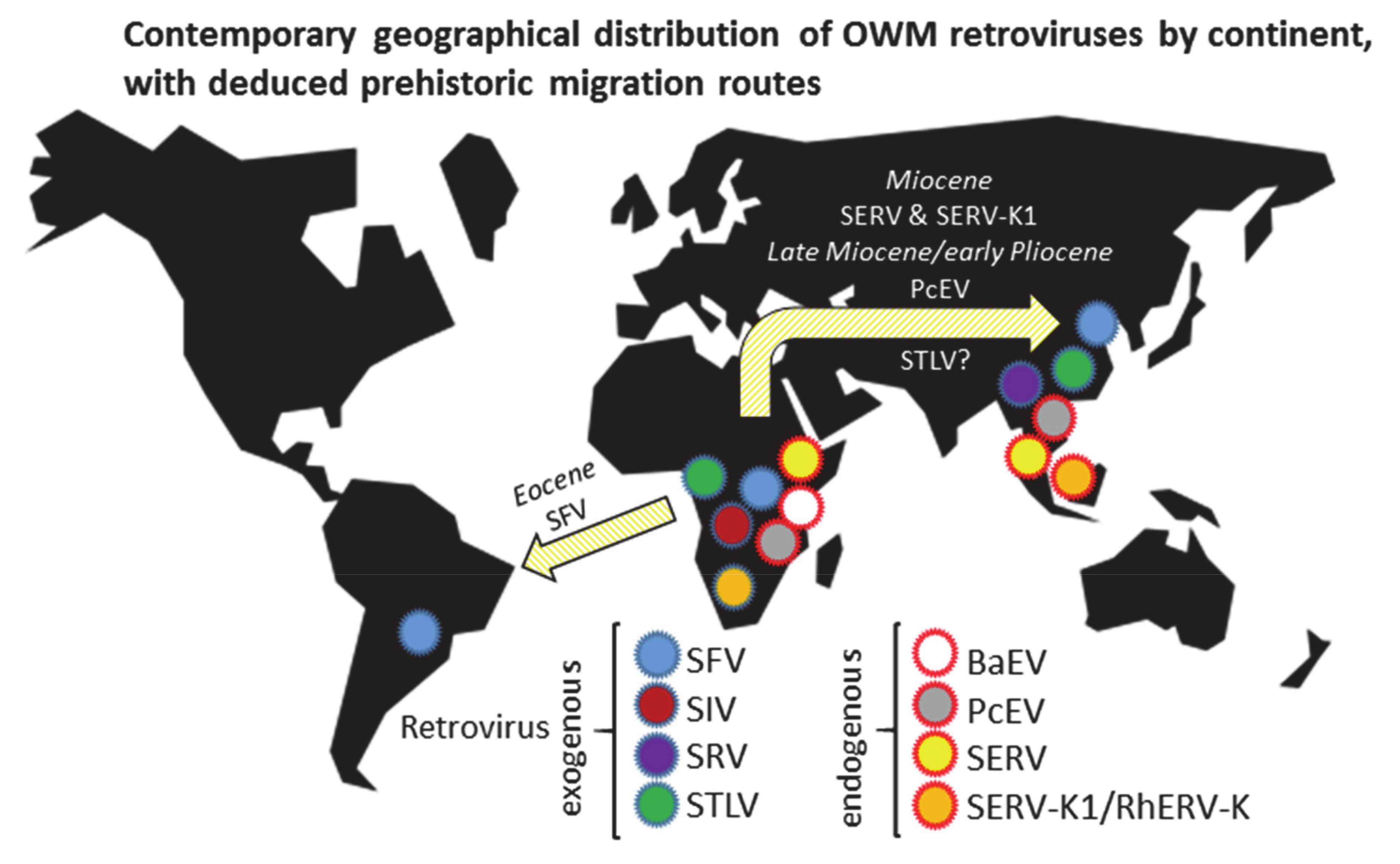{kind=link}
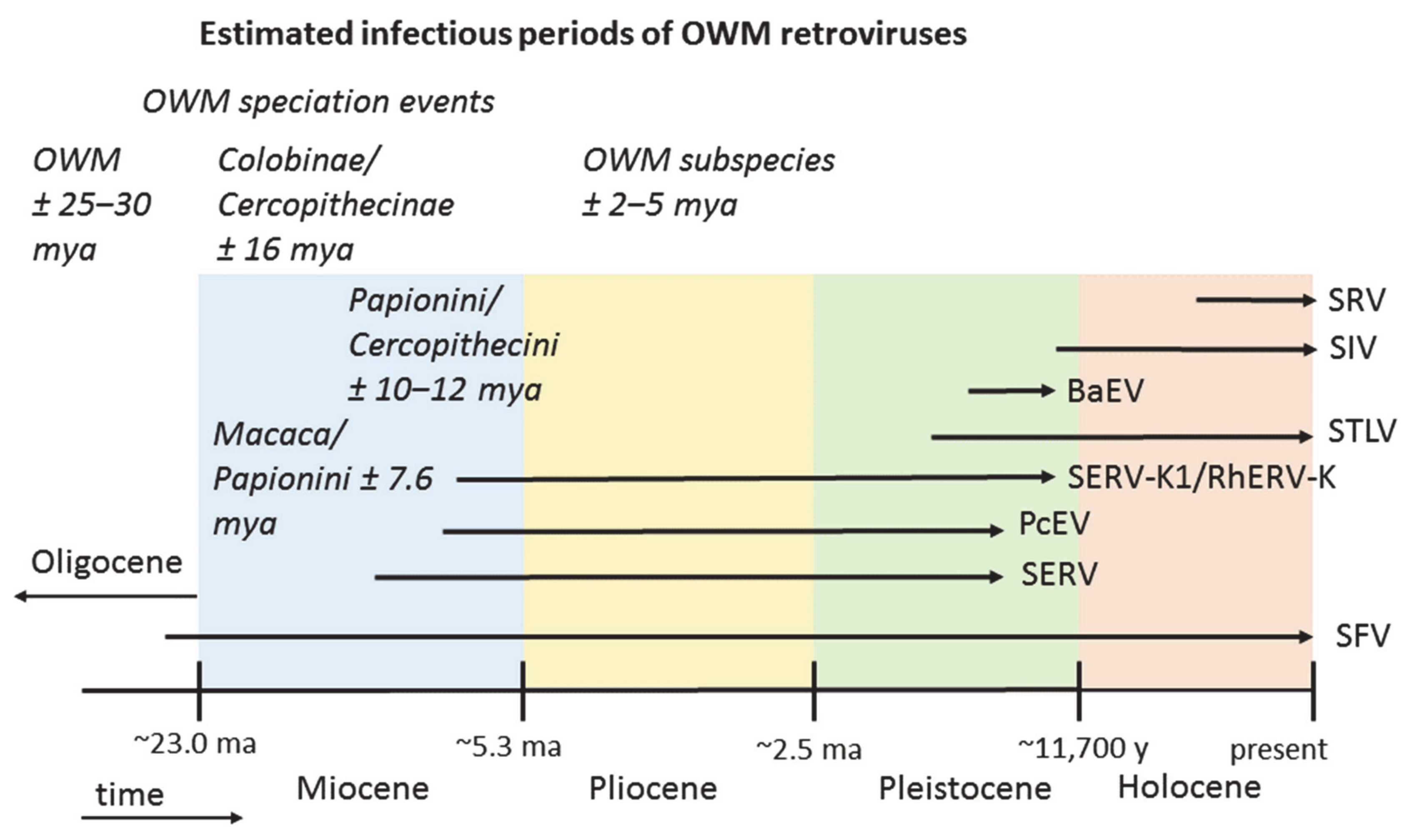{kind=link}
{kind=link}
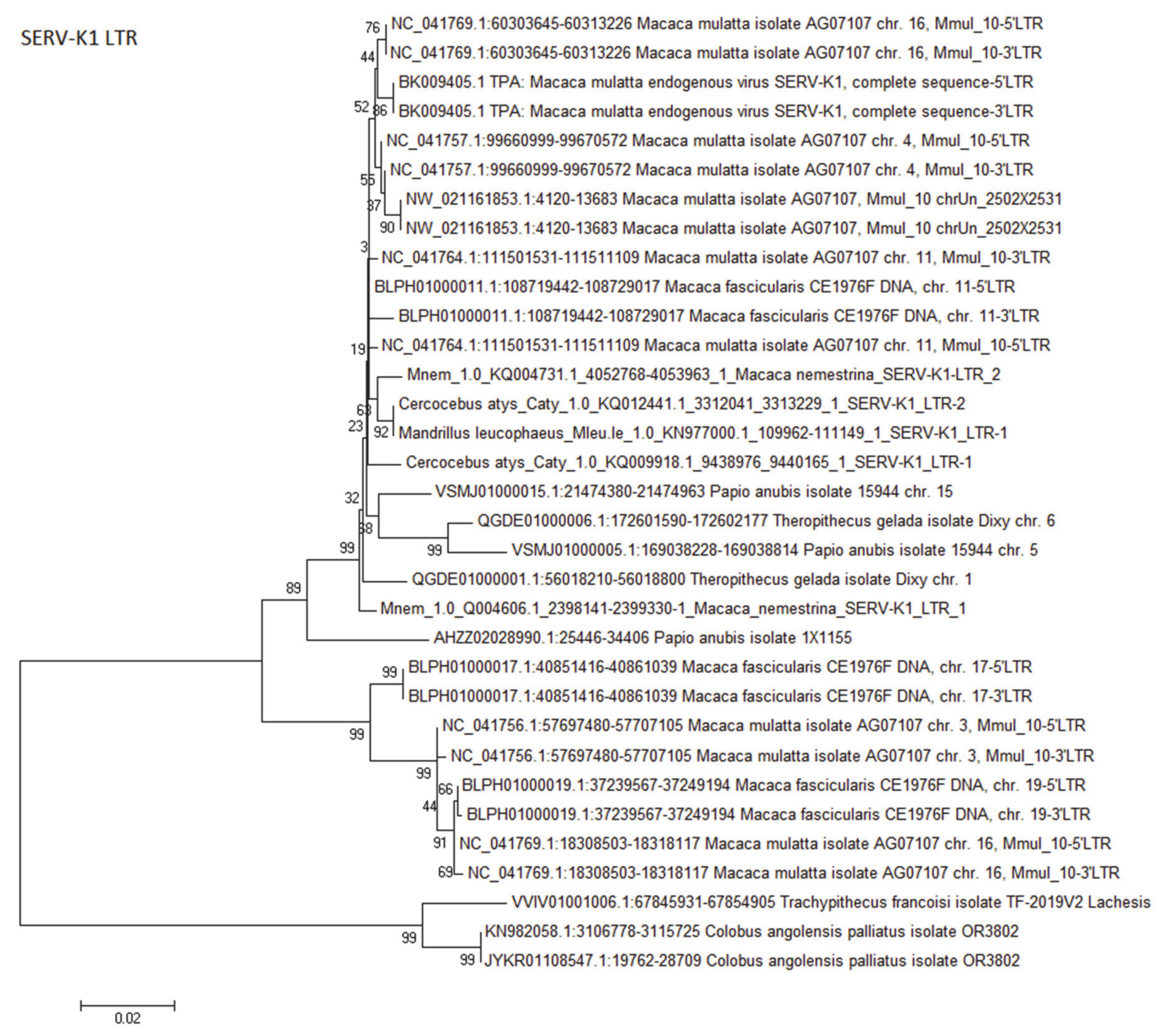{kind=link}
Table 1.
Overview of Old World primate species naturally infected with exogenous retroviruses.
| Retrovirus Species Genus | African OWM Species + | Asian OWM Species + | Other Primate Species + | References | ||
|---|---|---|---|---|---|---|
| Cercopithecinae | Colobinae | Cercopithecinae | Colobinae | |||
| Simian foamy virus (SFV) 1 Simii-spumavirus | Cercopithecini: Chlorocebus Erythrocebus Papionini: Cercocebus Lophocebus Macaca Mandrillus Papio Theropithecus | Colobus Procolobus | Papionini: Macaca | Pygathrix Trachypithecus | Gorilla Hylobates Pan Pongo | [50,53,54,55,56] |
| Simian immuno-deficiency virus (SIV) 2 Lentivirus | Cercopithecini: Allenopithecus Cercopithecus Chlorocebus Miopithecus Papionini: Cercocebus Lophocebus Mandrillus | Colobus Piliocolobus Procolobus | None | None | Gorilla Homo (HIV) Pan | [54,57,58,59,60,61,62] |
| Simian type D retrovirus 3 (SRV) Betaretrovirus | None | None | Papionini: Macaca | Semnopithecus? | None | [63,64,65,66,67] |
| Simian T-lymphotropic virus (STLV) Deltavirus | Cercopithecini: Allenopithecus Cercopithecus Chlorocebus Erythrocebus Miopithecus Papionini: Cercocebus Lophocebus Macaca Mandrillus Papio | Piliocolobus Procolobus | Papionini: Macaca | Presbytis | Gorilla Homo (HTLV) Hylobates Pan Pongo | [22,24,54,68,69,70,71,72,73,74,75,76,77] |
1 SFV is also widespread in New World monkeys (NWM) species [51,55,78]. 2 Three natural infections of a yellow, an olive and a chacma baboon with a Chlorocebus (African green monkey, AGM) SIV strain, respectively, have been reported [79,80,81]. SIV isolated from an Erythrocebus monkey likely also results from SIVagm cross-species transmission [82]. 3 Simian type D betaretroviruses are recombinants with a betaretroviral gag-pol sequence and a gammaretrovirus env gene.
Table 2.
Overview of OWM species harboring OWM-specific endogenous retrovirus genomes.
| Retrovirus Species Genus | African OWM Species + | Asian OWM Species + | References | ||
|---|---|---|---|---|---|
| Cercopithecinae | Colobinae | Cercopithecinae | Colobinae | ||
| Baboon endogenous virus (BaEV) Gammaretrovirus | Cercopithecini: Chlorocebus Papionini: Cercocebus Mandrillus Lophocebus Papio Theropithecus | None | None | None | [130] Figure A1 |
| Papio cynocephalus endogous virus (PcEV) Gammaretrovirus | Papionini: Lophocebus Papio Theropithecus | Colobus | Papionini: Macaca | None | [131] Figure A1 |
| Simian endogenous retrovirus (SERV) Betaretrovirus (Simian type D) | Cercopithecini: Cercopithecus Chlorocebus Erythrocebus Miopithecus Papionini: Macaca | None | Papionini: Macaca | Rhinopithecus | [38,132,133] Figure A1 |
| Simian endogenous retrovirus-K1 (SERV-K1/ RhERV-K) Betaretrovirus | Papionini: Cercocebus Mandrillus Papio Theropithecus | Colobus | Papionini: Macaca | Trachypithecus | Figure A2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
van der Kuyl, A.C. Contemporary Distribution, Estimated Age, and Prehistoric Migrations of Old World Monkey Retroviruses. Epidemiologia 2021, 2, 46-67. https://0-doi-org.brum.beds.ac.uk/10.3390/epidemiologia2010005
AMA Style
van der Kuyl AC. Contemporary Distribution, Estimated Age, and Prehistoric Migrations of Old World Monkey Retroviruses. Epidemiologia. 2021; 2(1):46-67. https://0-doi-org.brum.beds.ac.uk/10.3390/epidemiologia2010005
Chicago/Turabian Stylevan der Kuyl, Antoinette C. 2021. "Contemporary Distribution, Estimated Age, and Prehistoric Migrations of Old World Monkey Retroviruses" Epidemiologia 2, no. 1: 46-67. https://0-doi-org.brum.beds.ac.uk/10.3390/epidemiologia2010005