Unveiling the Influence of Non-Toxic Fluorinated Ionic Liquids Aqueous Solutions in the Encapsulation and Stability of Lysozyme



Abstract
:1. Introduction
2. Materials and Experimental Methodology
2.1. Materials
2.2. Cytotoxicity Assays
2.3. Preparation of Solutions for Protein Assays
2.4. UV–Visible Spectrophotometry
2.5. Encapsulation Efficiency
2.6. Lysozyme Activity
2.7. Differential Scanning Calorimetry (DSC)
3. Results and Discussion
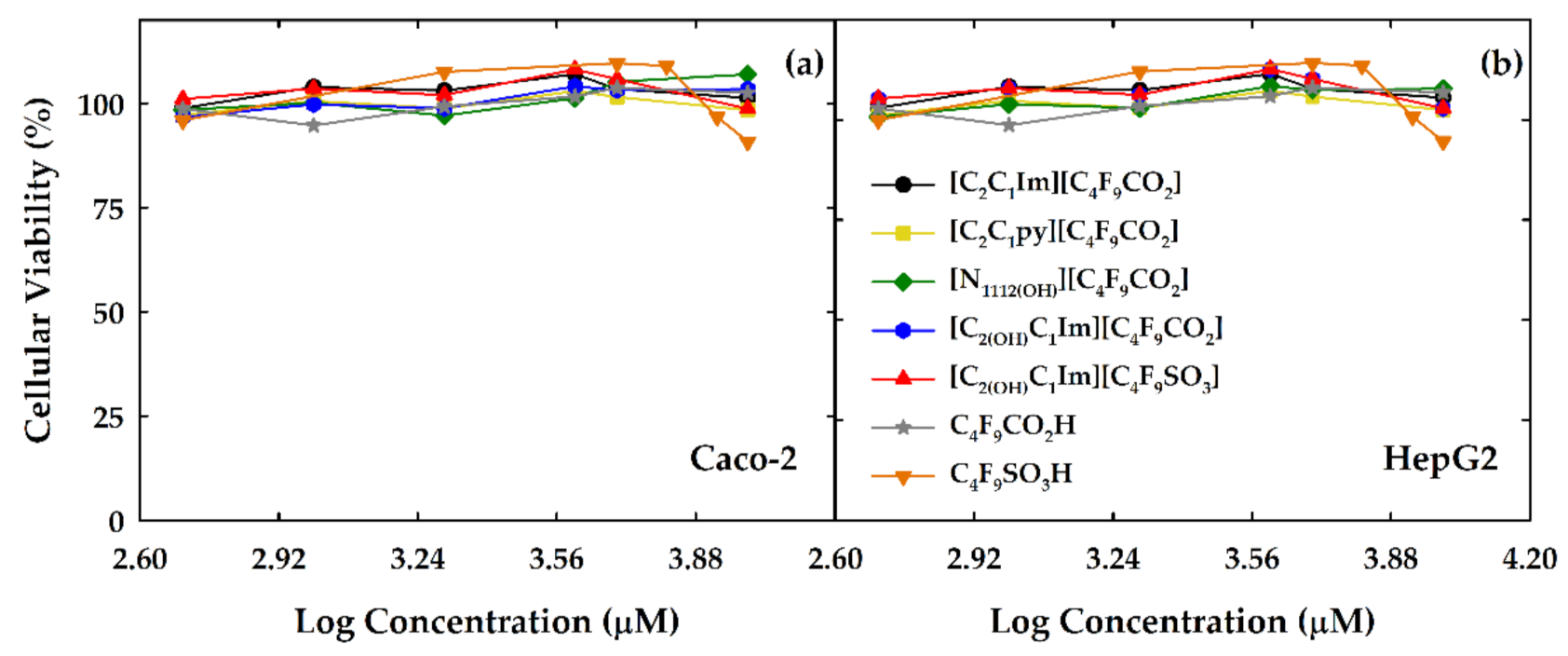
3.1. Cytotoxicity of FILs
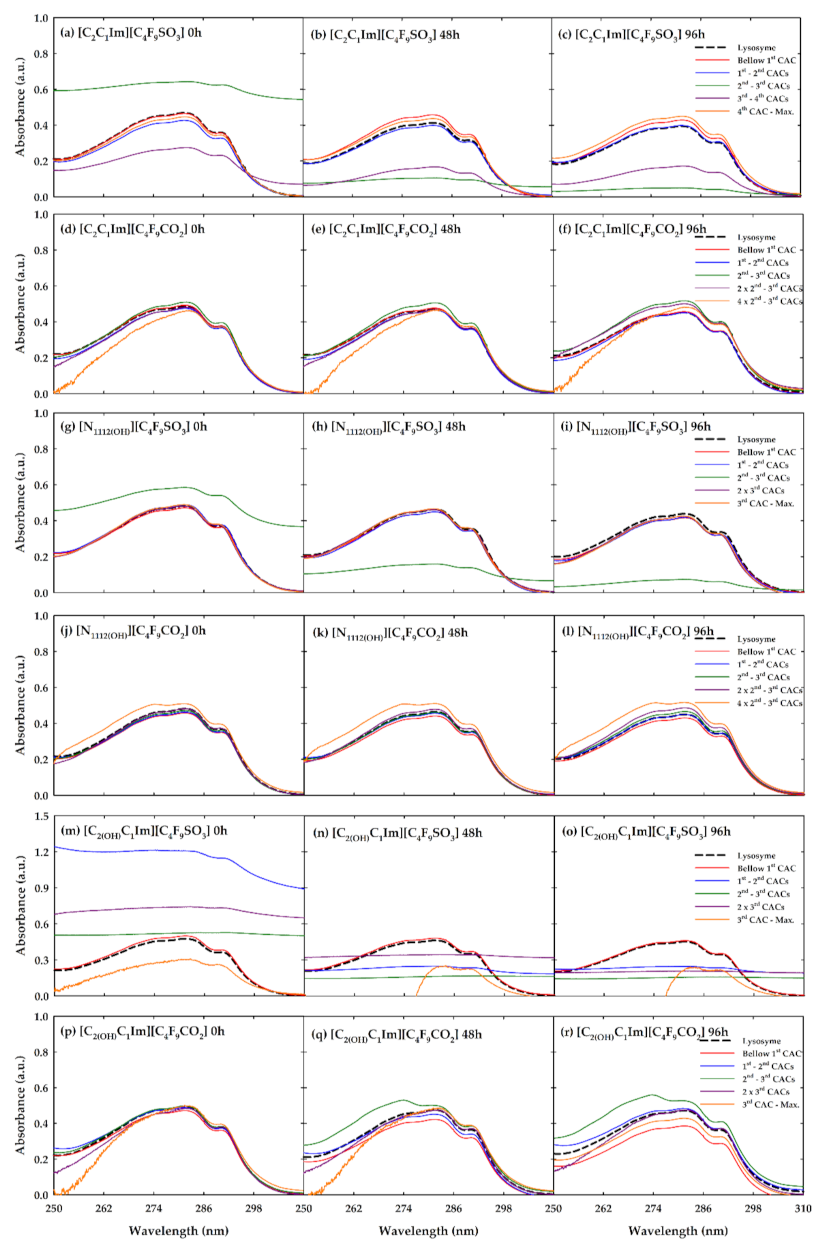
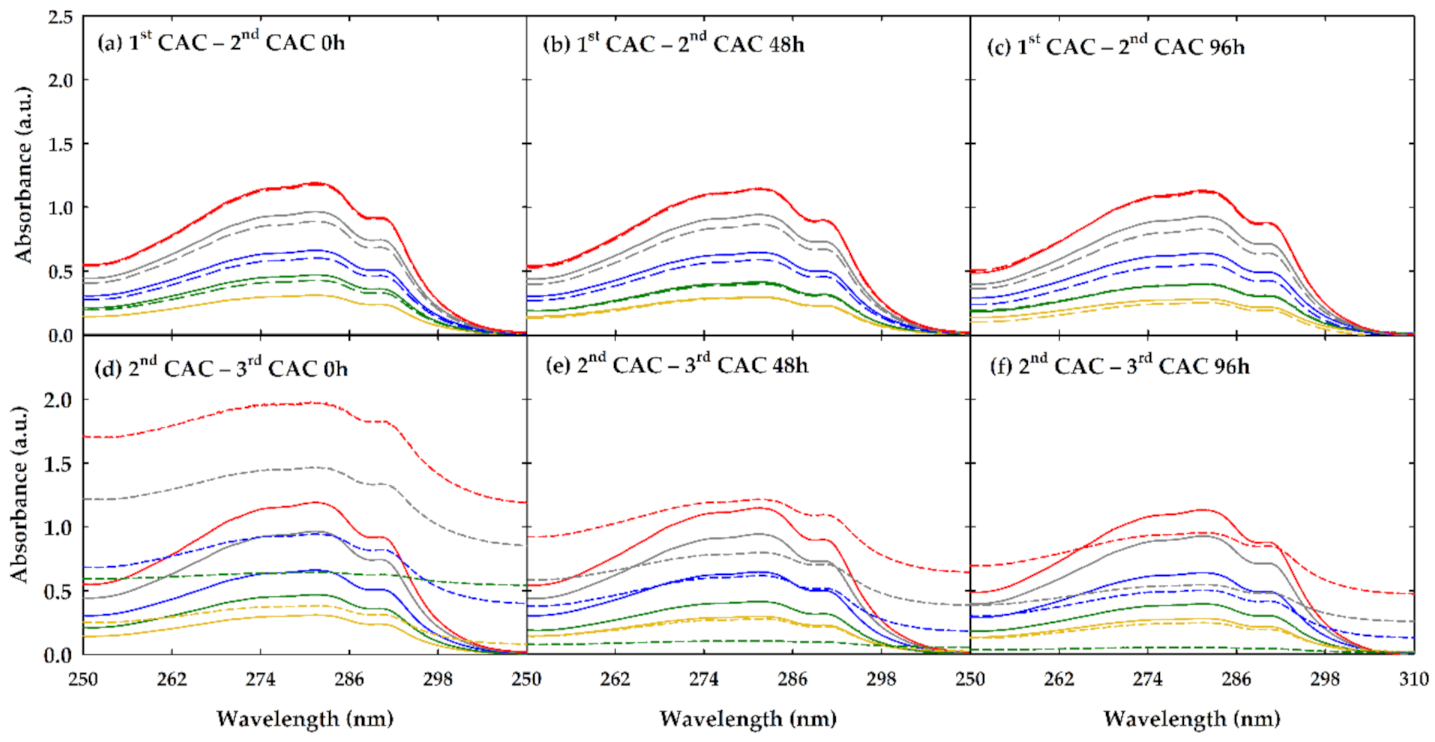
3.2. Absorption Measurements
3.3. Encapsulation of Lysozyme by FILs
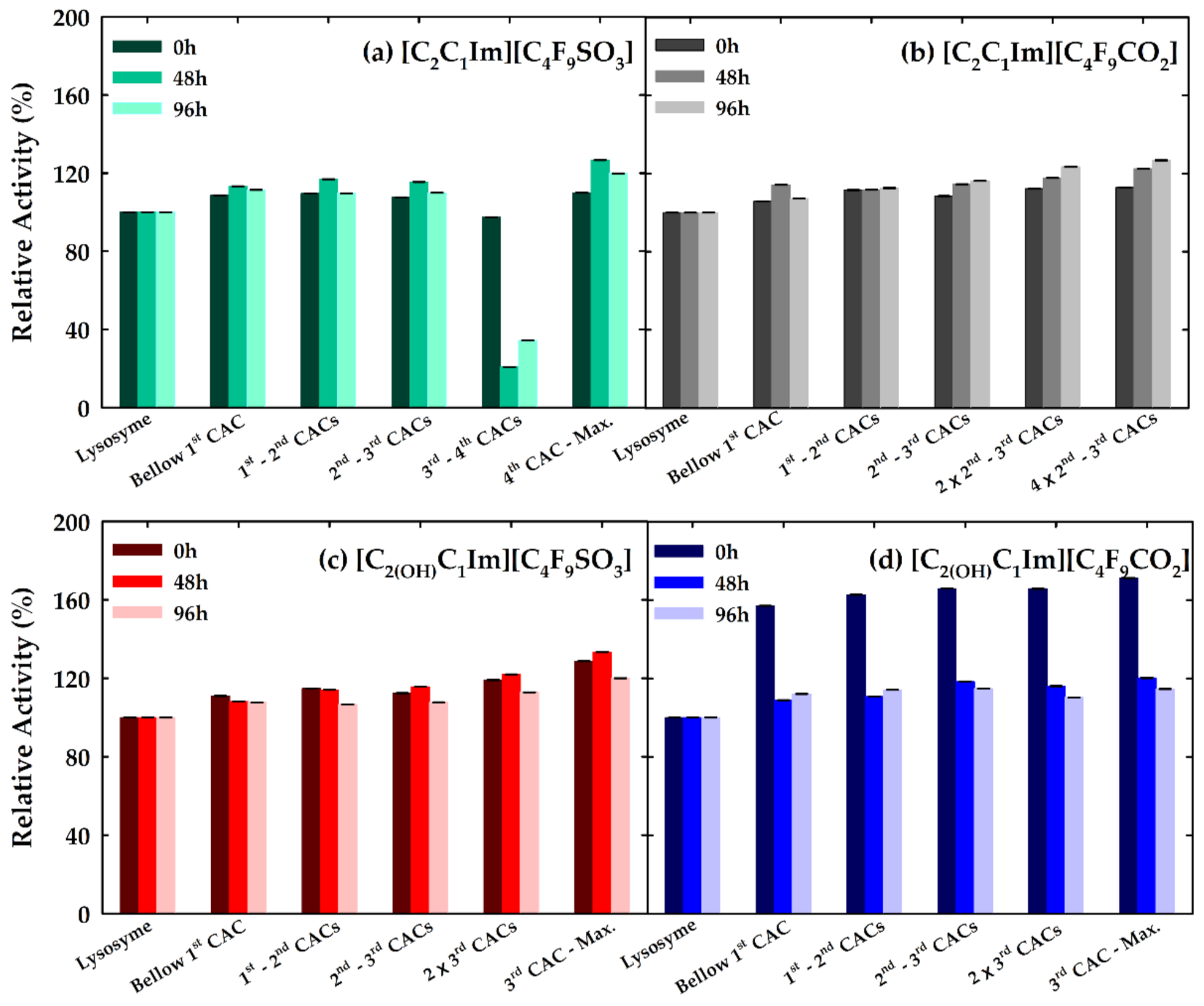
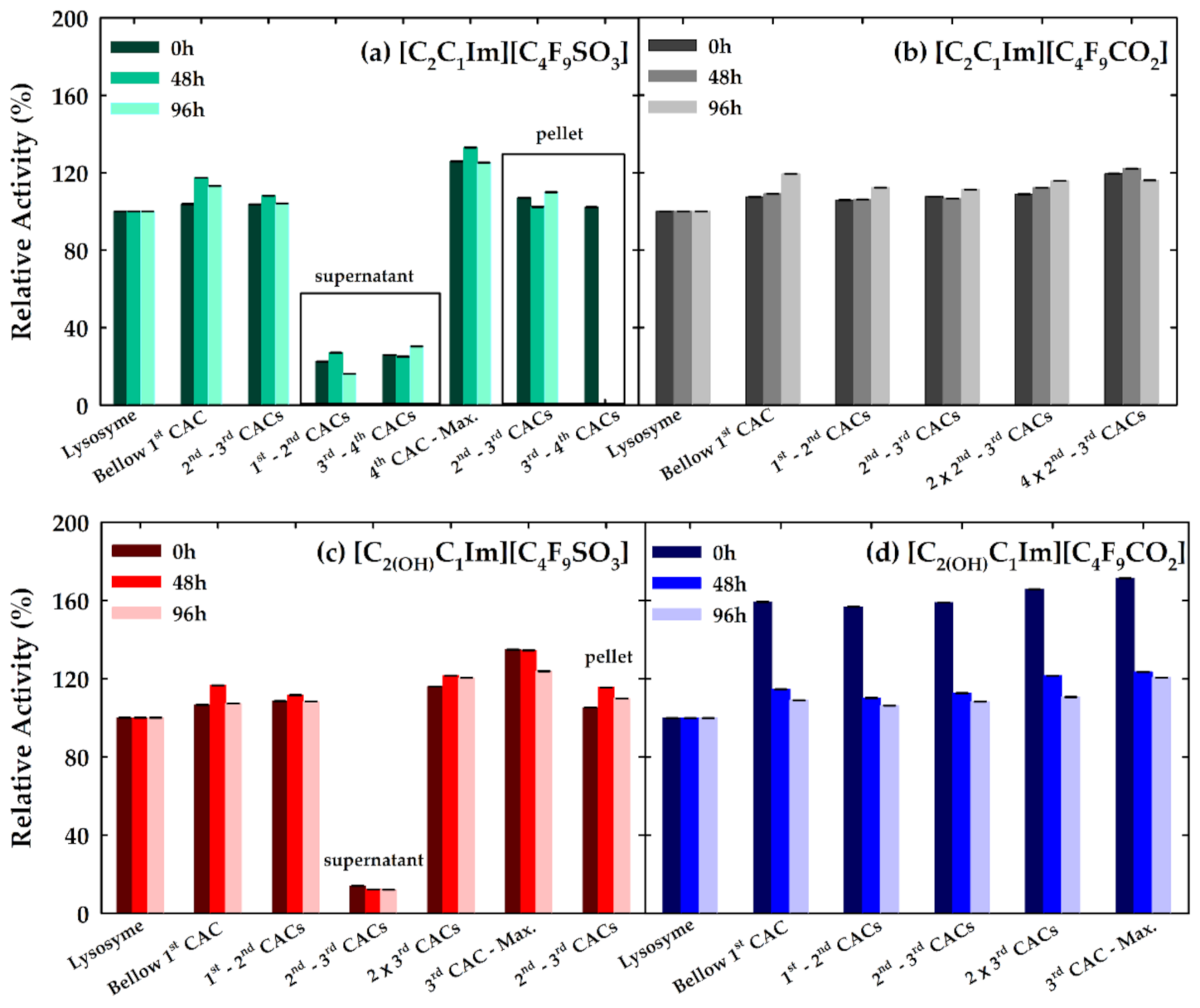
3.4. Effect of FILs on the Bioactivity of Lysozyme
3.5. Influence of FILs in the Thermal Stability of Lysozyme
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Balcão, V.M.; Vila, M.M.D.C. Structural and functional stabilization of protein entities: State-of-the-art. Adv. Drug Deliv. Rev. 2015, 93, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Bisht, M.; Venkatesu, P. Biocompatibility of ionic liquids towards protein stability: A comprehensive overview on the current understanding and their implications. Int. J. Biol. Macromol. 2017, 96, 611–651. [Google Scholar] [CrossRef] [PubMed]
- Schröder, C. Proteins in ionic liquids: Current status of experiments and simulations. Top Curr. Chem. 2017, 375, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, L.K.; Pereira, J.F.B.; Campos, W.F.; Silva, E.C.; Moutinho, C.G.; Vila, M.M.D.C.; Oliveira, J.M.; Teixeira, J.A.; Balcão, V.M.; Tubino, M. Insights into protein-ionic liquid interactions aiming at macromolecule delivery systems. J. Brazil. Chem. Soc. 2018, 29, 1983–1998. [Google Scholar] [CrossRef]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process. Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Callewaert, L.; Michiels, C.W. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef]
- Singh, G.; Kaur, M.; Singh, D.; Kesavan, A.K.; Kang, T.S. Antimicrobial colloidal complexes of lysozyme with bio-based surface active ionic liquids in aqueous medium. J. Phys. Chem. B 2020, 124, 3791–3800. [Google Scholar] [CrossRef]
- Egorova, K.S.; Gordeev, E.G.; Ananikov, V.P. Biological activity of ionic liquids and their application in pharmaceutics and medicine. Chem. Rev. 2017, 117, 7132–7189. [Google Scholar] [CrossRef] [PubMed]
- Schindl, A.; Hagen, M.L.; Muzammal, S.; Gunasekera, H.A.D.; Croft, A.K. Proteins in ionic liquids: Reactions, applications, and futures. Front. Chem. 2019, 7, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisht, M.; Kumar, A.; Venkatesu, P. Analysis of the driving force that rule the stability of lysozyme in alkylammonium-based ionic liquids. Int. J. Biol. Macromol. 2015, 81, 1074–1081. [Google Scholar] [CrossRef]
- Satish, L.; Rana, S.; Arakha, M.; Rout, L.; Ekka, B.; Jha, S.; Dash, P.; Sahoo, H. Impact of imidazolium-based ionic liquids on the structure and stability of lysozyme. Spectrosc. Lett. 2016, 49, 383–390. [Google Scholar] [CrossRef]
- Mandal, B.; Mondal, S.; Pan, A.; Moulik, S.P.; Ghosh, S. Physicochemical study of the interaction of lysozyme with surface active ionic liquid 1-butyl-3-methylimidazolium octylsulfate [BMIM] [OS] in aqueous and buffer media. Coll. Surf. A 2015, 484, 345–353. [Google Scholar] [CrossRef]
- Kumari, M.; Dohare, N.; Maurya, N.; Dohare, R.; Patel, R. Effect of 1-methyl-3-octyleimmidazolium chloride on the stability and activity of lysozyme: A spectroscopic and molecular dynamics studies. J. Biomol. Struct. Dyn. 2017, 35, 2016–2030. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Singh, U.K.; Beg, I.; Alanazi, A.M.; Khan, A.A.; Patel, R. Effect of cations and anions of ionic liquids on the stability and activity of lysozyme: Concentration and temperature effect. J. Mol. Liq. 2018, 272, 253–263. [Google Scholar] [CrossRef]
- Rather, M.A.; Dar, T.A.; Singh, L.R.; Rather, G.M.; Bhat, M.A. Structural-functional integrity of lysozyme in imidazolium based surface active ionic liquids. Int. J. Biol. Macromol. 2020, 156, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kaur, M.; Kaur, H.; Kang, T.S. Synthesis and complexation of a new caffeine based surface active ionic liquid with lysozyme in aqueous medium: Physicochemical, computational and antimicrobial studies. J. Mol. Liq. 2021, 325, 115156. [Google Scholar] [CrossRef]
- Pereiro, A.B.; Araújo, J.M.M.; Martinho, S.; Alves, F.; Nunes, S.; Matias, A.; Duarte, C.M.M.; Rebelo, L.P.N.; Marrucho, I.M. Fluorinated ionic liquids: Properties and applications. ACS Sustain. Chem. Eng. 2013, 1, 427–439. [Google Scholar] [CrossRef]
- Ferreira, M.L.; Pastoriza-Gallego, M.J.; Araújo, J.M.M.; Canongia Lopes, J.N.; Rebelo, L.P.N.; Piñeiro, M.M.; Shimizu, K.; Pereiro, A.B. Influence of nanosegregation on the phase behavior of fluorinated ionic liquids. J. Phys. Chem. C 2017, 121, 5415–5427. [Google Scholar] [CrossRef]
- Pereiro, A.B.; Pastoriza-Gallego, M.J.; Shimizu, K.; Marrucho, I.M.; Canongia Lopes, J.N.; Piñeiro, M.M.; Rebelo, L.P.N. On the formation of a third, nanostructured domain in ionic liquids. J. Phys. Chem. B 2013, 117, 10826–10833. [Google Scholar] [CrossRef]
- Teixeira, F.S.; Vieira, N.S.M.; Cortes, O.A.; Araújo, J.M.M.; Marrucho, I.M.; Rebelo, L.P.N.; Pereiro, A.B. Phase equilibria and surfactant behavior of fluorinated ionic liquids with water. J. Chem. Thermodyn. 2015, 82, 99–107. [Google Scholar] [CrossRef]
- Pereiro, A.B.; Araújo, J.M.M.; Teixeira, F.S.; Marrucho, I.M.; Piñeiro, M.M.; Rebelo, L.P.N. Aggregation behavior and total miscibility of fluorinated ionic liquids in water. Langmuir 2015, 31, 1283–1295. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Bastos, J.C.; Hermida-Merino, C.; Pastoriza-Gallego, M.J.; Rebelo, L.P.N.; Piñeiro, M.M.; Araújo, J.M.M.; Pereiro, A.B. Aggregation and phase equilibria of fluorinated ionic liquids. J. Mol. Liq. 2019, 285, 386–396. [Google Scholar] [CrossRef]
- Ferreira, M.L.; Araújo, J.M.M.; Vega, L.F.; Llovell, F.; Pereiro, A.B. Functionalization of fluorinated ionic liquids: A combined experimental-theoretical study. J. Mol. Liq. 2020, 302, 112489. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Stolte, S.; Araújo, J.M.M.; Rebelo, L.P.N.; Pereiro, A.B.; Markiewicz, M. Acute aquatic toxicity and biodegradability of fluorinated ionic liquids. ACS Sustain. Chem. Eng. 2019, 7, 3733–3741. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Bastos, J.C.; Rebelo, L.P.N.; Matias, A.; Araújo, J.M.M.; Pereiro, A.B. Human cytotoxicity and octanol/water partition coefficients of fluorinated ionic liquids. Chemosphere 2019, 216, 576–586. [Google Scholar] [CrossRef]
- Alves, M.; Vieira, N.S.M.; Rebelo, L.P.N.; Araújo, J.M.M.; Pereiro, A.B.; Archer, M. Fluorinated ionic liquids for protein drug delivery systems: Investigating their impact on the structure and function of lysozyme. Int. J. Pharmaceut. 2017, 526, 309–320. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Castro, P.J.; Marques, D.F.; Araújo, J.M.M.; Pereiro, A.B. Tailor-made fluorinated ionic liquids for protein delivery. Nanomaterials 2020, 10, 1594. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.M.S.; Araújo, J.M.M.; Martins, I.C.; Pereiro, A.B.; Archer, M. Insights into the interaction of bovine serum albumin with surface-active ionic liquids in aqueous solution. J. Mol. Liq. 2021, 322, 114537. [Google Scholar] [CrossRef]
- Fukumoto, K.; Yoshizawa, M.; Ohno, H. Room temperature ionic liquids from 20 natural amino acids. J. Am. Chem. Soc. 2005, 127, 2398–2399. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Reis, P.M.; Shimizu, K.; Cortes, O.A.; Marrucho, I.M.; Araújo, J.M.M.; Esperança, J.M.S.S.; Lopes, J.N.C.; Pereiro, A.B.; Rebelo, L.P.N. A thermophysical and structural characterization of ionic liquids with alkyl and perfluoroalkyl side chains. RSC Adv. 2015, 5, 65337–65350. [Google Scholar] [CrossRef]
- Araújo, J.M.M.; Florindo, C.; Pereiro, A.B.; Vieira, N.S.M.; Matias, A.A.; Duarte, C.M.M.; Rebelo, L.P.N.; Marrucho, I.M. Cholinium-based ionic liquids with pharmaceutically active anions. RSC Adv. 2014, 4, 28126–28132. [Google Scholar] [CrossRef]
- Vieira, N.S.M.; Luís, A.; Reis, P.M.; Carvalho, P.J.; Lopes da Silva, J.A.; Esperança, J.M.S.S.; Araújo, J.M.M.; Rebelo, L.P.N.; Freire, M.G.; Pereiro, A.B. Fluorination effects on the thermodynamic, thermophysical and surface properties of ionic liquids. J. Chem. Thermodyn. 2016, 97, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Petkovic, M.; Seddon, K.R.; Rebelo, L.P.N.; Pereira, C.S. Ionic liquids: A pathway to environmental acceptability. Chem. Soc. Rev. 2011, 40, 1383–1403. [Google Scholar] [CrossRef] [PubMed]
- Patinha, D.J.S.; Tomé, L.C.; Florindo, C.; Soares, H.R.; Coroadinha, A.S.; Marrucho, I.M. New low-toxicity cholinium-based ionic liquids with perfluoroalkanoate anions for aqueous biphasic system implementation. ACS Sustain. Chem. Eng. 2016, 4, 2670–2679. [Google Scholar] [CrossRef]
- García-Lorenzo, A.; Tojo, E.; Tojo, J.; Teijeira, M.; Rodríguez-Berrocal, F.J.; González, M.P.; Martínez-Zorzano, V.S. Cytotoxicity of selected imidazolium-derived ionic liquids in the human Caco-2 cell line. Sub-structural toxicological interpretation through a QSAR study. Green Chem. 2008, 10, 508–516. [Google Scholar] [CrossRef]
- Matias, A.; Nunes, S.L.; Poejo, J.; Mecha, E.; Serra, A.T.; Madeira, P.J.A.; Bronze, M.R.; Duarte, C.M.M. Antioxidant and anti-inflammatory activity of a flavonoid-rich concentrate recovered from Opuntia ficus-indica juice. Food Funct. 2014, 5, 3269–3280. [Google Scholar] [CrossRef] [PubMed]
- Wilkening, S.; Stahl, F.; Bader, A. Comparison of primary human hepatocytes and hepatoma cell line Hepg2 with regard to their biotransformation properties. Drug Metab. Dispos. 2003, 31, 1035–1042. [Google Scholar] [CrossRef]
- Gorrochategui, E.; Pérez-Albaladejo, E.; Casas, J.; Lacorte, S.; Porte, C. Perfluorinated chemicals: Differential toxicity, inhibition of aromatase activity and alteration of cellular lipids in human placental cells. Toxicol. Appl. Pharm. 2014, 277, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Lim, X. Tainted water: The scientists tracing thousands of fluorinated chemicals in our environment. Nature 2019, 566, 26–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segawa, S.I.; Sugihara, M. Characterization of the transition state of lysozyme unfolding. II. Effects of the intrachain crosslinking and the inhibitor binding on the transition state. Biopolymers 1984, 23, 2489–2498. [Google Scholar] [CrossRef]
- Vrikkis, R.M.; Fraser, K.J.; Fujita, K.; MacFarlane, D.R.; Elliott, G.D. Biocompatible ionic liquids: A new approach for stabilizing proteins in liquid formulation. J. Biomech. Eng. 2009, 131, 074514. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cations | |
|---|---|
 1-Ethyl-3-methylimidazolium [C2C1Im]+ |  (2-Hydroxyethyl)trimethylammonium [N1112(OH)]+ |
 1-Ethyl-3-methylimidazolium [C2C1py]+ |  1-(2-Hydroxyethyl)-3-methylimidazolium [C2(OH)C1Im]+ |
| Anions | |
 Perfluorobutanesulfonate [C4F9SO3]‾ |  Perfluoropentanoate [C4F9CO2]‾ |
| Acids | |
 Perfluorobutanesulfonic acid C4F9SO3H |  Perfluoropentanoic acid C4F9CO2H |
| FILs | Below 1st CAC | 1st–2nd CACs | 2nd–3rd CACs | 3rd–4th CACs | Above 4th CAC |
|---|---|---|---|---|---|
| [C2C1Im][C4F9SO3] | 0.0030 | 0.0109 | 0.0245 | 0.0384 | 0.3636 |
| [C2C1Im][C4F9CO2] | 0.0039 | 0.0184 | 0.0784 | 0.1567 | 0.3134 |
| [C2C1py][C4F9CO2] | 0.0029 | 0.0089 | 0.0222 | 0.0975 | 0.3582 |
| [N1112(OH)][C4F9CO2] | 0.0017 | 0.0297 | 0.0843 | 0.1686 | 0.3372 |
| [N1112(OH)][C4F9SO3] | 0.0033 | 0.0104 | 0.0447 | 0.1502 | 0.3100 |
| [C2(OH)C1Im][C4F9SO3] | 0.0042 | 0.0230 | 0.0456 | 0.1072 | 0.3065 |
| [C2(OH)C1Im][C4F9CO2] | 0.0036 | 0.0159 | 0.0356 | 0.0928 | 0.1788 |
| FILs | Time | Below 1st CAC | 1st–2nd CACs | 2nd–3rd CACs | 3rd–4th CACs | Above 4th CAC |
|---|---|---|---|---|---|---|
| [C2C1Im][C4F9SO3] | 0 h | 1 | 1 | 72.1 ± 3.4 2 | 68.7 ± 19.5 2 | 1 |
| 48 h | 86.3 ± 0.8 2 | 1 | ||||
| 96 h | 75.5 ± 0.1 2 | |||||
| [C2(OH)C1Im][C4F9SO3] | 0 h | 1 | 1 | 78.4 ± 3.9 2 | 1 | 1 |
| 48 h | 80.5 ± 4.4 2 | |||||
| 96 h | 82.3 ± 8.5 2 |
| FILs | 0 | Below 1st CAC | 1st–2nd CACs | 2nd–3rd CACs | 3rd–4th CACs | |
| [C2C1Im][C4F9SO3] | Tm (°C) | 74.31 | 70.67 | 1 | 73.69 | 72.97 |
| ΔH (kJ/mol) | 526.9 | 363.1 | 1 | 458.0 | 434.6 | |
| [C2(OH)C1Im][C4F9SO3] | Tm (°C) | 74.31 | 67.55 | 73.92 | 72.42 | 1 |
| ΔH (kJ/mol) | 526.9 | 337.1 | 543.2 | 456.3 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, M.L.; Vieira, N.S.M.; Araújo, J.M.M.; Pereiro, A.B. Unveiling the Influence of Non-Toxic Fluorinated Ionic Liquids Aqueous Solutions in the Encapsulation and Stability of Lysozyme. Sustain. Chem. 2021, 2, 149-166. https://0-doi-org.brum.beds.ac.uk/10.3390/suschem2010010
Ferreira ML, Vieira NSM, Araújo JMM, Pereiro AB. Unveiling the Influence of Non-Toxic Fluorinated Ionic Liquids Aqueous Solutions in the Encapsulation and Stability of Lysozyme. Sustainable Chemistry. 2021; 2(1):149-166. https://0-doi-org.brum.beds.ac.uk/10.3390/suschem2010010
Chicago/Turabian StyleFerreira, Margarida L., Nicole S. M. Vieira, João M. M. Araújo, and Ana B. Pereiro. 2021. "Unveiling the Influence of Non-Toxic Fluorinated Ionic Liquids Aqueous Solutions in the Encapsulation and Stability of Lysozyme" Sustainable Chemistry 2, no. 1: 149-166. https://0-doi-org.brum.beds.ac.uk/10.3390/suschem2010010