Are Structurally Modified Galactomannan Derivatives Biologically Active?


 , , ,
, , ,  and
and
Abstract
:1. Introduction
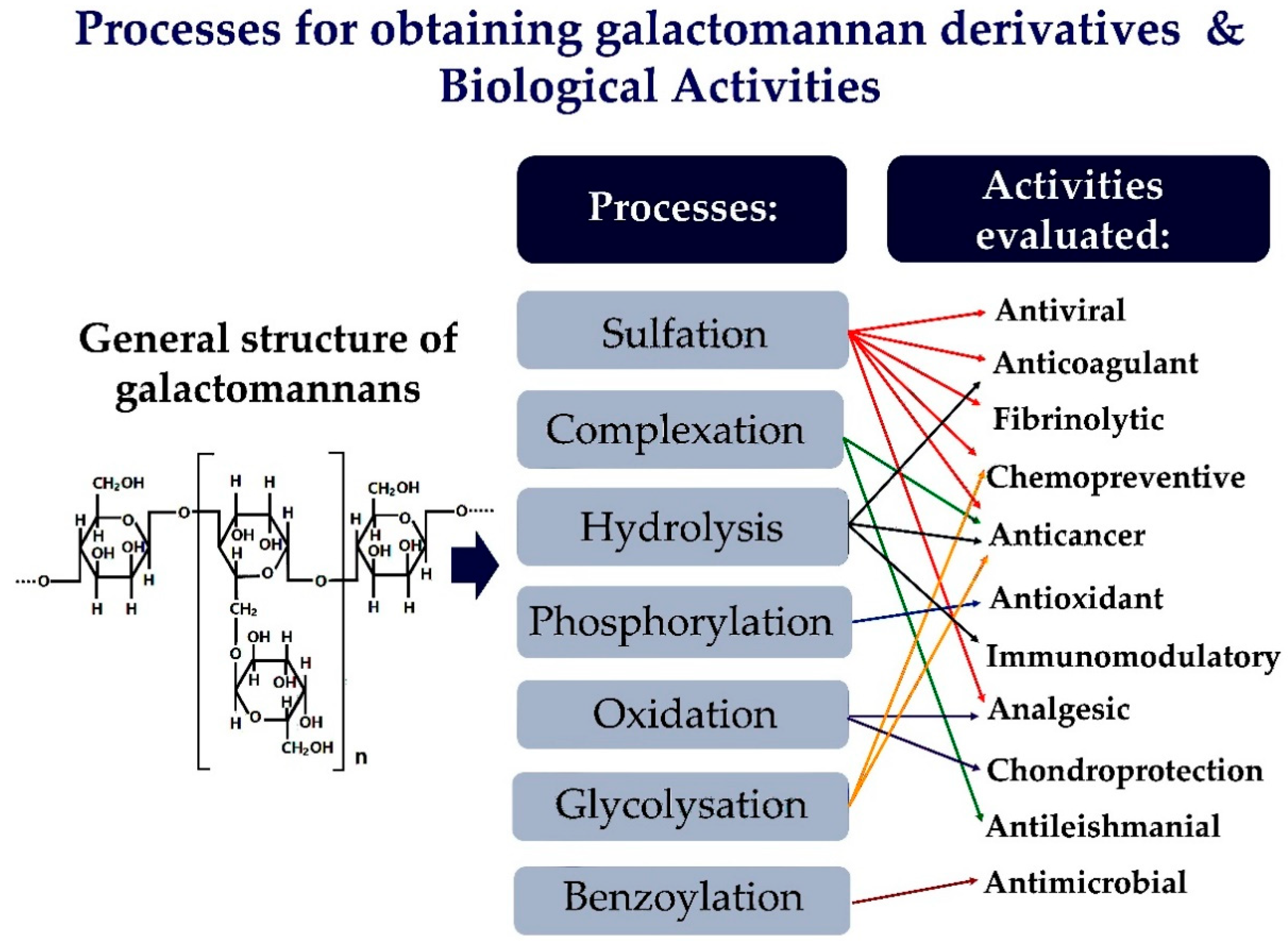
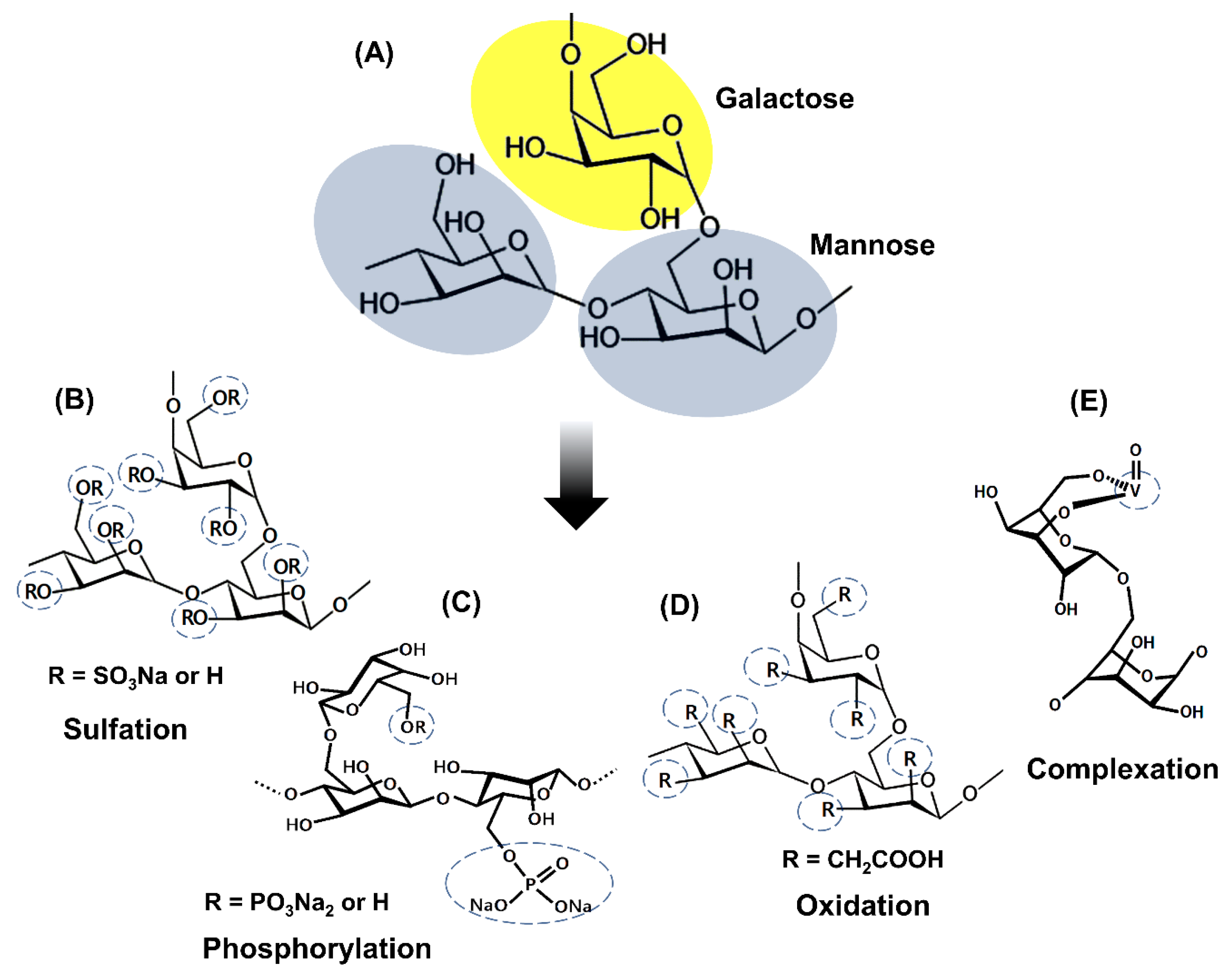
2. Strategies for Obtaining Galactomannan Derivatives
3. Pharmacological Activities of Galactomannan Derivatives
3.1. Antiviral Activity
3.2. Antimicrobial Activity
3.3. Anticoagulant and Fibrinolytic Activities
3.4. Chemopreventive Activity
3.5. Anticancer Activity
3.6. Antioxidant Activity
3.7. Analgesia and Chondroprotection Activities
3.8. Immunomodulatory Activity
3.9. Antileishmanial Activity
4. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rossi, B.; Campia, P.; Merlini, L.; Brasca, M.; Pastori, N.; Farris, S.; Melone, L.; Punta, C.; Galante, Y.M. An aerogel obtained from chemo-enzymatically oxidized fenugreek galactomannans as a versatile delivery system. Carbohydr. Polym. 2016, 144, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Bassi, P.; Kaur, G. Fenugreek gum derivatives with improved bioadhesion and controlled drug release: In vitro and in vivo characterization. J. Drug Deliv. Sci. Technol. 2015, 29, 42–54. [Google Scholar] [CrossRef]
- Braz, L.; Grenha, A.; Corvo, M.C.; Lourenço, J.P.; Ferreira, D.; Sarmento, B.; Costa, A.M.R. Synthesis and characterization of Locust Bean Gum derivatives and their application in the production of nanoparticles. Carbohydr. Polym. 2018, 181, 974–985. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Zhang, J.; Zhao, B.; Yao, J.; Wang, Y. Structure-antioxidant relationships of sulfated galactomannan from guar gum. Int. J. Biol. Macromol. 2010, 46, 59–66. [Google Scholar] [CrossRef]
- Muschin, T.; Budragchaa, D.; Kanamoto, T.; Nakashima, H.; Ichiyama, K.; Yamamoto, N.; Shuqin, H.; Yoshida, T. Chemically sulfated natural galactomannans with specific antiviral and anticoagulant activities. Int. J. Biol. Macromol. 2016, 89, 415–420. [Google Scholar] [CrossRef]
- Cunha de Padua, M.M.; Cadena, S.M.S.C.; Petkowicz, C.L.O.; Martinez, G.R.; Noleto, G.R. Galactomannan from Schizolobium amazonicum seed and its sulfated derivatives impair metabolism in HepG2 cells. Int. J. Biol. Macromol. 2017, 101, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, V.D.; Jani, G.K.; Moradiya, N.G.; Randeria, N.P.; Nagar, B.J.; Naikwadi, N.N.; Variya, B.C. Galactomannan: A versatile biodegradable seed polysaccharide. Int. J. Biol. Macromol. 2013, 60, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Chrestani, F.; Sierakowski, M.R.; Uchoa, D.E.A.; Nozawa, C.; Sassaki, G.L.; Gorin, P.A.J.; Ono, L. In vitro antiherpetic and antirotaviral activities of a sulfate prepared from Mimosa scabrella galactomannan. Int. J. Biol. Macromol. 2009, 45, 453–457. [Google Scholar] [CrossRef]
- Mestechkina, N.M.; Shcherbukhin, V.D.; Bannikova, G.E.; Varlamov, V.P.; Drozd, N.N.; Tolstenkov, A.S.; Makarov, V.A.; Tikhonov, V.E. Anticoagulant activity of low-molecular-weight sulfated derivatives of galactomannan from Cyamopsis tetragonoloba (L.) seeds. Appl. Biochem. Microbiol. 2008, 44, 111–116. [Google Scholar] [CrossRef]
- Hussein, M.M.D.; Helmy, W.A.; Salem, H.M. Biological activities of some galactomannans and their sulfated derivatives. Phytochemistry 1998, 48, 479–484. [Google Scholar] [CrossRef]
- Dos Santos, V.R.F.; Souza, B.W.S.; Teixeira, J.A.; Vicente, A.A.; Cerqueira, M.A. Relationship between galactomannan structure and physicochemical properties of films produced thereof. J. Food Sci. Technol. 2015, 52, 8292–8299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, R.R.; Magalhães, H.S.; Souza, J.R.R.; Trevisan, M.T.S.; Vieira, Í.G.P.; Feitosa, J.P.A.; Araújo, T.G.; Ricardo, N.M.P.S. Exploring the potential of Dimorphandra gardneriana galactomannans as drug delivery systems. Ind. Crop. Prod. 2015, 69, 284–289. [Google Scholar] [CrossRef]
- Ponzini, E.; Natalello, A.; Usai, F.; Bechmann, M.; Peri, F.; Müller, N.; Grandori, R. Structural characterization of aerogels derived from enzymatically oxidized galactomannans of fenugreek, sesbania and guar gums. Carbohydr. Polym. 2019, 207, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Ancona, D.; Pacheco-Aguirre, J.; Castellanos-Ruelas, A.; Chel-Guerrero, L. Microencapsulation of papain using carboxymethylated flamboyant (Delonix regia) seed gum. Innov. Food Sci. Emerg. Technol. 2011, 12, 67–72. [Google Scholar] [CrossRef]
- Bassi, P.; Kaur, G. Bioadhesive vaginal drug delivery of nystatin using a derivatized polymer: Development and characterization. Eur. J. Pharm. Biopharm. 2015, 96, 173–184. [Google Scholar] [CrossRef]
- Xie, J.-H.; Wang, Z.-J.; Shen, M.-Y.; Nie, S.-P.; Gong, B.; Li, H.-S.; Zhao, Q.; Li, W.-J.; Xie, M.-Y. Sulfated modification, characterization and antioxidant activities of polysaccharide from Cyclocarya paliurus. Food Hydrocoll. 2016, 53, 7–15. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, H.; Chen, T.; Zhang, H.; Liang, J.; Kong, W.; Yao, J.; Zhang, J.; Wang, J. Synthesis and structure characterization of sulfated galactomannan from fenugreek gum. Int. J. Biol. Macromol. 2019, 125, 1184–1191. [Google Scholar] [CrossRef]
- Barddal, H.P.O.; Faria, F.A.M.; Nogueira, A.V.; Iacomini, M.; Cipriani, T.R. Anticoagulant and antithrombotic effects of chemically sulfated guar gum. Int. J. Biol. Macromol. 2020, 45, 604–610. [Google Scholar] [CrossRef]
- Noleto, G.R.; Petkowicz, C.L.O.; Mercê, A.L.R.; Noseda, M.D.; Méndez-Sánchez, S.C.; Reicher, F.; Oliveira, M.B.M. Two galactomannan preparations from seeds from Mimosa scabrella (bracatinga): Complexation with oxovanadium (IV/V) and cytotoxicity on HeLa cells. J. Inorg. Biochem. 2009, 103, 749–757. [Google Scholar] [CrossRef]
- Wang, J.; Yang, T. Synthesis and characterization of phosphorylated galactomannan: The effect of DS on solution conformation and antioxidant activities. Carbohydr. Polym. 2014, 113, 325–335. [Google Scholar] [CrossRef]
- Lavazza, M.; Formantici, C.; Langella, V.; Monti, D.; Pfeiffer, U.; Galante, Y.M. Oxidation of galactomannan by laccase plus TEMPO yields an elastic gel. J. Biotechnol. 2011, 156, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Adriazola, I.O.; Amaral, A.E.; Amorim, J.C.; Correia, B.L.; Petkowicz, C.L.O.; Mercê, A.L.R.; Noleto, G.R. Macrophage activation and leishmanicidal activity by galactomannan and its oxovanadium (IV/V) complex in vitro. J. Inorg. Biochem. 2014, 132, 45–51. [Google Scholar] [CrossRef]
- Godoi, A.M.; Faccin-Galhardi, L.C.; Lopes, N.; Nozawa, C.; Almeida, R.R.; Ricardo, N.M.P.S.; Linhares, R.E.C. Characterization and antiherpetic activity of native and chemically sulfated polysaccharide from Adenanthera pavonina. Curr. Pharm. Biotechnol. 2015, 16, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Ba, J.; Gao, Y.; Xu, Q.; Qin, M. Research Development of Modification of Galactomannan Gums from Plant Resources. Adv. Mater. Res. 2012, 482–484, 1628–1631. [Google Scholar] [CrossRef]
- Castro, R.R.; Silva, C.M.M.; Nunes, R.M.; Cunha, P.L.R.; Paula, R.C.M.; Feitosa, J.P.A.; Girão, V.C.C.; Pompeu, M.M.L.; Leite, J.A.D.; Rocha, F.A.C. Structural characteristics are crucial to the benefits of guar gum in experimental osteoarthritis. Carbohydr. Polym. 2016, 150, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Ono, L.; Wollinger, W.; Rocco, I.M.; Coimbra, T.L.M.; Gorin, P.A.J.; Sierakowski, M.R. In vitro and in vivo antiviral properties of sulfated galactomannans against yellow fever virus (BeH111 strain) and dengue 1 virus (Hawaii strain). Antivir. Res. 2003, 60, 201–208. [Google Scholar] [CrossRef]
- Gemin, E.; Ferreira, C.E.O.; Sierakowski, M.R.; Jorge, T.R.; Joineau, M.E.G.; Ono, L. In vitro anti-HSV-1 activity of a chemically sulfated galactomannan from Leucaena leucocephala seeds. J. Basic Appl. Pharm. Sci. 2010, 31, 165–170. [Google Scholar]
- Lopes, N.; Faccin-Galhardi, L.C.; Espada, S.F.; Pacheco, A.C.; Ricardo, N.M.P.S.; Linhares, R.E.C.; Nozawa, C. Sulfated polysaccharide of Caesalpinia ferrea inhibits herpes simplex virus and poliovirus. Int. J. Biol. Macromol. 2013, 60, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.M.M.; Morais, S.M.; Silva, A.R.A.; Barroso, N.D.; Pontes Filho, T.R.; Araujo, F.M.D.C.; Vieira, I.G.P.; Lima, D.M.; Guedes, M.I.F. Antiviral and Antioxidant Activities of Sulfated Galactomannans from Plants of Caatinga Biome. Evid. Based Complement. Altern. Med. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Cumpstey, I. Chemical Modification of Polysaccharides. Int. Sch. Res. Not. Org. Chem. 2013, 2013, 1–27. [Google Scholar] [CrossRef]
- Chen, W.L.; Chen, H.L.; Guo, G.W.; Huang, Y.C.; Chen, C.Y.; Tsai, Y.; Huang, K.F.; Chao-Hsun Yang, C.H. Locust bean gum galactomannan hydrolyzed by thermostable β-D-mannanase may reduce the secretion of pro-inflammatory factors and the release of granule constituents. Int. J. Biol. Macromol. 2018, 114, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Noleto, G.R.; Mercê, A.L.R.; Iacomini, M.; Gorin, P.A.J.; Soccol, V.T.; Oliveira, M.B. Effects of a lichen galactomannan and its vanadyl (IV) complex on peritoneal macrophages and leishmanicidal activity. Mol. Cell. Biochem. 2002, 233, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.E.; Petkowicz, C.L.O.; Mercê, A.L.R.; Iacomini, M.; Martinez, G.R.; Rocha, M.E.M.; Cadena, S.M.S.C.; Noleto, G.R. Leishmanicidal activity of polysaccharides and their oxovanadium (IV/V) complexes. Eur. J. Med. Chem. 2015, 90, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Cunha-de Padua, M.M.; Cadena, S.M.S.C.; Petkowicz, C.L.O.; Martinez, G.R.; Rocha, M.E.M.; Mercê, A.L.R.; Noleto, G.R. Toxicity of native and oxovanadium (IV/V) galactomannan complexes on HepG2 cells is related to impairment of mitochondrial functions. Carbohydr. Polym. 2017, 173, 665–675. [Google Scholar] [CrossRef]
- Gamal-Eldeen, A.M.; Amer, H.; Helmy, W.A. Cancer chemopreventive and anti-inflammatory activities of chemically modified guar gum. Chem. Biol. Interact. 2006, 161, 229–240. [Google Scholar] [CrossRef]
- Andrews, P.R.; Craik, D.J.; Martin, J.L. Functional Group Contributions to Drug-Receptor Interactions. J. Med. Chem. 1984, 27, 1648–1657. [Google Scholar] [CrossRef]
- Mao, F.; Ni, W.; Xu, X.; Wang, H.; Wang, J.; Ji, M.; Li, J. Chemical Structure-Related Drug-Like Criteria of Global Approved Drugs. Molecules 2016, 21, 75. [Google Scholar] [CrossRef] [Green Version]
- Godoi, A.M.; Faccin-Galhardi, L.C.; Lopes, N.; Rechenchoski, D.Z.; Almeida, R.R.; Ricardo, N.M.P.S.; Nozawa, C.; Linhares, R.E.C. Antiviral activity of sulfated polysaccharide of Adenanthera pavonina against Poliovirus in HEp-2 Cells. Evid. Based Complement. Altern. Med. 2014, 2014, 712634. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Kato, A.; Kobayashit, K. Bifunctional Lysozyme-Galactomannan Conjugate Having Excellent Emulsifying Properties and Bactericidal Effect. J. Agric. Food Chem. 1992, 40, 735–739. [Google Scholar] [CrossRef]
- Nakamura, S.; Kato, A. Multi-functional biopolymer prepared by covalent attachment of galactomannan to egg-white proteins through naturally occurring Maillard reaction. Nahrung 2000, 44, 201–206. [Google Scholar] [CrossRef]
- Das, D.; Ara, T.; Dutta, S.; Mukherjee, A. New water-resistant biomaterial biocide film based on guar gum. Bioresour. Technol. 2011, 102, 5878–5883. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.; Gorin, P.A.J.; Reicher, F.; Sierakowski, M.R. An active heparinoid obtained by sulphation of a galactomannan extracted from the endosperm of Senna macranthera seeds. Carbohydr. Polym. 2001, 46, 165–169. [Google Scholar] [CrossRef]
- Naithani, R.; Huma, L.C.; Moriarty, R.M.; McCormick, D.L.; Mehta, R.G. Comprehensive review of cancer chemopreventive agents evaluated in experimental carcinogenesis models and clinical trials. Curr. Med. Chem. 2008, 15, 1044–1071. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Benny, I.S.; Gunasekar, V.; Ponnusami, V. Review on Application of Xanthan Gum in Drug Delivery. Int. J. PharmTech Res. 2014, 6, 1322–1326. [Google Scholar]
- Pleissner, D.; Kümmerer, K. Green Chemistry and Its Contribution to Industrial Biotechnology. Adv. Biochem. Eng. Biotechnol. 2020, 173, 281–298. [Google Scholar]
- Cavalu, S.; Kamel, E.; Laslo, V.; Fritea, L.; Costea, T.; Antoniac, I.V.; Vasile, E.; Antoniac, A.; Semenescu, A.; Mohan, A.; et al. Eco-friendly, Facile and Rapid Way for Synthesis of Selenium Nanoparticles. Production, structural and morphological characterisation. Rev. Chim. 2017, 68, 2963–2966. [Google Scholar] [CrossRef]
- Cavalu, S.; Antoniac, I.V.; Fritea, L.; Mates, L.M.; Milea, C.; Laslo, V.; Vicas, S.; Mohan, A. Surface modifications of the titanium mesh for cranioplasty using selenium nanoparticles coating. J. Adhes. Sci. Technol. 2018, 32, 2509–2522. [Google Scholar] [CrossRef]
- Islan, G.A.; Mukherjee, A.; Castro, G.R. Development of biopolymer nanocomposite for silver nanoparticles and Ciprofloxacin controlled release. Int. J. Biol. Macromol. 2015, 72, 740–750. [Google Scholar] [CrossRef]
- Teixeira, G.F.D.; Vieira-Neto, A.E.; da Costa, F.N.; Silva, A.R.A.; Campos, A.R. Antinociceptive effect of (-)-α-bisabolol in nanocapsules. Biomed. Pharmacother. 2017, 91, 946–950. [Google Scholar] [CrossRef]
- Srivastav, A.K.; Dhiman, N.; Tiwari, R.; Arjaria, N.; Prakash, J.; Jagdale, P.; Ayanur, A.; Singh, D.; Satyakam Patnaik, S.; Kumar, M. Sub-acute oral exposure of zinc oxide nanoparticles causes alteration in iron homeostasis through acute phase response: A protective effect by surface modification. J. Trace Elem. Med. Biol. 2018, 270–287. [Google Scholar] [CrossRef] [PubMed]
- Pu, C.; Tang, W.; Li, X.; Li, M.; Sun, Q. Stability enhancement efficiency of surface decoration on curcumin-loaded liposomes: Comparison of guar gum and its cationic counterpart. Food Hydrocoll. 2019, 87, 29–37. [Google Scholar] [CrossRef]
- Bosio, V.E.; Basu, S.; Abdullha, F.; Villalba, M.E.C.; Güida, J.A.; Mukherjee, A.; Castro, G.R. Encapsulation of Congo Red in carboxymethyl guar gum-alginate gel microspheres. React. Funct. Polym. 2014, 82, 103–110. [Google Scholar] [CrossRef]
- Verma, S.; Ahuja, M. Carboxymethyl sesbania gum: Synthesis, characterization and evaluation for drug delivery. Int. J. Biol. Macromol. 2017, 98, 75–83. [Google Scholar] [CrossRef]
- Seeli, D.S.; Prabaharan, M. Guar gum oleate-graft-poly(methacrylic acid) hydrogel as a colon-specific controlled drug delivery carrier. Carbohydr. Polym. 2017, 158, 51–57. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Galactomannan Sources and/or Polysaccharides | Processes for Obtaining Derivatives | Chemical Analysis | Biological Activity/Properties | References |
|---|---|---|---|---|
| Mimosa scabrella seeds | Sulfation | SEC-MALS | Antiviral | [26] |
| Leucaena leucocephala seeds | Sulfation | SEC-MALS | Antiviral | [26] |
| Mimosa scabrella seeds | Sulfation | FTIR, 13C NMR | Antiviral | [8] |
| Leucaena leucocephala seeds | Sulfation | FTIR, GLC | Antiviral | [27] |
| Caesalpinia ferrea seeds | Sulfation | FTIR, 1H, 13C NMR | Antiviral | [28] |
| Adenanthera pavonina seeds | Sulfation | FTIR, GLC, IV | Antiviral | [23,38] |
| Adenanthera pavonina L. seeds | Sulfation | FTIR, UV-VIS | Antiviral | [29] |
| Caesalpinia ferrea Mart. seeds | Sulfation | FTIR, UV-VIS | Antiviral | [29] |
| Dimorphandra gardneriana seeds | Sulfation | FTIR, UV-VIS | Antiviral | [29] |
| Trigonella foenum-graecum seeds, Fenugreek gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Antiviral | [5] |
| Cyamopsis tetragonolobus seeds, Guar gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Antiviral | [5] |
| Caesalpinia spinosa, Tara gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Antiviral | [5] |
| Ceratonia siliqua L. seeds, Locust bean gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Antiviral | [5] |
| Cyamopsis tetragonolobus seeds, Guar gum | Maillard conjugation | UV-VIS, SDS PAGE | Antimicrobial | [39,40] |
| Cyamopsis tetragonolobus seeds, Guar gum | Benzoylation | FTIR, 13C NMR, XRD, TGA, C, H, N analysis. | Antimicrobial | [41] |
| Leucaena sp. seeds | Sulfation | PC, UV-VIS | Anticoagulant and fibrinolytic | [10] |
| Medicago sativa seeds | Sulfation | PC, UV-VIS | Anticoagulant and fibrinolytic | [10] |
| Glycine max seed hulls | Sulfation | PC, UV-VIS | Anticoagulant and fibrinolytic | [10] |
| Phoenix dactylifera seeds | Sulfation | PC, UV-VIS | Anticoagulant and fibrinolytic | [10] |
| Senna macranthera seeds | Sulfation | FTIR, 13C NMR, UV-VIS, GPC | Anticoagulant | [42] |
| Cyamopsis tetragonoloba seeds, Guar gum | Sulfation | GPC, IV, UV-VIS | Anticoagulant | [9] |
| Trigonella foenum-graecum seeds, Fenugreek gum | Sulfation | FTIR, 1H, 13C NMR, SPR | Anticoagulant | [5] |
| Cyamopsis tetragonolobus seeds, Guar gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Anticoagulant | [5] |
| Caesalpinia spinosa, Tara gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Anticoagulant | [5] |
| Ceratonia siliqua L. seeds, Locust bean gum | Sulfation | FTIR, 1H, 13C NMR, OR, GPC, SPR | Anticoagulant | [5] |
| Cyamopsis tetragonolobus seeds, Guar gum | C-glycosylation and sulfation | UV-VIS, PC | Chemopreventive | [35] |
| Cyamopsis tetragonoloba seeds, Guar gum | Sulfation | FTIR, 13C NMR, SEC-MALS | Antioxidant | [4] |
| Adenanthera pavonina L. seeds | Sulfation | FTIR, UV-VIS | Antioxidant | [29] |
| Caesalpinia ferrea Mart. seeds | Sulfation | FTIR, UV-VIS | Antioxidant | [29] |
| Dimorphandra gardneriana seeds | Sulfation | FTIR, UV-VIS | Antioxidant | [29] |
| Cyamopsis tetragonoloba seeds, Guar gum | Phosphorylation | FTIR, 13C NMR, XPS, GC–MS, SEC-MALS | Antioxidant | [20] |
| Schizolobium amazonicum seeds | Sulfation | FTIR, HPSEC, 13C NMR, GLC, UV-VIS | Anticancer | [6] |
| Cyamopsis tetragonolobus seeds, Guar gum | C-glycosylation and sulfation | UV-VIS, PC | Anticancer | [35] |
| Cyamopsis tetragonolobus seeds, Guar gum | C-glycosylation | UV-VIS, PC | Anticancer | [35] |
| Schizolobium amazonicum seeds | Partial hydrolysis | FTIR, HPSEC, GLC, 13C NMR, 51V NMR, PT | Anticancer | [34] |
| Schizolobium amazonicum seeds | Partial hydrolysis and complexation with oxovanadium | FTIR, HPSEC, GLC, 13C NMR, 51V NMR, PT | Anticancer | [34] |
| Mimosa scabrella seeds | Complexation with oxovanadium | FTIR, HPSEC, GLC, 51V NMR, PT | Anticancer | [19] |
| Cyamopsis tetragonoloba seeds, Protein-free guar gum | Sulfation | FTIR, 1H, 13C NMR, IV, PT, GPC | Analgesia and chondroprotection | [25] |
| Cyamopsis tetragonoloba seeds, Protein-free guar gum | Oxidation | FTIR, 1H, 13C NMR, IV, PT, GPC | Analgesia and chondroprotection | [25] |
| Ceratonia siliqua L. seeds, Locust bean gum | Hydrolysis | HPSEC, HPLC | Immunomodulatory | [38] |
| Mimosa scabrella seeds | Complexation with oxovanadium | FTIR, PT, 51V NMR, GC-MS | Immunomodulatory | [22] |
| Mimosa scabrella seeds | Complexation with oxovanadium | FTIR, PT, 51V NMR, GC-MS | Antileishmanial | [22] |
| Ramalina celastri | Complexation with vanadium | FTIR, PT | Immunomodulatory | [39] |
| Ramalina celastri | Complexation with vanadium | FTIR, PT | Antileishmanial | [39] |
| Ramalina celastri | Complexation with oxovanadium | PT, 51V NMR | Immunomodulatory | [33] |
| Ramalina celastri | Complexation with oxovanadium | PT, 51V NMR | Antileishmanial | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geronço, M.S.; da Silveira Ramos, I.F.; da Silva Filho, E.C.; dos Santos Rizzo, M.; Ribeiro, A.B.; da Costa, M.P. Are Structurally Modified Galactomannan Derivatives Biologically Active? Polysaccharides 2021, 2, 1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2010001
Geronço MS, da Silveira Ramos IF, da Silva Filho EC, dos Santos Rizzo M, Ribeiro AB, da Costa MP. Are Structurally Modified Galactomannan Derivatives Biologically Active? Polysaccharides. 2021; 2(1):1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2010001
Chicago/Turabian StyleGeronço, Maurycyo Silva, Igor Frederico da Silveira Ramos, Edson Cavalcanti da Silva Filho, Márcia dos Santos Rizzo, Alessandra Braga Ribeiro, and Marcilia Pinheiro da Costa. 2021. "Are Structurally Modified Galactomannan Derivatives Biologically Active?" Polysaccharides 2, no. 1: 1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2010001