
Physicochemical Properties of Chitosan from Two Commonly Reared Edible Cricket Species, and Its Application as a Hypolipidemic and Antimicrobial Agent



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cricket Chitin Extraction
2.2. Chitin Demineralization and Deproteinization
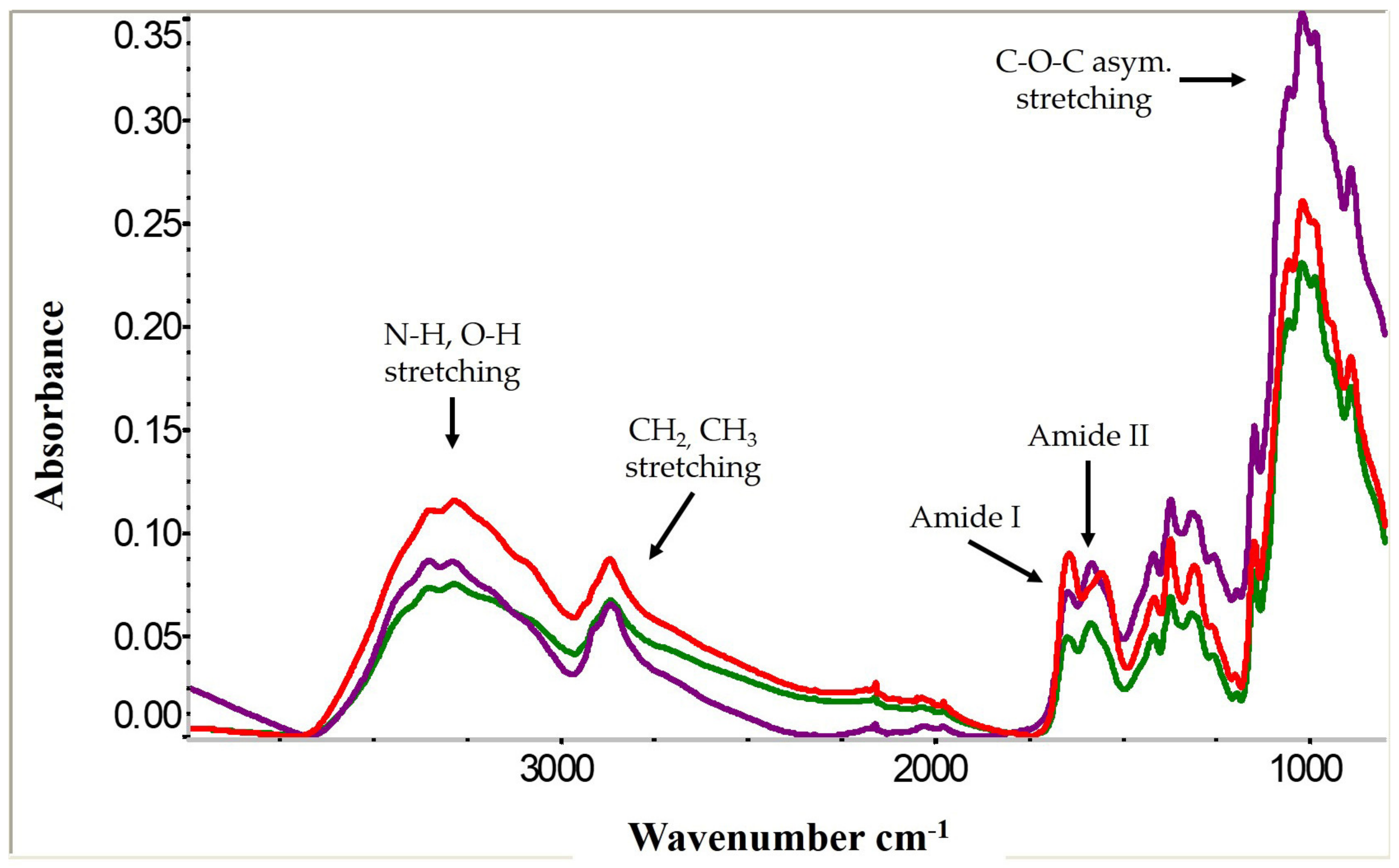
2.3. Chitosan Conversion and Characterization by Fourier Transform Infrared Spectroscopy (FTIR) and Degree of Deacetylation
2.4. Molecular Weight Determination
2.5. Anti-Obesity Effects (Lipid-Binding Capacity) of Cricket Chitosan
2.6. Determination of Antimicrobial Activity
2.6.1. Bacterial Stock Cultures and Sample Preparation
2.6.2. Antibacterial Determination
2.7. Statistical Analysis
3. Results and Discussion
3.1. Cricket Chitin and Chitosan Processing
3.2. Cricket Chitin and Chitosan Characterization
3.3. Molecular Weight Determination
3.4. Lipid-Binding Capacity
3.5. Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sivashankari, P.R.; Prabaharan, M. 5-Deacetylation modification techniques of chitin and chitosan. In Chitosan Based Biomaterials; Jennings, J.A., Bumgardner, J.D., Eds.; Woodhead Publishing: Cambridge, UK, 2017; Volume 1, pp. 117–133. [Google Scholar]
- Kumar, M.N.V.R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Zargar, V.; Asghari, M.; Dashti, A. A Review on Chitin and Chitosan Polymers: Structure, Chemistry, Solubility, Derivatives, and Applications. ChemBioEng Rev. 2015, 2, 204–226. [Google Scholar] [CrossRef]
- No, H.K.; Meyers, S.P. Preparation and Characterization of Chitin and Chitosan—A Review. J. Aquat. Food Prod. Technol. 1995, 4, 27–52. [Google Scholar] [CrossRef]
- Majtán, J.; Bíliková, K.; Markovič, O.; Gróf, J.; Kogan, G.; Šimúth, J. Isolation and characterization of chitin from bumblebee (Bombus terrestris). Int. J. Biol. Macromol. 2007, 40, 237–241. [Google Scholar] [CrossRef]
- Kaya, M.; Lelešius, E.; Nagrockaitė, R.; Sargin, I.; Arslan, G.; Mol, A.; Baran, T.; Can, E.; Bitim, B. Differentiations of Chitin Content and Surface Morphologies of Chitins Extracted from Male and Female Grasshopper Species. PLoS ONE 2015, 10, e0115531. [Google Scholar] [CrossRef]
- Kaya, M.; Sofi, K.; Sargin, I.; Mujtaba, M. Changes in physicochemical properties of chitin at developmental stages (larvae, pupa and adult) of Vespa crabro (wasp). Carbohydr. Polym. 2016, 145, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Zelencova, L.; Erdoǧan, S.; Baran, T.; Kaya, M.; Erdogan, S. Chitin extraction and chitosan production from Chilopoda (Scolopendra cingulata) with identification of physicochemical properties. Polym. Sci. Ser. A 2015, 57, 437–444. [Google Scholar] [CrossRef]
- Greven, H.; Kaya, M.; Sargin, I.; Baran, T.; Kristensen, R.M.; Sørensen, M.V. Characterisation of chitin in the cuticle of a velvet worm (Onychophora). Turk. J. Zooöl. 2019, 43, 416–424. [Google Scholar] [CrossRef]
- Kaya, M.; Sargin, I.; Sabeckis, I.; Noreikaite, D.; Erdonmez, D.; Salaberria, A.M.; Labidi, J.; Baublys, V.; Tubelytė, V. Biological, mechanical, optical and physicochemical properties of natural chitin films obtained from the dorsal pronotum and the wing of cockroach. Carbohydr. Polym. 2017, 163, 162–169. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Erdoğan, S.; Menteş, A.; Özüsağlam, M.A.; Çakmak, Y.S. Physicochemical comparison of chitin and chitosan obtained from larvae and adult Colorado potato beetle (Leptinotarsa decemlineata). Mater. Sci. Eng. C 2014, 45, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.-S.; Shin, C.-S.; Shin, W.-S. Characteristics of cricket (Gryllus bimaculatus) chitosan and chitosan-based nanoparticles. Food Sci. Biotechnol. 2018, 27, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Ibitoye, B.E.; Lokman, I.H.; Hezmee, M.N.M.; Goh, Y.M.; Zuki, A.B.Z.; Jimoh, A.A. Extraction and physicochemical characterization of chitin and chitosan isolated from house cricket. Biomed. Mater. 2017, 13, 025009. [Google Scholar] [CrossRef] [Green Version]
- Liceaga, A.M. Approaches for Utilizing Insect Protein for Human Consumption: Effect of Enzymatic Hydrolysis on Protein Quality and Functionality. Ann. Entomol. Soc. Am. 2019, 112, 529–532. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.; Liceaga, A. Effect of microwave-assisted enzymatic hydrolysis of cricket (Gryllodes sigillatus) protein on ACE and DPP-IV inhibition and tropomyosin-IgG binding. J. Funct. Foods 2020, 64, 103634. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Berezina, N.; Hubert, A.; Berro, F.; Levon, J.-G.; Le Roux, K.; Socolsky, C. Chitin, Hydrolysate and Production of at Least One Desired Product from Insects by Means of Enzymatic Hydrolysis, Comprising a Combination of Steps Performed Prior to the Enzymatic Hydrolysis. U.S. Patent Application No. 15/541,162, 2019. [Google Scholar]
- Berezina, N.; Hubert, A.; Berro, F.; Levon, J.-G.; Le Roux, K.; Socolsky, C. Chitin, Hydrolysate and Method for the Production of One or More Desired Products by Means of Enzymatic Hydrolysis, Including Pre-Treatment with an Oxidising Agent. U.S. Patent Application No. 15/541,174, 2018. [Google Scholar]
- Berezina, N.; Hubert, A.; Berro, F.; Levon, J.-G.; Le Roux, K.; Socolsky, C. Chitin, Hydrolysate and Method for the Production of One or More Desired Products from Insects by Means of Enzymatic Hydrolysis. U.S. Patent Application No. 15/541,186, 2018. [Google Scholar]
- Kim, M.-W.; Song, Y.-S.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Park, Y.-K.; Kang, S.H.; Kim, S.-A.; Choi, C.; Jung, W.-J. Production of chitin and chitosan from the exoskeleton of adult two-spotted field crickets (Gryllus bimaculatus). Entomol. Res. 2017, 47, 279–285. [Google Scholar] [CrossRef]
- Jin, Q.; Yu, H.; Wang, X.; Li, K.; Li, P. Effect of the molecular weight of water-soluble chitosan on its fat-/cholesterol-binding capacities and inhibitory activities to pancreatic lipase. PeerJ 2017, 5, e3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.; Liu, P.; Zhang, J.; Chen, J. Biological activities of chitosan and chitooligosaccharides. Food Hydrocoll. 2011, 25, 170–179. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.-T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as Antimicrobial Agent: Applications and Mode of Action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef]
- Luna, G.C.; Martin-Gonzalez, F.S.; Mauer, L.; Liceaga, A. Cricket (Acheta domesticus) protein hydrolysates’ impact on the physicochemical, structural and sensory properties of tortillas and tortilla chips. J. Insects Food Feed. 2021, 7, 109–120. [Google Scholar] [CrossRef]
- Percot, A.; Viton, A.C.; Domard, A. Optimization of Chitin Extraction from Shrimp Shells. Biomacromolecules 2003, 4, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Elshaarawy, R.F.M.; Mustafa, F.H.A.; Herbst, A.; Farag, A.E.M.; Janiak, C. Surface functionalization of chitosan isolated from shrimp shells, using salicylaldehyde ionic liquids in exploration for novel economic and ecofriendly antibiofoulants. RSC Adv. 2016, 6, 20901–20915. [Google Scholar] [CrossRef]
- Sajomsang, W.; Gonil, P. Preparation and characterization of α-chitin from cicada sloughs. Mater. Sci. Eng. C 2010, 30, 357–363. [Google Scholar] [CrossRef]
- Czechowska-Biskup, R.; Jarosińska, D.; Rokita, B.; Ulański, P.; Rosiak, J.M. Determination of degree of deacetylation of chitosan-comparision of methods. Prog. Chem. Appl. Chitin Deriv. 2012, 17, 5–20. [Google Scholar]
- Domard, A.; Rinaudo, M. Preparation and characterization of fully deacetylated chitosan. Int. J. Biol. Macromol. 1983, 5, 49–52. [Google Scholar] [CrossRef]
- Czechowska-Biskup, R.; Wach, R.A.; Rosiak, J.M.; Ulański, P. Procedure for determination of the molecular weight of chitosan by viscometry. Prog. Chem. Appl. Chitin its Deriv. 2018, XXIII, 45–54. [Google Scholar] [CrossRef]
- Roberts, G.A.; Domszy, J.G. Determination of the viscometric constants for chitosan. Int. J. Biol. Macromol. 1982, 4, 374–377. [Google Scholar] [CrossRef]
- Chen, R.H.; Tsaih, M.L. Effect of temperature on the intrinsic viscosity and conformation of chitosans in dilute HCl solution. Int. J. Biol. Macromol. 1998, 23, 135–141. [Google Scholar] [CrossRef]
- Panith, N.; Wichaphon, J.; Lertsiri, S.; Niamsiri, N. Effect of physical and physicochemical characteristics of chitosan on fat-binding capacities under in vitro gastrointestinal conditions. LWT 2016, 71, 25–32. [Google Scholar] [CrossRef]
- Zhou, K.; Xia, W.; Zhang, C.; Yu, L. (Lucy) In vitro binding of bile acids and triglycerides by selected chitosan preparations and their physico-chemical properties. LWT 2006, 39, 1087–1092. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Deering, A.J.; Liceaga, A.M. New Insights into the Antimicrobial Properties of Hydrolysates and Peptide Fractions Derived from Chia Seed (Salvia hispanica L.). Probiotics Antimicrob. Proteins 2020, 12, 1571–1581. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Mentes, A.; Sezen, G. Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan Orthoptera species (Insecta). Biotechnol. Bioprocess Eng. 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Hu, M.; Gurtler, J.B. Selection of Surrogate Bacteria for Use in Food Safety Challenge Studies: A Review. J. Food Prot. 2017, 80, 1506–1536. [Google Scholar] [CrossRef]
- Kurita, K. Chitin and Chitosan: Functional Biopolymers from Marine Crustaceans. Mar. Biotechnol. 2006, 8, 203–226. [Google Scholar] [CrossRef] [PubMed]
- Berezina, N.; Hubert, A. Marketing and Regulations of Chitin and Chitosan from Insects. Chitin Chitosan 2019, 2019, 477–489. [Google Scholar] [CrossRef]
- Zhang, M.; Haga, A.; Sekiguchi, H.; Hirano, S. Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 2000, 27, 99–105. [Google Scholar] [CrossRef]
- Purkayastha, D.; Sarkar, S. Physicochemical Structure Analysis of Chitin Extracted from Pupa Exuviae and Dead Imago of Wild Black Soldier Fly (Hermetia illucens). J. Polym. Environ. 2019, 28, 445–457. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, Y.; Han, Q.; Ji, L.; Zhang, H.; Fei, Z.; Wang, Y. Comparison of the physicochemical, rheological, and morphologic properties of chitosan from four insects. Carbohydr. Polym. 2019, 209, 266–275. [Google Scholar] [CrossRef]
- Mohan, K.; Ganesan, A.R.; Muralisankar, T.; Jayakumar, R.; Sathishkumar, P.; Uthayakumar, V.; Chandirasekar, R.; Revathi, N. Recent insights into the extraction, characterization, and bioactivities of chitin and chitosan from insects. Trends Food Sci. Technol. 2020, 105, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Marei, N.H.; El-Samie, E.A.; Salah, T.; Saad, G.R.; Elwahy, A.H. Isolation and characterization of chitosan from different local insects in Egypt. Int. J. Biol. Macromol. 2016, 82, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baran, T. Description of a new surface morphology for chitin extracted from wings of cockroach (Periplaneta americana). Int. J. Biol. Macromol. 2015, 75, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Kumirska, J.; Czerwicka, M.; Kaczyński, Z.; Bychowska, A.; Brzozowski, K.; Thöming, J.; Stepnowski, P. Application of Spectroscopic Methods for Structural Analysis of Chitin and Chitosan. Mar. Drugs 2010, 8, 1567–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Jiang, C.; Yang, Q. Extraction and Characterization of Chitin from the Beetle Holotrichia parallela Motschulsky. Molecules 2012, 17, 4604–4611. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.-K.; Kong, B.-G.; Jeong, Y.-I.; Lee, C.H.; Nah, J.-W. Physicochemical characterization of α-chitin, β-chitin, and γ-chitin separated from natural resources. J. Polym. Sci. Part A Polym. Chem. 2004, 42, 3423–3432. [Google Scholar] [CrossRef]
- Ai, H.; Wang, F.; Yang, Q.; Zhu, F.; Lei, C. Preparation and biological activities of chitosan from the larvae of housefly, Musca domestica. Carbohydr. Polym. 2008, 72, 419–423. [Google Scholar] [CrossRef]
- Song, C.; Yu, H.; Zhang, M.; Yang, Y.; Zhang, G. Physicochemical properties and antioxidant activity of chitosan from the blowfly Chrysomya megacephala larvae. Int. J. Biol. Macromol. 2013, 60, 347–354. [Google Scholar] [CrossRef]
- Khayrova, A.; Lopatin, S.; Varlamov, V. Black Soldier Fly Hermetia illucens as a Novel Source of Chitin and Chitosan. Int. J. Sci. 2019, 8, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Raafat, D.; Sahl, H.G. Chitosan and its antimicrobial potential—A critical literature survey. Microb. Biotechnol. 2009, 2, 186–201. [Google Scholar] [CrossRef] [Green Version]
- Varun, T.K.; Senani, S.; Jayapal, N.; Chikkerur, J.; Roy, S.; Tekulapally, V.B.; Gautam, M.; Kumar, N. Extraction of chitosan and its oligomers from shrimp shell waste, their characterization and antimicrobial effect. Veter. World 2017, 10, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Czechowska-Biskup, R.; Rokita, B.; Ulanski, P.; Rosiak, J. Radiation-induced and sonochemical degradation of chitosan as a way to increase its fat-binding capacity. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2005, 236, 383–390. [Google Scholar] [CrossRef]
- Dimzon, I.K.D.; Ebert, J.; Knepper, T.P. The interaction of chitosan and olive oil: Effects of degree of deacetylation and degree of polymerization. Carbohydr. Polym. 2013, 92, 564–570. [Google Scholar] [CrossRef]
- Kaya, M.; Sargin, I.; Mulerčikas, P.; Labidi, J.; Salaberria, A.M.; Cakmak, Y.S.; Kazlauskaitė, S.; Erdonmez, D.; Baublys, V. Conversion of Waste Parasitic Insect (Hylobius abietis L.) into Antioxidative, Antimicrobial and Biodegradable Films. J. Renew. Mater. 2019, 7, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Fraise, A.; Wilkinson, M.; Bradley, C.; Oppenheim, B.; Moiemen, N. The antibacterial activity and stability of acetic acid. J. Hosp. Infect. 2013, 84, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Halstead, F.D.; Rauf, M.; Moiemen, N.S.; Bamford, A.; Wearn, C.M.; Fraise, A.P.; Lund, P.A.; Oppenheim, B.A.; Webber, M.A. The Antibacterial Activity of Acetic Acid against Biofilm-Producing Pathogens of Relevance to Burns Patients. PLoS ONE 2015, 10, e0136190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibañez-Peinado, D.; Ubeda-Manzanaro, M.; Martínez, A.; Rodrigo, D. Antimicrobial effect of insect chitosan on Salmonella Typhimurium, Escherichia coli O157:H7 and Listeria monocytogenes survival. PLoS ONE 2020, 15, e0244153. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Composition Analysis (%) | A. domesticus | G. sigillatus |
|---|---|---|
| Whole crickets, ground | ||
| Protein | 67.4 ± 1.5 1 | 56.8 ± 0.01 2 |
| Ash | 4.0 ± 0.90 1 | 18.1 ± 0.60 2 |
| Chitin 3 | 5.7 ± 0.10 | 3.4 ± 0.10 |
| Chitin, extracted and purified | ||
| Nitrogen | <6.9 | 7.1 |
| Protein (calculated) 4 | ND | 1.4 |
| Ash | 0.09 | <0.01 |
| Conversion Time (Hours) | A. domesticus3 | G. sigillatus3 | |
|---|---|---|---|
| DDA (%) 1 | |||
| 2 | 72.5 ± 1.0 a | 73.5 ± 1.4 a | |
| 4 | 76.3 ± 1.3 b | 74.9 ± 1.3 a | |
| 6 | 79.1 ± 1.9 c | 77.2 ± 1.8 b | |
| 10 | 79.4 ± 1.3 c | 81.3 ± 1.1 c | |
| Chitosan yield (%) 2 | |||
| 2 | 76.0 ± 6.7 ab | 65.0 ±1.6 a | |
| 4 | 77.3 ± 1.9 ab | 63.7 ± 1.2 a | |
| 6 | 80.5 ± 2.1 b | 60.3 ± 3.3 a | |
| 10 | 69.0 ± 2.2 a | 62.3 ± 0.9 a | |
| Chitosan molecular weight 4 | - | 344 kDa | 524 kDa |
| Lipid-Binding Capacity (g Oil per g Chitosan) 1 | |||
|---|---|---|---|
| DDA (%) | A. domesticus | G. sigillatus | Shrimp |
| ~72 | 210.8 ± 21.1 | 163.5 ± 17.8 | 168.5 ± 36.8 |
| ~76 | 221.8 ± 25.4 | 159.0 ± 15.9 | - |
| ~80 | 168.7 ± 10.2 | 180.5 ± 21.6 | - |
| Bacteria Growth Inhibition (%) 1 | ||||
|---|---|---|---|---|
| Inhibitors | Degree of Deacetylation (%) | |||
| 72 | 76 | 80 | ||
| Escherichia coli ATCC 25922 | ||||
| A. domesticus chitosan | - | 26.6 ± 0.7 a | 65.6 ± 2.5 b | 89 ± 1.2 c |
| G. sigillatus chitosan | - | 33.8 ± 1 a | 64.2 ± 2.7 b | 94.2 ± 0.9 c |
| Commercial shrimp chitosan | 21.4 ± 2.4 | - | - | - |
| Acetic acid 3 | 5.4 ± 0.2 | - | - | - |
| Listeria innocua ATCC 33090 | ||||
| A. domesticus chitosan | - | 100 2 | 100 | 100 |
| G. sigillatus chitosan | - | 100 | 100 | 100 |
| Commercial shrimp chitosan | 100 | - | - | - |
| Acetic acid 3 | 37.7 ± 0.7 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malm, M.; Liceaga, A.M. Physicochemical Properties of Chitosan from Two Commonly Reared Edible Cricket Species, and Its Application as a Hypolipidemic and Antimicrobial Agent. Polysaccharides 2021, 2, 339-353. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020022
Malm M, Liceaga AM. Physicochemical Properties of Chitosan from Two Commonly Reared Edible Cricket Species, and Its Application as a Hypolipidemic and Antimicrobial Agent. Polysaccharides. 2021; 2(2):339-353. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020022
Chicago/Turabian StyleMalm, Morgan, and Andrea M. Liceaga. 2021. "Physicochemical Properties of Chitosan from Two Commonly Reared Edible Cricket Species, and Its Application as a Hypolipidemic and Antimicrobial Agent" Polysaccharides 2, no. 2: 339-353. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020022