
Dextran: Sources, Structures, and Properties


Unidad Professional Interdisciplinaria de Biotecnología, Instituto Politécnico Nacional, Av. Acueducto s/n, Barrio La Laguna Ticoman, Ciudad de México 07340, Mexico
Polysaccharides 2021, 2(3), 554-565; https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030033
Submission received: 21 May 2021
/
Revised: 14 June 2021
/
Accepted: 28 June 2021
/
Published: 1 July 2021
(This article belongs to the Collection Current Opinion in Polysaccharides)
Abstract
:Dextran is an exopolysaccharide (EPS) synthesized by lactic acid bacteria (LAB) or their enzymes in the presence of sucrose. Dextran is composed of a linear chain of d-glucoses linked by α-(1→6) bonds, with possible branches of d-glucoses linked by α-(1→4), α-(1→3), or α-(1→2) bonds, which can be low (<40 kDa) or high molecular weight (>40 kDa). The characteristics of dextran in terms of molecular weight and branches depend on the producing strain, so there is a great variety in its properties. Dextran has commercial interest because its solubility, viscosity, and thermal and rheological properties allow it to be used in food, pharmaceutical, and research areas. The aim of this review article is to compile the latest research (in the past decade) using LAB to synthesize high or low molecular weight dextran. In addition, studies using modified enzymes to produce dextran with specific structural characteristics (molecular weights and branches) are addressed. On the other hand, special attention is paid to LAB extracted from unconventional sources to expose their capacities as dextran producers and their possible application to compete with the only commercial strain (Leuconostoc mesenteroides NRRL B512).
1. Introduction
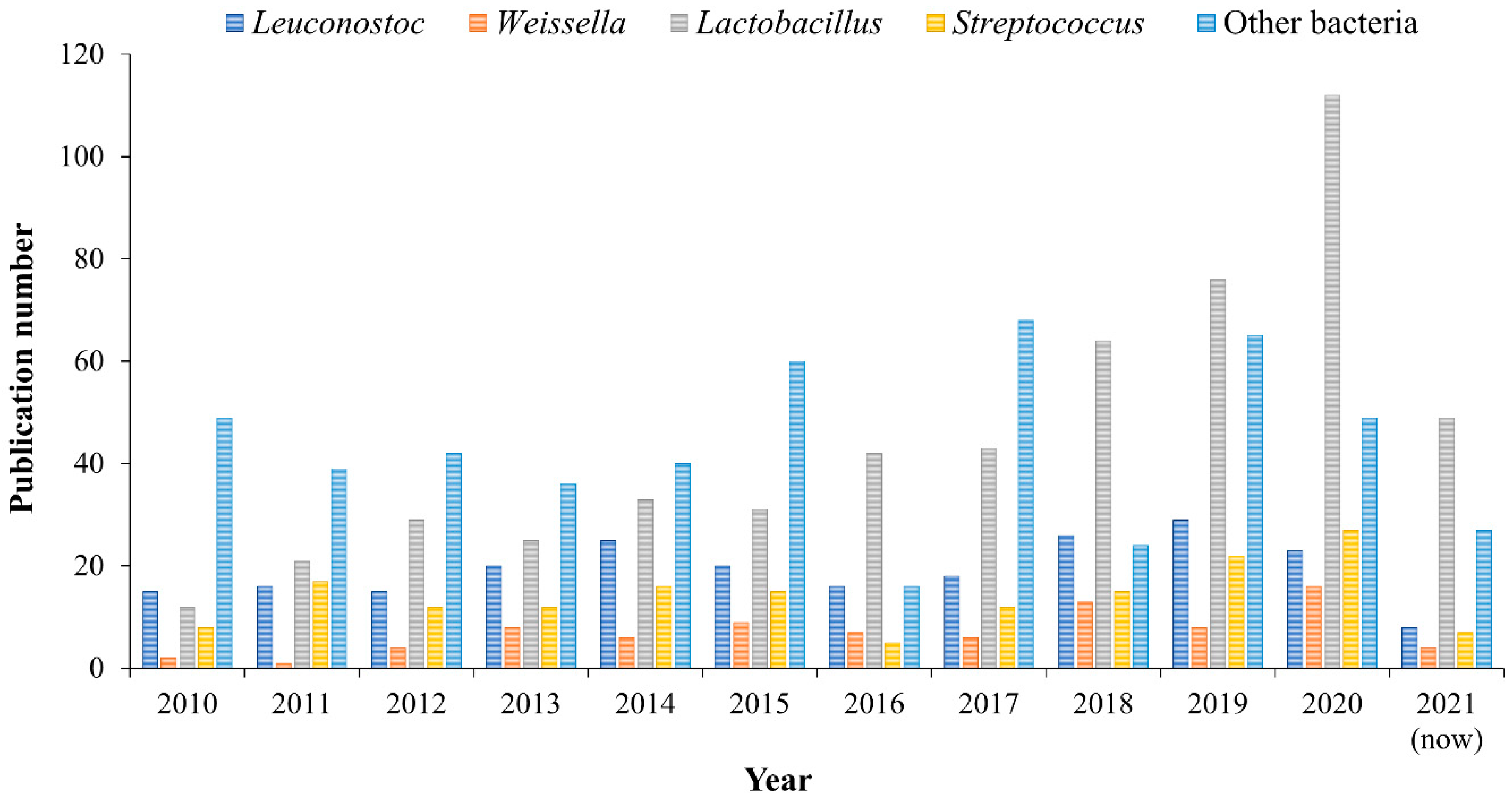
Lactic acid bacteria (LAB) are microorganisms that produce lactic acid as the main or only product of carbohydrate fermentation (heterofermentation or homofermentation, respectively). The nutritional requirements are complex, since they are based on vitamins, minerals, fatty acids, amino acids, peptides, and carbohydrates, which are usually in their natural habitats [1]. LAB have been isolated from dairy foods, meats, cereals, vegetables, soil, water, and vaginal waste. According to their characteristics and taxonomy, LAB include bacteria belonging to the genera Aerococcus, Alloiococcus, Carnobacterium, Dolosigranulum, Enterococcus, Globicatella, Lactococcus, Lactobacillus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Vaiscoccus, and Weiscoccus [2]. LAB are considered probiotic bacteria because they can be incorporated into food to improve the consumer′s intestinal microbial balance, and they are also generally recognized as safe (GRAS) because they are not pathogenic for humans [3]. On the other hand, they are responsible for a great diversification of flavors and textures of food products, which is why they are mainly used to produce different fermented products such as yogurt, cheese, sourdoughs, pickles, sausages, and soy products [4]. In addition, some LAB can produce extracellular polysaccharides (called exopolysaccharides, EPS) that are repeat units of sugars such as glucose, galactose, and rhamnose, which are secreted during bacterial growth [5]. EPS can be classified into two groups depending on the units that comprise it. Heteropolysaccharides consist of different monosaccharide units, for example, xanthan and gellan. Homopolysaccharides are composed of repeating units of a single type of monosaccharides (e.g., glucose or fructose), for example, glucans and fructans. Levan and inulin are the fructans produced by LAB, while the most commonly produced glucans are cellulose, pullulan, curdlan, mutan, alternan, and dextran [6]. These natural polysaccharides have been used as carriers, encapsulants, thickeners, binders, lubricants, and additives in the pharmaceutical and food industries [7]. However, the most important EPS for medical and industrial use is dextran, which was initially believed to be synthesized only by Leuconostoc mesenteroides, but subsequent research reported its segregation by another type of LAB (see Section 2) [8]. The literature on the identification or characterization of dextrans produced by LAB has been increasing in the past decade, as can be seen in Figure 1. Therefore, the aim of this review is to show the advances that have been made in the discovery and characterization of new dextrans, their structural characteristics (molecular weight, links, and branches), and a brief description of their possible applications in medical, food, and research areas. In addition, emphasis is placed on extraction sources for dextran-producing bacterial strains.
2. Synthesis of Dextran
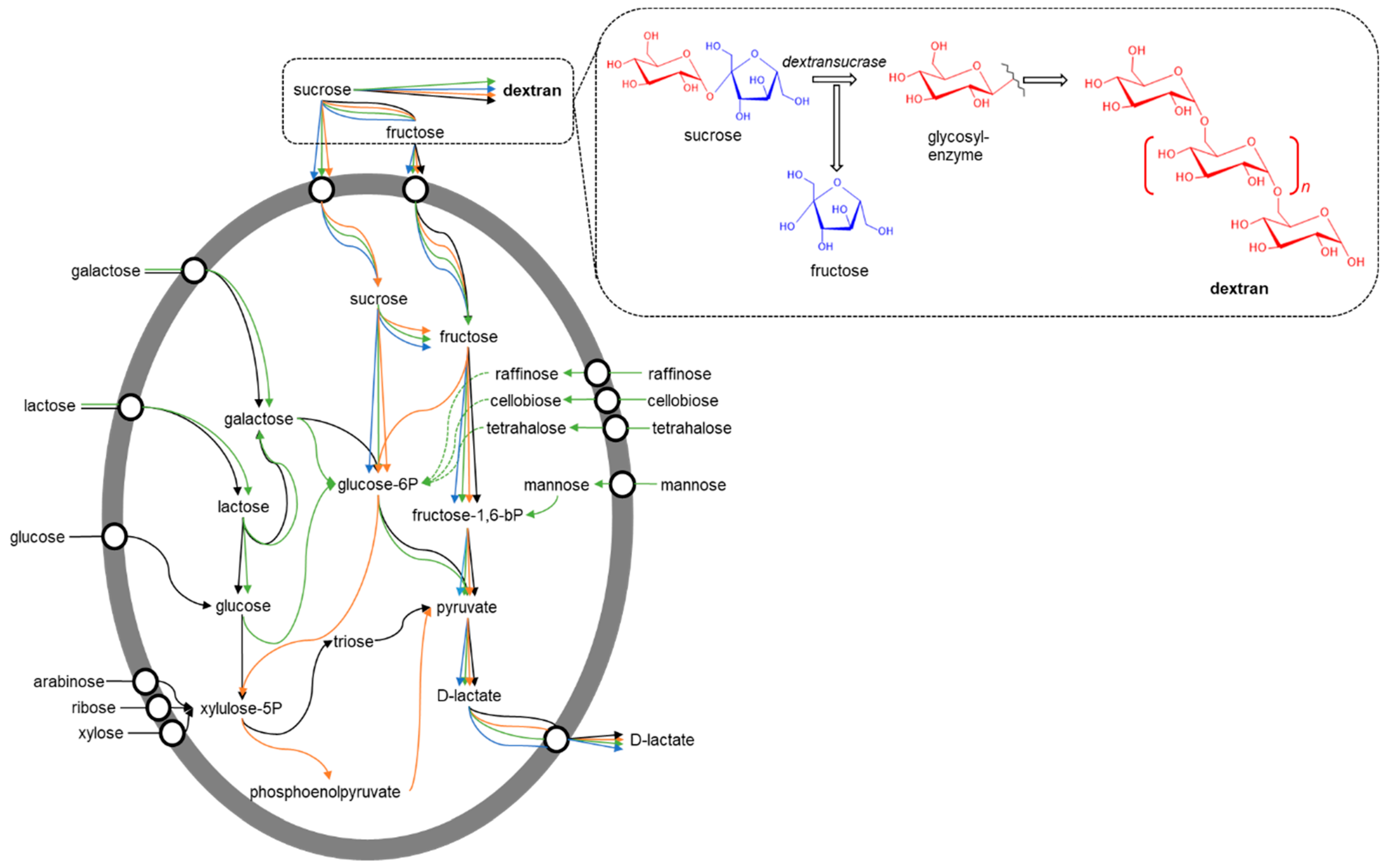
Dextran is synthesized in a particular way by LAB when exposed to a medium with sucrose as a carbon source [8]. In some LAB (e.g., Lactobacillus), sucrose can enter the cell directly via the phosphotransferase system (PTS) and metabolize to form d-lactate or become dextran [9]. Bacterial dextransucrases, located extracellularly, are responsible for hydrolyzing sucrose in its fructose and glucose monomers, forming an intermediate with glucose (glycosyl-enzyme) to later carry out their polymerization and form dextran [10], while the resulting fructose enters the bacteria through PTS to meet its metabolic demand [11], as shown in Figure 2. LAB that report dextran production are mainly of the genus Leuconostoc, Weissella, Lactobacillus, and Streptococcus [10], which have been isolated from different plant sources (e.g., Agave salmiana and pummelo) [12,13] and fermented products (e.g., rice batter, cabbage, idli batter, and pickles) [14,15,16,17]. However, dextran can also be synthesized via enzymatic, directly using dextransacarases (sucrose: 1,6-α-d-glucan 6-α-d-glucosyltransferase, EC 2.4.1.5) [18], which polymerize the glucoses of the sucrose in dextran, as shown at the top of Figure 2.
In industry, dextran is obtained through the culture of Leuconostoc mesenteroides NRRL B512, because it is considered a bacteria generally recognized as safe (GRAS) and very stable [21]. The fermentation of the NRRL B512 strain is carried out in a sucrose medium, which is nourished with yeast extracts, malt extracts, casein, peptone, and tryptone; in addition, low levels of calcium and phosphate are added [22,23,24,25]. During fermentation, the pH drops from 7 to 5 due to the generation of lactic acid; therefore, non-ionic detergents are usually added to maintain the stability of the bacteria and its enzymes [8]. In the clinical area, dextran is usually obtained from the acid hydrolysis (e.g., sulfuric and hydrochloric acids) of the native dextran from Leuconostoc mesenteroides NRRL B512, which allows controlling the molecular weights of the resulting dextrans [26,27].
3. Characteristics of Dextran
Dextran is a complex glucan formed by a main chain of d-glucoses linked by α-(1→6) bonds with possible branches of d-glucoses with α-(1→4), α-(1→3), or α-(1→2) bonds [28], as shown in Figure 3. Dextrans have molecular weight of up to 440 MDa [29], and they can be classified into two types according to the length of their chains—those with molecular weight greater than 40 kDa are simply called dextrans [8], while those with molecular weight less than 40 kDa can be called oligodextrans [30]. However, some authors name high molecular weight dextran, low molecular weight dextran, and just dextran to generalize (as in this review) [31].
Some reports affirm the synthesis of dextran is affected by the amount of substrate—they already found the highest dextran production using sucrose between 10% and 20% [12,32], because sucrose causes an inhibitory effect that affects the production of the EPS [33]. However, the variations in the molecular weight and the types and proportions of branches in each dextran depend on the producing strain (or enzyme) and the fermentation (or synthesis) conditions, making each glucan complex and different [15,34]. Table 1 compiles studies that report the synthesis of dextrans by different bacteria, in which a variation in the molecular weight and the branches including dextrans produced by bacteria of the same genus is appreciated. For example, the genus Leuconostoc mesenteroides generally synthesizes dextrans with a main chain linked by α-(1→6) bonds and branches with α-(1→3) bonds [12]; however, the study by Sawale and Lele [16] reported that the UICT/L18 strain Leuconostoc mesenteroides synthesized a branched dextran with α-(1→4) bonds. Siddiqui et al. [35] stated that the Leuconostoc mesenteroides KIBGE-IB22 strain produced a branched dextran with α-(1→3) and β-(2→6) bonds. However, most of the dextrans synthesized by LAB (i.e., Leuconostoc, Lactobacillus, and Weissella) have only α-(1→6) and α-(1→3) bonds with percentages between 52% and 97% and 3% and 48%, respectively.
There are other factors that affect both the molecular weight and the branching of dextran; for example, if fermentations with more than 25 °C are used, dextrans with greater branching are produced [36,37], while at temperatures lower than 25 °C, they are obtained with higher molecular weight [23,38]. In addition, with the increase in the concentration of sucrose, the yield of dextran decreases, but so does its degree of branching [37,39]. The commercial dextran synthesized by Leuconostoc mesenteroides NRRL B512 became the most important glucan due to its stability caused by composition of 95% α-(1→6) bonds and 5% branches with α-(1→3) bonds [21].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Dextrans synthesized by lactic acid bacteria (LAB) isolated from different sources.
| LAB | Substrates | Dextran | Reference | |||
|---|---|---|---|---|---|---|
| Genus | Subspecies | Source | Molecular Weight | Linkages | ||
| Leuconostoc mesenteroides | SD1 | Agave salmiana | Sucrose 10% | α-(1→6) 93% α-(1→3) 7% | [12] | |
| SD23 | Agave salmiana | Sucrose 10% | α-(1→6) 95% α-(1→3) 5% | |||
| SF2 | Agave salmiana | Sucrose 10% | α-(1→6) 94% α-(1→3) 6% | |||
| SF3 | Agave salmiana | Sucrose 10% | α-(1→6) 74% α-(1→3) 26% | |||
| NRRL B512 | Milk permeate 5% | <10 kDa | [31] | |||
| NRRL B512 | Sucrose 3% | <40 kDa | ||||
| NRRL B512 | Molasses | [40] | ||||
| NRRL B512 | Cheese whey 6% | |||||
| NRRL B512 | Molasses + Cheese whey 2–10% | |||||
| CM9 | Camel milk | Sucrose 2% | 230 MDa | [29] | ||
| CM30 | Camel milk | Sucrose 2% | 390 MDa | |||
| SM34 | Sheep milk | Sucrose 2% | 210 MDa | |||
| RTF10 | Meat products | Sucrose 2% | 440 MDa | |||
| BA08 | Fermented rice batter | Whey + Sucrose 5% | α-(1→6) 93% α-(1→3) 7% | [14] | ||
| KIBGE-IB22 | Indigenous source | Sucrose 10% | 15–20 MDa | α-(1→6) α-(1→3) β-(2→6) | [35] | |
| KIBGE-IB22M20 | Mutant | Sucrose 10% | 25–40 MDa | α-(1→6) α-(1→3) | ||
| BD1710 | Tomato juice + Sucrose 15% | 635 kDa | α-(1→6) 94% α-(1→3) 6% | [41] | ||
| ATCC 10830 | Residual pineapple juice + Sucrose 15% | 960 kDa | [42] | |||
| AA1 | Fermented cabbage | Sucrose 10% | 10–40 MDa | [15] | ||
| NRRL B-1149 | Sucrose 10% + Maltose 5% | α-(1→6) 52% α-(1→3) 48% | [43] | |||
| UICT/L18 | Fermented idli batter | Sucrose 22% | 970 kDa | α-(1→6) α-(1→4) | [16] | |
| Leuconostoc carnosum | CUPV411 | Apple must | Sucrose 2% | 358 MDa | α-(1→6) α-(1→3) | [44] |
| Leuconostoc citreum | SK24.002 | Fermented pickles | Sucrose 10% | 46 MDa | α-(1→6) 56% α-(1→3) 44% | [17] |
| Leuconostoc sp. | LS1 | Sauerkraut | Sucrose 15% | [45] | ||
| LI1 | Idli batter | Sucrose 15% | ||||
| Lactobacillus mali | CUPV271 | Ropy slime of cooked ham | Sucrose 2% | 123 MDa | α-(1→6) α-(1→3) | [44] |
| Lactobacillus sakei | MN1 | Meat products | Sucrose 2% | 170 MDa | [29] | |
| Lactobacillus plantarum | DM5 | Ethnic fermented beverage | Sucrose 5% | α-(1→6) 87% α-(1→3) 13% | [46] | |
| LS3 | Stool samples | Sucrose 15% | [47] | |||
| Lactobacillus gasseri | LV1 | Vaginal swabs | Sucrose 15% | |||
| LV2 | Vaginal swabs | Sucrose 15% | ||||
| LS1 | Stool samples | Sucrose 15% | ||||
| Lactobacillus acidophilus | LV3 | Vaginal swabs | Sucrose 15% | |||
| LV4 | Vaginal swabs | Sucrose 15% | ||||
| LV5 | Vaginal swabs | Sucrose 15% | ||||
| Lactobacillus fermentum | LS2 | Stool samples | Sucrose 15% | |||
| Lactobacillus satsumensis | NRRL B-59839 | Water kefir grains | Sucrose 20% | α-(1→6) 55% α-(1→3) 45% | [48] | |
| Weissella cibaria | 27 | Kimchi | Sucrose 20% | 12 MDa | α-(1→6) | [49] |
| 10M | Sucrose 0.5 M | 5–40 MDa | [50] | |||
| YB-1 | Pickle cabbage | Sucrose 5% | 390 kDa | α-(1→6) 96% α-(1→3) 4% | [51] | |
| RBA12 | Pummelo | Sucrose 2% | α-(1→6) 97% α-(1→3) 3% | [13] | ||
| 11GM-2 | Sour milk | Sucrose 20% | >20 MDa | α-(1→6) 95% α-(1→3) 5% | [52] | |
| JAG8 | Apple peel | Sucrose 10% | 800 kDa | [53] | ||
| JAG8 | Apple peel | Sucrose 2% | 177 kDa | α-(1→6) 93% α-(1→3) 7% | [54] | |
| MG1 | Sucrose 10% | α-(1→6) | [55] | |||
| CMGDEX3 | Cabbage | Sucrose 15% | >2 MDa | α-(1→6) 97% α-(1→3) 3% | [56] | |
| Weissella confusa | PP29 | Romanian yoghurt | Sucrose 8% | 120–870 kDa | α-(1→6) 96% α-(1→3) 4% | [57] |
| PP29 | Romanian yoghurt | Milk + Sucrose 8% | 120–250 kDa | α-(1→6) 96% α-(1→3) 4% | ||
| R003 | Sugar cane juice | Sucrose 10% | 10 MDa | α-(1→6) 97%α-(1→3) 3% | [58] | |
| QS813 | Sourdough starters | Sucrose 5% | 160 MDa | α-(1→6) 97% α-(1→3) 3% | [59] | |
| A3/2-1 | Fermented cassava | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | [52] | |
| A4/2-1 | Fermented cassava | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| F3/2-2 | Fermented cassava | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| E5/2-1 | Fermented cassava | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| G3/2-2 | Fermented cassava | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| 8CS-2 | Sour milk | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| 11GU-1 | Sour milk | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| 11GT-2 | Sour milk | Sucrose 10% | >20 MDa | α-(1→6) 97% α-(1→3) 3% | ||
| K1-Lb5 | Kimchi | Sucrose 20% | 1158 kDa | α-(1→6) α-(1→3) | [60] | |
| Cab3 | Fermented cabbage | Sucrose 5% | [61] | |||
| Weissella sp. | TN610 | Pear | Sucrose 4% | 180 kDa | α-(1→6) 96% α-(1→3) 4% | [62] |
Some researchers prefer the enzymatic use to produce dextran due to the advantages it presents over fermentation; for example, microbial enzymes can be easily modified by molecular engineering, they do not need growth factors such as yeast and meat extracts, and they produce specific and pure metabolites, which translates into a reduction in processing costs [63,64]. Table 2 shows high and low molecular weight dextrans produced via enzymatic. In these studies, most of the enzymes have been modified to synthesize high molecular weight dextrans (up to 23 MDa) [65]; however, enzymes have also been designed to directly produce low molecular weight dextrans or even to achieve the synergic interaction between enzymes to obtain low molecular weight dextrans with varied molecular weights [66]. Most of the characteristics of dextrans produced via enzymatic depend on the type of enzyme (source or method of obtaining it); however, the molecular weights are also directly related to the concentration of the substrate [67]. On the other hand, the dextrans obtained by this via report a variation between α-(1→6) and α-(1→3) bonds; specifically, they show a decrease in the percentage of α-(1→6) bonds compared to the dextrans obtained by fermentation. It even allowed obtaining totally linear dextrans [65] and with α-(1→6) and α-(1→2) bonds [68]. In general, the modification or transformation of enzymes by engineering makes it possible to obtain dextrans with desired characteristics.
On the other hand, low molecular weight dextrans are obtained mainly through acid hydrolysis of a high molecular weight dextran; however, there are studies that use enzymatic hydrolysis to produce them [66,70], as shown in the Table 3. In these studies, enzymes derived from LAB were modified or cloned to hydrolyze high molecular weight dextrans to shorter chain dextrans (up to 500 Da) [71]. The dextrans obtained specifically presented α-(1→6) and α-(1→2) bonds in different proportions depending on the enzyme and the substrate (dextran). Furthermore, the cloning of enzymes allowed them to be used not only for dextran hydrolysis, but also to polymerize high molecular weight dextrans from short chain dextrans [72].
4. Properties of Dextran
Variations in dextran characteristics (e.g., molecular weight and branching) cause its properties to be different [15,34]. The main chain of dextran with α-(1→6) bonds adopts a helical shape, which is modified by the presence of branches (α-(1→2), α-(1→3) or α-(1→4)), such that the linear structure of glucan is repeatedly folded [73,74,75].
The solubility and rheological properties of dextran are affected by its molecular weight and branching [76]. The solubility of polymers refers to the interaction of the molecule with water through interactions by hydrogen bridges [77]. Some research asserts that if the dextran molecule were totally linear (without branches), it would be totally soluble, because its hydroxy groups (–OH) would be exposed to interact with water molecules [78]. Other investigations affirm that the greater the number of branches, the greater the solubility of the dextran due to the increase in amorphous areas in the molecule that favor water adsorption and retention [73,74,75]. There are even reports that, in general, all low molecular weight polysaccharides have a higher solubility compared to long chain polysaccharides [43]. There is no direct relationship between the characteristics of the molecule and the variation of the properties [14,35,41,68,79]. However, regardless of the degree of solubility, dextrans are considered soluble EPS due to their ability to incorporate large amounts of water and form hydrogels [80].
The rheology and viscosity of polymers show their behavior as flow or deformation under an applied force, respectively [81], which is associated with –OH groups that easily interact with other molecules through hydrogen bonds, which are they break during shear [80]. Generally, the viscosity of dextran is directly related to the concentration and the shear rate, which means that at low concentrations they have a Newtonian behavior (independent of the shear rate) and at high concentrations their behavior is non-Newtonian (or pseudoplastic) [29]. Other studies show that the viscosity is also in direct relation to the molecular weight of the dextran, since as one increases, the other increases [82].
On the other hand, the flexibility of the polymers is determined as a function of the temperature; however, the temperatures vary depending on the intermolecular forces, crystallinity, and the size of the polymer [83]. Linear amorphous polymers have characteristics like glass at low temperatures—that is, little flexibility due to the zero mobility of the polymer chains [84]. With increasing temperature, they tend to become leathery (at the glass transition temperature, Tg), then rubbery and finally melt (at the melting temperature, Tm) [83]. During this transformation process, polymers show their most flexible point [84]. In crystalline polymers, the Tg is high due to intermolecular forces between the polymer chains. In short chain polymers, the Tm is low because the entropy is low, whereas long chains tend to be less mobile with high entropies, so the Tm is high [83].
5. Concluding Remarks and Future Perspectives
The production of dextran occurs mainly by a fermentation with LAB in a medium with sucrose; however, the enzymatic route has been used because it is a direct method in which other products or metabolites are not produced. The enzymatic pathway has also allowed the modification of enzymes to produce dextrans with specific desired characteristics. The characteristics of dextran depend on the LAB or enzyme of origin, which makes each dextran unique in terms of molecular structure, molecular weight, and branching, which cause variations in the viscosity and flexibility, and thermal and rheological properties. In addition, these properties vary depending on the temperature, concentration, and force applied to each dextran, which allows its application in different areas such as food, pharmaceutics, cosmetics, and research.
In medicine, high molecular weight dextran (between 40 and 70 kDa) has been used as an extender, anticoagulant, antithrombotic, osmotic agent, and intravenous plasma lubricant; in addition, it is used as a cryopreservative for vaccines and organs [8,28]. In the cosmetic industry, dextran has been used as a thickening and moisturizing agent, and its reducing property allows it to be used as an anti-aging agent. In the research area, dextran is useful to generate chromatography matrices, immobilize biosensors, generate nanoparticles, and form emulsions [28].
However, the most explored application of dextran is in the food industry, as it is used in baking and confectionery due to its moisturizing, stabilizing, and preserving effects, improving the flavor, texture, and consistency of ice creams, sweets, breads, flours, and jellies. In meat, vegetable, and cheese products, it has been added to retard oxidation; therefore, they are preservatives of texture, aroma, and flavor [8,15,28,85]. In addition, dextran has been proposed to be used as coatings or biodegradable film-forming agents [86,87], and as potential prebiotics (low molecular weight dextrans) [71,88].
The versatility of dextran has attracted attention in the past decade, and for this reason, the sources of obtaining LAB and the manipulation of enzymes that produce it have increased in such a way that the variety of dextrans has also increased its applications. However, the full characterization of each dextran produced is still incomplete and it would be worth studying so that they could compete with commercial dextran from Leuconostoc mesenteroides NRRL B512.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Voidarou, C.; Antoniadou, M.; Rozos, G.; Tzora, A.; Skoufos, I.; Varzakas, T.; Lagiou, A.; Bezirtzoglou, E. Fermentative foods: Microbiology, biochemistry, potential human health benefits and public health issues. Foods 2021, 10, 1–27. [Google Scholar]
- Mozzi, F. Lactic acid bacteria. In Encyclopedia of Food Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 501–508. ISBN 9780123849533. [Google Scholar]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. Open Access 2018, 6, 89–94. [Google Scholar]
- Narvhus, J.A.; Axelsson, L. Lactic acid bacteria. In Encyclopedia of Food Sciences and Nutrition; Academic Press: San Diego, CA, USA, 2003; pp. 3465–3472. ISBN 012227055X. [Google Scholar]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from lactic acid bacteria: Perspectives and challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef]
- Laws, A.; Gu, Y.; Marshall, V. Biosynthesis, characterisation, and design of bacterial exopolysaccharides from lactic acid bacteria. Biotechnol. Adv. 2001, 19, 597–625. [Google Scholar] [CrossRef]
- Yadav, H.; Karthikeyan, C. Natural polysaccharides: Structural features and properties. In Polysaccharide Carriers for Drug Delivery; Maiti, S., Jana, S., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; pp. 1–17. ISBN 9780081025536. [Google Scholar]
- Heinze, T.; Liebert, T.; Heublein, B.; Hornig, S. Functional polymers based on dextran. In Polysaccharides II; Klemm, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 199–291. ISBN 9783540371021. [Google Scholar]
- Prechtl, R.M.; Janßen, D.; Behr, J.; Ludwig, C.; Küster, B.; Vogel, R.F.; Jakob, F. Sucrose-induced proteomic response and carbohydrate utilization of lactobacillus sakei TMW 1.411 during dextran formation. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kothari, D.; Das, D.; Patel, S.; Goyal, A. Dextran and food application. In Polysaccharides; Gopal Ramawat, K., Mérillon, J.-M., Eds.; Springer International Publishing: Gewerbestrasse, AG, Switzerland, 2014; pp. 735–752. ISBN 9783319162973. [Google Scholar]
- Besrour-Aouam, N.; Fhoula, I.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Najjari, A.; Prieto, A.; Ruas-Madiedo, P.; López, P.; Ouzari, H.-I. The role of dextran production in the metabolic context of Leuconostoc and Weissella Tunisian strains. Carbohydr. Polym. 2020, 253, 117254. [Google Scholar] [CrossRef]
- Castro-Rodríguez, D.; Hernández-Sánchez, H.; Yáñez-Fernández, J. Structural characterization and rheological properties of dextran produced by native strains isolated of Agave salmiana. Food Hydrocoll. 2019, 90, 1–8. [Google Scholar]
- Baruah, R.; Maina, N.H.; Katina, K.; Juvonen, R.; Goyal, A. Functional food applications of dextran from Weissella cibaria RBA12 from pummelo (Citrus maxima). Int. J. Food Microbiol. 2016, 242, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Lule, V.K.; Singh, R.; Pophaly, S.D.; Poonam Tomar, S.K. Production and structural characterisation of dextran from an indigenous strain of Leuconostoc mesenteroides BA08 in whey. Int. J. Dairy Technol. 2016, 69, 520–531. [Google Scholar] [CrossRef]
- Aman, A.; Siddiqui, N.N.; Qader, S.A.U. Characterization and potential applications of high molecular weight dextran produced by Leuconostoc mesenteroides AA1. Carbohydr. Polym. 2012, 87, 910–915. [Google Scholar] [CrossRef]
- Sawale, S.D.; Lele, S.S. Statistical optimization of media for dextran production by Leuconostoc sp., isolated from fermented Idli batter. Food Sci. Biotechnol. 2010, 19, 471–478. [Google Scholar] [CrossRef]
- Miao, M.; Huang, C.; Jia, X.; Cui, S.W.; Jiang, B.; Zhang, T. Physicochemical characteristics of a high molecular weight bioengineered α-D-glucan from Leuconostoc citreum SK24.002. Food Hydrocoll. 2015, 50, 37–43. [Google Scholar] [CrossRef]
- Alcalde, M.; Plou, F.J.; Martín, M.T.; Remaud, M.; Monsan, P.; Ballesteros, A. Stability in the presence of organic solvents of dextransucrase from Leuconostoc mesenteroides NRRL B-512F immobilized in calcium-alginate beads. Prog. Biotechnol. 1998, 15, 535–540. [Google Scholar]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Ajdić, D.; McShan, W.M.; McLaughlin, R.E.; Savić, G.; Chang, J.; Carson, M.B.; Primeaux, C.; Tian, R.; Kenton, S.; Jia, H.; et al. Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 2002, 99, 14434–14439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BeMiller, J.N. Dextran. In Encyclopedia of Food Sciences and Nutrition; Academic Press: San Diego, CA, USA, 2003; pp. 1772–1773. ISBN 012227055X. [Google Scholar]
- DeBelder, A.N. Dextran. In Industrial gums-Polysaccharides and their derivates; Whistler, R.L., BeMiller, J.N., Eds.; Academic Press, Inc.: London, UK, 1993; pp. 399–425. ISBN 0127462538. [Google Scholar]
- Jeanes, A. Dextrans and pullulans: Industrially significant α-D-Ggucans. In Extracellular Microbial Polysaccharides; Sandford, P.A., Allen, L., Eds.; American Chemical Society: Washington, DC, USA, 1977; pp. 284–298. ISBN 9780841203723. [Google Scholar]
- Koepsell, H.J.; Tsuchiya, H.M. Enzymatic synthesis of dextran. J. Bacteriol. 1952, 63, 293–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, H.M.; Koepsell, H.J.; Corman, J.; Bryant, G.; Bogard, M.O.; Feger, V.H.; Jackson, R.W. The effect of certain cultural factors on production of dextransucrase by Leuconostoc mesenteroides. J. Bacteriol. 1952, 64, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Wolff, I.A.; Mehltretter, C.L.; Mellies, R.L.; Watson, P.R.; Hofreiter, B.T.; Patrick, P.L.; Rist, C.E. Production of clinical-type dextran-Partial hydrolytic depolymerization and fractionation of the dextran from Leuconostoc mesenteroides strain NRRL B-512. Ind. Eng. Chem. 1954, 46, 370–377. [Google Scholar] [CrossRef]
- Zief, M.; Brunner, G.; Metzendorf, J. Fractionation of partially hydrolyzed dextran. J. Am. Pharm. Assoc. Am. 1956, 48, 119–121. [Google Scholar] [CrossRef]
- Bhavani, A.L.; Nisha, J. Dextran-The polysaccharide with versatile uses. Int. J. Pharma Bio Sci. 2010, 1, 569–573. [Google Scholar]
- Zarour, K.; Llamas, M.G.; Prieto, A.; Rúas-Madiedo, P.; Dueñas, M.T.; de Palencia, P.F.; Aznar, R.; Kihal, M.; López, P. Rheology and bioactivity of high molecular weight dextrans synthesised by lactic acid bacteria. Carbohydr. Polym. 2017, 174, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.H.; Killen, D.A.; Jolly, P.C.; Kirtley, J.H. Low molecular weight dextran in vascular surgery: Prevention of early thrombosis following arterial recosntruction in 85 cases. Ann. Surg. 1966, 163, 764–770. [Google Scholar] [CrossRef]
- Esmaeilnejad-Moghadam, B.; Mokarram, R.R.; Hejazi, M.A.; Khiabani, M.S.; Keivaninahr, F. Low molecular weight dextran production by Leuconostoc mesenteroides strains: Optimization of a new culture medium and the rheological assessments. Bioact. Carbohydr. Diet. Fibre 2019, 18, 100181. [Google Scholar] [CrossRef]
- Sarwat, F.; Qader, S.A.U.; Aman, A.; Ahmed, N. Production & characterization of a unique dextran from an indigenous Leuconostoc mesenteroides CMG713. Int. J. Biol. Sci. 2008, 4, 379–386. [Google Scholar]
- Martinez-Espindola, J.P.; Lopez-Munguia, C.A. On the kinetics of dextransucrase and dextran synthesis in batch reactors. Biotechnol. Lett. 1985, 7, 483–486. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Lazado, C.C. Nutritional impacts on fish mucosa: Immunostimulants, pre- and probiotics. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: London, UK, 2015; pp. 211–272. ISBN 9780124171862. [Google Scholar]
- Siddiqui, N.N.; Aman, A.; Silipo, A.; Qader, S.A.U.; Molinaro, A. Structural analysis and characterization of dextran produced by wild and mutant strains of Leuconostoc mesenteroides. Carbohydr. Polym. 2014, 99, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Sabatie, J.; Choplin, L.; Moan, M.; Doublier, J.L.; Paul, F.; Monsan, P. The effect of synthesis temperature on the structure of dextran NRRL B 512F. Carbohydr. Polym. 1988, 9, 87–101. [Google Scholar] [CrossRef]
- Braswell, E.; Goodmatu, A.; Kurt, G. Studies on the enzymatic synthesis of dextran. Part II. J. Polym. Sci. 1962, 61, 143–154. [Google Scholar] [CrossRef]
- Hamdy, M.; Gardner, E.; Stahly, G.; Weiser, H. Factors affecting production and clarification of dextran. Ohio J. Sci. 1954, 54, 317–328. [Google Scholar]
- Hellman, N.N.; Tsuchiya, H.M.; Rogovin, S.P.; Lamberts, B.L.; Tobin, R.; Glass, C.A.; Stringer, C.S.; Jackson, R.W.; Senti, F.R. Controlled enzymatic synthesis of dextran-Conditions for producing clinically suitable molecular weight. Ind. Eng. Chem. 1955, 47, 1593–1598. [Google Scholar] [CrossRef]
- Moosavi-Nasab, M.; Gavahian, M.; Yousefi, A.R.; Askari, H. Fermentative production of dextran using food industry wastes. Int. J. Nutr. Food Eng. 2010, 4, 1921–1923. [Google Scholar]
- Han, J.; Hang, F.; Guo, B.; Liu, Z.; You, C.; Wu, Z. Dextran synthesized by Leuconostoc mesenteroides BD1710 in tomato juice supplemented with sucrose. Carbohydr. Polym. 2014, 112, 556–562. [Google Scholar] [CrossRef]
- Vega, J.; Sibaja, M.; Lopretti, M. Biosíntesis de dextranos de alto peso molecular mediante la inoculación con Leuconostoc síntesis y caracterización de hierro-dextranos. Innotec 2012, 7, 55–58. [Google Scholar]
- Shukla, R.; Shukla, S.; Bivolarski, V.; Iliev, I.; Ivanova, I.; Goyal, A. Structural characterization of insoluble dextran produced by Leuconostoc mesenteroides NRRL B-1149 in the presence of maltose. Food Technol. Biotechnol. 2011, 49, 291–296. [Google Scholar]
- Llamas-Arriba, M.G.; Puertas, A.I.; Prieto, A.; López, P.; Cobos, M.; Miranda, J.I.; Marieta, C.; Ruas-Madiedo, P.; Dueñas, M.T. Characterization of dextrans produced by Lactobacillus mali CUPV271 and Leuconostoc carnosum CUPV411. Food Hydrocoll. 2019, 89, 613–622. [Google Scholar] [CrossRef]
- Subathra Devi, C.; Reddy, S.; Mohanasrinivasan, V. Fermentative production of dextran using Leuconostoc spp. isolated from fermented food products. Front. Biol. 2014, 9, 244–253. [Google Scholar] [CrossRef]
- Das, D.; Goyal, A. Characterization and biocompatibility of glucan: A safe food additive from probiotic Lactobacillus plantarum DM5. J. Sci. Food Agric. 2014, 94, 683–690. [Google Scholar] [CrossRef]
- Kareem, A.J.; Salman, J.A.S. Production of dextran from locally lactobacillus spp. isolates. Rep. Biochem. Mol. Biol. 2019, 8, 278–286. [Google Scholar]
- Côté, G.L.; Skory, C.D.; Unser, S.M.; Rich, J.O. The production of glucans via glucansucrases from Lactobacillus satsumensis isolated from a fermented beverage starter culture. Appl. Microbiol. Biotechnol. 2013, 97, 7265–7273. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.J.; Chen, Z.; Chen, P.T.; Ng, I.S. Production, characterization and antibacterial activity of exopolysaccharide from a newly isolated Weissella cibaria under sucrose effect. J. Biosci. Bioeng. 2018, 126, 769–777. [Google Scholar] [CrossRef]
- Hu, Y.; Gänzle, M.G. Effect of temperature on production of oligosaccharides and dextran by Weissella cibaria 10 M. Int. J. Food Microbiol. 2018, 280, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Chen, Y.; Wang, C.; Yang, R.; Bin, X. Purification and characterization of exopolysaccharide produced by Weissella cibaria YB-1 from pickle Chinese cabbage. Int. J. Biol. Macromol. 2018, 120, 1315–1321. [Google Scholar] [CrossRef]
- Malang, S.K.; Maina, N.H.; Schwab, C.; Tenkanen, M.; Lacroix, C. Characterization of exopolysaccharide and ropy capsular polysaccharide formation by Weissella. Food Microbiol. 2015, 46, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Tingirikari, J.M.R.; Kothari, D.; Shukla, R.; Goyal, A. Structural and biocompatibility properties of dextran from Weissella cibaria JAG8 as food additive. Int. J. Food Sci. Nutr. 2014, 65, 686–691. [Google Scholar] [CrossRef]
- Rao, T.J.M.; Goyal, A. A novel high dextran yielding Weissella cibaria JAG8 for cereal food application. Int. J. Food Sci. Nutr. 2013, 64, 346–354. [Google Scholar] [CrossRef]
- Galle, S.; Schwab, C.; Dal Bello, F.; Coffey, A.; Gänzle, M.G.; Arendt, E.K. Influence of in-situ synthesized exopolysaccharides on the quality of gluten-free sorghum sourdough bread. Int. J. Food Microbiol. 2012, 155, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.Z.; Siddiqui, K.; Arman, M.; Ahmed, N. Characterization of high molecular weight dextran produced by Weissella cibaria CMGDEX3. Carbohydr. Polym. 2012, 90, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Rosca, I.; Petrovici, A.R.; Peptanariu, D.; Nicolescu, A.; Dodi, G.; Avadanei, M.; Ivanov, I.C.; Bostanaru, A.C.; Mares, M.; Ciolacu, D. Biosynthesis of dextran by Weissella confusa and its In vitro functional characteristics. Int. J. Biol. Macromol. 2018, 107, 1765–1772. [Google Scholar] [CrossRef]
- Netsopa, S.; Niamsanit, S.; Sakloetsakun, D.; Milintawisamai, N. Characterization and rheological behavior of dextran from Weissella confusa R003. Int. J. Polym. Sci. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Liu, N.; Huang, W.; Cheng, X.; Wang, F.; Zhang, B.; Chen, J.; Jiang, H.; Omedi, J.O.; Li, Z. Syneresis rate, water distribution, and microstructure of wheat starch gel during freeze-thaw process: Role of a high molecular weight dextran produced by Weissella confusa QS813 from traditional sourdough. Cereal Chem. 2018, 95, 117–129. [Google Scholar]
- Park, J.H.; Ahn, H.J.; Kim, S.G.; Chung, C.H. Dextran-like exopolysaccharide-producing Leuconostoc and Weissella from kimchi and its ingredients. Food Sci. Biotechnol. 2013, 22, 1047–1053. [Google Scholar] [CrossRef]
- Shukla, S.; Goyal, A. 16S rRNA-based identification of a glucan-hyperproducing weissella confusa. Enzyme Res. 2011, 2011, 250842. [Google Scholar] [CrossRef] [Green Version]
- Bejar, W.; Gabriel, V.; Amari, M.; Morel, S.; Mezghani, M.; Maguin, E.; Fontagné-Faucher, C.; Bejar, S.; Chouayekh, H. Characterization of glucansucrase and dextran from Weissella sp. TN610 with potential as safe food additives. Int. J. Biol. Macromol. 2013, 52, 125–132. [Google Scholar] [CrossRef]
- Hara, K.Y.; Araki, M.; Okai, N.; Wakai, S.; Hasunuma, T.; Kondo, A. Development of bio-based fine chemical production through synthetic bioengineering. Microb. Cell Fact. 2014, 13, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of microbial enzymes in food industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Dobruchowska, J.M.; Pijning, T.; López, C.A.; Kamerling, J.P.; Dijkhuizen, L. Residue Leu940 has a crucial role in the linkage and reaction specificity of the glucansucrase GTF180 of the probiotic bacterium lactobacillus reuteri 180. J. Biol. Chem. 2014, 289, 32773–32782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, W.; Zhang, H.; Zhang, Y.; Hu, X. Biosynthesis of oligodextrans with different Mw by synergistic catalysis of dextransucrase and dextranase. Carbohydr. Polym. 2014, 112, 387–395. [Google Scholar] [CrossRef]
- Falconer, D.J.; Mukerjea, R.; Robyt, J.F. Biosynthesis of dextrans with different molecular weights by selecting the concentration of Leuconostoc mesenteroides B-512FMC dextransucrase, the sucrose concentration, and the temperature. Carbohydr. Res. 2011, 346, 280–284. [Google Scholar] [CrossRef]
- Vettori, M.H.P.B.; Franchetti, S.M.M.; Contiero, J. Structural characterization of a new dextran with a low degree of branching produced by Leuconostoc mesenteroides FT045B dextransucrase. Carbohydr. Polym. 2012, 88, 1440–1444. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Shi, Q.; Maina, N.H.; Juvonen, M.; Tenkanen, M.; Goyal, A. Weissella confusa Cab3 dextransucrase: Properties and in vitro synthesis of dextran and glucooligosaccharides. Carbohydr. Polym. 2014, 101, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Luo, J.; Pinelo, M.; Wan, Y. Directing filtration to narrow molecular weight distribution of oligodextran in an enzymatic membrane reactor. J. Memb. Sci. 2018, 555, 268–279. [Google Scholar] [CrossRef] [Green Version]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.W.; Gibson, G.R.; Rastall, R.A. The prebiotic effect of α-1,2 branched, low molecular weight dextran in the batch and continuous faecal fermentation system. J. Funct. Foods 2013, 5, 1938–1946. [Google Scholar] [CrossRef]
- Brison, Y.; Fabre, E.; Moulis, C.; Portais, J.C.; Monsan, P.; Remaud-Siméon, M. Synthesis of dextrans with controlled amounts of α-1,2 linkages using the transglucosidase GBD-CD2. Appl. Microbiol. Biotechnol. 2010, 86, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Balani, K.; Verma, V.; Agarwal, A.; Narayan, R. Physical, thermal, and mechanical properties of polymers. In Biosurfaces: A Materials Science and Engineering Perspective; Balani, K., Verma, V., Agarwal, A., Narayan, R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 329–344. ISBN 9781118299975. [Google Scholar]
- Monnerie, L. Mechanical properties of polymeric materials. In Statistical Models for the Fracture of Disordered Media; Herrmann, H.J., Roux, S., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 1990; pp. 66–76. ISBN 9781483296128. [Google Scholar]
- Whistler, R.L. Solubility of polysaccharides and their behavior in solution. In Carbohydrates in Solution; Horace, S.I., Ed.; American Chemical Society: Washington, DC, USA, 1973; pp. 242–255. ISBN 9780841201781. [Google Scholar]
- Guo, M.Q.; Hu, X.; Wang, C.; Ai, L. Polysaccharides: Structure and solubility. In Solubility of Polysaccharides; Xu, Z., Ed.; Intech Open: London, UK, 2017; pp. 7–21. ISBN 9789535136507. [Google Scholar]
- Antoniou, E.; Tsianou, M. Solution properties of dextran in water and in formamide. J. Appl. Polym. Sci. 2012, 125, 1681–1692. [Google Scholar] [CrossRef]
- Suner, S.S.; Sahiner, M.; Sengel, S.B.; Rees, D.J.; Reed, W.F.; Sahiner, N. Responsive biopolymer-based microgels/nanogels for drug delivery applications. In Stimuli Responsive Polymeric Nanocarriers for Drug Delivery Applications: Volume 1: Types and Triggers; Hamdy Makhlouf, A.S., Abul-Thabit, N.Y., Eds.; Woodhead Publishing: Duxford, UK, 2018; pp. 453–500. ISBN 9780081019979. [Google Scholar]
- Gil, E.C.; Colarte, A.I.; El Ghzaoui, A.; Durand, D.; Delarbre, J.L.; Bataille, B. A sugar cane native dextran as an innovative functional excipient for the development of pharmaceutical tablets. Eur. J. Pharm. Biopharm. 2008, 68, 319–329. [Google Scholar] [CrossRef]
- Campos, F.D.S.; Ferrari, L.Z.; Cassimiro, D.L.; Ribeiro, C.A.; De Almeida, A.E.; Gremião, M.P.D. Effect of 70-kDa and 148-kDa dextran hydrogels on praziquantel solubility. J. Therm. Anal. Calorim. 2016, 123, 2157–2164. [Google Scholar] [CrossRef] [Green Version]
- Morris, B.A. Rheology of polymer melts. In The Science and Technology of Flexible Packaging; Morris, B.A., Ed.; William Andrew Publishing: Oxford, UK, 2017; pp. 121–147. ISBN 9780323242738. [Google Scholar]
- Masuelli, M.A. Dextrans in aqueous solution. Experimental review on intrinsic viscosity measurements and temperature effect. J. Polym. Biopolym. Phys. Chem. 2013, 1, 13–21. [Google Scholar]
- Seymour, R.B.; Carraher, C.E. Thermal properties of polymers. In Structure—Property Relationships in Polymers; Seymour, R.B., Carraher, C.E., Eds.; Springer: Boston, MA, USA, 1984; pp. 83–93. ISBN 978-1-4684-4748-4. [Google Scholar]
- Van De Velde, K.; Kiekens, P. Biopolymers: Overview of several properties and consequences on their applications. Polym. Test. 2002, 21, 433–442. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.; Zannini, E.; Czerny, M.; Arendt, E.K. Influence of dextran-producing Weissella cibaria on baking properties and sensory profile of gluten-free and wheat breads. Int. J. Food Microbiol. 2014, 172, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Moncayo-Martínez, D.C.; Buitrago-Hurtado, G.; Néstor, Y.; Algecira-Enciso, A. Películas comestibles a base de un biopolímero tipo dextrana Edible films based of dextran biopolymer. Agron. Colomb. 2016, 34, 107–109. [Google Scholar]
- Díaz-Montes, E.; Yáñez-Fernández, J.; Castro-Muñoz, R. Dextran/chitosan blend film fabrication for bio-packaging of mushrooms (Agaricus bisporus). J. Food Process. Preserv. 2021. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.; Brison, Y.; Remaud-Simeon, M.; Monsan, P.; Gibson, G.R.; Rastall, R.A. In vitro fermentation of linear and α-1,2-branched Dextrans by the human Fecal microbiota. Appl. Environ. Microbiol. 2011, 77, 5307–5315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Number of publications related to dextran synthesis by lactic acid bacteria (source: Scopus; keywords: Bacteria, Leuconostoc, Weissella, Lactobacillus, and Streptococcus; accessed: 3 May 2021).
Figure 1.
Number of publications related to dextran synthesis by lactic acid bacteria (source: Scopus; keywords: Bacteria, Leuconostoc, Weissella, Lactobacillus, and Streptococcus; accessed: 3 May 2021).

Figure 2.
General carbohydrate pathways and dextran synthesis in lactic acid bacteria: Leuconostoc (black), Weissella (orange), Lactobacillus (blue), and Streptococcus (green). Adapted from [9,10,11,19,20].

Figure 3.
Structural model of dextran. Backbone formed by d-glucose units with α-(1→6) bonds and different branching bonds: (a) α-(1→4), (b) α-(1→3), and (c) α-(1→2).
Figure 3.
Structural model of dextran. Backbone formed by d-glucose units with α-(1→6) bonds and different branching bonds: (a) α-(1→4), (b) α-(1→3), and (c) α-(1→2).

Table 2.
Dextrans synthesized by enzymes isolated from different sources.
| Enzyme | Microorganisms | Substrates | Dextran | Reference | |||
|---|---|---|---|---|---|---|---|
| Type | Obtaining | Genus | Subspecies | Molecular Weight | Linkages | ||
| Glucansucrase GTF180 | Isolated | Leuconostoc reuteri | 180 | Maltose 100 mM + Sucrose 100 mM | ~23 MDa | α-(1→6) 78% α-(1→3) 22% | [65] |
| Glucansucrase L940G | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~17 MDa | α-(1→6) 85% α-(1→3) 15% | [65] | |
| Glucansucrase L940C | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~17 MDa | α-(1→6) 74% α-(1→3) 26% | [65] | |
| Glucansucrase L940A | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~19 MDa | α-(1→6) 84% α-(1→3) 16% | [65] | |
| Glucansucrase L940S | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~20 MDa | α-(1→6) 84% α-(1→3) 16% | [65] | |
| Glucansucrase L940M | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~19 MDa | α-(1→6) 72% α-(1→3) 28% | [65] | |
| Glucansucrase L940E | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~19 MDa | α-(1→6) 73% α-(1→3) 27% | [65] | |
| Glucansucrase L940F | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~20 MDa | α-(1→6) 93% α-(1→3) 7% | [65] | |
| Glucansucrase L940W | Mutated | 180 | Maltose 100 mM + Sucrose 100 mM | ~6 MDa | α-(1→6) 100% | [65] | |
| Dextransucrase B-512FMC | Mutated | Leuconostoc mesenteroides | B-512FMC | Sucrose 20 mM | 20–341 kDa | - | [67] |
| Dextransucrase B-512FMC | Mutated | B-512FMC | Sucrose 50 mM | 49–431 kDa | - | [67] | |
| Dextransucrase B-512FMC | Mutated | B-512FMC | Sucrose 100 mM | 63–514 kDa | - | [67] | |
| Dextransucrase B-512FMC | Mutated | B-512FMC | Sucrose 200 mM | 126–787 kDa | - | [67] | |
| Dextransucrase B-512FMC | Mutated | B-512FMC | Sucrose 1000 mM | 1645 kDa | - | [67] | |
| Dextransucrase FT045B-Dextranase | Isolated | Leuconostoc mesenteroidesPenicillium | FT045Bsp. | Sucrose 400 mM | ~92 kDa | α-(1→6) 98% α-(1→2) 2% | [68] |
| Dextransucrase (DE3)/pET28-dexYG | Engineered | Escherichia coli | BL21 | Sucrose 10% | 5 kDa | - | [66] |
| Dextransucrase (DE3)/pET28-dexYG-Dextranase | Engineered | Escherichia coliPenicillium aculeatum | BL21- | Sucrose 10% | 10–20 kDa | - | [66] |
| Dextransucrase WcCab3 | Isolated | Weissella confusa | Cab3 | Sucrose 5% | 178 kDa | α-(1→6) 97% α-(1→3) 3% | [69] |
Table 3.
Dextrans synthesized by hydrolysis enzymatic.
| Enzyme | Microorganisms | Substrates | Dextran | Reference | |||
|---|---|---|---|---|---|---|---|
| Type | Obtaining | Genus | Subspecies | Molecular Weight | Linkages | ||
| Dextranase | Isolated | Penicillium | Sp. | Dextran 40 kDa | 5–8 kDa | - | [70] |
| α-1,2 transglucosidase | Engineered | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa | 0.5 kDa | α-(1→6) 75% α-(1→2) 25% | [71] |
| α-1,2 transglucosidase | Engineered | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa | 1 kDa | α-(1→6) 68% α-(1→2) 32% | [71] |
| Transglucosidase GBD–CD2 | Cloned | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa + Sucrose 292 mM | 10 kDa | α-(1→6) 62% α-(1→2) 38% | [72] |
| Transglucosidase GBD–CD2 | Cloned | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa + Sucrose 292 mM | 40 kDa | α-(1→6) 63% α-(1→2) 37% | [72] |
| Transglucosidase GBD–CD2 | Cloned | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa + Sucrose 292 mM | 70 kDa | α-(1→6) 62–67% α-(1→2) 33–38% | [72] |
| Transglucosidase GBD–CD2 | Cloned | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa + Sucrose 292 mM | 70 kDa | α-(1→6) 81–88% α-(1→2) 12–19% | [72] |
| Transglucosidase GBD–CD2 | Cloned | Leuconostoc mesenteroides | NRRL B-1299 | Dextran 70 kDa + Sucrose 292 mM | 2000 kDa | α-(1→6) 64% α-(1→2) 36% | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Díaz-Montes, E. Dextran: Sources, Structures, and Properties. Polysaccharides 2021, 2, 554-565. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030033
AMA Style
Díaz-Montes E. Dextran: Sources, Structures, and Properties. Polysaccharides. 2021; 2(3):554-565. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030033
Chicago/Turabian StyleDíaz-Montes, Elsa. 2021. "Dextran: Sources, Structures, and Properties" Polysaccharides 2, no. 3: 554-565. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030033