
Polysaccharide Vaccines: A Perspective on Non-Typhoidal Salmonella


1
Department of Biochemistry, Microbiology and Immunology, University of Saskatchewan, Saskatoon, SK S7N 5E5, Canada
2
Vaccine and Infectious Disease Organization, Saskatoon, SK S7N 5E3, Canada
3
David Braley Center for Antibiotic Discovery, M.G. DeGroote Institute for Infectious Disease Research, Department of Biochemistry and Biomedical Sciences, McMaster University, Hamilton, ON L8S 4L8, Canada
*
Author to whom correspondence should be addressed.
Polysaccharides 2021, 2(3), 691-714; https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030042
Submission received: 14 July 2021
/
Revised: 16 August 2021
/
Accepted: 26 August 2021
/
Published: 11 September 2021
(This article belongs to the Collection Current Opinion in Polysaccharides)
Abstract
:Polysaccharides are often the most abundant antigens found on the extracellular surfaces of bacterial cells. These polysaccharides play key roles in interactions with the outside world, and for many bacterial pathogens, they represent what is presented to the human immune system. As a result, many vaccines have been or currently are being developed against carbohydrate antigens. In this review, we explore the diversity of capsular polysaccharides (CPS) in Salmonella and other selected bacterial species and explain the classification and function of CPS as vaccine antigens. Despite many vaccines being developed using carbohydrate antigens, the low immunogenicity and the diversity of infecting strains and serovars present an antigen formulation challenge to manufacturers. Vaccines tend to focus on common serovars or have changing formulations over time, reflecting the trends in human infection, which can be costly and time-consuming. We summarize the approaches to generate carbohydrate-based vaccines for Salmonella, describe vaccines that are in development and emphasize the need for an effective vaccine against non-typhoidal Salmonella strains.
1. Salmonellosis
Salmonellosis is one of the most common foodborne diseases worldwide [1]. Symptoms of Salmonellosis include abdominal cramps, diarrhea, nausea, vomiting, chills, fever, and headache, and typically develop 6–72 h after ingestion. These symptoms can last up to 2–14 days depending on the initial concentration of bacteria ingested, serotype, age and the immune status of the host [2,3]. Salmonellosis in humans is primarily caused by non-typhoidal Salmonella (NTS), consisting of >1000 different serovars, with serovars Enteritidis and Typhimurium being the most common [4]. NTS infections can cause bacteremia in rare cases (i.e., 3–10%), which can lead to further complications, such as endocarditis, septic shock, infection of the liver, spleen, biliary tract and urinary tract, mesenteric lymphadenitis, pneumonia, osteomyelitis, pulmonary and brain abscess, empyema, meningitis, CNS infections and death [3]. In addition, perforation of the gut and necrosis of Peyer’s patches can result in toxic encephalopathy [5]. The risk of bacteremia is higher in immunosuppressed individuals, infants, and the elderly, and the number of infections has risen in recent times in correlation with increased incidences of the human immunodeficiency virus (HIV) and malaria.
The global surveillance of Salmonellosis is primarily conducted by the US Centers for Disease Control and Prevention (CDC) and the World Health Organization (WHO). In the United States, Salmonella strains cause 1.35 million reported infections, 26,500 hospitalizations and 420 deaths annually [6]. According to the United States Department of Health and Human Services, Salmonella infections are the leading cause of hospitalizations and death among foodborne pathogens. For Canadians, travel to endemic regions was the most common factor for reported NTS infections [4], specifically, travel to Central and South America, the Caribbean, Mexico and the Dominican Republic have seen the highest rates of infection [4]. Between 2009 and 2013, a yearly average of 6500 infections was reported throughout Canada, and in 2018, 7300 infections were reported, accounting for 48% of all notifiable isolates reported within the year [4]. In developing countries, Salmonella is responsible for 20% of childhood diarrheal infections and the mortality rate can reach as high as 24%, with certain serotypes having higher mortality rates than others [3]. However, 60–80% of Salmonellosis cases are thought to be either not diagnosed at all or are classified as sporadic cases [2,4].
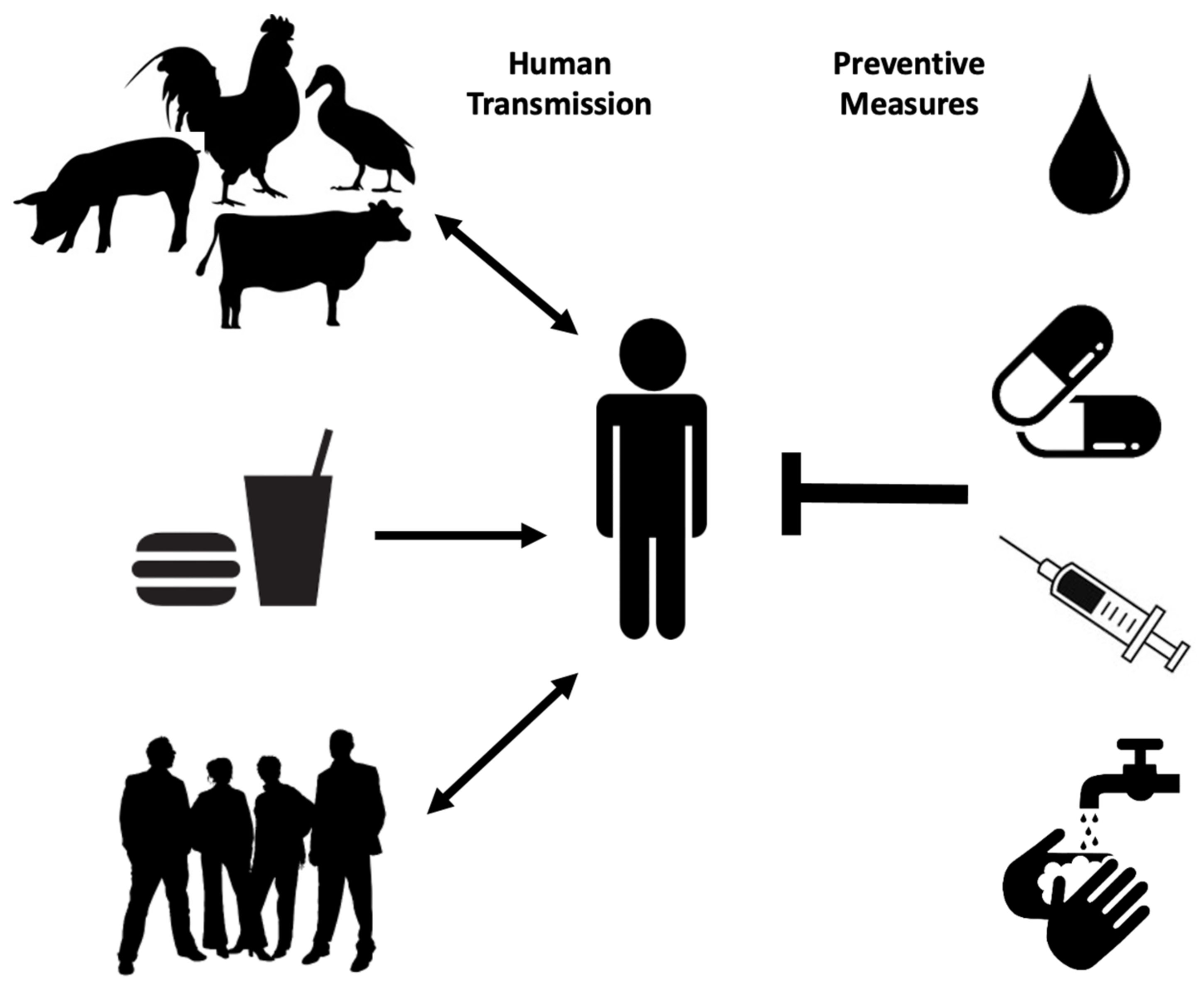
Human transmission occurs through ingestion of Salmonella-contaminated food (i.e., raw or undercooked poultry) or drinks, contact with infected animals, especially food animals and pets, and animal feed (Figure 1). Person-to-person transmission through the fecal–oral route is also common and is facilitated by inadequate handwashing [3]. Preliminary diagnosis of salmonellosis in Canada is mainly based on symptoms and travel history and can be confirmed by serotyping (i.e., whole genome sequencing) each specimen from stool, rectal swabs, vomit, urine, deep tissue wounds, or sterile sites at the public health laboratories [4,7]. Treatment is based on symptoms and typically includes electrolyte replacement and rehydration, due to fluid loss that occurs through diarrhea and vomiting [2]. Commonly used first-line antibiotics include fluoroquinolones for adults and azithromycin for children. Ceftriaxone is used as an alternative first-line treatment [8]. However, antimicrobial therapy is usually reserved for immunocompromised patients, infants, children, the elderly and for severe cases of gastroenteritis, as unnecessary administration of antibiotics can select for resistant strains and contributes to prolonged shedding [2].
Multidrug-resistant Salmonella strains have emerged as far back as 1989 [3]. As of 1997, more than 70% of clinical isolates showed resistance to ampicillin, tetracycline, sulphonamides, streptomycin, and chloramphenicol, with few strains even resistant to nalidixic acid, trimethoprim, sulfamethoxazole, and kanamycin [9]. Ceftiofur resistance is considered significant because it is often correlated with resistance to other antibiotics commonly used, such as ceftriaxone, cefoxitin, amoxicillin-clavulanic acid, and ampicillin. As extra-intestinal Salmonellosis in young children and pregnant women is treated with Ceftriaxone, it is recognized as a drug of very high importance to human medicine. Thus, patients infected with even moderately Ceftriaxone-resistant Salmonella strains (i.e., Salmonella serovar Heidelberg) are at elevated risk of ceftriaxone therapy failure [10]. Proper sanitation, access to clean water supplies and treatment of infected individuals are among the best prophylactic measures to prevent the spread of Salmonellosis especially in endemic regions (Figure 1) [3]. A vaccine against common NTS serovars would also represent a big step in reducing the global incidence of Salmonellosis and bacteremia.
2. Capsular Polysaccharides
Extracellular polysaccharides are carbohydrate-based polymers synthesized and secreted into the external environment by bacteria [11]. They include lipooligosaccharides (LOS), lipopolysaccharides (LPS), capsular polysaccharides (CPS), teichoic acids, lipoteichoic acids, glycoproteins, and peptidoglycans [11]. They can be present on the cell surface as slime with no visible attachment (often called exopolysaccharides; EPS), or as CPS covalently linked to the bacterial cell surface or anchored to the cell surface via lipid A (LOS and LPS) [12]. In rare cases, CPS can be released from the cell due to poor stability of the phosphodiester interactions between the CPS and the cell surface phospholipid molecules. Conversely, at times, EPS can also remain tightly associated with the cell surface [13]. Therefore, the nomenclature does not always match the biological reality.
CPS are highly hydrated molecules containing 95–99% water and are composed of regularly repeating units of one or more monosaccharides linked via glycosidic bonds. They can take the form of homo- or heteropolymers. Homopolymers are composed of identical monosaccharides, while heteropolymers are composed of at least two different monosaccharides [14]. CPS are incredibly diverse in terms of their structure and composition because they can be composed of different monosaccharides connected by varying glycosidic linkages and modified with a diverse array of organic and inorganic molecules [12,14,15]. The structural diversity and the presence of immunological epitopes on CPS depend on a multitude of factors, including the type of monosaccharides, variations in the glycosidic linkages, the presence of organic and inorganic molecules and the introduction of side chains, configuration of the anomeric center of the sugar molecule, and enantiomeric form of the sugar [12,16].
During capsule biosynthesis, activated precursor molecules (monophosphate and diphosphate sugars) are assembled by inner membrane enzymes to form the growing polysaccharide chain. Following this, in the case of LPS assembly, the seven transport proteins Lpt A–G move the newly synthesized polysaccharide through the periplasm and across the outer membrane to the cell surface [17,18]. Studies have indicated the presence of transenvelope assembly complexes that coordinate simultaneous biosynthesis, export and translocation of the polysaccharide [19]. In some instances, identical translocation pathways are used for certain capsular polysaccharides and LPS O-Antigens [20].
CPS producing bacteria are highly diverse and include both animal and plant pathogens such as Escherichia coli, Salmonella serovar Typhi, non-typhoidal Salmonella serovars, Shigella species, Neisseria species, Vibrio species, Streptococcus species, Cryptococcus neoformans, Staphylococcus aureus, Campylobacter jejuni, Bacteroides fragilis, Haemophilus influenza, Aeromonas salmonicida, Acetobacter methanolicus, Burkholderia mallei, and Francisella tularensis [16,21,22,23,24,25,26,27,28].
2.1. Capsular Polysaccharides—Classification
Historically, the classification of CPS started with E. coli, which has over 80 different CPS, known as K antigens [29]. Certain K antigens have identical polysaccharide chains and differ only in the modifications of the polysaccharides [30]. According to the classification system introduced by Whitfield and Roberts (Table 1), a CPS can be classified into four categories [19]. Group 1 CPS are acidic polysaccharides containing uronic acid and tend to be similar in structure and are often inaccurately classified as colanic acid [19]. The most prominent feature distinguishing group 1 CPS and colanic acid is that most wild-type bacteria are unable to produce colanic acid when grown at 37 °C on common lab media, whereas they can produce CPS [19].
Group 2 capsules are linked to the cell surface through α-glycerophosphatidic acid. In some bacteria, cytidine 5′-monophospho-3-deoxy-d-manno-2-octulosonic acid (CMP-KDO) provides a link between the phospholipid and the polysaccharide. The expression of group 2 capsules is significantly influenced by the growth temperature, and they are not typically expressed below 20 °C. Group 2 capsules harbour an α-glycerophosphate moiety at the terminus [31]. Group 3 capsules are largely similar to group 2 capsules but differ in not being regulated by growth temperature. Both group 2 and 3 capsules are co-expressed with colanic acid and many different types of O-serogroups. Although there is only a hypothetical α-glycerophosphate moiety at the terminus of group 3 capsules, genetic data support the presence of a common anchor for both group 2 and group 3 capsules. Despite the lack of information regarding the group 3 capsule biosynthesis pathway, functionally interchangeable CPS homologs have been identified between groups 2 and 3 capsules [31].
When bacteria produce a group 4 capsule as the only serotype-specific polysaccharide, it is classified as the O-Antigen of that isolate [19]. Group 4 CPS were previously classified as group 1 CPS and as a result, became known as the ‘O-Antigen capsules’ [31]. Group 4 CPS, despite many similarities, are far more diverse than group 1 CPS. One example is the group 4 representative K40 antigen that was assigned to group 1 due to the presence of acetamido sugars in its repeat unit. K40 antigen was then shown to be surface expressed as smooth LPS and as an unlinked O-Antigen capsule and was reclassified into group 4 [12,32]. This confusion in classification has arisen since the K40 antigen is co-expressed with a neutral LPS linked polymer such as O8 or O9 antigen [33]. Group 1 and 4 antigens are subgrouped into KLPS and capsular K antigens, where KLPS is made up of low molecular weight K antigenic oligosaccharides containing few repeat units that are linked to the cell surface through lipid A. KLPS, however, are different from LPS with the serological O-Antigen found on the same cell [19]. Capsular K antigens are made up of high molecular weight antigens that form the capsule structure responsible for masking O-Antigen in serotyping. These capsular K antigens are not linked to lipid A core as in LPS molecules [33].
During translocation, group 1 and 4 capsule assembly occurs at the periplasmic face of the plasma membrane and uses the Wzx pathway, while the assembly of group 2 and 3 capsules occurs at the cytoplasmic face of the plasma membrane and an ABC transporter is used for translocation. During the assembly of capsules from all four groups, the sequential action of glycosyltransferase enzymes joins individual repeat units together to elongate the polysaccharide [33].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Classification of capsules. Classification is based on Whitfield and Roberts 1999 research article [34]. Reprinted with permission from ref [34]. Copyright obtained 2021, John Wiley and Sons.
| Characteristic | Group 1 | Group 2 | Group 3 | Group 4 |
|---|---|---|---|---|
| Thermostability | Yes | No | No | Yes |
| Thermoregulated (expressed below 20 °C) | No | Yes | No | No |
| Co-expressed with colanic acid | No | Yes | Yes | Yes |
| Co-expressed with O serogroups | Limited | Many | Many | Mainly O8, O9. Can be none |
| Former K antigen group | 1A | II | I/II or III | IB (O-Antigen capsules) |
| Capsule assembly | Periplasmic face of the plasma membrane | Cytoplasmic face of the plasma membrane | Cytoplasmic face of the plasma membrane | Periplasmic face of the plasma membrane |
| Export system | Wzx pathway | ABC transporter | ABC transporter | Wzx pathway |
| Translocation proteins | Wza, Wzc | KpsD, KpsE (may be KpsF) | KpsD (may be KpsE) | Unknown |
| Polymerization system | Wzy dependent | Processive | Processive | Wzy dependent |
| Direction of polymer chain growth | Reducing terminus | Non-reducing terminus | Non-reducing terminus (hypothetical) | Reducing terminus |
| Terminal lipid moiety | Lipid A core in KLPS; unknown for capsular K antigen | α-Glycerophosphate | α-Glycerophosphate (hypothetical) | Lipid A core in KLPS; unknown for capsular K antigen |
| Elevated levels of CMP-Kdo synthetase | No | Yes | No | No |
| Positively regulated by the Rcs system | Yes | No | No | No |
2.2. Capsular Polysaccharides—Function
As a major cell-surface component of many bacterial species, CPS can provide diverse functions. CPS often contribute to virulence, with innate and adaptive immune evasion; they also provide barrier protection, desiccation resistance, and act as lubricating agents and reduce friction over solid surfaces. For example, CPS facilitate swarming by Proteus mirabilis on solid surfaces [14]. It was observed that capsulated E. coli, Acinetobacter calcoaceticus, and Erwinia stewartii strains are more resistant to desiccation than non-capsulated strains [34]. In addition, CPS can allow bacterial adherence to host cells, surfaces, and to each other, thereby facilitating biofilm formation and colonization [35]. Biofilm formation is ubiquitous and can provide bacteria with nutritional advantages as well as protection from bacteriophage and phagocytic protozoa [12,36].
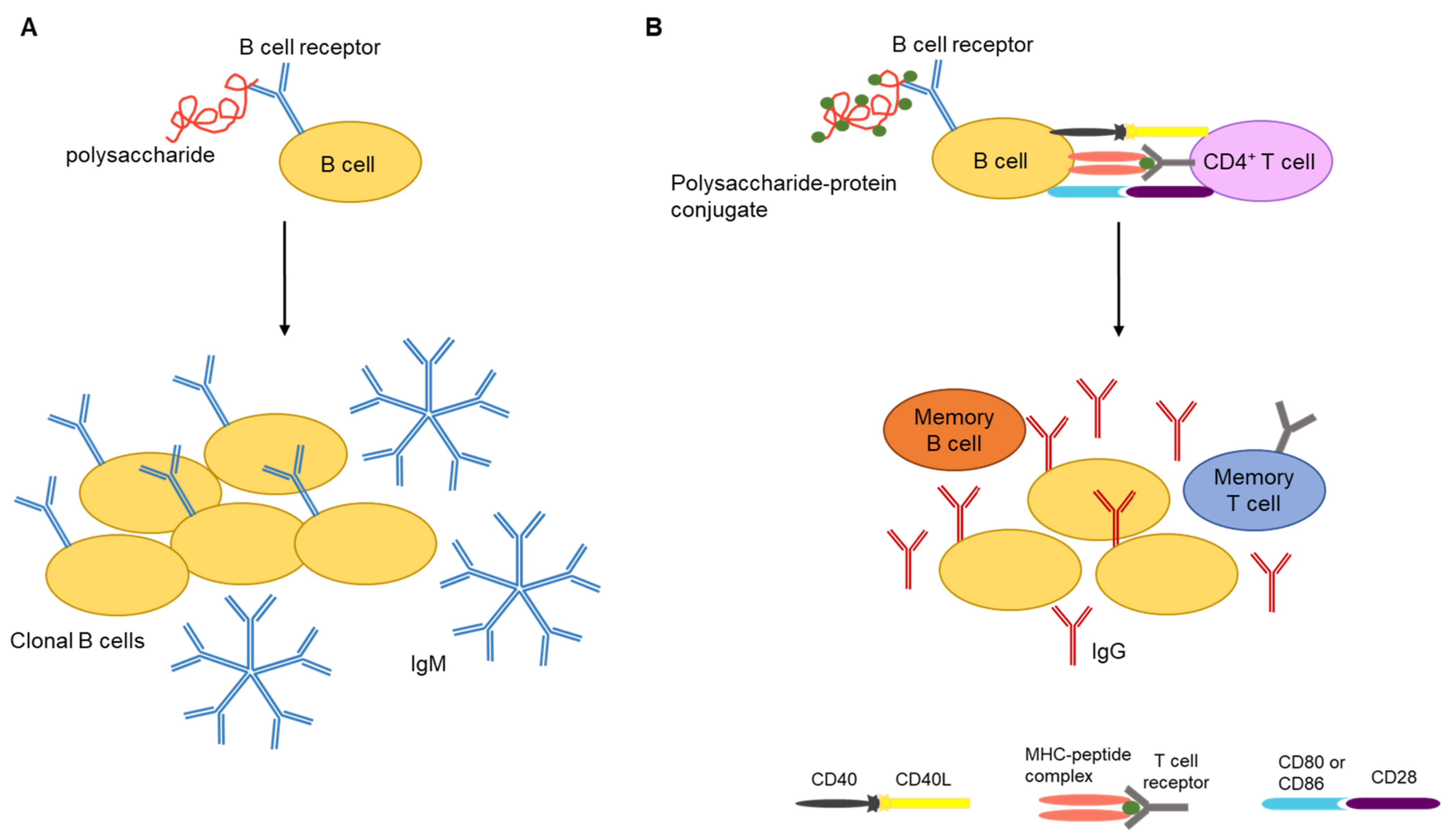
Studies have shown that pure carbohydrate antigens are T-cell-independent activators of B cells. Consequently, CPS antigens often induce immunoglobulin M (IgM) responses without a detectable level of IgG production (Figure 2A). The absence of antibody class switching from IgM to IgG and the inability to produce higher titers of antibody upon subsequent exposure to the antigen are hallmarks of T-cell-independent antigens [16]. However, conjugation of polysaccharides to carrier proteins can lead to T-cell activation, memory B-cell production, and the development of polysaccharide-specific immune responses (Figure 2B) [37].
2.3. Capsular Polysaccharides—Regulation
Despite the lack of concrete evidence in the mechanisms behind the regulation of CPS expression, it is hypothesized that reduced external osmolarity can act as a signal to increase CPS production [12]. In support of this argument, it has been shown that reduced external osmolarity leads to increased expression of Vi antigen in S. Typhi and expression of anionic polysaccharide alginate by Pseudomonas aeruginosa [38,39]. In E. coli, the expression of genes for the biosynthesis of EPS colanic acid is increased in the face of harsh environments and desiccation [34]. In general, the expression of a given type of CPS is associated with a unique infection. For example, K1 antigen-expressing E. coli are the primary cause of neonatal meningitis [12]. Remarkably, chemically identical capsular polysaccharides can be produced by different genera of bacteria. For example, E. coli K1 CPS is identical to Neisseria meningitides group B CPS, and E. coli K18, K22, and K100 CPS are identical to Haemophilus influenza serotype b CPS [12,40].
2.4. Capsular Polysaccharides—Salmonella and Other Selected Bacterial Species
There are two types of CPS found in Salmonella—the Vi capsule (also known as Vi antigen) produced by Salmonella serovar Typhi, and the O-Antigen capsule produced by NTS. The Vi antigen is composed of O-acetylated α-1,4-linked N-acetylgalactosaminuronic acid residues and is attached to the cell surface via a reducing terminal lipid structure [41]. The Vi capsule increases virulence and disease severity of S. Typhi infection by interfering with complement deposition and subsequent neutrophil chemotaxis and bacterial clearance [5,42]. Deletion of genes for Vi capsule biosynthesis can remarkably enhance neutrophil chemotaxis in vitro. In addition, the presence of the Vi capsule prevents complement activation through the alternative pathway, as C3b is unable to deposit on the capsule surface and promotes opsonophagocytosis [42,43]. It has also been demonstrated that the Vi capsule promotes immune evasion by masking LPS molecules and preventing recognition through pathogen recognition receptors (PRR). As a consequence, Vi-positive S. Typhi does not induce neutrophil influx and can disseminate systemically and cause a persistent infection [5]. Furthermore, the capsule-mediated suppression of TNF-α production by human monocytes leads to low serum concentrations of pyrogenic cytokines and thereby prevents the development of septic shock [44,45].
The O-Antigen capsule produced by NTS is co-regulated with cellulose and fimbriae to be part of the extracellular matrix (ECM). The group 4 O-Antigen capsules were shown to be important for attachment to surfaces, colonization, and desiccation resistance [23]. S. Typhimurium O-Antigen capsule was shown to be expressed both at lower and higher temperatures indicating that the capsule may play a role in bacterial survival inside and outside the host, as described for other bacterial species, such as Hyphomonas strain MHS-3 [46,47]. Salmonella O-Ag capsule was initially hypothesized to be a CPS that was distinct from very long chain LPS O-Ag and assembled by the yih operons [23]. However, in our recent study, we hypothesize that O-Ag capsule and very-long-chain LPS O-Ag are the same CPS and demonstrated that the yihUTSRQP operon is not directly involved in capsule assembly [47].
Campylobacter jejuni produces LOS and CPS that are genetically and biochemically similar to polysaccharide capsules in other Gram-negative bacteria [26]. Due to the presence of hypervariable polysaccharide biosynthesis genes, the structure of these polysaccharide monomers can vary between C. jejuni strains. Groups 2 and 3 CPS are part of the Penner antigen used for C. jejuni serotyping. It has been shown that C. jejuni CPS can be released to the surrounding environment in either naïve or lipid-free form depending on the presence or absence of deoxycholate; a component of bile salt. Even though the specific purpose of released CPS is unknown, it is regarded as important for the survival and pathogenesis of C. jejuni in foreign environments [26]. Treatment of C. jejuni CPS with phospholipase release a lipid-free product to the medium indicating that these CPS are substituted with a phospholipid in place of lipid A [26].
Another well-characterized CPS is produced by Staphylococcus aureus. Clinical isolates of S. aureus are classified into eight groups based on their CPS. The type 8 CPS is the most prevalent among bacteremic isolates and confers resistance to complement-mediated opsonization and phagocytosis [28]. Bacteroides fragilis produced zwitterionic CPS (contains both positive and negative charge) that can activate CD4+ T cells [16]. It has also been shown that administration of B. fragilis CPS alone protects Wistar rats against subsequent infection with B. fragilis [48].
Polysaccharide capsules, associated with the cell surface, can regulate interactions between the bacterium and the environment. Although CPS are often associated with Gram-negative bacteria, many Gram-positive bacteria synthesize lipoteichoic acids or teichoic acids containing sugar alcohols with phosphodiester linkages. These sugar alcohols play a key role in the attachment of bacteria to surfaces and their interactions with host innate and adaptive immune systems [49].
3. Polysaccharide Vaccines
Since the late 20th century, polysaccharide vaccines have helped save the lives of millions of people worldwide. However, there are several limitations associated with pure polysaccharide vaccines. Mainly these include hyporesponsiveness and poor immunogenicity in children under the age of two [50]. In addition, the response for polysaccharide antigens among two to five-year-old children, the elderly, and the immunocompromised individuals are not optimal, and these populations remain at an elevated risk of contracting bacterial diseases [37]. Furthermore, due to hyporesponsiveness, pure polysaccharide vaccines are unable to provide long-lasting herd immunity, which is a key element in the prevention of invasive diseases. Herd immunity is achieved when the majority of a community is immunized against a given infectious disease, such that even those who have not received immunization are protected as the spread of the disease is contained [51].
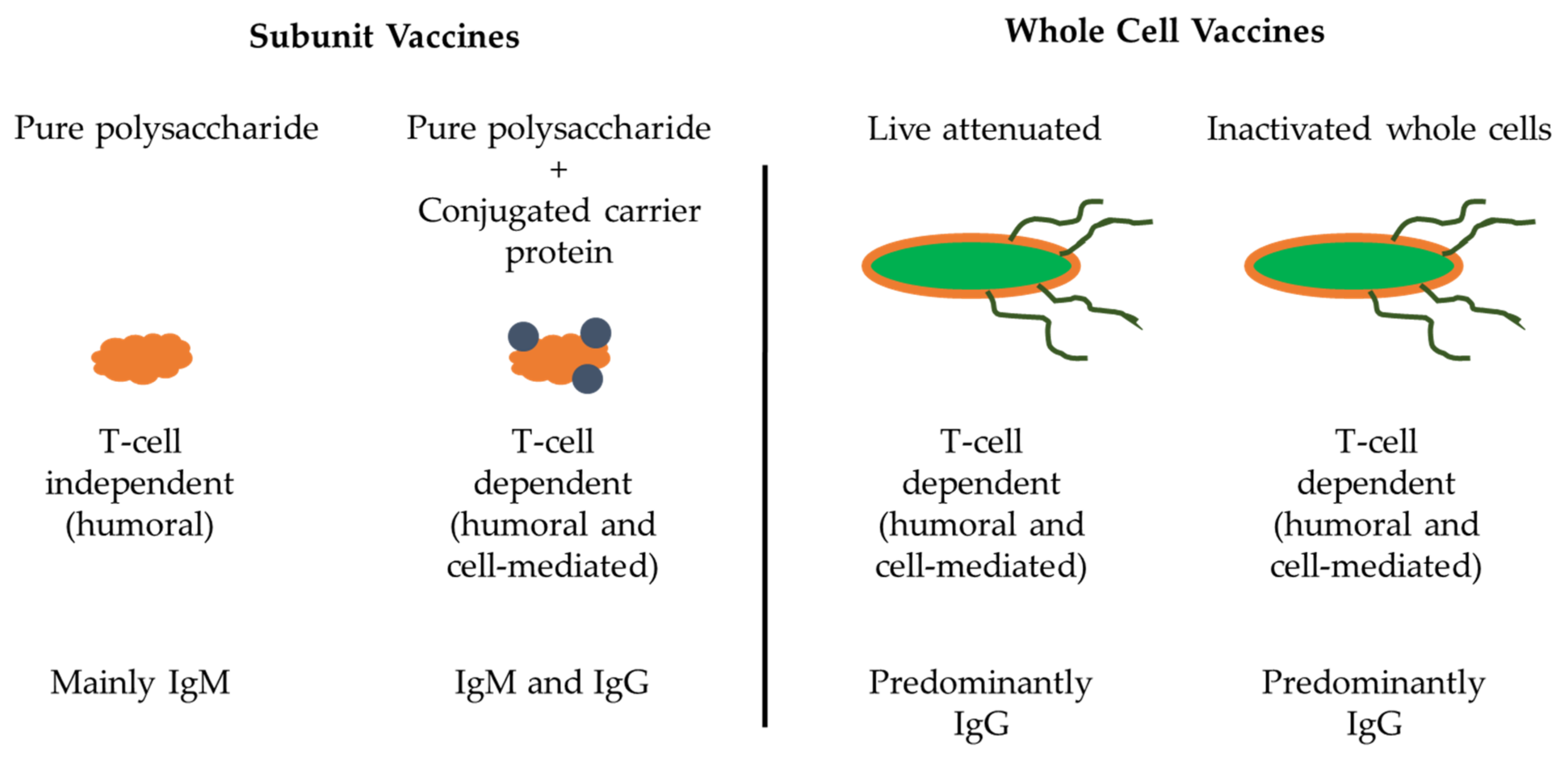
Polysaccharide vaccines are classified as inactivated subunit vaccines (Figure 3). To date pure polysaccharide vaccines have been produced for four encapsulated bacteria: Streptococcus pneumonia (pneumococcus), Neisseria meningitidis (meningococcus), Haemophilus influenzae type b (Hib) and S. Typhi [52]. The adaptive immune response to polysaccharide antigens is usually T-cell independent, implying that polysaccharides are capable of activating B cells in the absence of T helper cells [53]. However, the lack of T-cell induction and in turn failure of B-cell stimulation by T cells results in an absence of immunoglobulin class switching, induction of memory B cells, affinity maturation, prolonged antibody response, which often leads to hyporesponsiveness to subsequent vaccination and/or to natural infection.
Polysaccharide capsules can have numerous identical epitopes near each other and can crosslink immunoglobulin receptors on the surface of B cells for B-cell activation. However, these polysaccharides are not processed and presented on MHC class II molecules for T helper cell activation [54]. Although polysaccharide antigens are unable to induce memory B-cell production, they can activate previously formed memory cells, leading to the terminal differentiation of memory cells into plasma cells. However, this depletes the reservoir of memory B cells specific for a given polysaccharide antigen and could result in a condition called hyporesponsiveness, which is the inability to respond to subsequent exposure to the same antigen due to lack of memory B cells [55]. Repeated administration of the same polysaccharide antigen (s) can exacerbate this situation. In contrast, repeated administration of most protein vaccines leads to a booster effect resulting in progressively higher antibody titers. In addition, antibodies produced in response to polysaccharide antigens are predominantly IgM, which have less functional activity than IgG antibodies [55].
Marginal zone (MZ) B cells, which are produced in the spleen, B1 cells, and naïve B cells to a lesser extent, are the most important at recognizing polysaccharide antigens. B1 and MZ B cells express a large number of CD21 receptors and can bind to complement coated polysaccharides [55,56]. Individuals with congenital asplenia, splenic hypofunction or who have undergone splenectomy are highly susceptible to encapsulated bacterial diseases, as they have a depleted number of functional MZ B cells and are defective at removing opsonized bacteria [55]. Furthermore, the immune response to polysaccharide antigens is age dependent as children under the age of two are also susceptible to encapsulated bacteria since the MZ is not fully developed prior to this age and is unable to facilitate the development of B cells [57]. In addition, the presence of maternal antibodies and the effect of suppresser T cells also contribute to the lack of response to polysaccharide antigens in infants [37]. However, some zwitterionic polysaccharides such as capsules from B. fragilis, S. pneumonia serotypes 1 and 3, and N. meningitides serogroup A possess characteristics of T-cell-dependent antigens and are presented in an MHC class II-dependent manner. These antigens are immunogenic in even infants and young children [58,59].
To address the concerns of T-cell-independent immunogenicity of pure polysaccharide vaccines, polysaccharide antigens were chemically conjugated to carrier proteins in the late 1980s, resulting in polysaccharide–protein conjugates that were T-cell dependent and highly immunogenic [60]. The first polysaccharide–protein conjugate vaccine was produced for H. influenzae type-b. A conjugate vaccine for S. pneumonia was produced in 2000 and for N. meningitides in 2005. When these vaccines were introduced to the immunization schedules of multiple countries, it resulted in a significant reduction in the disease burden associated with encapsulated bacteria [61,62]. Conjugate vaccines have the advantage of inducing memory B-cell production, induction of immunity in children under the age of 2, and the capacity to overcome hyporesponsiveness [62,63].
Although immunogenic, the magnitude of the antibody response to conjugate vaccines and their persistence is age dependent. It was observed that in infants and toddlers, the serum antibody concentration following immunization with conjugate vaccines declined rapidly even though the immunological memory lasted much longer [64,65]. Despite multiple doses being administered to infants, the effectiveness of MenC and Hib vaccines declined within one year from initial immunization in the absence of a booster dose after the age of one [61,66]. The importance of persistent serum antibodies was demonstrated in vaccine failures where despite being immunized and having memory B cells, some children succumbed to meningococcal and Hib infections [67,68]. This is explained by the fact that encapsulated bacteria can invade and cause disease within a few hours, while memory B cells require several days to mount a protective antibody response [65,69]. Thus, protection against encapsulated bacterial infections requires high titers of persistent serum antibodies as well as induction of immune memory.
3.1. Polysaccharide Vaccines in Use
Current vaccines often have bi-, tri-, tetra-, and/or multivalent formulations, where the given number of separate protein or polysaccharide antigens are combined in a single vaccine. Although pure polysaccharide vaccines have been effective at saving lives for years, conjugation of polysaccharide antigens from N. meningitidis, S. pneumoniae, H. influenzae type b, and S. Typhi have generated T-cell-dependent antigens that are effective even in very young children [70].
3.1.1. Neisseria meningitidis
The first polysaccharide vaccine against N. meningitidis was used in the United States in 1984 and was given to all the new military personnel upon entering basic training. Upon the administration of this tetravalent polysaccharide vaccine, no cases of meningococcal infections related to serogroup Y or W135 were reported in the United States military [37]. However, this vaccine was poorly immunogenic among infants and young children [37]. A quadrivalent vaccine containing polysaccharide antigens from N. meningitidis serogroup A, C, Y, and W135 was initially licensed in the United States in 1981. A bivalent vaccine containing polysaccharide antigens from serogroup A and C is also available in some countries [52].
Serogroup C (MenC) conjugate vaccine containing either tetanus toxoid or inactivated diphtheria toxin, CRM197, was first licensed in the United Kingdom in 1999, while a quadrivalent A, C, Y, and W135 diphtheria toxoid conjugate vaccine became first available in North America in 2005 [71]. In addition, several other vaccine candidates have been tested, including A, C, Y, and W135 antigens conjugated to tetanus toxoid or conjugated to CRM197 [52]. As the serogroup B capsular polysaccharide is highly similar to cell surface glycoproteins on fetal brain tissue, which results in tolerance and reduced immunogenicity, the development of a pure or conjugate polysaccharide-based vaccine remains a significant hurdle in vaccine development against N. meningitides serogroup B [71]. One study compared the effectiveness between the meningococcal serogroups A and C pure polysaccharide vaccine (MACP) with the conjugate vaccine against serogroup C (MCC) since the use of MACP vaccine had raised concerns that this vaccine is leading to hyporesponsiveness to serogroup C polysaccharide antigen [72]. In this study, individuals who received a primary immunization of MACP were given a second immunization of either MACP or MCC. Those who received MCC produced significantly higher titers of IgG antibody than the individuals who received MACP as the second dose. In addition, the response to the second MACP vaccine was considerably lower than that following the first immunization. This supported the observation that repeated vaccination with the MACP vaccine is ineffective and leads to hyporesponsiveness towards the meningococcal serogroup C polysaccharide antigens in adults. However, it was possible to overcome the MACP induced hyporesponsiveness with subsequent vaccination with MCC [72].
Antibody titers induced in infants after immunization with N. meningitides serogroup C (MenC) vaccine fell below the threshold of protection in 50% of vaccinees by one year of age and only 12% of vaccinees maintained protection at four years of age [73]. There is very little evidence that immunization of infants with the MenC vaccine confers protection beyond one year, as antibody titers decrease rapidly following infant immunization [74]. When administered after one year of age, the MenC vaccine induced a more persistent antibody response, but sustained levels of protection were not observed until much later in childhood in many children [66]. In addition, the decline in protective immunity following infant immunization was also noted for the H. influenzae type b vaccine [52].
3.1.2. Streptococcus pneumonia
A pure polysaccharide hexavalent vaccine against S. pneumonia was first licensed in 1947 but was eventually withdrawn due to low sales and lack of acceptance [37]. A polyvalent vaccine containing 14 polysaccharide antigens became available in the United States in 1977 and was recommended for at-risk populations including the elderly, immunocompromised patients, and individuals with chronic cardiac, pulmonary, or renal diseases [75]. The vaccine had 93% efficacy following three years from initial immunization among immunocompetent individuals under 55 years of age, but only a 46% efficacy in individuals over 85 years of age [75]. Furthermore, the vaccine’s efficacy was shown to decline over time. However, this can be explained by the waning immunity and the exposure to additional pneumococcal serotypes over the years [75,76].
The current polyvalent pneumococcal vaccine was first licensed in 1983, is made of 23 immunologically distinct polysaccharide antigens and is effective in preventing disease in immunocompetent individuals [75]. Nonetheless, upon the introduction of the Hib conjugate vaccine in 1988, the disease burden among children in the United States declined by 50% per year from 1988 to 1991 [75]. In addition, several other pneumococcal vaccines are also in use. These include the seven-valent conjugate vaccine (PCV7) that contains the serotypes 4, 6B, 9V, 14, 18C, 19F, and 23F, and the ten-valent conjugate vaccine additionally containing serotypes 1, 5, and 7F. The eleven-valent and thirteen-valent conjugate vaccines have also been tested and the thirteen-valent vaccine is licensed in the United States [52,71]. As there are multiple pathogenic S. pneumonia serotypes, the development and conjugation of each polysaccharide to carrier proteins remains a complex and challenging process [71].
3.1.3. Haemophilus influenzae
A pure polysaccharide vaccine for Haemophilus influenzae type b (Hib) was first used in the United States in 1985, but due to poor efficacy in children younger than 18 months of age and immunological variability in older children, the vaccine was withdrawn from use in 1988 [77]. The pure polysaccharide Hib vaccine elicited an age-dependent antibody response, where older children and adults developed protective levels of antibodies with long-lasting efficacy [78,79], while younger children generated protective, but lower levels of antibody titers that decreased after one year from initial immunization. In addition, infants, being the most susceptible to Hib infection, were non-responsive to the pure polysaccharide Hib vaccine [77,79]. In clinical trials, a reduction in the nasal carriage with vaccine-specific S. pneumoniae serotypes in children was also observed following immunization with two multivalent pneumococcal conjugate vaccines [37,80]. Furthermore, in older children and adults, even a single dose of a conjugate vaccine leads to highly persistent anti-capsule antibodies [65,66].
In 1989, the pure Hib polysaccharide vaccine was replaced by a polysaccharide–protein conjugate vaccine linking the Hib polysaccharide polyribosylribitol phosphate to diphtheria toxoid, tetanus toxoid, or meningococcal outer membrane protein [52]. Subsequent immunizations with pure polysaccharide Hib or Hib conjugate vaccines do not generate booster responses in adults. However, multiple immunizations with Hib conjugate vaccine elicit booster responses in infants with elevated antibody titers [77,81,82]. The United Kingdom introduced the Hib conjugate vaccine to the infant immunization schedule in 1992 [52]. These vaccines are administered either alone or in combination with other vaccines for the protection of preschool children [52]. As a result, meningitis and other infections caused by Hib have been eliminated in countries that have incorporated the Hib conjugate vaccines into the infant immunization schedules [77,83].
3.1.4. Shigella
The Shigella polysaccharide conjugate vaccine is based on the O-specific polysaccharide (O-SP) domain of LPS. It has been shown that serum IgG to O-SP induced protective immunity to Shigellosis [84]. To increase immunogenicity in young children, O-SP was covalently linked to carrier proteins. In one clinical study, it was shown that conjugation of Shigella sonnei and Shigella flexneri 2a O-SP to succinylated P. aeruginosa exotoxin A or Diptheria toxin mutants was highly immunogenic, and resulted in high titers of serum IgG against O-SP within one week of immunization [84]. Out of the different vaccine candidates, the P. aeruginosa conjugate vaccine resulted in the highest IgG titers and the highest efficacy was observed for S. sonnei conjugate vaccine at 74% [84,85]. Following immunization with S. sonnei conjugate vaccine, IgG and secretory IgA antibody was detected in urine, implying that this vaccine was able to induce mucosal immunity [86]. In addition, a single immunization with S. sonnei conjugated to P. aeruginosa exoprotein A (rEPA) conferred specific protection against shigellosis in young adults [87].
3.1.5. Staphylococcus aureus and Vibrio cholera
S. aureus type 5 and type 8 capsular polysaccharides are of high clinical importance. Immunization with S. aureus capsular polysaccharide conjugate vaccines generated high titers of specific antibodies. These antibodies were able to recognize variations in capsular polysaccharides that occur among clinically significant S. aureus isolates [88]. In addition, the V. cholera O1 serotype Inaba polysaccharide and cholera toxin conjugate vaccine were shown to induce serum anti-LPS IgG with bactericidal activity. V. cholera specific antibodies, whether acquired through natural immunity following colonization by the bacteria, or after immunization, are all directed against the LPS molecules [89]. Furthermore, antibodies generated against the O-specific polysaccharides of V. cholera serotype Inaba were able to cross recognize polysaccharides from V. cholera serotype Ogawa [89].
3.1.6. Typhoidal Salmonella Vaccines
The development of an effective Salmonella vaccine for humans remains a pressing issue. Salmonella can survive both extracellularly and intracellularly within monocytes and macrophages. This is a significant challenge for vaccine development, as both humoral and cell-mediated immunity are needed to efficiently target and clear both extracellular and intracellular bacteria, respectively [90,91]. Antibodies mediate pathogen killing through opsonization, thereby facilitating phagocytosis and preventing the spread of the disease [92]. T cells, on the other hand, are involved in destroying Salmonella residing within phagocytic cells [93,94]. The requirement of both arms of the immune system has been demonstrated through animal studies, where mice lacking T cells were unable to control Salmonella infections [95]. In other studies, passive transfer of specific antibodies provided significant clearance of Salmonella, in vivo [90,96].
The first inactivated whole-cell vaccine against S. Typhi was introduced to British and United States military personnel in 1896 [53]. During the next 100 years, this vaccine dramatically reduced the number of deaths associated with typhoid fever [53,97]. Although the inactivated whole-cell vaccine had an efficacy of 73% over three years post-immunization, it also had high reactogenicity and therefore it was discontinued from use [98,99,100]. In recent years, several clinical trials have tested the efficacy and type of immune response mounted following immunization with inactivated whole-cell Salmonella vaccines and had failed to find a significant cellular response. Additionally, individuals with pre-acquired natural immunity expressed a transient suppression of cellular immunity, questioning the usefulness of whole-cell vaccine administration in endemic regions [101,102].
Currently, there are three vaccine types against S. Typhi: (1) live attenuated, (2) pure Vi polysaccharide capsule-based, and (3) Vi-protein conjugates (Table 2). Non-specific chemical mutagenesis of the S. Typhi Ty21 strain was used to generate the live-attenuated vaccine Ty21. Ty21 does not express Vi capsule and as a result, the immune response to live-attenuated Ty21 strain is not directed against the Vi capsule [103]. The Ty21 vaccine requires three doses to induce the best protective immunity and in combination, the three doses have an efficacy of 51% [98,104]. This vaccine is not thermostable over long periods and requires a cold chain. This is a setback as the vaccine is most needed in developing countries where access to refrigeration is difficult [53]. It has been shown that freeze-drying can increase the thermal stability of the vaccine, but this measure is not employed widely [105]. Despite the Ty21vaccine being licensed for use with adults and children over five years of age, seroconversion was greatly reduced in young children compared to adults [106]. Nonetheless, the Ty21 vaccine can induce T-cell immunity and has shown cross-protection against S. Paratyphi B in clinical studies. In addition, in vitro cross-protection has been shown for S. Paratyphi A and B [107,108]. As the antibody response is mostly directed against the O:9 O-Antigen epitope of S. Typhi, it was shown that the Ty21 vaccine can elicit cross-protective immunity against invasive non-typhoidal Salmonella (iNTS) and NTS serovars also expressing the O:9 or O:9,12 epitopes [53,109,110].
The pure Vi polysaccharide capsule-based vaccine generates primarily an antibody response directed towards the Vi capsule. However, recent evidence indicates that the immune response could also be due to other Salmonella components in the vaccine [110]. Unfortunately, the vaccine is considered to be non-immunogenic in infants, due to the Vi capsule being a T-independent antigen and the serum antibody response being mostly IgM driven. Therefore, the pure Vi vaccine is only licensed for children over 2 years of age [53]. In contrast to the Ty21 vaccine, the Vi capsule-based vaccine requires a single dose and has an efficacy of 55% over three years. In one study only 2 out of 32 subjects generated a sufficient amount of IgG towards the Vi polysaccharide capsule-based vaccine [111]. However, in typhoid fever endemic regions, immunization with this vaccine resulted in 55% and 75% efficacy rates among adults and children over the age of 5 years, respectively [37].
The Vi polysaccharide capsule-based vaccine also requires a cold chain to prolong stability and effectiveness [53]. Additionally, vaccination with the Vi capsule-based vaccine leads to the production of circulating plasmablasts (immature plasma cells) expressing systemic homing receptors (L selectin), while vaccination with the Ty21 vaccine leads to the expression of mucosal homing receptors (α4β7), as with natural infection [110]. Thus, vaccination with the Vi capsule will not direct immune cells specifically to the intestinal mucosal region, rather they will be distributed systemically, and consequently, the immune cells will not have a targeted response at the intestinal site of infection. No clinical trials have been performed to date to examine the efficacy and protection of these two types of vaccines combined [53].
In recent years, several different Vi protein conjugate vaccines have been licensed, especially in Asia, to counter the low immune response mounted to the pure polysaccharide vaccine (Table 2). The importance of antibodies in clearing S. Typhi infections was demonstrated by efficacy data for Vi capsule-based vaccine and Vi protein conjugate vaccine candidate phase III studies [98,104,112,113]. A phase III study of a glycoconjugate vaccine combining S. Typhi Vi capsule with P. aeruginosa exoprotein A (Vi-rEPA), conducted over a decade ago in Vietnamese children between two and five years of age, found a 91% efficacy after 27 months and 89% efficacy after 46 months from primary immunization [53]. Thus, protein conjugation holds promise in developing an effective vaccine against S. Typhi infections.
Although several vaccines against S. Typhi are being developed (Table 2), no new vaccine against S. Typhi has been introduced to the market in recent times. The delay in seeing new vaccines in the market is mainly due to a lack of commercial interest in developing vaccines against typhoid fever and iNTS disease since these are largely diseases of low-income countries, which have little money to invest in vaccines [114]. New vaccines are expected to provide improved protection and efficacy, especially in young children, and to reduce the cost of delivery; these vaccines are to be administered as part of the national Expanded Programs on Immunization [53]. Vaccines for S. Typhi and/or S. Paratyphi are to be given at nine months, as disease peaks at two years of age, and vaccines for iNTS are to be given between two and four months of age, prior to the disease peak at one year of age [115,116,117].
4. Non-Typhoidal Salmonella Vaccines
Although a humoral response is sufficient to clear most S. Typhi infections, both antibody and cell-mediated immunity are required for proper clearance of NTS. This has been shown in field studies in sub-Saharan Africa, where the age-related prevalence of iNTS disease in children declined with the acquisition of specific antibodies [116]. In addition, serum antibodies have shown in vitro bactericidal activity and oxidative killing of iNTS [116,118]. It has also been demonstrated that O-Antigen-based conjugate vaccine candidates induce sufficient protection against otherwise lethal doses of Salmonella in mice [119,120]. Passive transfer of monoclonal antibodies specific for O-Antigen was also able to confer protection against lethal doses of Salmonella challenge [121,122]. Immunization of animals with a heat-killed invasive African S. Typhimurium sequence type 313 strain led to the majority of antibodies mounted against the O-Antigen [123]. On the other hand, the importance of cell-mediated immunity in clearing NTS is apparent through the association between HIV infection and predisposition to NTS, especially when the CD4+ count is below 200 cells/uL [124]. HIV has been shown to increase susceptibility to NTS through dysregulation of cytokine response, humoral immunity, and disruption of the integrity of the gastrointestinal mucosa [94,125]. Therefore, vaccine candidates that induce protective mucosal immunity, preventing gastroenteritis and NTS invasion from the gastrointestinal tract, would be highly beneficial for HIV-infected individuals [53].
4.1. Glycoconjugate Vaccines for NTS
There are several different types of NTS vaccine candidates currently being tested, which fall into three main categories: glycoconjugate, live attenuated, and subunit vaccines (Table 2). Glycoconjugate vaccines covalently link a poorly immunogenic antigen, such as a bacterial surface polysaccharide, to a protein carrier molecule to activate CD4+ T cells. For glycoconjugation, most Salmonella vaccine strategies use the common carrier proteins tetanus toxoid, diphtheria toxoid, nontoxic recombinant diphtheria toxoid, or P. aeruginosa exoprotein A (rEPA) [53]. In one study, S. Typhimurium O-Antigens 4 and 12 covalently linked to bovine serum albumin (BSA) were administered with Freund’s adjuvant, leading to the production of high O4 specific antibody titers in rabbits, comparable to those elicited by immunization with heat-killed bacteria [126].
It was proposed that glycoconjugate vaccines using Salmonella carbohydrates linked to Salmonella proteins could be more effective at inducing Salmonella-specific antibodies since antibodies can be produced against both Salmonella proteins and carbohydrates [127,128]. In addition, conjugation to Salmonella proteins could lead to the generation of a Salmonella-specific T-cell response. Conjugation of S. Typhimurium O:4 polysaccharide to outer membrane (OM) porins led to better protection of mice upon challenge, as compared to using porins alone or O:4 conjugated diphtheria toxoid [128]. As flagellin is the only cell-surface protein antigen typically used for typing Salmonella, it was investigated for the ability to generate protective immunity. Immunization of mice with flagellin alone, or in conjugation with the O-Antigen leads to the induction of protection [119,129,130]. For example, immunization with O:4,5 and O:9 conjugated to Salmonella flagellin in iNTS vaccine candidates led to enhanced protective immunity, as compared to either antigen alone [119,131]. Flagellin is the primary ligand for TLR5 activation and can signal to the innate immune system and lead to immunomodulatory effects in mice [130,132]. One concern is that in some Salmonella serovars, such as S. Typhimurium, flagellin can have phase variable expression and is not constitutively expressed during infection [53]. Another concern is due to the expression of multiple types of flagellin, immunization against one flagellin may not provide cross-protection against infection with a strain expressing a different type of flagellin.
4.2. Live-Attenuated Vaccines for NTS
Live-attenuated vaccines can induce both humoral and cellular immunity, and thereby facilitate Salmonella-specific T-cell responses. These vaccines are the easiest to deliver (orally) and can induce strong mucosal immune responses [110]. In addition, the availability of numerous Salmonella antigens on live bacteria can yield broad protective immune responses and even allow cross recognition between Salmonella serovars. The primary challenge with developing live-attenuated vaccines is in balancing the level of immunogenicity with a proper and sufficient level of attenuation [53]. Currently, several live-attenuated S. Typhimurium strains are being tested for their efficacy in preclinical and Phase I clinical studies (Table 2). Most of these strains contain mutations or deletions in genes coding for secretion systems and virulence.
Additionally, recombinant attenuated Salmonella vaccines (RASV) have been used as vectors to deliver vaccine antigens for various other bacterial pathogens. RASV expressing pneumococcal surface protein A (PspA) elicited an IgG response with a predominant rPspA specific IgG1 (Th2-type) and Salmonella LPS and OMP specific IgG2a (Th1-type) responses [133]. Similarly, oral immunization of mice with RASV vectors expressing genes from seven different serotypes of CPS from S. pneumoniae elicited a Th1-type immune response and subsequent in vitro clearance of the S. pneumoniae expressing the corresponding CPS serotype [134]. Immunization with RASV vectors expressing Salmonella and M. tuberculosis effector protein SopE resulted in similar observations [135]. Furthermore, RASVs with regulated delayed attenuation have seen significantly higher anti-PspA responses (i.e., IgG, IL-4, and INF-γ), compared to the controls [136]. These vaccines express wild-type phenotypes at immunization and are attenuated after colonization of the host tissues, by inactivating the virulence genes [136]. Thus, RASVs not only facilitate the induction of an immune response against the vaccine antigen from other bacteria but also induce protective responses against Salmonella-specific antigens [137,138,139].
4.3. Subunit Vaccines
Protein-based subunit vaccines are composed of multiple antigenic epitopes and can have broad coverage. A subunit approach is currently in use for multivalent pneumococcal and meningococcal conjugate vaccines. In another strategy to develop broad coverage, cross-protective vaccines can use highly conserved protein antigens produced through recombinant technology [140,141,142]. Proper antigens can be selected using bioinformatics analysis of whole-genome sequences and reverse vaccinology [143]. Examples of highly conserved Salmonella protein antigens include flagellin and porins (i.e., OmpC, F, and D). Both recombinant and purified protein vaccine candidates have been tested against conserved protein epitopes [2]. Subunit vaccines can induce both T-cell responses and antibody production; a balanced Th1-Th2 approach is vital to proper clearance of Salmonella. One of the downsides of subunit vaccines developed through recombinant technology is that it is difficult to maintain and preserve the proper conformation of proteins, especially when there are several membrane-spanning domains. This can result in the induction of a poor antibody response. One approach is to purify proteins from whole Salmonella cells rather than relying on recombinant proteins [142].
4.4. Other Vaccines Currently Being Developed for NTS
As glycoconjugate vaccines primarily rely on the production of antibodies specific for surface carbohydrate moieties, it may not be sufficient to efficiently deal with the growing iNTS disease. The development of new live-attenuated, protein-based, or Generalized Modules for Membrane Antigens (GMMA)-based vaccines will serve to provide broader protection by activating Salmonella-specific T-cell responses [53,144,145,146]. Vaccine development has been hindered due to a lack of understanding regarding specific antigenic epitopes of Salmonella and preservation of proper protein confirmations. GMMA technology (i.e., outer membrane blebs) is used to deliver outer membrane proteins and surface polysaccharides in the correct orientation and conformation to induce protective immunity. In addition, the membrane components of GMMAs have adjuvant activity by co-delivering multiple pathogen-associated molecular patterns (PAMPs) along with the target antigens [2]. In our hands, immunization of mice with GMMAs isolated from colanic acid overproducing strains was sufficient to significantly reduce Salmonella colonization [47]. It is possible that the development of a GMMA-CPS-based vaccine against NTS can be explored in the future.
5. Looking Ahead
The primary goal of this review was to provide a perspective of the current state of NTS vaccine development and to encourage vaccine candidate testing and clinical development. Despite the type of vaccine developed, in terms of delivery, time and the associated cost of administration, it will be advantageous to develop vaccines that require only one or two doses to mount an optimal immune response, compared to the required three doses of the current Ty21 vaccine [53]. Although the current vaccine pipeline is looking weak for NTS, there are candidates not listed in official databases, therefore the prospects are stronger than they appear. In our opinion, there needs to be a greater commitment of scientific expertise and availability of resources to accelerate vaccine candidates, especially taking S. Typhi as a proof of principle for vaccine development. Less expensive conjugate methods and technological advancements should be considered to facilitate vaccine accessibility in low-income countries. Subunit vaccines, especially those using glycoconjugate technologies, are costly to produce and since most vaccines are to be used in low-income countries, affordability remains a key factor in vaccine development. To balance the cost-effectiveness, it has been proposed that multivalent vaccines composed of 5 to 6 antigens could provide cross-protection against the majority of the iNTS serovars [127]. Furthermore, a better understanding of the regional burden of NTS/iNTS infections will enhance informed policy decisions.
An NTS vaccine would be both beneficial as a vaccine for travelers in developed countries, and as an early childhood vaccine in endemic regions. Although an NTS vaccine would require no new implementation provided the vaccine is introduced to childhood vaccination schedules, vaccination in high-disease-burden regions may require additional infrastructure for storage. One of the biggest hurdles is that commercial attractiveness for NTS vaccines remains low, given the demand remains relatively low in middle- and high-income countries. Thus, it should be noted that the limited candidates in the pipeline are a reflection of a lack of interest and resources to drive clinical testing, rather than a reflection of the feasibility of developing an NTS vaccine.
Author Contributions
Conceptualization, S.R.P. and A.P.W.; writing—original draft preparation, S.R.P.; writing—review and editing, S.R.P. and A.S.S.; funding acquisition, A.P.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a New Investigator Establishment Grant from the Saskatchewan Health Research Foundation (Grant #2588 to A.P.W.), Natural Sciences and Engineering Research Council (NSERC) Discovery grant (Grant #2017-05737 to A.P.W.), the Jarislowsky Chair in Biotechnology (A.P.W.), and the University of Saskatchewan (Graduate research fellowships to S.R.P. and A.S.S.) The funders had no role in the decision to publish or preparation of the manuscript. VIDO receives operational funding from the Government of Saskatchewan through Innovation Saskatchewan and the Ministry of Agriculture. Published as VIDO manuscript #946.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella-host interactions—Modulation of the host innate immune system. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Product Development for Vaccines Advisory Committee (PD-VAC) Meeting. 2015. Available online: http://www.who.int/immunization/research/meetings_workshops/pdvac/en/ (accessed on April 2021).
- Pathogen Safety Data Sheet: Infectious Substances—Salmonella Enterica Spp. Available online: http://www.phac-aspc.gc.ca/lab-bio/res/psds-ftss/salmonella-ent-eng.php (accessed on April 2021).
- FoodNet Canada Annual Report 2016. Available online: https://www.canada.ca/en/public-health/services/surveillance/foodnet-canada/publications/foodnet-canada-annual-report-2016.html (accessed on April 2021).
- De Jong, H.K.; Parry, C.M.; van der Poll, T.; Wiersinga, W.J. Host-pathogen interaction in invasive salmonellosis. PLoS Pathog. 2012, 8, e1002933. [Google Scholar] [CrossRef] [Green Version]
- Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on April 2021).
- Salmonella Outbreak Investigations: Timeline for Reporting Cases. Available online: http://www.cdc.gov/salmonella/reportingtimeline.html (accessed on April 2021).
- Antibiotic Resistance Threats in the United States. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwjiqq6U0sDwAhVH_54KHbTICiUQFjAOegQIEhAD&url=https%3A%2F%2Fwww.cdc.gov%2Fdrugresistance%2Fpdf%2Fthreats-report%2F2019-ar-threats-report-508.pdf&usg=AOvVaw1709sEktvf-ovR38riUreg (accessed on April 2021).
- Brisabois, A.; Cazin, I.; Breuil, J.; Collatz, E. Surveillance of antibiotic resistance in Salmonella. Eurosurveillance 1997, 2, 181. [Google Scholar] [CrossRef] [PubMed]
- Salmonella Heidelberg Ceftiofur-Related Resistance in Human and Retail Chicken Isolates. Available online: http://www.phac-aspc.gc.ca/cipars-picra/heidelberg/heidelberg-eng.php (accessed on April 2021).
- Upreti, R.K.; Kumar, M.; Shankar, V. Bacterial glycoproteins: Functions, biosynthesis and applications. Proteomics 2003, 3, 363–379. [Google Scholar] [CrossRef]
- Roberts, I.S. The biochemistry and genetics of capsular polysaccharide production in Bacteria. Annu. Rev. Microbiol. 1996, 50, 285–315. [Google Scholar] [CrossRef]
- Troy, F.A.; Frerman, F.E.; Heath, E.C. The biosynthesis of capsular polysaccharide in aerobacter aerogenes. J. Biol. Chem. 1971, 246, 118–133. [Google Scholar] [CrossRef]
- De Rezende, C.E.; Anriany, Y.; Carr, L.E.; Joseph, S.W.; Weiner, R.M. Capsular polysaccharide surrounds smooth and rugose types of Salmonella enterica serovar Typhimurium DT104. Appl. Environ. Microb. 2005, 71, 7345–7351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costerton, J.W.; Irvin, R.T.; Cheng, K.-J. The bacterial glycocalyx in nature and disease. Annu. Rev. Microbiol. 1981, 35, 299–324. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Kasper, D.L. The love-hate relationship between bacterial polysaccharides and the host immune system. Nat. Rev. Immunol. 2006, 6, 849–858. [Google Scholar] [CrossRef]
- Owens, T.W.; Taylor, R.J.; Pahil, K.S.; Bertani, B.R.; Ruiz, N.; Kruse, A.C.; Kahne, D. Structrual basis of unidirectional export of lipopolysaccharide to the cell surface. Nature 2019, 567, 550–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Orlando, B.J.; Liao, M. Structural basis of lipopolysaccharide extraction by the LptB2FGC complex. Nature 2019, 567, 486–490. [Google Scholar] [CrossRef]
- Whitfield, C. Biosynthesis and assembly of capsular polysaccharides in escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef]
- Whitfield, C.; Amor, P.A.; Koplin, R. Modulation of the surface architecture of gram-negative bacteria by the action of surface polymer: Lipid a-core ligase and by determinants of polymer chain length. Mol. Microbiol. 1997, 23, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Sukupolvi-Petty, S.; Grass, S.; StGeme, J.W., III. The haemophilus influenzae type B hcsA and hcsB gene products facilitate transport of capsular polysaccharide across the outer membrane and are essential for virulence. J. Bacteriol. 2006, 188, 3870–3877. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.S.; Brotcke, A.; Henry, T.; Margolis, J.J.; Chan, K.; Monack, D.M. In vivo negative selection screen identifies genes required for francisella virulence. Proc. Natl. Acad. Sci. USA 2007, 104, 6037–6042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.L.; White, A.P.; Snyder, S.D.; Martin, S.; Heiss, C.; Azadi, P.; Surette, M.; Kay, W.W. Salmonella produces an o-antigen capsule regulated by agfd and important for environmental persistence. J. Bacteriol. 2006, 188, 7722–7730. [Google Scholar] [CrossRef] [Green Version]
- DeShazer, D.; Waag, D.M.; Fritz, D.; Woods, D.E. Identification of a Burkholderia mallei polysaccharide gene cluster by subtractive hybridization and demonstration that the encoded capsule is an essential virulence determinant. Microb. Pathog. 2001, 30, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Laxalt, K.A.; Kozel, T.R. Chemotaxigenesis and activation of the alternative complement pathway by encapsulated and non-encapsulated cryptococcus neoformans. Infect. Immun. 1979, 26, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Karlyshev, A.V.; Wren, B.W. Detection and initial characterization of novel capsular polysaccharide among diverse campylobacter jejuni strains using alcian blue dye. J. Clin. Microbiol. 2001, 39, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.L.; Roche, A.M.; Gould, J.M.; Chim, K.; Ratner, A.J.; Weiser, J.N. Capsule enhances pneumococcal colonization by limiting mucus-mediated clearance. Infect. Immun. 2007, 75, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, J.-M.; Vann, W.F.; Karakawa, W.W. Purification and characterization of staphylococcus aureus type 8 capsular polysaccharide. Infect. Immun. 1984, 45, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Orskov, F.; Orskov, I. Escherichia coli serotyping and disease in man and animals. Can. J. Microbiol. 1992, 38, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Vann, W.F.; Soderstrom, T.; Egan, W.; Tsui, F.-P.; Schneerson, R.; Orskov, I.; Orskov, F. Serological, chemical, and structural analyses of the escherichia coli cross-reactive capsular polysaccharides K13, K20, and K23. Infect. Immun. 1983, 39, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Goldman, R.C.; White, D.; Orskov, F.; Orskov, I.; Rick, P.D.; Lewis, M.S.; Bhattacharjee, A.K.; Leive, L. A surface polysaccharide of Escherichia coli O111 contains o-antigen and inhibits agglutination of cells by O-Antiserum. J. Bacteriol. 1982, 151, 1210–1221. [Google Scholar] [CrossRef] [Green Version]
- Amor, P.A.; Whitfield, C. Molecular and functional analysis of genes required for expression of group 1B K antigens in Escherichia coli: Characterization of the his-region containing gene clusters for multiple cell-surface polysaccharides. Mol. Microbiol. 1997, 26, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Roberts, I.S. Structure, assembly and regulation of expression of capsules in Escherichia coli. Mol. Microbiol. 1999, 31, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Ophir, T.; Gutnick, D.L. A role for exopolysaccharides in the protection of microorganisms from desiccation. Appl. Environ. Microb. 1994, 60, 740–745. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Cheng, K.-J.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial biofilms in nature and disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef]
- Ledeboer, N.A.; Jones, B.D. Exopolysaccharide sugars contribute to biofilm formation by Salmonella enterica serovar typhimurium on Hep-2 cells and chicken intestinal epithelium. J. Bacteriol. 2005, 187, 3214–3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, R.K.; Fattom, A. Capsular polysaccharide-protein conjugate vaccines and intravenous immunoglobulins. Expert Opin. Investig. Drugs. 1998, 7, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Pickard, D.; Li, J.; Roberts, M.; Maskell, D.; Hone, D.; Levine, M.; Dougan, G.; Chatfield, S. Characterization of defined ompR mutants of salmonella typhi: OmpR is involved in the regulation of vi polysaccharide expression. Infect. Immun. 1994, 62, 3984–3993. [Google Scholar] [CrossRef] [Green Version]
- Berry, A.; DeVault, J.D.; Chakrabarty, A.M. High osmolarity is a signal for enhanced algD transcription in mucoid and nonmucoid pseudomonas aeruginosa strains. J. Bacteriol. 1989, 171, 2312–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grados, O.; Ewing, W.H. Antigenic relationship between escherichia coli and neisseria meningitidis. J. Infect. Dis. 1970, 122, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Liston, S.D.; Ovchinnikova, O.G.; Whitfield, C. Unique lipid anchor attaches vi antigen capsule to the surface of Salmonella enterica serovar Typhi. Proc. Natl. Acad. Sci. USA 2016, 113, 6719–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wangdi, T.; Lee, C.-Y.; Spees, A.M.; Yu, C.; Kingsbury, D.D.; Winter, S.E.; Hastey, C.J.; Wilson, R.P.; Heinrich, V.; Baumler, A.J. The vi capsular polysaccharide enables salmonella enterica serovar typhi to evade microbe-guided neutrophil chemotaxis. PLoS Pathog. 2014, 10, e1004306. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.P.; Winter, S.E.; Spees, A.M.; Winter, M.G.; Nishimori, J.H.; Sanchez, J.F.; Nuccio, S.-P.; Crawford, R.W.; Tukel, C.; Baumler, A.J. The vi capsular polysaccharide prevents complement receptor 3-mediated clearance of Salmonella enterica serotype typhi. Infect. Immun. 2011, 79, 830–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Tukel, C.; Akcelik, M.; Baumler, A.J. Capsule-mediated immune evasion: A new hypothesis explaining aspects of typhoid fever pathogenesis. Infect. Immun. 2006, 74, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Hirose, K.; Ezaki, T.; Miyake, M.; Li, T.; Khan, A.Q.; Kawamura, Y.; Yokoyama, H.; Takami, T. Survival of vi-capsulated and vi-deleted salmonella typhi strains in cultured macrophage expressing different levels of CD14 antigen. FEMS Microbiol. Lett. 1997, 147, 259–265. [Google Scholar] [CrossRef]
- Quintero, E.; Weiner, R.M. Evidence for the adhesive function of the exopolysaccharide of hyphomonas strain MHS-3 in its attachment to surfaces. Appl. Environ. Microb. 1995, 61, 1897–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokaribo, A.S.; Perera, S.R.; Sereggela, Z.; Krochak, R.; Balezantis, L.R.; Xing, X.; Lam, S.; Deck, W.; Attah-Poku, S.; Abbott, D.W.; et al. A GMMA-CPS-based vaccine for non-typhoidal salmonella. Vaccines 2021, 9, 165. [Google Scholar] [CrossRef]
- Kasper, D.L.; Onderdonk, A.B.; Crabb, J.; Bartlett, J.G. Protective efficacy of immunization with capsular antigen against experimental infection with bacteroides fragilis. J. Infect. Dis. 1979, 140, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Kenzel, S.; Henneke, P. The innate immune system and its relevance to neonatal sepsis. Curr. Opin. Infect. Dis. 2006, 19, 264–270. [Google Scholar] [CrossRef]
- Wilder-Smith, A. Meningococcal vaccines: A neglected topic in travel medicine? Expert Rev. Vaccines 2009, 8, 1343–1350. [Google Scholar] [CrossRef]
- State Exemption Levels Low, National Vaccination Rates High. Available online: https://www.cdc.gov/media/releases/2015/p0827-vaccination-rates.html (accessed on April 2021).
- Pollard, A.J.; Perrett, K.P.; Beverley, P.C. Maintaining protection against invasive bacteria with protein-polysaccharide conjugate vaccines. Nat. Rev. Immunol. 2009, 9, 213–220. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive salmonella disease. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesinski, G.B.; Westerink, M.A.J. Novel vaccine strategies to t-independent antigens. J. Microbiol. Methods. 2001, 47, 135–149. [Google Scholar] [CrossRef]
- Blanchard-Rohner, G.; Pollard, A. Long-term protection after immunization with protein-polysaccharide conjugate vaccines in infancy. Expert Rev. Vaccines 2011, 10, 673–684. [Google Scholar] [CrossRef]
- Kruetzmann, S.; Rosado, M.M.; Weber, H.; Germing, U.; Tournilhac, O.; Peter, H.-H.; Berner, R.; Peters, A.; Boehm, T.; Plebani, A.; et al. Human immunoglobulin M memory B cells controlling streptococcus pneumoniae infections are generated in the spleen. J. Exp. Med. 2003, 197, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Kruschinski, C.; Zidan, M.; Debertin, A.S.; Von Horsten, S.; Pabst, R. Age-dependent development of the splenic marginal zone in human infants is associated with different causes of death. Hum. Pathol. 2004, 35, 113–121. [Google Scholar] [CrossRef]
- Jokhdar, H.; Borrow, R.; Sultan, A.; Adi, M.; Riley, C.; Fuller, E.; Baxter, D. Immunologic hyporesponsiveness to serogroup c but not serogroup a following repeated meningococcal a/c polysaccharide vaccination in Saudi Arabia. Clin. Diagn. Lab. Immun. 2004, 11, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Kalka-Moll, W.M.; Tzianabos, A.O.; Bryant, P.W.; Niemeyer, M.; Ploegh, H.L.; Kasper, D.L. Zwitterionic polysaccharides stimulate T cells by MHC class II-dependent interactions. J. Immunol. 2002, 169, 6149–6153. [Google Scholar] [CrossRef] [Green Version]
- Makela, P.H.; Kayhty, H. Evolution of conjugate vaccines. Expert Rev. Vaccines 2002, 1, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, M.E.; McVernon, J.; Andrews, N.J.; Heath, P.T.; Slack, M.P. Estimating haemophilus influenzae type b vaccine effectiveness in England and wales by use of the screening method. J. Infect. Dis. 2003, 188, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, P.T.; McVernon, J. The UK hib vaccine experience. Arch. Dis. Child. 2002, 86, 396–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, M.E.; Andrews, N.J.; Trotter, C.L.; Kaczmarski, E.B.; Miller, E. Herd immunity form meningococcal serogroup C conjugate vaccination in England: Database analysis. BMJ 2003, 326, 365–366. [Google Scholar] [CrossRef] [Green Version]
- Borrow, R.; Andrews, N.; Findlow, H.; Waight, P.; Southern, J.; Crowley-Luke, A.; Stapley, L.; England, A.; Findlow, J.; Miller, E. Kinetics of antibody persistence following administration of a combination meningococcal serogroup C and haemophilus influenzae type b conjugate vaccine in healthy infants in the United Kingdom primed with a monovalent meningococcal serogroup C vaccine. Clin. Vaccine Immunol. 2010, 17, 154–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snape, M.D.; Kelly, D.F.; Salt, P.; Green, S.; Snowden, C.; Diggle, L.; Borkowski, A.; Yu, L.-M.; Moxon, R.; Pollard, A.J. Serogroup C meningococcal glycoconjugate vaccine in adolescents: Persistence of bactericidal antibodies and kinetics of the immune reponse to a booster vaccine more than 3 years after immunization. Clin. Infect. Dis. 2006, 43, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Snape, M.D.; Kelley, D.F.; Lewis, S.; Banner, C.; Kibwana, L.; Moore, C.E.; Diggle, L.; John, T.; Yu, L.-M.; Borrow, R.; et al. Seroprotection against serogroup C meningococcal disease in adolescents in the United Kingdom: Observational study. BMJ 2008, 336, 1487–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auckland, C.; Gray, S.; Borrow, R.; Andrews, N.; Goldblatt, D.; Ramsay, M.; Miller, E. Clinical and immunologic risk factors for meningococcal c conjugate vaccine failure in the United Kingdom. J. Infect. Dis. 2006, 194, 1745–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McVernon, J.; Johnson, P.D.R.; Pollard, A.J.; Slack, M.P.E.; Moxon, E.R. Immunologic memory in haemophilus influenzae type b conjugate vaccine failure. Arch. Dis. Child. 2003, 88, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Blanchard-Rohner, G.; Snape, M.D.; Kelly, D.F.; John, T.; Morant, A.; Yu, L.-M.; Borkowski, A.; Ceddia, F.; Borrow, R.; Siegrist, C.-A.; et al. The magnitude of the antibody and memory b cell responses during priming with a protein-polysaccharide conjugate vaccine in human infants is associated with the persistence of antibody and the intensity of booster response. J. Immunol. 2008, 180, 2165–2173. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines, vaccination, and vaccinology. J. Infect. Dis. 2003, 187, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Nair, M. Protein conjugate polysaccharide vaccines: Challenges in development and global implementation. Indian J. Community Med. 2012, 37, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Richmond, P.; Kaczmarski, E.; Borrow, R.; Findlow, J.; Clark, S.; McCann, R.; Hill, J.; Barker, M.; Miller, E. Meningococcal C polysaccharide vaccine induces immunologic hyporesponsiveness in adults that is overcome by meningococcal C conjugate vaccine. J. Infect. Dis. 2000, 181, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Snape, M.D.; Pollard, A.J. Meningococcal polysaccharide-protein conjugate vaccines. Lancet Infect. Dis. 2005, 5, 21–30. [Google Scholar] [CrossRef]
- Trotter, C.L.; Andrews, N.J.; Kaczmarski, E.B.; Miller, E.; Ramsay, M.E. Effectiveness of meningococcal serogroup C conjugate vaccine 4 years after introduction. Lancet 2004, 364, 365–367. [Google Scholar] [CrossRef]
- Shapiro, E.D.; Berg, A.T.; Austrian, R.; Schroeder, D.; Parcells, V.; Margolis, A.; Adair, R.K.; Clemens, J.D. the protective efficacy of polyvalent pneumococcal polysaccharide vaccine. N. Engl. J. Med. 1991, 325, 1453–1460. [Google Scholar] [CrossRef]
- Halloran, M.E.; Haber, M.; Longini, I.M., Jr.; Struchiner, C.J. Direct and indirect effects in vaccine efficacy and effectiveness. Am. J. Epidemiol. 1991, 133, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.B.; Chu, C.; Schneerson, R. Hypothesis for vaccine development: Protective immunity to enteric diseases caused by nontyphoidal salmonella and shigellae may be conferred by serum igG antibodies to the O-specific polysaccharide of their lipopolysaccharides. Clin. Infect. Dis. 1992, 15, 346–361. [Google Scholar] [CrossRef]
- Rodrigues, L.P.; Schneerson, R.; Robbins, J.B. Immunity to hemophilus influenzae type b. i. The isolation, and some physicochemical, serologic and biologic properties of the capsular polysaccharide of hemophilus influenzae type b. J. Immunol. 1971, 107, 1071–1080. [Google Scholar] [PubMed]
- Smith, D.H.; Peter, G.; Ingram, D.L.; Harding, A.L.; Anderson, P. Responses of children immunized with the capsular polysaccharide of hemophilus influenzae type B. Pediatrics 1973, 52, 637–644. [Google Scholar]
- Dagan, R.; Melamed, R.; Muallem, M.; Piglansky, L.; Greenberg, D.; Abramson, O.; Mendelman, P.M.; Bohidar, N.; Yagupsky, P. Reduction of nasopharyngeal carriage of pneumococci during the second year of life by a heptavalent conjugate pneumococcal vaccine. J. Infect. Dis. 1996, 174, 1271–1278. [Google Scholar] [CrossRef]
- Parke, J.C.; Schneerson, R.; Relmer, C.; Black, C.; Welfare, S.; Bryla, D.; Levi, L.; Pavliakova, D.; Cramton, T.; Schulz, D.; et al. Clinical and immunologic reponses to haemophilus influenzae type b-tetanus toxoid conjugate vaccine in infants injected at 3,5,7 and 18 months of age. J. Pediatr. 1991, 118, 184–190. [Google Scholar] [CrossRef]
- Claesson, B.A.; Trolfors, B.; Lagergard, T.; Taranger, J.; Bryla, D.; Otterman, G.; Cramton, T.; Yang, Y.; Reimer, C.B.; Robbins, J.B.; et al. Clinical and immunologic responses to capsular polysaccharide of haemophilus influenzae type b alone or conjugated to tetanus toxoid in 18- to 23-month-old children. J. Pediatr. 1988, 112, 695–702. [Google Scholar] [CrossRef]
- Adams, W.G.; Deaver, K.A.; Cochi, S.L.; Plikaytis, B.D.; Zell, E.R.; Broome, C.V.; Wenger, J.D. Decline of childhood haemophilus influenzae type b (Hib) disease in the Hib vaccine era. JAMA—J. Am. Med. Assoc. 1993, 269, 221–226. [Google Scholar] [CrossRef]
- Passwell, J.H.; Harlev, E.; Ashkenazi, S.; Chu, C.; Miron, D.; Ramon, R.; Farzan, N.; Shiloach, J.; Bryla, D.A.; Majadly, F.; et al. Safety and immunogenicity of improved shigella o-specific polysaccharide-protein conjugate vaccines in adults in Israel. Infect. Immun. 2001, 69, 1351–1357. [Google Scholar] [CrossRef] [Green Version]
- Ferreccio, C.; Prado, V.; Ojeda, A.; Cayyazo, M.; Abrego, P.; Guers, L.; Levine, M.M. Epidemiologic patterns of acute diarrhea and endemic shigella infections in children in a poor periurban setting in Santiago, Chile. Am. J. Epidemiol. 1991, 134, 614–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.; Orr, N.; Robin, G.; Slepon, R.; Ashkenazi, S.; Ashkenazi, I.; Shemer, J. Detection of antibodies to shigella lipopolysaccharide in urine after natural shigella infection or vaccination. Clin. Diagn. Lab. Immun. 1996, 3, 451–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.; Ashkenazi, S.; Green, M.S.; Gdalevich, M.; Robin, G.; Slepon, R.; Yavzori, M.; Orr, N.; Block, C.; Asshkenazi, I.; et al. Double-blinded vaccine-controlled randomised efficacy trial of an investigational shigella sonnei conjugate vaccine in young adults. Lancet 1997, 349, 155–159. [Google Scholar] [CrossRef]
- Fattom, A.I.; Sarwar, J.; Basham, L.; Ennifar, S.; Naso, R. Antigenic determinants of staphylococcus aureus type 5 and type 8 capsular polysaccharide vaccines. Infect. Immun. 1998, 66, 4588–4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K.; Taylor, D.N.; Bryla, D.A.; Robbins, J.B.; Szu, S.C. Phase 1 evaluation of vibrio cholera O1, Serotype Inaba, polysaccharide-cholera toxin conjugates in adult volunteers. Infect. Immun. 1998, 66, 3095–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastroeni, P.; Villarreal-Ramos, B.; Hormaeche, C.E. Adoptive transfer of immunity to oral challenge with virulent salmonellae in innately susceptible BALB/c mice requires both immune serum and T cells. Infect. Immun. 1993, 61, 3981–3984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salerno-Goncalves, R.; Pasetti, M.F.; Sztein, M.B. Characterization of CD8+ effector T cell responses in volunteers immunized with salmonella enterica serovar typhi strain Ty21a typhoid vaccine. J. Immunol. 2002, 169, 2196–2203. [Google Scholar] [CrossRef] [Green Version]
- Mastroeni, P.; Grant, A.; Restif, O.; Maskell, D. A dynamic view of the spread and intracellular distribution of Salmonella enterica. Nat. Rev. Microbiol. 2009, 7, 73–80. [Google Scholar] [CrossRef]
- Blanden, R.V.; Mackaness, G.B.; Collins, F.M. Mechanisms of acquired resistance in mouse typhoid. J. Exp. Med. 1966, 124, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Mackaness, G.B.; Blanden, R.V.; Collins, F.M. Host-parasite relations in mouse typhoid. J. Exp. Med. 1966, 124, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Mastroeni, P.; Harrison, J.; De Hormaeche, R.D.; Hormaeche, C.E. Salmonella typhimurium aroA, htrA, and aroD htrA mutants cause progressive infections in athymic (nu/nu) BALB/c mice. Infect. Immun. 1997, 65, 1566–1569. [Google Scholar] [CrossRef] [Green Version]
- McSorley, S.J.; Jenkins, M.K. Antibody is required for protection against virulent but not attenuated salmonella enterica serovar typhimurium. Infect. Immun. 2000, 68, 3344–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, P.R.; Simmons, J.S. The effectiveness of vaccines used for the prevention of typhoid fever in the United States Army and Navy. Am. J. Public Health 1934, 24, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Engels, E.A.; Falagas, M.E.; Lau, J.; Bennish, M.L. Typhoid fever vaccines: A meta-analysis of studies on efficacy and toxicity. BMJ 1998, 316, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanoff, B.; Levine, M.M.; Lambert, P.H. Vaccination against typhoid fever: Present status. Bull. World Health Organ. 1994, 72, 957–971. [Google Scholar] [PubMed]
- Wahdan, M.H.; Sippel, J.E.; Mikhail, I.A.; Rahka, A.E.; Anderson, E.S.; Sparks, H.A.; Cvjetanovic, B. Controlled field trial of a typhoid vaccine prepared with a nonmotile mutant of salmonella typhi Ty2. Bull. World Health Organ. 1975, 52, 69–73. [Google Scholar]
- Nath, T.R.; Malaviya, A.N.; Kumar, R.; Balakrishnan, K.; Singh, B.P. A study of the efficacy of typhoid vaccine in inducing humoral and cell-mediated immune responses in human volunteers. Clin. Exp. Immunol. 1977, 30, 38–43. [Google Scholar] [PubMed]
- Rajagopalan, P.; Kumar, R.; Malaviya, A.N. a study of humoral and cell-mediated immune response following typhoid vaccination in human volunteers. Clin. Exp. Immunol. 1982, 47, 275–282. [Google Scholar] [PubMed]
- Germanier, R.; Furer, E. Isolation and characterization of Gal E mutant Ty 21a of Salmonella typhi: A candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef]
- Fraser, A.; Paul, M.; Goldberg, E.; Acosta, C.J.; Leibovici, L. Typhoid fever vaccines: Systematic review and meta-analysis of randomised controlled trials. Vaccine 2007, 25, 7848–7857. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, S.; Martin, R.; Saxena, A.; Pham, B.; Chiueh, G.; Osorio, M.; Kopecko, D.; Xu, D.Q.; Lechuga-Ballesteros, D.; Truong-Le, V. Room temperature stabilization of oral, live attenuated salmonella enterica serovar typhi-vectored vaccines. Vaccine 2011, 29, 2761–2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryz, S.J., Jr.; Vanprapar, N.; Thisyakorn, U.; Olanratmanee, T.; Losonsky, G.; Levine, M.M.; Chearskul, S. Safety and immunogenicity of Salmonella typhi Ty21a vaccine in young Thai children. Infect. Immun. 1993, 61, 1149–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakkanen, S.H.; Kantele, J.M.; Kantele, A. Cross-reactive gut-directed immune response against salmonella enterica serovar paratyphi A and B in typhoid fever and after oral Ty21 a typhoid vaccination. Vaccine 2012, 30, 6047–6053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, R.; Simon, R.; Zafar, S.J.; Levine, M.M.; Sztein, M.B. Live oral typhoid vaccine Ty21a induces cross-reactive humoral immune responses against Salmonella enterica serovar paratyphi A and S. Paratyphi B in humans. Clin. Vaccine Immunol. 2012, 19, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakkanen, S.H.; Kantele, J.M.; Herzog, C.; Kantele, A. Cross-reactive immune response elicited by parenteral vi polysaccharide typhoid vaccine against non-typhoid Salmonellae. Vaccine 2014, 32, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Kantele, A.; Pakkanen, S.H.; Siitonen, A.; Karttunen, R.; Kantele, J.M. Live oral typhoid salmonella typhi Ty21a—A surrogate vaccine against non-typhoid Salmonella? Vaccine 2010, 30, 7238–7245. [Google Scholar] [CrossRef] [PubMed]
- Tacket, C.O.; Pasetti, M.F.; Sztein, M.B.; Livio, S.; Levine, M.M. Immune responses to an oral typhoid vaccine strain that is modified to constitutively express vi capsular polysaccharide. J. Infect. Dis. 2004, 190, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.C.; Ho, V.A.; Khiem, H.B.; Trach, D.D.; Bay, P.V.; Thanh, T.C.; Kossaczka, Z.; Bryla, D.A.; Shiloach, J.; Robbins, J.B.; et al. The efficacy of a Salmonella typhi Vi conjugate vaccine in two-to-five-year-old children. N. Engl. J. Med. 2001, 344, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Thiem, V.D.; Lin, F.-Y.; Canh, D.G.; Son, N.H.; Anh, D.D.; Mao, N.D.; Chu, C.; Hunt, S.W.; Robbins, J.B.; Schneerson, R.; et al. The Vi conjugate typhoid vaccine is safe, elicits protective levels of igG anti-vi, and is compatible with routine infant vaccines. Clin. Vaccine Immunol. 2011, 18, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLennan, C.A. Vaccines for low-income countries. Semin. Immunol. 2013, 25, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podda, A.; Saul, A.; Arora, R.; Bhutta, Z.; Sinha, A.; Gaind, R.; Singhal, T.; Saha, S.; Brooks, A.; Martin, L.B.; et al. Conjugated vaccines for enteric fever: Proceedings of a meeting organized in New Delphi, India in 2009. J. Infect. Dev. Ctries. 2010, 4, 404–411. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Gondwe, E.N.; Msefula, C.L.; Kingsley, R.A.; Thomson, N.R.; White, S.A.; Goodall, M.; Pickard, D.J.; Graham, S.M.; Dougan, G.; et al. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J. Clin. Investig. 2008, 118, 1553–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feasey, N.A.; Archer, B.N.; Heyderman, R.S.; Sooka, A.; Dennis, B.; Gordon, M.A.; Keddy, K.H. Typhoid fever and invasive nontyphoid Salmonellosis, Malawi and South Africa. Emerg. Infect. Dis. 2010, 16, 1448–1451. [Google Scholar] [CrossRef] [PubMed]
- Gondwe, E.N.; Molyneux, M.E.; Goodall, M.; Graham, S.M.; Mastroeni, P.; Drayson, M.T.; MacLennan, C.A. Importance of antibody and complement for oxidative burst and killing of invasive nontyphoidal Salmonella by blood cells in Africans. Proc. Natl. Acad. Sci. USA 2010, 107, 3070–3075. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; Tennant, S.M.; Wang, J.Y.; Schmidlein, P.J.; Lees, A.; Ernst, R.K.; Pasetti, M.F.; Galen, J.E.; Levine, M.M. Salmonella enterica serovar enteritidis core O polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. enteritidis. Infect. Immun. 2011, 79, 4240–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, D.C.; Robbins, J.B.; Szu, S.C. Protection of Mice against Salmonella typhimurium with an O-specific polysaccharide-protein conjugate vaccine. Infect. Immun. 1992, 60, 4679–4686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlin, N.I.A.; Svenson, S.B.; Lindberg, A.A. Role of monoclonal O-antigen antibody epitope specificity and isotype in protection against experimental mouse typhoid. Microb. Pathog. 1987, 2, 171–183. [Google Scholar] [CrossRef]
- Singh, S.P.; Williams, Y.U.; Benjamin, W.H.; Klebba, P.E.; Boyd, D. Immunoprotection by monoclonal antibodies to the porins and lipopolysaccharide of Salmonella typhimurium. Microb. Pathog. 1996, 21, 249–263. [Google Scholar] [CrossRef]
- Rondini, S.; Lanzilao, L.; Necchi, F.; O’Shaughnessy, C.M.; Micoli, F.; Saul, A.; MacLennan, C.A. Invasive African salmonella typhimurium induces bactericidal antibodies against O-antigens. Microb. Pathog. 2013, 63, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A.; Banda, H.T.; Gondwe, M.; Gordon, S.B.; Boeree, M.J.; Walsh, A.L.; Corkill, J.E.; Hart, A.; Gilks, C.F.; Molyneux, M.E. Non-typhoidal Salmonella bacteraemia among HIV-infected Malawian adults: High mortality and frequent recrudescence. AIDS 2002, 16, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, G.M.; Neu, H.C. Agents responsible for infection in chronic granulomatous disease of childhood. J. Pediatr. 1975, 86, 415–417. [Google Scholar] [CrossRef]
- Svenson, S.B.; Lindberg, A.A. Artificial Salmonella vaccines: Salmonella typhimurium O-antigen-specific oligosaccharide-protein conjugates elicit protective antibodies in rabbits and mice. Infect. Immun. 1981, 32, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; Levine, M.M. Glycoconjugate vaccine strategies for protection against invasive Salmonella infections. Hum. Vaccines Immunother. 2012, 8, 494–498. [Google Scholar] [CrossRef] [Green Version]
- Svenson, S.B.; Nurminen, M.; Lindberg, A.A. Artificial Salmonella vaccines: O-antigenic oligosaccharide-protein conjugates induce protection against infection with Salmonella typhimurium. Infect. Immun. 1979, 25, 863–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, J.S.; Kim, J.S.; Jang, J.I.; Kim, B.-H.; Yoo, S.Y.; Choi, J.H.; Bang, L.-S.; Lee, I.S.; Park, Y.K. Enhancement of host immune responses by oral vaccination to Salmonella enterica serovar typhimurium harboring both FliC and FljB flagella. PLoS ONE 2013, 8, e74850. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Wang, J.Y.; Boyd, M.A.; Tulapurkar, M.E.; Ramachandran, G.; Tennant, S.M.; Pasetti, M.; Galen, J.E.; Levine, M.M. Sustained protection in mice immunized with fractional doses of Salmonella enteritidis core and O polysaccharide-flagellin glycoconjugates. PLoS ONE 2013, 8, e64680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobat, S.; Flores-Langarica, A.; Hitchcock, J.; Marshall, J.L.; Kingsley, R.A.; Goodall, M.; Gil-Cruz, C.; Serre, K.; Leyton, D.L.; Letran, S.E.; et al. Soluble flagellin, FliC, induces an Ag-specific Th2 response, yet promotes T-bet-regulated Th1 clearance of Salmonella typhimurium infection. J. Immunol. 2011, 41, 1606–1618. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Khan, M.; Ball, J.; Toellner, K.-M.; Serre, K.; Mohr, E.; MacLennan, I.C.M. Responses to the soluble flagellar protein FliC are Th2, while those to FliC on Salmonella are Th2. J. Immunol. 2004, 34, 2986–2995. [Google Scholar]
- Kang, H.Y.; Srinivasan, J.; Curtiss, R., III. Immune responses to recombinant pneumococal PspA antigen delivered by live attenuated Salmonella enterica serovar typhimurium vaccine. Infect. Immun. 2002, 70, 1739–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Liu, Q.; Bian, X.; Wang, S.; Curtiss, R., 3rd; Kong, Q. Synthesis and delivery of streptococcus pneumoniae capsular polysaccharides by recombinant attenuated Salmonella vaccines. Proc. Natl. Acad. Sci. USA 2021, 118, e2013350118. [Google Scholar] [CrossRef]
- Juarez-Rodriguez, M.D.; Arteaga-Cortes, L.T.; Kader, R.; Curtiss, R., 3rd. Live attenuated Salmonella vaccines against mycobacterium tuberculosis with antigen delivery via the type III secretion system. Infect. Immun. 2012, 80, 798–814. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, S.; Scarpellini, G.; Gunn, B.; Xin, W.; Wanda, S.-Y.; Roland, K.L.; Curtis, R., 3rd. Evaluation of new generation of Salmonella enterica serovar typhimurium vaccines with regulated delayed attenuation to induce immune responses against PspA. Proc. Natl. Acad. Sci. USA 2009, 106, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kong, Q.; Curtis, R., 3rd. New technologies in developing recombinant attenuated Salmonella vaccine vectors. Microb. Pathog. 2013, 58, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, Y.; Scarpellini, G.; Kong, W.; Shi, H.Y.; Baek, C.-H.; Gunn, B.; Wanda, S.-Y.; Roland, K.L.; Zhang, X.; et al. Salmonella vaccine vectors displaying delayed antigen synthesis in vivo to enhance immunogenicity. Infect. Immun. 2010, 78, 3969–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.Y.; Curtis, R., 3rd. Immune responses dependent on antigen location in recombinant attenuated Salmonella typhimurium vaccines following oral immunization. FEMS Immunol. Med. Mic. 2003, 37, 99–104. [Google Scholar] [CrossRef]
- Gil-Cruz, C.; Bobat, S.; Marshall, J.L.; Kingsley, R.A.; Ross, E.A.; Henderson, I.R.; Leyton, D.L.; Coughlan, R.E.; Khan, M.; Jensen, K.T.; et al. The porin ompd from nontyphoidal salmonella is a key target for a protective B1b cell antibody response. Proc. Natl. Acad. Sci. USA 2009, 106, 9803–9808. [Google Scholar] [CrossRef] [Green Version]
- Secundino, I.; Lopez-Macias, C.; Cervantes-Barragan, L.; Gil-Cruz, C.; Rios-Sarabia, N.; Pastelin-Palacios, R.; Villasis-Keever, M.A.; Becker, I.; Puente, J.L.; Calva, E.; et al. Salmonella porins induce a sustained, lifelong specific bactericidal antibody memory response. Immunology 2005, 117, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Gonzalez, R.M.; Maldonado-Bernal, C.; Ramirez-Cruz, N.E.; Rios-Sarabia, N.; Beltran-Nava, J.; Castanon-Gonzalez, J.; Castillo-Torres, N.; Palma-Aguirre, J.A.; Carrera-Camargo, M.; Lopez-Macias, C.; et al. Induction of cellular immune response and anti-Salmoneall enterica serovar typhi bactericidal antibodies in healthy volunteers by immunization with a vaccine candidate against typhoid fever. Immunol. Lett. 2004, 93, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Rappuoli, R. Reverse vaccinology: Developing vaccines in the era of genomics. Immunity 2010, 33, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Fiorino, F.; Pettini, E.; Koeberling, O.; Ciabattini, A.; Pozzi, G.; Martin, L.B.; Medaglini, D. Long-term anti-bacterial immunity against systemic infection by salmonella enterica serovar typhimurium elicited by a gmma-based vaccine. Vaccines 2021, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Bjarnarson, S.P.; Arcuri, M.; Pindi, A.A.A.; Magnusdottir, G.J.; Necchi, F.; Benedetto, R.D.; Carducci, M.; Schiavo, F.; Giannelli, C.; et al. Short vi-polysaccharide abroages t-independent immune response and hyporesponsiveness elicited by long Vi-CRM197 conjugate vaccine. Proc. Natl. Acad. Sci. USA 2020, 117, 24443–24449. [Google Scholar] [CrossRef]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Palmieri, E.; Balocchi, C.; Gasperini, G.; et al. GMMA is a versatile platform to design effective multivalent combination vaccines. Vaccines 2020, 8, 540. [Google Scholar] [CrossRef]
Figure 1.
Transmission and prevention of Salmonella infections. Human transmission of Salmonella primarily occurs through ingestion of contaminated food or water, contact with infected animals or animal feed and person-to-person transmission. Dual arrows are shown between humans and animals because transmission can occur in both directions. The best preventive measures to reduce the incidence of human infections include proper sanitation, treatment of infected individuals, vaccination (when possible) and access to clean water.
Figure 1.
Transmission and prevention of Salmonella infections. Human transmission of Salmonella primarily occurs through ingestion of contaminated food or water, contact with infected animals or animal feed and person-to-person transmission. Dual arrows are shown between humans and animals because transmission can occur in both directions. The best preventive measures to reduce the incidence of human infections include proper sanitation, treatment of infected individuals, vaccination (when possible) and access to clean water.

Figure 2.
Antibody production in response to polysaccharide antigens. (A) Polysaccharide antigens alone lead to IgM production, while (B) polysaccharide–protein conjugated antigens lead to IgG production and immune memory.
Figure 2.
Antibody production in response to polysaccharide antigens. (A) Polysaccharide antigens alone lead to IgM production, while (B) polysaccharide–protein conjugated antigens lead to IgG production and immune memory.

Figure 3.
General concepts for development of vaccines. Pure polysaccharide vaccines are T-cell-independent activators of B-cells and their use usually results in an antibody only response (i.e., IgM). Polysaccharide–protein conjugate vaccines typically generate both humoral and cell-mediated immune responses. Both live-attenuated (i.e., mutagenesis) and inactivated whole-cell (i.e., heat or chemically inactivated) vaccines mount a mixed cell-mediated and humoral response dominated by IgG antibodies.
Figure 3.
General concepts for development of vaccines. Pure polysaccharide vaccines are T-cell-independent activators of B-cells and their use usually results in an antibody only response (i.e., IgM). Polysaccharide–protein conjugate vaccines typically generate both humoral and cell-mediated immune responses. Both live-attenuated (i.e., mutagenesis) and inactivated whole-cell (i.e., heat or chemically inactivated) vaccines mount a mixed cell-mediated and humoral response dominated by IgG antibodies.

Table 2.
Human vaccines currently licensed and vaccine candidates being developed for S. Typhi, S. Paratyphi A, and non-typhoidal Salmonella a.
Table 2.
Human vaccines currently licensed and vaccine candidates being developed for S. Typhi, S. Paratyphi A, and non-typhoidal Salmonella a.
| Description | Developer | Academic/Commercial | Phase |
|---|---|---|---|
| Vaccines Licensed for S. Typhi | |||
| Neotyf, Typhoral, Vivotif (Ty21a live attenuated) | Johnson & Johnson | Commercial | Marketed |
| Bio Typh (Vi capsular polysaccharide) | BioMed | Commercial | Marketed |
| Typhim Vi, Typhyvax (Vi capsular polysaccharide) | Sanofi (Pasteur Merieux) | Commercial | Marketed |
| Typherix (Vi capsular polysaccharide) | GlaxoSmithKline | Commercial | Marketed |
| Peda-Typh (conjugate) | BioMed | Commercial | Marketed |
| Typbar-TCV (conjugate) | Bharat Biotech | Commercial | Marketed |
| Typhoid vaccine | Microgen | Commercial | Marketed |
| Vi Polysaccharide typhoid vaccine | China National Pharmaceutical (Beijing Tiantan Biological) | Commercial | Marketed |
| Vi-rEPA (Vi conjugate) | Lanzhou Institute (China) | Commercial | Marketed |
| Tyvax VI plus | VHB Life Sciences | Commercial | Marketed |
| Typhoid vaccine | Zydus Cadila (Zydus Vaccicare) | Commercial | Marketed |
| Tyrix Vi | SK Holdings | Commercial | Marketed |
| Shantyph | Sanofi (Shantha Biotechnics) | Commercial | Marketed |
| Typhobox, Typhovax | Green Cross | Commercial | Marketed |
| Zerotyph | Boryung | Commercial | Marketed |
| vax-TyVi | Finlay Institute | Commercial | Marketed |
| Typhoid-Kovax | Sanofi | Commercial | Marketed |
| Hepatyrix | GlaxoSmithKline | Commercial | Marketed |
| VIVAXIM | Sanofi | Commercial | Marketed |
| Biovac Typhoid | Wockhardt | Commercial | Marketed |
| Typho-Vi | Bio-Med | Commercial | Marketed |
| Vaccines in Development in Recent Years for S. Typhi | |||
| Vi-DT typhoid conjugate vaccine | Bio Farma | Commercial | Phase I |
| Typhetec | Prokarium | Commercial | Phase I |
| OmpC and OmpF | Institute Mexicano del Seguro Social | Academic | Phase 1 in Mexico |
| Enteric fever vaccine | Prokarium | Commercial | Phase II |
| M01ZH09 (live attenuated) | Emergent BioSolutions | Commercial | Phase II |
| CVD 909 (live attenuated) | University of Maryland | Academic | Phase II |
| Ty800 (live attenuated) | Avant Immunotherapeutics | Commercial | Phase II |
| Vi-rEPA (Vi conjugate) | National Health Institute | Academic | Phase III |
| Vi-CRM (Vi conjugate) | Biological E | Commercial | Phase III |
| Vi conjugated to fusion protein PsaA-PdT | Harvard Medical School | Academic | Preclinical |
| O:9-DT (conjugate) | International Vaccine Institute (IVI) | Academic | Preclinical |
| Salmonella typhi + paratyphi vaccine | Prokarium | Commercial | Research/Preclinical |
| Salmonella Vaccine Project | Affinivax | Commercial | Research/Preclinical |
| Vi conjugated to fusion protein PsaA-PdT | Harvard Medical School | Academic | Research/Preclinical |
| Ty21a typhoid vaccine expressing Shigella LPS | Protein Potential LLC. | Commercial | Research/Preclinical |
| O:2,12-DT + Vi-DT | International Vaccine Institute (IVI) | Academic | Research/Preclinical |
| O:2,12-CRM197 + Vi-CRM197 | Biological E | Commercial | Research/Preclinical |
| Vaccines in Development in Recent Years for S. Paratyphi | |||
| CVD 1902 (live attenuated) | University of Maryland, Bharat Biotech | Commercial | Phase I |
| Paratyphoid A conjugate vaccine | Lanzhou Institute (China) | Academic | Phase II |
| O:2-TT (conjugate) | Technology transfer from NIH to Lanzhou Institute (China) | Commercial | Phase II |
| O:2,12-DT + Vi-DT | International Vaccine Institute (IVI) | Academic | Research/Preclinical |
| O:2,12-CRM197 + Vi-CRM197 | Biological E | Commercial | Research/Preclinical |
| O:2-TT (conjugate) | Technology transfer from NIH to Chengdu Institute (China) | Academic | Preclinical |
| O:2-TT (conjugate) | Changchun Institute of Biological Products | Commercial | Preclinical |
| NTS Vaccines in Development in Recent Years | |||
| WT05—aroC and ssaV mutants (live attenuated) | Microscience, Wokingham Berkshire | Academic | Phase 1 |
| NTS Vaccine | University of Maryland | Academic | Research/Preclinical |
| CVD 1921 and CVD 1941 NTS Project (live attenuated) | University of Maryland, Bharat Biotec, Wellcome Trust | Commercial | Research/Preclinical |
| Bivalent iNTS-GMMA | GlaxoSmithKline (Novartis Vaccine Institute for Global Health; NVGH) | Commercial | Research/Preclinical |
| Bivalent Conjugate (O:1,4[5],12-CRM197 + O:1,9,12-CRM197) | GlaxoSmithLine (Novartis Vaccine Institute for Global Health; NVGH NVGH) | Commercial | Research/Preclinical |
| OmpD | University of Birmingham | Academic | Research/Preclinical |
| O:4,5/O:9-flagellin (conjugate) | University of Maryland | Academic | Preclinical |
| O:4, 12-TT (conjugate) | National Institutes for Health NIH) | Academic | Preclinical |
| Os-po (O:4 porin conjugate) | National Bacteriology Laboratory, Stockholm | Preclinical | |
| S. Typhimurium ruvB mutant (live attenuated) | Seoul National University | Academic | Preclinical |
| Salmonella hfq deletion mutant (live attenuated) | Indian Institute of Science Bangalore | Academic | Preclinical |
| SA186 (live attenuated) | Istituto Superiore di Sanita Roma | Academic | Preclinical |
| MT13 (live attenuated) | KIIT University Odisha | Academic | Preclinical |
| DNA adenine methylase mutants (live attenuated) | University of California, Santa Barbara | Academic | Preclinical |
| live attenuated, regulated delayed attenuation | Arizona State University | Academic | Preclinical |
a This is not a comprehensive list of all the vaccine candidates that are being tested, especially since this list does not reflect vaccine candidates tested in research studies. This information was compiled from the Boston Consulting Group—Vaccine pipeline information web page (https://vaccinesforamr.org/review-of-pathogens/vaccine-pipeline-information/#Salmonella_non-typhoidal, accessed on April 2021), National Institute of Health—U.S. National Library of Medicine ClinicalTrials.gov web page (https://clinicaltrials.gov/ct2/home, accessed on April 2021), and the MacLennan et al., 2014 research article [53].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Perera, S.R.; Sokaribo, A.S.; White, A.P. Polysaccharide Vaccines: A Perspective on Non-Typhoidal Salmonella. Polysaccharides 2021, 2, 691-714. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030042
AMA Style
Perera SR, Sokaribo AS, White AP. Polysaccharide Vaccines: A Perspective on Non-Typhoidal Salmonella. Polysaccharides. 2021; 2(3):691-714. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030042
Chicago/Turabian StylePerera, Sumudu R., Akosiererem S. Sokaribo, and Aaron P. White. 2021. "Polysaccharide Vaccines: A Perspective on Non-Typhoidal Salmonella" Polysaccharides 2, no. 3: 691-714. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2030042