Tau Protein in Lung Smooth Muscle Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
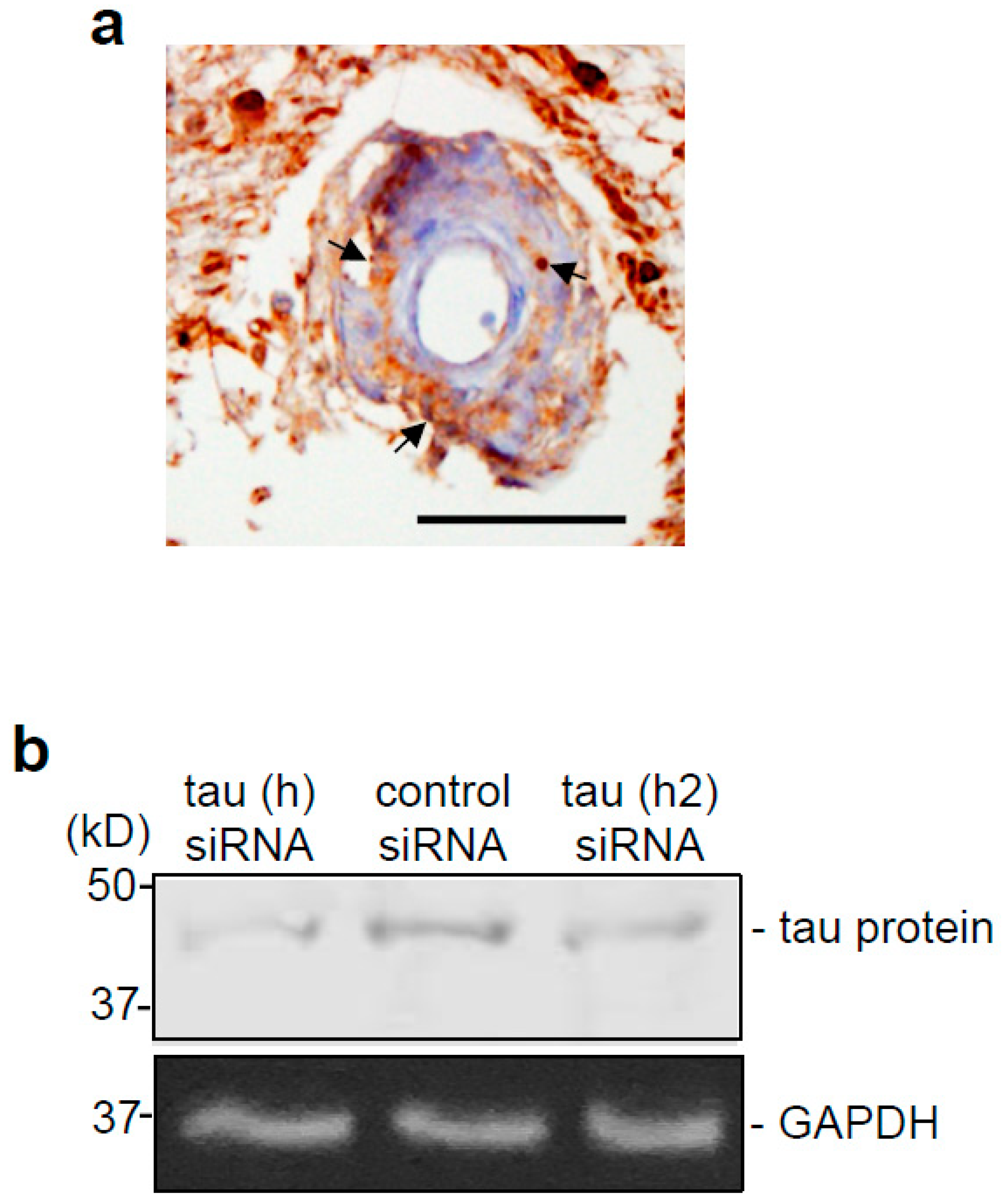
2.1. Histology
2.2. Cell Culture
2.3. Western Blotting
2.4. Immunofluorescence Analysis
2.5. Statistical Analysis
3. Results
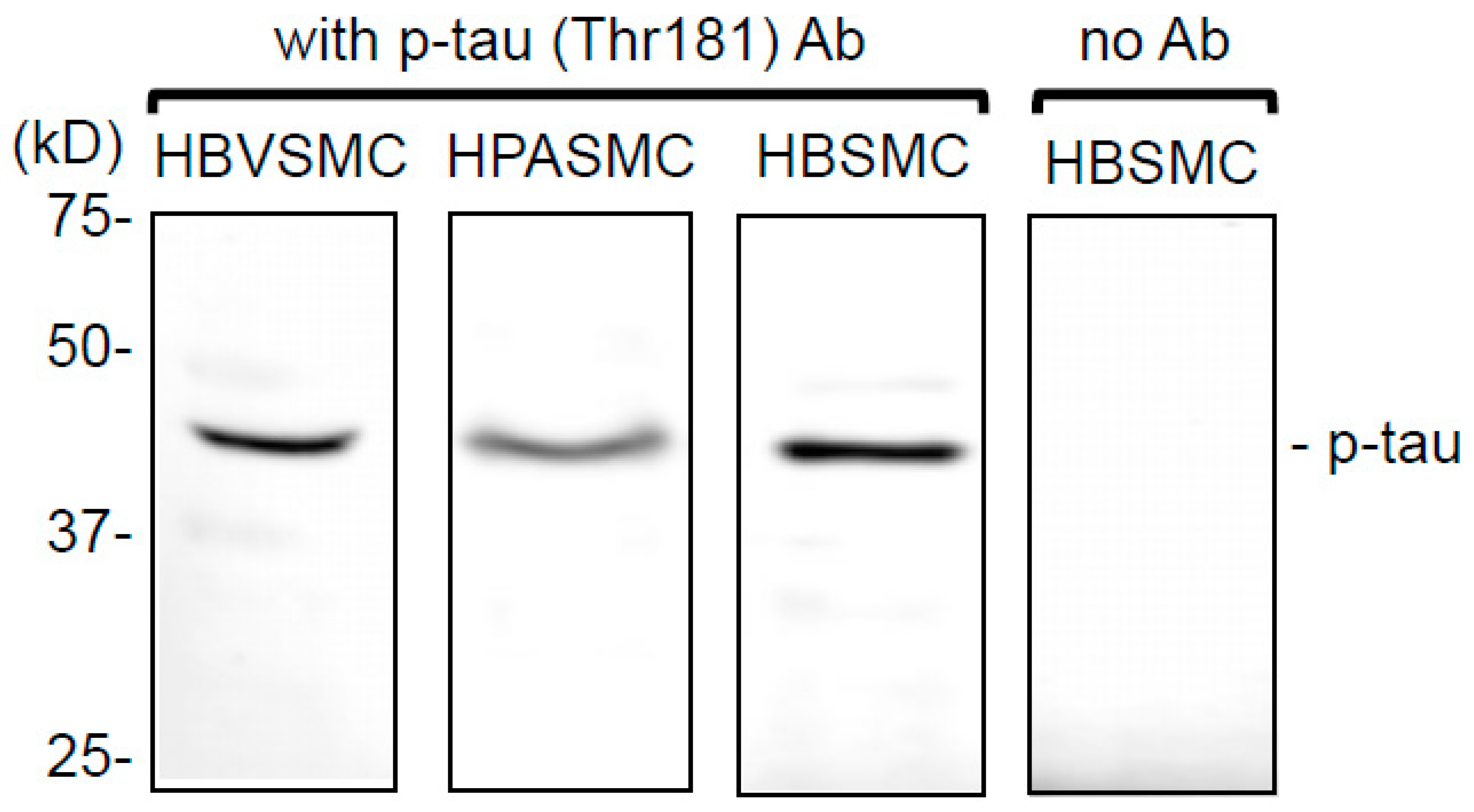
3.1. Tau Protein is Expressed and Constitutively Phosphorylated at Threonine 181 in Smooth Muscle Cells
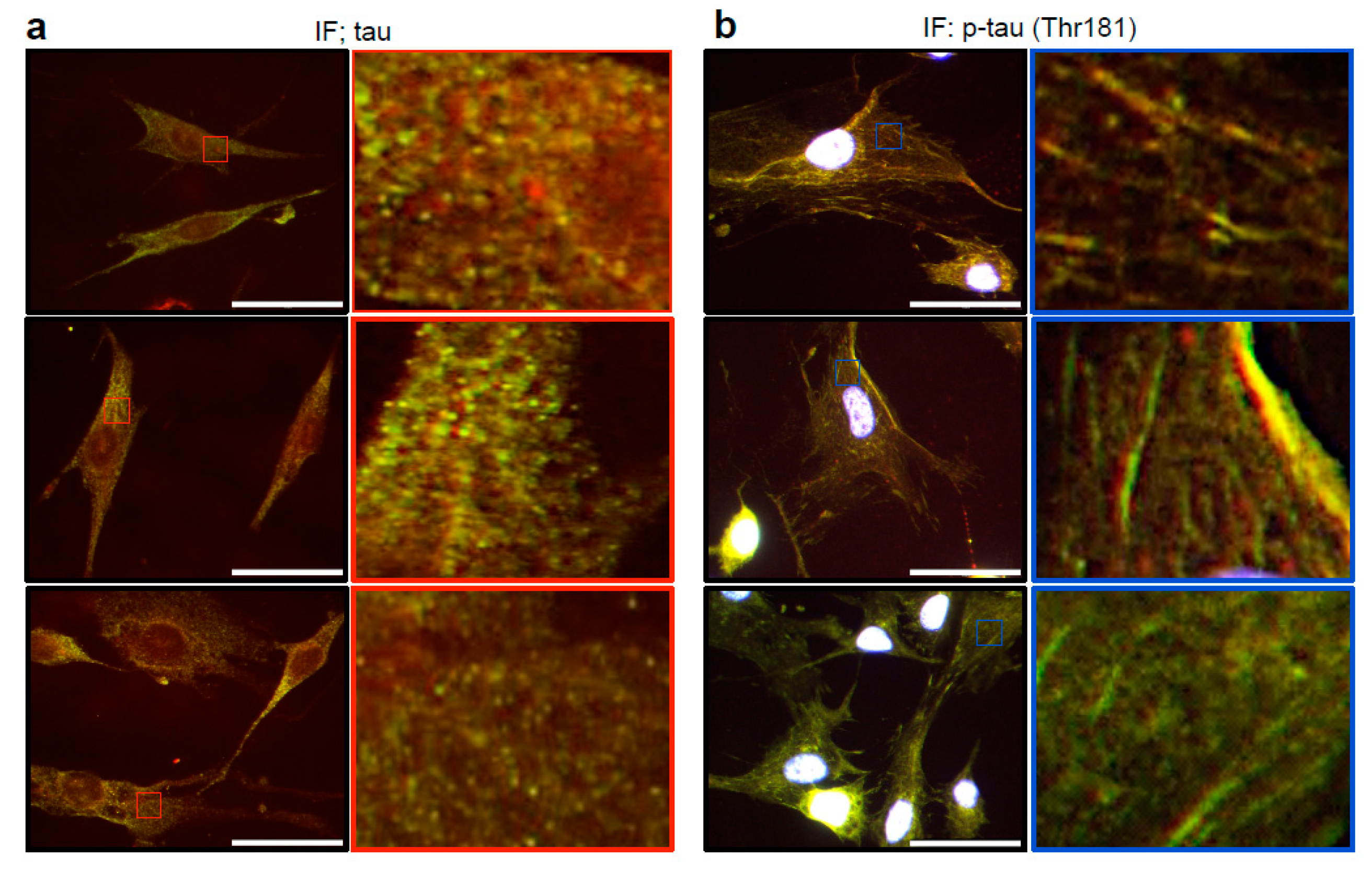
3.2. Cytoskeletal Organization of Tau and Phosphorylated Tau in Lung Vascular Smooth Muscle Cells
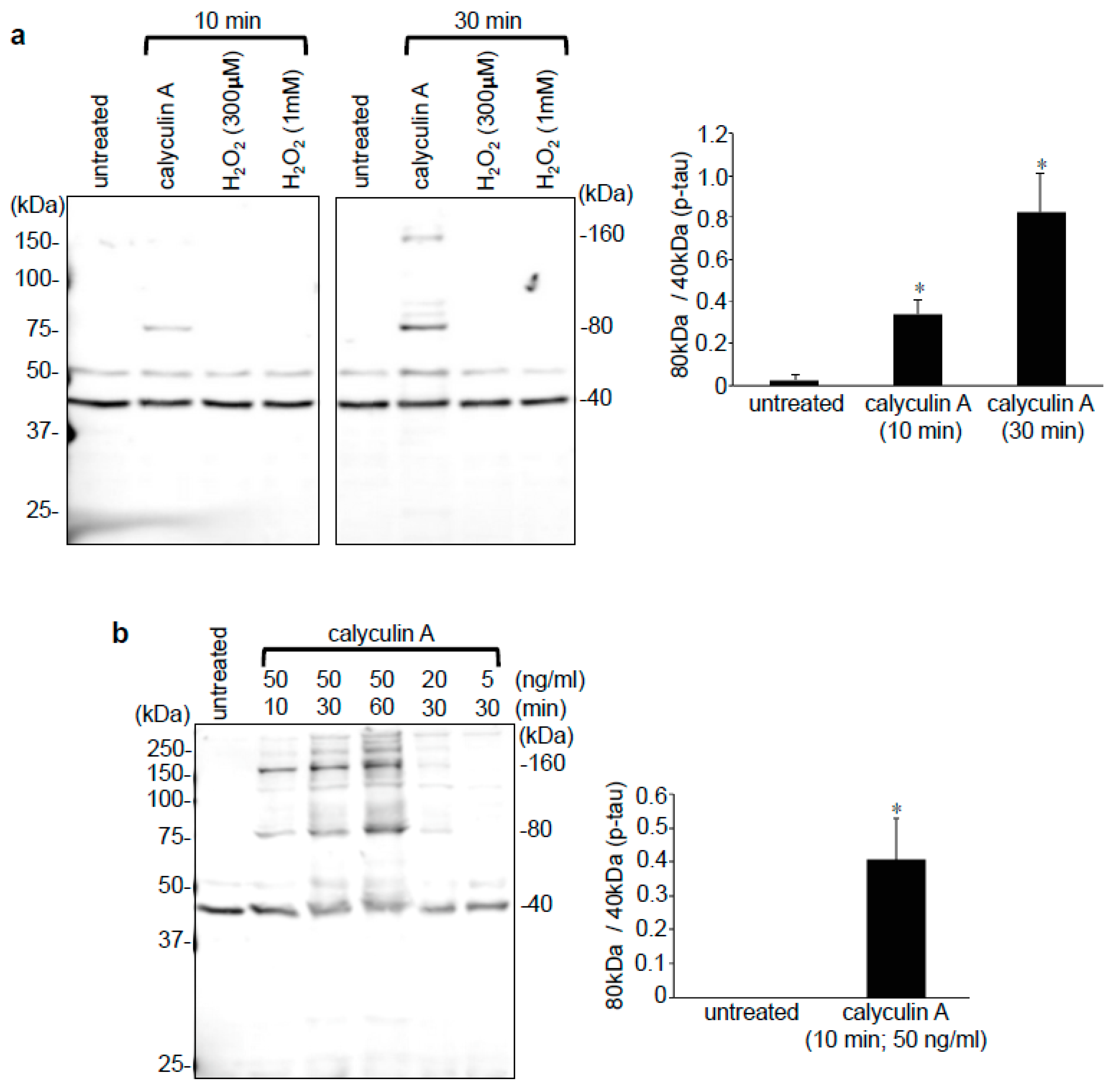
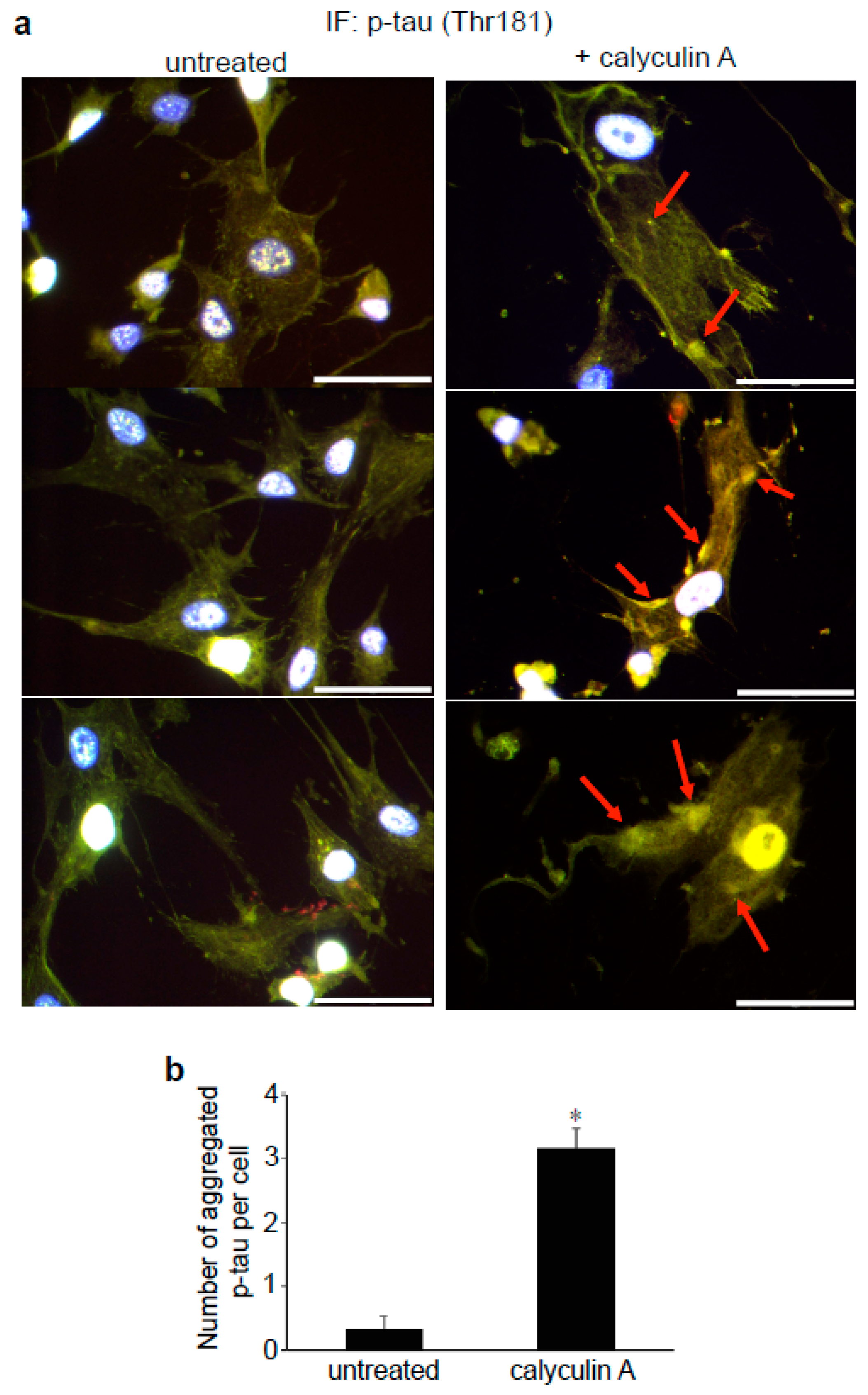
3.3. Effects of Calyculin A in Lung Vascular and Airway Smooth Muscle Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Grundke-Iqbal, I.; Iqbal, K.; Tung, Y.C.; Quinlan, M.; Wisniewski, H.M.; Binder, L.I. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc. Natl. Acad. Sci. USA 1986, 83, 4913–4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, L.I.; Frankfurter, A.; Rebhun, L.I. The distribution of tau in the mammalian central nervous system. J. Cell Biol. 1985, 101, 1371–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, L.; Sun, X.; Austin, T.O.; Muralidharan, H.; Jean, D.C.; Liu, M.; Yu, W.; Baas, P.W. Tau does not stabilize axonal microtubules but rather enables them to have long labile domains. Curr. Biol. 2018, 28, 2181–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, L.D.; Wassmer, T.; Fraser, G.; Smith, J.; Perkinton, M.; Billinton, A.; Livesey, F.J. Extracellular monomeric and aggregated tau efficiently enter human neurons through overlapping but distinct pathways. Cell Rep. 2018, 22, 3612–3624. [Google Scholar] [CrossRef] [Green Version]
- Drubin, D.G.; Kirschner, M.W. Tau protein function in living cells. J. Cell Biol. 1986, 103, 2739–2746. [Google Scholar] [CrossRef] [Green Version]
- Weingarten, M.D.; Lockwood, A.H.; Hwo, S.Y.; Kirschner, M.W. A protein factor essential for microtubule assembly. Proc. Natl. Acad. Sci. USA 1975, 72, 1858–1862. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Sen, N. Tauopathy: A common mechanism for neurodegeneration and brain aging. Mech. Ageing Dev. 2019, 178, 72–79. [Google Scholar] [CrossRef]
- Mudher, A.; Colin, M.; Dujardin, S.; Medina, M.; Dewachter, I.; Alavi Naini, S.M.; Mandelkow, E.M.; Mandelkow, E.; Buee, L.; Goedert, M.; et al. What is the evidence that tau pathology spreads through prion-like propagation? Acta Neuropathol. Commun. 2017, 5, 99. [Google Scholar] [CrossRef]
- Mukrasch, M.D.; von Bergen, M.; Biernat, J.; Fischer, D.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. The “jaws” of the tau-microtubule interaction. J. Biol. Chem. 2017, 282, 12230–12239. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Guo, J.L.; Changolkar, L.; Stieber, A.; McBride, J.D.; Silva, L.V.; He, Z.; Zhang, B.; Gathagan, R.J.; Trojanowski, J.Q.; et al. Pathological tau strains from human brains recapitulate the diversity of Tauopathies in nontransgenic mouse brain. J. Neurosci. 2017, 37, 11406–11423. [Google Scholar] [CrossRef]
- Rossi, G.; Dalprà, L.; Crosti, F.; Lissoni, S.; Sciacca, F.L.; Catania, M.; Di Fede, G.; Mangieri, M.; Giaccone, G.; Croci, D.; et al. A new function of microtubule-associated protein tau: Involvement in chromosome stability. Cell Cycle 2008, 7, 1788–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, G.; Conconi, D.; Panzeri, E.; Redaelli, S.; Piccoli, E.; Paoletta, L.; Dalprà, L.; Tagliavini, F. Mutations in MAPT gene cause chromosome instability and introduce copy number variations widely in the genome. J. Alzheimers Dis. 2013, 33, 969–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ittner, L.M.; Ke, Y.D.; Delerue, F.; Bi, M.; Gladbach, A.; van Eersel, J.; Wölfing, H.; Chieng, B.C.; Christie, M.J.; Napier, I.A.; et al. Dendritic function of tau mediates amyloid-beta toxicity in Alzheimer’s disease mouse models. Cell 2010, 142, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, G.V.W.; Hartigan, J.A. Tau protein in normal and Alzheimer’s disease brain: An update. J. Alzheimers Dis. 1999, 1, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Vuono, R.; Winder-Rhodes, S.; de Silva, R.; Cisbani, G.; Drouin-Ouellet, J. The role of tau in the pathological process and clinical expression of Huntington's disease. Brain 2015, 138, 1907–1918. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Nogales, M.; Cabrera, J.R.; Santos-Galindo, M.; Hoozemans, J.J.; Ferrer, I.; Rozemuller, A.J.; Hernández, F.; Avila, J.; Lucas, J.J. Altered levels and isoforms of tau and nuclear membrane invaginations in Huntington's disease. Front. Cell. Neurosci. 2019, 13, 574. [Google Scholar] [CrossRef]
- Ballatore, C.; Lee, V.M.; Trojanowski, J.Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat. Rev. Neurosci. 2007, 8, 663–672. [Google Scholar] [CrossRef]
- Chin, S.S.; Goldman, J.E. Glial inclusions in CNS degenerative diseases. J. Neuropathol. Exp. Neurol. 1996, 55, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Feany, M.B.; Dickson, D.W. Neurodegenerative disorders with extensive tau pathology: A comparative study and review. Ann. Neurol. 1996, 40, 139–148. [Google Scholar] [CrossRef]
- Komori, T. Tau-positive glial inclusions in progressive supranuclear palsy, corticobasal degeneration and Pick’s disease. Brain Pathol. 1999, 9, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Poorkaj, P.; Bird, T.D.; Wijsman, E.; Nemens, E.; Garruto, R.M.; Anderson, L.; Andreadis, A.; Wiederholt, W.C.; Raskind, M.; Schellenberg, G.D. Tau is a candidate gene for chromosome 17 frontotemporal dementia. Ann. Neurol. 1998, 43, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mandelkow, E. Tau in physiology and pathology. Nat. Rev. Neurosci. 2016, 17, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Šimić, G.; Babić Leko, M.; Wray, S.; Harrington, C.; Delalle, I.; Jovanov-Milošević, N.; Bažadona, D.; de Silva, R.; Di Giovanni, G.; Wischik, C.; et al. Tau protein hyperphosphorylation and aggregation in Alzheimer’s disease and other tauopathies, and possible neuroprotective strategies. Biomolecules 2016, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, V.; Lu, Y.; Steinhilb, M.L.; Oldham, S.; Shulman, J.M.; Feany, M.B. TOR-mediated cell-cycle activation causes neurodegeneration in a Drosophila tauopathy model. Curr. Biol. 2006, 16, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Steinhilb, M.L.; Dias-Santagata, D.; Mulkearns, E.E.; Shulman, J.M.; Biernat, J.; Mandelkow, E.M.; Feany, M.B. S/P and T/P phosphorylation is critical for tau neurotoxicity in Drosophila. J. Neurosci. Res. 2007, 85, 1271–1278. [Google Scholar] [CrossRef]
- Ingelson, M.; Vanmechelen, E.; Lannfelt, L. Microtubule-associated protein tau in human fibroblasts with the Swedish Alzheimer mutation. Neurosci. Lett. 1996, 220, 9–12. [Google Scholar] [CrossRef]
- Thurston, V.C.; Zinkowski, R.P.; Binder, L.I. Tau as a nucleolar protein in human nonneural cells in vitro and in vivo. Chromosoma 1996, 105, 20–30. [Google Scholar] [CrossRef]
- Sotiropoulos, I.; Galas, M.C.; Silva, J.M.; Skoulakis, E.; Wegmann, S.; Maina, M.B.; Blum, D.; Sayas, C.L.; Mandelkow, E.M.; Mandelkow, E.; et al. Atypical, non-standard functions of the microtubule associated tau protein. Acta Neuropathol. Commun. 2017, 5, 91. [Google Scholar] [CrossRef] [Green Version]
- Spillantini, M.G.; Goedert, M. Tau pathology and neurodegeneration. Lancet Neurol. 2013, 12, 609–622. [Google Scholar] [CrossRef]
- Bou Samra, E.; Buhagiar-Labarchede, G.; Machon, C.; Guitton, J.; Onclercq-Delic, R.; Green, M.R.; Alibert, O.; Gazin, C.; Veaute, X.; Amor-Gueret, M. A role for tau protein in maintaining ribosomal DNA stability and cytidine deaminase-deficient cell survival. Nat. Commun. 2017, 8, 693. [Google Scholar] [CrossRef] [PubMed]
- Tsartsalis, S.; Xekardaki, A.; Hof, P.R.; Kovari, E.; Bouras, C. Early Alzheimer-type lesions in cognitively normal subjects. Neurobiol. Aging 2018, 62, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Bell, M.; Lyons, D.N.; Rodriguez-Rivera, J.; Ingram, A.; Fontaine, S.N.; Mechas, E.; Chen, J.; Wolozin, B.; LeVine, H.; et al. Pathological tau promotes neuronal damage by impairing ribosomal function and decreasing protein synthesis. J. Neurosci. 2016, 36, 1001–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, B.; Hemberg, M.; Lewis, J.; Feany, M.B. Tau promotes neurodegeneration through global chromatin relaxation. Nat. Neurosci. 2014, 7, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Hamdane, M.; Bretteville, A.; Sambo, A.V.; Schindowski, K.; Begard, S.; Delacourte, A.; Bertrand, P.; Buee, L. p25/Cdk5-mediated retinoblastoma phosphorylation is an early event in neuronal cell death. J. Cell Sci. 2005, 118, 1291–1298. [Google Scholar] [CrossRef] [Green Version]
- Burnouf, S.; Martire, A.; Derisbourg, M.; Laurent, C.; Belarbi, K.; Leboucher, A.; Fernandez-Gomez, F.J.; Troquier, L.; Eddarkaoui, S.; Grosjean, M.E.; et al. NMDA receptor dysfunction contributes to impaired brain-derived neurotrophic factor-induced facilitation of hippocampal synaptic transmission in a tau transgenic model. Aging Cell 2013, 12, 11–23. [Google Scholar] [CrossRef]
- Lerea, K.M.; Venjara, A.Y.; Olson, S.C.; Kelly, M.R. Threonine phosphorylation of integrin beta3 in calyculin A-treated platelets is selectively sensitive to 5’-iodotubercidin. Biochim. Biophys. Acta 2007, 1773, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, M.; Toyoda, H.; Saito, M.; Morita, K.; Tawara, I.; Deguchi, K.; Kuno, T.; Shima, H.; Nagao, M.; Shirakawa, S. Calyculin A and okadaic acid inhibit human platelet aggregation by blocking protein phosphatases types 1 and 2A. Cell. Signal. 1994, 6, 59–71. [Google Scholar] [CrossRef]
- Takai, A.; Eto, M.; Hirano, K.; Takeya, K.; Wakimoto, T.; Watanabe, M. Protein phosphatase 1 and 2A and their naturally occurring inhibitors: Current topics in smooth muscle physiology and chemical biology. J. Physiol. Sci. 2018, 68, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Boban, M.; Babić Leko, M.; Miškić, T.; Hof, P.R.; Šimić, G. Human neuroblastoma SH-SY5Y cells treated with okadaic acid express phosphorylated high molecular weight tau-immunoreactive protein species. J. Neurosci. Methods 2019, 319, 60–68. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shults, N.V.; Seeherman, S.; Sariipek, N.E.; Rybka, V.; Marcocci, L.; Gychka, S.G.; Ibrahim, Y.F.; Suzuki, Y.J. Tau Protein in Lung Smooth Muscle Cells. J. Respir. 2021, 1, 30-39. https://0-doi-org.brum.beds.ac.uk/10.3390/jor1010003
Shults NV, Seeherman S, Sariipek NE, Rybka V, Marcocci L, Gychka SG, Ibrahim YF, Suzuki YJ. Tau Protein in Lung Smooth Muscle Cells. Journal of Respiration. 2021; 1(1):30-39. https://0-doi-org.brum.beds.ac.uk/10.3390/jor1010003
Chicago/Turabian StyleShults, Nataliia V., Sarah Seeherman, Nurefsan E. Sariipek, Vladyslava Rybka, Lucia Marcocci, Sergiy G. Gychka, Yasmine F. Ibrahim, and Yuichiro J. Suzuki. 2021. "Tau Protein in Lung Smooth Muscle Cells" Journal of Respiration 1, no. 1: 30-39. https://0-doi-org.brum.beds.ac.uk/10.3390/jor1010003