
Structural and Functional Thymic Biomarkers Are Involved in the Pathogenesis of Thymic Epithelial Tumors: An Overview


1
Department of Pathology, IRCCS Regina Elena National Cancer Institute, 00144 Rome, Italy
2
Department of Pathology, San Camillo-Forlanini Hospital, 00152 Rome, Italy
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Immuno 2022, 2(2), 408-429; https://0-doi-org.brum.beds.ac.uk/10.3390/immuno2020025
Submission received: 29 March 2022
/
Revised: 4 May 2022
/
Accepted: 9 May 2022
/
Published: 23 May 2022
Abstract
:The normal human thymus originates from the third branchial cleft as two paired anlages that descend into the thorax and fuse on the midline of the anterior–superior mediastinum. Alongside the epithelial and lymphoid components, different types of lymphoid accessory cells, stromal mesenchymal and endothelial cells migrate to, or develop in, the thymus. After reaching maximum development during early postnatal life, the human thymus decreases in size and lymphocyte output drops with age. However, thymic immunological functions persist, although they deteriorate progressively. Several major techniques were fundamental to increasing the knowledge of thymic development and function during embryogenesis, postnatal and adult life; these include immunohistochemistry, immunofluorescence, flow cytometry, in vitro colony assays, transplantation in mice models, fetal organ cultures (FTOC), re-aggregated thymic organ cultures (RTOC), and whole-organ thymic scaffolds. The thymic morphological and functional characterization, first performed in the mouse, was then extended to humans. The purpose of this overview is to provide a report on selected structural and functional biomarkers of thymic epithelial cells (TEC) involved in thymus development and lymphoid cell maturation, and on the historical aspects of their characterization, with particular attention being paid to biomarkers also involved in Thymic Epithelial Tumor (TET) pathogenesis. Moreover, a short overview of targeted therapies in TET, based on currently available experimental and clinical data and on potential future advances will be proposed.
1. Introduction
Complex organogenetic events lead to the development and maturation of the thymus. The human thymus originates from the third branchial clefts, as two paired anlages which descend into the thorax and fuse on the midline of the anterior–superior mediastinum. Along with the epithelial and lymphoid components, different types of lymphoid accessory cells, stromal mesenchymal and endothelial cells, migrate to or develop in the thymus [1]. As is well known, the human thymus reaches maximum development during early postnatal life, and then decreases in size, and lymphocyte output drops during young adulthood and throughout adult life. However, the functions of this primary lymphatic organ persist, although the immune system deteriorates with age [2,3]. The thymic morphological and functional characterization, first performed in the mouse, was then extended to humans. Several major techniques were fundamental to increasing the knowledge of thymic development and function during embryogenesis and postnatal life; these include immunohistochemistry, immunofluorescence, flow cytometry, in vitro colony assays, transplantation in mice models, fetal organ cultures (FTOC), re-aggregated thymic organ cultures (RTOC), and whole-organ thymic scaffolds [4,5,6,7,8,9,10,11]. We deal here first with a short overview of thymic development and, after a historical report regarding how the clinical observations led to the first insights into thymic defects, we report briefly on selected epithelial cell (EC) genes/transcription factors/pathways (such as Foxn1, CD205, Notch, keratins, Autoimmune Regulator transcription factor-AIRE-, and compartment specific antibodies) which were extensively explored in humans, as well as in mouse models. In addition, we also investigated the biomarkers in Thymic Epithelial Tumors (TET) and we report on the main laboratory findings of human TET-focused investigations on the same biomarkers. Moreover, we mention some recently identified biomarkers, such as the Pou domain, class 2, transcription factor 3 (POU2F3); and the main marker of a thymic-tuft cell (TTC) signature, which possibly indicates a further thymic epithelial cell (TEC) subtype of still largely unexplored significance. Lastly, we provide a short overview of currently available experimental and preclinical studies and a further overview of clinical trials testing targeted therapies. Old and recent experimental approaches have led to substantial progress in our knowledge of thymic development and to prospective advances in treatment of immune disfunctions and of TET.
2. A Short Overview of Thymic Development and Cellular Types
The human thymus derives from the 3rd pharyngeal pouches (PP). The development of the thymic microenvironment includes a series of molecular, morphological, phenotypical, and architectural changes, which lead to the establishment of the two main thymic compartments, cortex (C) and medulla (M); the changes also lead to the appearance of perivascular spaces (PVS) at the corticomedullary junction and the development of Hassall’s bodies/corpuscles (HCs) in the medulla. Molecular changes in this evolving process were studied extensively in mice, in mutant mice models, and more recently in humans through a variety of laboratory approaches, and are extensively reported in specialized literature [9,11,12,13,14,15,16]. These observations highlighted that mouse thymic developmental processes are conserved in humans. Several transcription factors (TBX1, HOXA3, EYA1, PAX1 and -9, SIX1 and-4) (not further mentioned here) are involved, before the forkhead transcription factor (Foxn1 gene) is first activated, in the early phases of development of the 3rd PP, thymic domain (TD) [17,18]. Early in embryogenesis, in fact, Foxn1, which is considered the master transcriptional regulator of TEC, is already present in the 3rd PP. Previously, the 3rd and the 4th PP were thought to be involved in the thymic development in humans; however, Foxn1 is only found in the 3rd PP [12], whereas both the 3rd and 4th PPs show expression of GCM2 (a member of the Glial Cells Missing- Gcm- transcription factor family), a specific factor of parathyroids [19]. At this stage, TEC with an undifferentiated phenotype, of endodermal origin (TEC progenitors—TECP), including bipotent progenitor cells, are located in the 3rd PP [20]. Later, in the migrated TD, cortical and medullary thymic epithelial cells (cTEC and mTEC), under Foxn1 input, form a cellular network, differentiate and, with the lymphoid cells, constitute different anatomical and functional microenvironments (Figure 1A–C). The cortex (Figure 1A,C) becomes the site of positive/negative selection of lymphocytes, whereas the medulla (Figure 1A–C) is the site where central tolerance is established [13,21,22]. In both humans and mice, thymocyte development occurs through the subsequent double negative (DN) CD4-CD8-stage, double positive (DP) CD4+CD8+ stage, and CD4+CD8− CD3+ or CD8+CD4−CD3+ (single positive, SP) stages [23]. More precisely, cTECs are involved in positive thymocyte selection, acting as Antigen-presenting cells (APC) by expressing self-peptides, presented by MHC molecules class I and II to DP thymocytes. On the other hand, mTEC together with dendritic cells (DCs) establish central tolerance by mediating the negative selection of autoreactive T cells. This process includes the expression by mTEC of the autoimmune regulator gene (AIRE) (Figure 1B). Moreover, mTEC promote the development of CD4+CD25+Foxp3+ (forkhead box P3) regulatory T cells (TReg cells) and of natural killer (NK) T cells [13,17]. During thymic ontogenesis, the Hassall’s Bodies/Corpuscles (HCs) appear in the M as concentric epithelial swirls (Figure 1A–C) when lymphopoiesis is already established. HCs, first described in the human thymus and later in the mouse, are structurally composed of mTEC. They show cornification similar to epidermal keratinocytes and, therefore, express CK1 (keratin1). Further markers include CKAE1/AE3 (Figure 2B), CK10 (Figure 2C), involucrin, Desmoglein3 (Dsg3), and tight-junction components. Additionally, HC-mTEC strongly express Cld3/4 (claudin 3/4), a marker of Aire+ mTEC. HCs, therefore, represent terminally differentiated Aire+ mTEC and are thought to play a role in thymocyte maturation. In fact, they promote the constitutive activation of neutrophils and plasmacytoid dendritic cells (pDC) bringing to the production of IFNα [24].
The presence of other intrathymic cell types, including mesenchymal, vascular (endothelial), and lymphoid cells, was already detected in the human thymus from mid-week 8 gestational age [6]. Mesenchymal derived signals, by direct and indirect interplay via TEC with thymocytes in development, play a role in thymic organogenesis, in the development/maintenance of epithelial stroma, and in thymocyte development and maturation [25]. Moreover, the corticomedullary junction (CMJ), the third thymus functional and anatomical compartment and a major transit pathway, includes perivascular spaces (PVS), which allow lymphoid progenitor cells to migrate into the thymus and mature T cells to emigrate from the thymus [7,15]. At the CMJ, PVS are delimited by a continuous layer of TEC (Figure 1A and Figure 2A,D). In the medulla, moreover, CD4+ and CD8+ single positive (SP) thymocytes acquire the ability to proliferate in response to T-cell Receptor (TCR) stimulation. They undergo licensing for cytokine production, and finally leave the thymus at the CMJ as Recent Thymus Emigrants (RTEs) [25].
Table 1 shows a summary of the major gene/transcription factors involved in thymic development and in the pathogenesis of autoimmune disease in TET, and (some of) their roles and available clinical characteristics (taken from human and animal studies).
3. Immunodeficiency as a Clue to Understanding Thymic Development and Functions
In the characterization of genes/pathways that are relevant for thymic function, several approaches led to the discovery of subsequent steps in thymic development. Historically, the clinical syndromes of immunodeficiency provided the first insights into thymic defects. In 1968, Angelo M. DiGeorge (an Italian-American pediatric endocrinologist) and Howard W. Lischner observed a congenital absence of thymus and parathyroids in several children; this was often associated with other malformations, such as hypoplasia of the mandible or of the aortic arc, and which resulted in heart disease, and T cell defects. These alterations appeared to be embryologically related [57] and were collectively called DiGeorge syndrome (DGS). Children affected by DGS were also frequently affected by psychiatric disorders and by feeding difficulties. Initially the disease was also defined as 3rd and 4th PP syndrome, given that the organs derived from these embryonal structures are affected. The DGS spectrum includes cases of complete DGS with athymia (1% of cases), characterized by Severe Combined Immunodeficiency (SCID)-like features, as well as partial DGS [27]. DGS is the most common chromosomal microdeletion syndrome in humans, and it affects approximately 0.25 in 1000 births [35,45]. DGS is characterized by a hemizygous deletion of chromosome 22q11 in 35 to 90% of patients. In a minority of patients, there is instead a deletion of 10p13-14. The syndrome has several degrees of severity, which correlate with variability in thymus size (residual thymic tissue may be present) [40] and is associated with immunodeficiency and T-cell lymphopenia in 75–80% of patients. The term “complete DiGeorge syndrome” is sometimes still utilized by immunologists to indicate cases with a complete absence of T cells. However, nowadays SCIDs are classified according to either the main pathway/molecular defect or on the basis of the specific immunological phenotype. The historical nomenclature is reserved for cases with no recognizable genetic cause of the phenotype [46,51]. However, the DGS phenotype was identified in cases associated with several different genetic and also with environmental (toxic) factors. Among the former, there are mutations in chromodomain helicase DNA-binding 7 (CHD7), which cause CHARGE syndrome (association of Coloboma, Heart defects, Atresia choanae, Retarded growth/development, Genital and Ear anomalies/deafness) [52]. Maternal diabetes was also identified, as well as fetal toxic conditions, such as retinoic acid deficiency [51,58]. A new classification schema of congenital immunodeficiencies was recently proposed [20], based on the occurrence of inborn errors occurring in human thymic stromal cells—including TEC, together with mesenchymal and vascular cells—or due to intrinsic hematopoietic cell defects. In the former group, molecular aberrations in the 3rd PP and/or TEC development and functions are at the basis of inborn errors in stromal cell defects, which are associated with the early onset of autoimmune disease and with T cell immunodeficiency. Thymic hypoplasia or aplasia is also associated. In some forms of SCID, however, the hematopoietic cells have an intrinsic defect, which impairs TEC differentiation/function and T cell development, due to a lack of crosstalk signals among thymocytes [28,29,30]. Among these conditions, there are genetic deficiencies (not further mentioned here) in RAG1/2, AK2, IL2RG, CD3D, ADA, RMRP, and ZAP70, which induce aberrant thymic architecture, impairment of TEC differentiation and AIRE expression, and reduced DC and FOXP3+ T regulatory cells [30,61,62]. They also cause autoimmune diseases [20,30]. The reader is advised to refer to several relevant already-cited literature sources for more information on these conditions.
4. Selected Developmental and Functional Genes in the Thymus
4.1. On the Role of Foxn1
Foxn1, located on chromosome 17q11.2 (Table 1), is a member of the Fox gene family, which comprises different winged-helix transcription factors implicated in development, metabolism, cancer, and aging [31]. Originally named winged-helix nude (Whn), Foxn1 deficiency was identified as responsible in both mice and humans for the Nude/SCID phenotype [32,51]. During fetal life, Foxn1 is expressed in epithelial cells, including liver, lung, intestine, kidney, and the urinary tract, whereas in postnatal life, Foxn1 expression is restricted to keratinocytes and to TEC [32]. From the 8th gestational week, Foxn1, in the migrated TD, drives the appearance of cTEC and mTEC from TEC with undifferentiated phenotypes, including bipotent progenitor cells (TECP) [33,34]. At this time, the first lymphoid precursors are detectable, and fundamental lympho–stromal crosstalk is established, mostly under the influence of Foxn1 [29]. A role is played also by the FOXN1 target genes CCL25 and CXCL12, which encode chemokines involved in the colonization of the thymus by lymphoid progenitors, in antigen processing and thymocyte selection [36] and by the Delta-like4 Notch ligand (Dll4), expressed by cTEC in the postnatal thymus, and necessary for progenitors to commit to T cell lineage [20]. Within the epidermis, hair follicles and nail beds, Foxn1 also plays a role in the regulation of terminal differentiation [37].
4.2. CD205 Is Foxn1-Related
DEC-205, also known as DEC205 or CD205, is a marker of cTEC in the adult thymus [28,38]. It is an integral membrane protein homologous to the macrophage mannose receptor and related receptors, which are able to bind carbohydrates and to mediate endocytosis [39]. It is an endocytic receptor, a type I C-type lectin-like molecule expressed at high level by cTEC, in the thymic medullary DC (CD11c+CD8+), and in subsets of peripheral DC [41]. It acts via a Foxn1-dependent mechanism and may provide a mechanism for the uptake and presentation of self-antigens for intrathymic and peripheral tolerance induction [41]. Moreover, a TECP population was found to simultaneously express the cTEC marker CD205 and the mTEC regulator Receptor Activator of NF-κB (RANK), therefore pointing to a role of CD205+ progenitors in the generation of mTEC [42].
4.3. Notch1
The Notch pathway is involved in embryogenesis and cellular differentiation [43] and is a major determinant of T cell lineage commitment and differentiation [23,44]. Notch1 is a gene which encodes a transmembrane receptor that regulates normal T cell development. In mammals, the Notch family members include five ligands, two from the Jagged family and three from the Delta-like family (Dll1, Dll3 and Dll4), and four receptors (Notch1-4). Among the Notch ligands, Dll4, in mice, promotes Notch1-dependent T-cell fate specification and maturation [47]. The Notch pathway is activated through the interaction between two adjacent cells, one cell carrying a ligand and the second adjacent carrying a receptor programmed to combine with the ligand. Notch1 was also shown to be involved in hematological malignancies and solid tumors, and it may act, according to the malignancy, as an oncogene or tumor suppressor, depending on the context [48]. The role of Notch in the normal thymic epithelium has yet to be completely clarified, although TEC show an activated Notch pathway [49]. Weak Notch-signaling is necessary to commit TECP towards the mTECP, which are also capable of self-renewal [50]. The expansion of early T-cell progenitors driven by cTEC occurs via the expression of Notch ligand Dll4 [53] and of IL-7 [54]. Based on immunohistochemistry and the quantitative confocal microscopy of fetal, postnatal and adult human and mouse thymus samples, a recent study showed that the spatial regulation of Notch ligand expression defines discrete Notch-signaling “niches” of the human fetal and postnatal thymus. Notch activation in vivo is induced in CD34+ progenitors and developing thymocytes in these particular cortical niches, thus ensuring the coordinated delivery of niche-specific Notch1 signals required for thymopoiesis [47].
4.4. Keratins: Not Only Structural Components of TEC
Keratins (CKs; K; intermediate filaments, ifs) are differentiation markers and cytoskeleton components [55], which are differentially expressed in TEC subsets. Moreover, in experimental models, it was shown that CKs are needed to maintain thymic structural integrity. In mouse models, in fact, the loss of K8/K18 alters TEC structure and organization. It also causes prominent cTEC depletion associated with increased apoptosis, along with a perturbed mTEC distribution without any change in their differentiation status [56]. Moreover, the role and distribution of keratins were strictly associated with the search of TECP and of their progressive differentiation steps towards cTEC and mTEC [56,59,60,63]. The phenotype of TECP in mice remains controversial [33,64,65,66] and only low numbers were detected using multiple antibodies and multi-color flow cytometry. In human fetal thymuses [67], before 16 weeks of gestation, cortical (cTEC) and medullary (mTEC) TEC exhibited homogenous staining for cytokeratins CK8 and CK5. After 16 weeks, a differential staining appears, with cTEC positive for CK8 and mTEC for CK5 and CK14. Interestingly, however, CK5+CK8+ TECP were reported in the fetal thymic cortex at all gestational ages, with a relatively high number from 12 to 16 weeks. However, the complex situation is evident in the controversial results: in the cultured human thymus model [9], the direct detection of TECP in human tissue sections is not currently feasible. Clinical outcome results of long-lasting immune reconstitution [68], however, suggested that TECP are likely also to be present within transplanted thymus slices [9]. In fact, the extensive study of the distribution and role of keratins in mouse and human thymus has provided variable results in the different experimental models and according to the different antibodies [9,56,69,70,71]. Recently, in a different approach, reconstituted human thymic scaffolds that form a unique 3D epithelial thymic network were used to establish a functional microenvironment which supported T cell development both ex vivo and in vivo. This approach provided insights that suggested clonogenic TEC were double positive for CK5-14 and CK8-18. Interestingly, these clonogenic TEC were capable of extensive expansion capacity in culture and showed a hybrid phenotype characterized by the co-expression of mesenchymal and epithelial markers. The use of human thymic scaffolds could contribute to a better understanding of the differentiation phases of TECP and their progeny, and to provide clues about how to treat different immunodeficiency diseases [11].
4.5. AIRE
The gene Aire, which is located in the locus 22.3 on the q-arm of chromosome 21 in humans [72], is a thymus key transcription factor, expressed in mTECs at the final maturation stage, and downregulated with age. In the mTEC, Aire regulates the processes of differentiation and expression of peripheral tissue-specific antigens but also controls intercellular interactions in the thymus. Moreover, Aire contributes to an increase in the diversity and transfer of presented antigens, thus increasing the repertoires of maturing thymocytes [50]. mTEC belonging to the antigen-presenting subtype of thymic cells (APCs) express—together with the AIRE protein—self-antigens with specific tissue-limited expression (tissue-specific antigens, TSAs), setting the basis for “promiscuous gene expression” (PGE) [73]. The role of AIRE, therefore, resides in the presentation of self-antigens in the thymus and in the establishment and maintenance of self-tolerance. AIRE enhances the capacity of the immune system to recognize self, altered-self, and non-self, and improves the specificity of adaptive immunity [74]. AIRE is mainly expressed in mouse MHCIIhigh mTECs, in a minority subset of thymic B cells, in CCR7+ PDL1--CD127+ medullary thymic Dendritic Cells (DCs), and in rare extrathymic DCs (extrathymic AIRE-expressing cells –eTACs) [75]. Moreover, FOXP3+ regulatory T cells (Tregs) develop [76] within the thymus and, by seeding the periphery, they identify autoreactive T cells escaping to negative selection [77,78]. In humans, AIRE is expressed in the nuclei of rare mTECs mainly in the vicinity of HCs (Figure 1B). Biallelic mutations in AIRE are responsible for the autoimmune polyglandular syndrome 1 type (APS1), also known as autoimmune polyendocrinopathy, an early onset multisystem autoimmune disease, which includes chronic mucocutaneous candidiasis, ectodermal dystrophy, hypoparathyroidism, and primary adrenal insufficiency (APECED) syndrome [79].
4.6. Compartment Specific Antibodies
Thymus compartment (Cortex and Medulla)-specific TEC markers include the proteasome β5t subunit (β5t), which is a component of the thymoproteasome that is expressed exclusively in cTEC, both in mice and humans [80,81,82] (Figure 3A). The thymoproteasome generates unique, optimized peptides [83] presented by major histocompatibility complex (MHC) class I molecules [84] to support positive selection, and, therefore, plays an essential role in adaptive immunity.
“Medullary” TEC markers were also identified and characterized: CD40, CK10 (Figure 2C), and Claudins (Cld family) (Figure 3C) [85,86,87]. The proliferation and differentiation of mEC is dependent on the tumor necrosis factor (TNF) superfamily cytokines, such as RANKL and CD40L, which are produced by lymphoid cells [88]. CD40 signaling controls the development of mTEC, which play a critical role in preventing autoimmunity [89]. The Claudin (Cld) family includes membrane proteins that play a crucial role in junctional complexes. In humans, Cld3+/4+ mTEC, which are scattered exclusively in the medulla, have a globular cell morphology (Figure 3C). Moreover, Cld4 is strongly expressed in mTEC around HCs, which are thought to represent terminally differentiated mEC [14,38,88]. Moreover, terminally differentiated mTEC express CK10 (Figure 2C) and involucrin [88].
4.7. Thymic Tuft Cells and POU2F3
A new insight into the heterogeneity of TEC was obtained through the identification of thymic tuft cells in mice (TTC) [90,91]. These are rare medullary TEC characterized by unique long and thick microvilli tufts) on their apical side. Discovered more than 60 years ago in other organs, their role is far from being clarified. They resemble the chemosensory tuft (‘brush’) cells first identified in mucosal tissues in the trachea and gastro-intestinal tract [92]. Miller reported that they derive from AIRE+ and AIRE—mTECs. Therefore, they are thought to be involved in specific antigen presentation, thus playing a role in preventing autoimmunity [90]. Like other tuft cells but unlike other TEC, TTC develop under the control of the Pou domain, class 2, transcription factor 3 (POU2F3), and express many other tuft cell markers, including IL-25, the protein kinase DCLK1, pro-inflammatory cyclooxygenases, which are proteins involved in acetylcholine metabolism, and taste transduction (e.g., Trpm5) [93]. On the other hand, TTC specifically express the markers, involved in antigen presentation, MHCII and CD74. Several additional roles for TTC, however, are under investigation [94]. Therefore, TTC could represent a newly recognized TEC subtype.
5. On the Role of Developmental and Functional Genes in Thymic Epithelial Tumors (TET)
The expression and significance in TET pathogenesis of genes/transcription factors (TF) involved in thymus development and function have yet to be completely clarified. Molecular analyses performed and reported in TET [95,96,97,98,99] have seldom revealed data related to these genes/pathways. We mention here mainly the genes/TF/signaling pathways involved in thymus development and function which were “explored” in TET by expression profiles, by whole genome analysis, by targeted gene panels and whole transcriptome sequencing, by RNA-seq and by morphological approaches (immunohistochemistry).
5.1. Foxn1 and CD205 in TET
An immunohistochemical study by D. Nonaka et al. [100] on TET and few samples of normal thymuses showed that Foxn1 was widely expressed as nuclear staining in all cases of type B thymoma and all but 1 case of type A thymoma, whereas its expression was generally focal in thymic carcinoma (TC) (76%). The expression of Foxn1 was identified in all cases of AB thymoma, the expression in the type A component being more heterogeneous than in the type B component. On the other hand, CD205 cytoplasmic expression in the form of coarse granular staining with membranous accentuation was strong and widespread in all cases of type B thymoma (100%), and in a majority of type A thymoma (89%), and heterogeneous with variable intensity in thymic carcinoma (TC) (59%). AB thymoma demonstrated widespread expression of CD205 in the type B component (100%), and variable expression in the type A component (94%). Neither Foxn1 nor CD205 were expressed in two cases of thymic neuroendocrine carcinoma, one case of high grade or one case of intermediate grade. According to A. Marx and colleagues (2011) [101], the finding of focal positivity for Foxn1 in TC raised the possibility that TC may be derived from a TECP. Shortly after, Weissferdt and Moran stained a series of TC with Foxn1 and Pax8. A total of 21 out of 31 cases expressed Foxn1, and 18 out of 21 cases also showed positivity to Pax8. Therefore, the combined positivity of these two markers was considered a relatively specific marker for TC, which could be useful in its differential diagnosis [102].
Some years later, in 2017, an interesting study by Y. Chen and colleagues [103] determined mRNA and protein levels by RT-qPCR and immunohistochemistry of Wnt4 (wingless-related integration site—WNT pathway) and Foxn1 in thymoma. Moreover, the authors analyzed the effect of thymoma cell apoptosis and tumor growth in nude mice after Wnt4 and Foxn1 downregulation. Thymoma cells were cultured and transfected with siRNA targeting the Wnt4, JNK, and Foxn1 genes. In addition, thymoma cells were inoculated into nude mice and the tumor growth was analyzed. Following immunohistochemistry, thymoma cells exhibited an abnormal expression of Wnt4 and FoxN1 mRNA and protein expression. The expression levels of WNT and Foxn1 increase with increasing malignancy, according to the 2015 WHO histological thymoma classification [104] and Masaoka/Masaoka–Koga stage [105,106]. In addition, the apoptosis of thymoma cells increased and tumor growth was inhibited after the downregulation of Wnt4 and Foxn1, therefore supporting a role of these genes in thymoma pathogenesis. Thus, the authors suggest exploring the possibility to treat thymoma by targeting Wnt4 and Foxn1.
5.2. Notch
In a different approach, S. Badve and colleagues [107] performed whole genome gene analysis of 34 thymoma analyzed with Partek Genomics SuiteH and Ingenuity Pathways Analysis (IPA). By correlating gene expression and clinical data, the authors identified the top cancer pathways associated with metastases. These included those related to amino acid metabolism, to cell cycle checkpoint proteins, and to Notch signaling. In fact, Notch signaling proteins showed high gene expression associated with a propensity towards metastatic disease. Moreover, JW Riess and colleagues [108] investigated a Tissue Micro Array (TMA) containing 68 TET and eight benign thymuses by fluorescent immunohistochemistry (AQUA) for insight into the activity of the sonic hedgehog, Notch and WNT pathways. The authors hypothesized that TC in particular (many of which have squamous differentiation) would overexpress Notch. In fact, Notch mutations and overexpression were previously reported in squamous cell lung and head and neck carcinomas [109,110]. However, in the reported study [108], the TMA included only three TC, and only the protein expression of Notch was tested, precluding any definitive analysis. Interestingly, in the Cancer Genome Atlas study of TET (TCGA-THYM) by the National Cancer Institute of the United States (NCI-US) and its associated worldwide clinical and research network, RNA-seq of 117 TET cases identified a higher expression of genes involved in cell morphogenesis, receptor tyrosine kinases, retinoic acid receptors, neuronal processes, as well as the WNT and SHH signaling pathways in GTF2I mutant tumors [111]. Moreover, in a recent study, Liang and colleagues performed a targeted gene panel sequencing and whole transcriptome sequencing of tumor tissue specimens from 27 Chinese TET patients, including 24 thymomas (A, AB, and B subtypes) and 3 thymic squamous cell carcinomas (TSQCCs) [112], and compared the results also with the TCGA analysis. They reported, from RNA-seq results, that genes related to thymus development (FGF7, FGF10 and CLDN4) were highly expressed in certain TET subtypes (including A-AB-B2 and B3 cases), suggesting that the developmental process of the thymus might be linked to the pathogenesis of these TET subtypes. A further report from P. Vodicka and colleagues [113], who investigated a series of 112 thymomas by mRNA expression analysis, identified activated Wnt pathways in thymoma. Moreover, the levels of mRNA expression corresponded with aggressive behavior of type B thymomas (increasing from B1 to B3) and with the Masaoka–Koga stage. It is well known that Wnt signaling is an important regulatory pathway in TEC developmental processes, acting by regulating Foxn1 expression. In addition, growing data indicate that, in particular, Wnt4 induction of both classical (canonical) and non-classical Wnt signaling plays an essential role in maintaining the thymic microenvironment. In the adult thymus, Wnt4, predominantly produced by mTEC and cTEC [114], controls thymopoiesis and thymus size by regulating TEC, thymocytes and their progenitor proliferation [115]. On the other hand, aberrant activation of Wnt/β-catenin signaling drives oncogenic transformation in a wide range of cancers, indicating this key pathway modulator as a possible and attractive therapeutic target in malignancies [116].
5.3. Keratins
Due to their role as differentiation markers for histogenetic and classification studies, cytokeratins were also investigated in the thymus and in TET. I. Fukai and colleagues [117], through IHC on frozen tumor tissue sections from 81 thymomas and 14 TC, found a significant heterogeneity in the expression of different cytokeratin subfamilies. However, a quantitative difference in CK13 expression in thymomas and a prevalence of CK18 expression in TC were also reported. Five pediatric thymuses were also investigated: the TEC network expressed CK8 and CK19. K8.12 monoclonal (mc) antibody, specific for the CK pair 13/16, labeled subcapsular-cortical and medullary TEC and recognized some intracortical subsets; 2D7, a specific antibody for CK13, labeled a subset of subcapsular-cortical TEC and labeled most of the mTEC. RGE53, which is specific for CK18, labeled only a discrete mTEC subset. Some years later, T-t Kuo [118], in formalin fixed, paraffin-embedded tissues (FFPE) of nine nonneoplastic thymuses (three pediatric and six from myasthenia gravis—MG—patients) found that TEC of all thymic compartments were CK19 positive; CK20, CK7, and CK14 were expressed by the subcapsular TEC, mEC, and HCs, but not by the cTEC. Only HCs consistently expressed CK13 and CK10; CK8 and CK18 were expressed by mTEC and HCs. Differences in EC staining among the outer and the inner part of the HCs were found [118]. In the same study, T-t. Kuo investigated the cytokeratin distribution, on FFPE material, in a series of 34 thymomas of different subtypes and found different CK profiles for these thymomas, classified according to their own working classification, and also compared with the (at that time) first WHO classification [119] including the A-AB-B subtypes of thymoma [118]. The studies mentioned provided further insights into the histogenesis of various thymoma subtypes in relation to the thymic TEC according to their respective CK profiles. CK pattern complexity in TET is not surprising, and it reflects the morphological and functional heterogeneity of TEC in the thymus [14].
5.4. AIRE
Among TET, about 30–40% of thymoma are associated with myasthenia gravis (MG): these cases apparently do not express the autoimmune regulator AIRE and have reduced or absent thymic myoid cells [120]. Furthermore, neoplastic TEC variably express striational antigen epitopes, such as epitopes of titin and various AChR subunits, together with reduced levels of MHC-class II [121]. In thymoma-associated MG (TAMG), the defective properties of neoplastic TEC alter thymocyte maturation, which interferes with their positive/negative selection, thereby causing defective intratumoral generation of Treg cells [122]. It is possible for the pathogenic (autoimmune) response to begin before, but rarely also after, thymoma resection, therefore sustaining TAMG [123]. A. Marx and colleagues in 2018 [124] reported the presence of desmin (+) myoid cells and of AIRE+ TEC in (some) “medullary islands” (MIs) in 50% of cases of B1 thymoma. The abnormalities observed could contribute to the complex pathogenesis of TAMG and were extensively discussed in a recent (2021) review by A. Marx and colleagues [75]. It is worthy of note that the absence or reduction of AIRE in TAMG represents a model with which to study AIRE-deficient thymopoiesis in humans, similar (but not identical) to the already mentioned genetic defect-associated disease APECED/APS1 [125]. As an additional approach, spontaneous and genetically engineered thymoma models in different animals and the transplantation of human/murine thymoma tissue into nude mice provide potential models to explore TAMG pathogenesis [126].
5.5. Compartment-Specific Antibodies
An immunohistochemical characterization of TET by compartment-specific antibodies was performed by several authors. Yamada and colleagues [127] investigated the histological diversity of type AB thymoma and found that such heterogeneity correlated with the expression of the functional cortical marker β5-thymoproteasome and an abundance of TdT-positive lymphocytes. Hayashi and colleagues [128] showed that the use, in a panel of biomarkers, of corticomedullary markers, such as β5-thymoproteasome, could be useful in the differential diagnosis of B3 thymoma versus TSQCCs.
Ströbel and colleagues reported 57 thymomas studied by IHC on FFPE specimens [87]. The cortical markers used were β5T (Figure 3B), PRSS16, and cathepsin V; the medullary markers were CD40, claudin-4 (Figure 3D), AIRE, and desmin; and involucrin and CK 10 were used as markers of terminal mTEC maturation. Irrespective of their histological subtype, most thymoma contained distinct areas with cortical and medullary differentiation. Type B1, type B2 and type AB thymoma showed marked bi-lineage differentiation, with a lack of terminal mTEC maturation in type AB. Type AB thymoma were unique as they showed areas where cells with either cortical or medullary differentiation were intimately ‘mixed’ at single-cell level. Type B3 and type A thymomas showed only abortive lineage differentiation and maturation. The authors concluded that thymomas show highly characteristic patterns of bi-lineage TEC differentiation that reflect the histological subtypes recognized by the WHO classification (WHO 2004) [129]. Moreover, the patterns observed suggest that thymoma arise from thymic precursors with a cortical/medullary maturation defect [87].
5.6. A TUFT Cell Signature in TET?
Y.Yamada and colleagues [130] recently reported that most TSQCCs strongly express tuft cell markers, including the tuft cell master regulator, POU2F3. In a more recent series of 94 thymomas from an Asian cohort, Yamada used immunohistochemistry and found that approximately half of subtypes A, AB, and B1 thymomas contain small numbers of TEC expressing POU2F3, while most types B2 and B3 thymomas are negative for this marker. At variance, the tuft cell marker L1CAM, expressed in 6/17 (35%) of their TSQCCs series, was never seen to be positive in thymomas [131]. The presence of scattered POU2F3(+) cells in types A and AB thymoma suggested a variable degree of medullary differentiation, as the historical hypothesis of the medullary differentiation of “spindle cell” thymomas (now named “type A”) suggested [132].
6. Targeted Therapies in TET: A Short Overview of Current Available Experimental and Clinical Data and of Perspectives Advances
In recent years, targeted therapies were increasingly applied to several cancer types. The infrequency of actionable mutations as demonstrated by the TCGA–THYM study [111], however, caused difficulties for the development of targeted therapies in TET. In particular, a preclinical/clinical targeting of the gene/transcription factors discussed earlier in the paper was rarely “explored” until now. Among the experimental models in cancer research, it is worth noting that the athymic nude mice, a murine strain bearing spontaneous loss-of-function of the Foxn1 gene (deletion) that causes a deteriorated or absent thymus, represents a widely used tool for the preclinical testing of anticancer drugs [133]; this suggests that this gene plays a crucial role in the genetic control of growth in several cancer types. Recently, patient derived xenograft (PDX) models were introduced in cancer research, since they mimic the microenvironment and primary tumor behavior in response to chemotherapeutics and provide insights into the efficacy of drug testing [134]. PDX can also be transplanted in the previously cited athymic nude mice. However, significant data on the use of PDX in TET are rare. Nevertheless, two xenograft tumor models (MP57 and T1889) were treated with Selinexor, an inhibitor of Exportin (XPO1), as described later [135].
In the meantime, however, an increasing amount of data is being collected, which come from experimental and preclinical studies and a limited number of clinical trials in TET. Here we provide an overview of the available in vitro cell systems and of preclinical and clinical studies in TET. It is worth noting that the molecular characterization of thymoma is hampered by the specific biological characteristics and the (variable) lymphocytic component of TET, and of thymoma in particular. To overcome this problem, Maury and colleagues [136] established primary cell cultures of TET and explored the activation of the Akt/mTOR pathway. The authors demonstrated the activation of Akt and mTOR in thymoma and the expression in all thymoma subtypes of Phospho-P70S6K (mTORC1 target). Moreover, they showed the inhibitory effects of rapamycin on thymoma EC proliferation. Selected clinical trials through the use of targeted therapies in TET were reviewed by Rajan and Zhao [137]: among a limited number of target agents, sunitinib [138] and everolimus [139,140] were used with some positive results in clinical trials. For the multitargeted tyrosine kinase inhibitor sunitinib, “CUSTOM” was the first completed “basket” clinical trial, testing the effects of targeted agents against specific molecular aberrations in advanced thoracic tumors of different histologies. In the sunitinib arm, one partial response was observed in a patient with a thymic malignancy [141]. In the phase II trial in advanced TET already mentioned, PDGFRA, FLT3, and KIT somatic variations were investigated and identified in 62% of patients with TC. However, the somatic variations were not predictive of response to sunitinib, and no c-KIT mutations were identified in the patients with response [138]. The mTOR inhibitor, everolimus, was used in a phase II trial of advanced TET cases which included an NGS molecular characterization of tumor tissue. Of note, among several pathogenic mutations found by the NGS study, a Notch1 variant of uncertain significance was described in a B1 thymoma. Patients with previously treated advanced TETs appeared to benefit from everolimus in the trial; however, no association between detectable tumor mutations and time-to-treatment failure was observed [140].
As far as epigenetic alterations in TET are concerned, Y. Wang and colleagues in 2014 showed that mutations of epigenetic regulatory genes are common in TC [142]. Moreover, in a phase I trial on solid tumors, the pan-histone deacetylases inhibitor (HDACi) belinostat, a hydroxamic acid HDAC inhibitor with potent antiproliferative and HDAC inhibitory activities in vitro, determined a minor response in a patient with metastatic thymoma [143]. Based on the first results, a phase II study of belinostat was performed and demonstrated limited activity in TET by the histone deacetylase inhibitor belinostat. Among the 41 patients enrolled (25 thymoma and 16 TC), two thymoma had partial remission (PR), whereas the others had stable disease (SD). No responses were observed in the TC group [144]. A phase I/II study on belinostat in combination with chemotherapy (PAC regimen) as a first-line therapy was subsequently performed, indicating that the combination is active and feasible in TET [145].
The role of miRNAs as oncogenes or tumor suppressor genes was recently investigated in TET. Radovich and colleagues found overexpression of a large miRNA cluster on chromosome 19q13.42 (C19MC) in type A and AB thymomas (absent in type B thymoma) resulting in activation of the PI3K/AKT pathway [146]. Enkner and colleagues also observed overexpression of C19MC in type A thymomas and absence of expression in TC. In addition, they found differential expression of a miRNA cluster on chromosome 14q32 (C14MC) between type A thymoma and TC with significant down regulation in TC, pointing to the possibility that the downregulation of C14MC could contribute to the pathogenesis of TC [99]. Based on previous findings related to mature MicroRNA [147], Bellissimo and colleagues showed that, in the thymic carcinoma cell line TC 1899, HDAC inhibition promoted miRNA-145-5p expression, reduced tumor phenotype, and sensitized neoplastic EC to chemotherapy and to epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor. Moreover, after Valproid Acid (VPA) treatment of TC1889 cells, the authors showed up-regulation of miRNA-145-5p and concomitant down-regulation of miRNA-145-5p target genes. Therefore, the antitumor effects of VPA were indicated by the induction of cell-cycle arrest and by the reduction in cell viability, colony-forming ability and migration capability [148].
As far as long non-coding RNAs (lncRNAs) are concerned, it is well known that they have a regulatory capacity acting as chromatin regulators, and as important cis and trans-acting modulators for the expression of protein-coding genes. Furthermore, they can mediate epigenetic modification by recruiting chromatin-remodelling complex to a specific chromatin locus. Tito and colleagues [149] recently showed that LINC00174 up-regulated and positively correlated with a 5-gene prognostic signature (including UBAC1, FEM1B, CCDC30, SYBU, and SCD5 genes) in TET; among these five prognostic gene signature positively correlated to LINC00174, and predicted to be targeted by miR-145-5p, the SCD5 gene (steroyl-Codesaturase) is involved in lipid metabolism, and specifically implicated in unsaturated fatty acids synthesis. SCD5 in TC1889 cells increased the amount of lipid droplets as well as cell migration. Moreover, J. Wang and colleagues [150] found that the LncRNA LOXL1-AS1 affects thymoma progression by the miR-525-5p-HSPA9 axis; the latter is overexpressed in TET and associated with a poor prognosis. In vitro experiments with the cell lines Ty-82 (derived from a metastatic undifferentiated TC) and Thy0517 cells (derived from AB thymoma) showed that downregulation of miR-525-5p promoted cell invasion. LOXL1-AS1 promoted HSPA9 expression as a sponge, targeting miR-525-5p. HSPA9 (mortalin) is a member of the heat shock protein (HSP) 70 family, which is mainly found in mitochondria and plays an important role in regulating cell growth and survival.
Recently, new data were obtained from a study based on genomic-clustering analysis of the TCGA data and on the use of the IU-TAB-1 cell line, derived from an AB thymoma [151]. The authors identified six molecular subtypes (TH1 to TH6) of TET, irrespective of the WHO classification, and the IU-TAB-1 cell line clustered into the TH4 molecular subtype. The cell line was found to be resistant to everolimus (mTORC1 inhibitor) and sensitive to nelfinavir (AKT1 inhibitor) across the endpoints considered. The resistance to everolimus was driven by upregulation of downstream signaling of KIT, PDGFRA, and PDGFB in the presence of mTORC1 inhibition, whereas the sensitivity to nelfinavir was due to the IU-TAB-1 cell line’s (characteristic) gain-of-function (GOF) mutation in PIK3CA and amplification of genes observed by array comparative genomic hybridization (aCGH), including AURKA, ERBB2, KIT, PDGFRA and PDGFB, the latter known to be an upregulator of AKT. Other in vitro candidates, tested for TH4 subtype in the IU-TAB-1 cell line, included panobinostat (histone deacetylase [HDAC] inhibitor) and bortezomib (proteasome inhibitor). Other TET cell lines at present available were reported in a recent paper dealing with the growing body of epigenetic studies in TET [152].
Among new targets to study, mesothelin and exportin1 (XPO1) represent examples of putative biologic targets expressed in TET. Mesothelin is a cell-surface glycoprotein, which is highly expressed in most TC [153]. Among several drugs being developed to target mesothelin, anetumab, an antibody–drug conjugate, is undergoing evaluation in patients with advanced TC (NCT03102320) [154]. XPO1, instead, is involved in nuclear export of tumor-suppressor proteins, including p53; the inhibition of XPO1 in TET cells was shown to induce anti-tumor activity in vitro and to suppress TET xenograft tumor growth. Moreover, the authors demonstrated that p53 loss and XPO1 amplification are involved in resistance to selinexor [135]. Selinexor (KPT-330), a small-molecule inhibitor of XPO1, is under evaluation in a phase II trial in patients with advanced TETs (NCT03193437) [137]. Five Asian patients with advanced or metastatic TET were treated in a phase I study of selinexor (SINE) [155].
New possibilities are now also being offered by the identification in the thymus and in TET of TTC and of their controller POU2F3 [130,131]. A study on the transcription factor POU2F3 recently identified a subset of small cell lung carcinoma (SCLC) having POU2F3 as master regulator of cell identity, in association with low or absent expression of neuroendocrine (NE) markers. Huang and colleagues reported that POU2F3high SCLC tumors and cell lines lack the expression of classical neuroendocrine lineage markers and instead express markers of the tuft cell lineage. POU2F3High cell lines are also available to test pathogenetic mechanisms and new therapeutic strategies in SCLC [156,157]. A molecular stratification of SCLC could correlate with therapeutic responsiveness in preclinical and clinical studies. [158]. A new field was, therefore, opened to investigate the molecular and clinical significance of a TTF signature also in TET.
7. Conclusions
From our overview, it appears that considerable evidence points to a role of developmental and functional genes in the pathogenesis of TET, as reported in thymic tumor specimen-based studies, by IHC and GEP, as well as by the multi-omics study TCGA–THYM, by targeted gene panels and whole transcriptome sequencing. Further insights into the pathogenesis of TET derived from data provided by thymoma EC primary cultures and by TET cell lines targeted by novel molecules/novel experimental therapeutic strategies. As mentioned, novel biomarkers are under investigation; however, the underlying pathogenetic mechanisms of TET development remain far from being clarified and further approaches will be needed in the future. From the complex observations reported in this overview, it is possible that, in the future, TET could be stratified not only on the basis of morphological features/classification, but also according to molecular subtypes and signaling networks. Moreover, the recent identification of thymic tuft cells once more points to the morphological and functional heterogeneity of TEC in the thymus and to their possible role as a source of tumors. Furthermore, without doubt, the previously cited new experimental approaches to thymic development, such as the reconstruction of the human thymus, including clonogenic TEC characterization, will also contribute to better understand the complex interplay among the different TEC and the mesenchymal and lymphoid thymic constituents in normal and diseased conditions.
Author Contributions
Conceptualization, all authors.; methodology, M.M.; software, M.M., E.G.; validation all authors.; formal analysis, M.M.; investigation, all authors.; resources, M.T.R.; data curation, E.G., M.T.R.; writing—original draft preparation, M.M.; writing—review and editing, M.M., E.G.; visualization all authors.; supervision M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors wish to thank Paola Bonfanti for her reading of the manuscript and Michael Kenyon for his review of the English language. This review article is dedicated to HK Müller-Hermelink, Würzburg, Germany for his long-lasting encouragement in the study of the thymus and of thymic epithelial tumors.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
EC: epithelial cells; TEC: thymic epithelial cells; C: cortex; M: medulla; HC: Hassall’s corpuscle; cTEC: cortical epithelial cells; mTEC: medullary epithelial cells; CMJ: corticomedullary junction; PVS: perivascular spaces; TET: thymic epithelial tumors; TD: thymic domain; PP: pharyngeal pouches; FTOC: fetal organ cultures; RTOC: re-aggregated thymic organ cultures; TECP: thymic epithelial progenitor cells; AIRE: autoimmune regulator transcription factor; eTACs: extathymic AIRE-expressing cells; DP: double positive; SP: single positive; TReg: regulatory T cells; RTE: recent thymic emigrants; POU2F3: Pou domain, class 2, transcription factor; TTC: thymic tuft cells; APC: antigen presenting cells; MHC: major histocompatibility complex; DC: dendritic cells; pDC: plasmacytoid DC; NK: natural killer; Foxp3: forkhead box P3; IFNα: interferon α; ADA: adenosin deaminase; TSA: tissue-specific antigens; PGE: promiscuous gene expression; DGS: diGeorge Syndrome; APS1: autoimmune polyglandular syndrome 1 type; APECED: autoimmune polyendocrinopathy, including chronic mucocutaneous candidiasis, ectodermal dystrophy, hypoparathyroidism and primary adrenal insufficiency; Cld: claudin family; TMA: tissue micro array; TCGA: the Cancer Genome Atlas; MG: myasthenia gravis; TAMG: thymoma-associated myasthenia gravis; TSQCC: thymic squamous cell carcinoma; Wnt: wingless-related integration site; TNF: tumor necrosis factor; β5t: proteasome β5t subunit; CK or K: keratins; if: intermediate filaments; IF: immunofluorescence; IHC: immunohistochemistry; Dll4: delta-like4 Notch ligand; Mls: medullary islands; GOF: gain-of-function; GEPgene expression profile; aCGH = array comparative genomic hybridization; SCID: severe combined immune deficiency; CHARGE syndrome: Coloboma, Heart defects, Atresia choanae, Retarded growth/development, Genital and Ear anomalies/deafness.
References
- Rezzani, R.; Bonomini, F.; Rodella, L.F. Histochemical and molecular overview of the thymus as site for T-cells development. Prog. Histochem. Cytochem. 2008, 43, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Aw, D.; Palmer, D.B. The origin and implication of thymic involution. Aging Dis. 2011, 2, 437–443. [Google Scholar] [PubMed]
- Gui, J.; Mustachio, L.M.; Su, D.M.; Craig, R.W. Thymus Size and Age-related Thymic Involution: Early Programming, Sexual Dimorphism, Progenitors and Stroma. Aging Dis. 2012, 3, 280–290. [Google Scholar] [PubMed]
- Masuda, K.; Itoi, M.; Amagai, T.; Minato, N.; Katsura, Y.; Kawamoto, H. Thymic anlage is colonized by progenitors restricted to T, NK, and dendritic cell lineages. J. Immunol. 2005, 174, 2525–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cupedo, T.; Nagasawa, M.; Weijer, K.; Blom, B.; Spits, H. Development and activation of regulatory T cells in the human fetus. Eur. J. Immunol. 2005, 35, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Heinly, C.S. Early human T cell development: Analysis of the human thymus at the time of initial entry of hematopoietic stem cells into the fetal thymic microenvironment. J. Exp. Med. 1995, 181, 1445–1458. [Google Scholar] [CrossRef]
- Mori, K.; Itoi, M.; Tsukamoto, N.; Kubo, H.; Amagai, T. The perivascular space as a path of hematopoietic progenitor cells and mature T cells between the blood circulation and the thymic parenchyma. Int. Immunol. 2007, 19, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Weerkamp, F.; Baert, M.R.; Brugman, M.H.; Dik, W.A.; de Haas, E.F.; Visser, T.P. Human thymus contains multipotent progenitors with T/B lymphoid, myeloid, and erythroid lineage potential. Blood 2006, 107, 3131–3137. [Google Scholar] [CrossRef]
- Hale, L.P.; Neff, J.; Cheatham, L.; Cardona, D.; Markert, M.L.; Kurtzberg, J. Histopathologic assessment of cultured human thymus. PLoS ONE 2020, 15, e0230668. [Google Scholar] [CrossRef]
- Piccinini, E.; Bonfanti, P. Disassembling and Reaggregating the Thymus: The Pros and Cons of Current Assays. Immunol. Toler. 2019, 1899, 129–142. [Google Scholar]
- Campinoti, S.; Gjinovci, A.; Ragazzini, R.; Zanieri, L.; Ariza-McNaughton, L.; Catucci, M. Reconstitution of a functional human thymus by postnatal stromal progenitor cells and natural whole-organ scaffolds. Nat. Commun. 2020, 11, 6372. [Google Scholar] [CrossRef]
- Farley, A.M.; Morris, L.X.; Vroegindeweij, E.; Depreter, M.L.; Vaidya, H.; Stenhouse, F.H. Dynamics of thymus organogenesis and colonization in early human development. Development 2013, 140, 2015–2026. [Google Scholar] [CrossRef] [Green Version]
- Alves, N.L.; Takahama, Y.; Ohigashi, I.; Ribeiro, A.R.; Baik, S.; Anderson, G. Serial progression of cortical and medullary thymic epithelial microenvironments. Eur. J. Immunol. 2014, 44, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Hamazaki, Y. Adult thymic epithelial cell (TEC) progenitors and TEC stem cells: Models and mechanisms for TEC development and maintenance. Eur. J. Immunol. 2015, 45, 2985–2993. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.; Jenkinson, W.E. Border control: Anatomical origins of the thymus medulla. Eur. J. Immunol. 2015, 45, 2203–2207. [Google Scholar] [CrossRef] [Green Version]
- Haunerdinger, V.; Moccia, M.D.; Opitz, L.; Vavassori, S.; Dave, H.; Hauri-Hohl, M.M. Novel Combination of Surface Markers for the Reliable and Comprehensive Identification of Human Thymic Epithelial Cells by Flow Cytometry: Quantitation and Transcriptional Characterization of Thymic Stroma in a Pediatric Cohort. Front. Immunol. 2021, 12, 740047. [Google Scholar] [CrossRef]
- Manley, N.R.; Richie, E.R.; Blackburn, C.C.; Condie, B.G.; Sage, J. Structure and function of the thymic microenvironment. Front. Biosci. 2011, 16, 2461–2477. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.; Manley, N.R. Mechanisms of thymus organogenesis and morphogenesis. Development 2011, 138, 3865–3878. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Farley, A.; Chen, L.; Kirby, B.J.; Kovacs, C.S.; Blackburn, C.C. Thymus-associated parathyroid hormone has two cellular origins with distinct endocrine and immunological functions. PLoS Genet. 2010, 6, e1001251. [Google Scholar] [CrossRef] [Green Version]
- Kreins, A.Y.; Maio, S.; Dhalla, F. Inborn errors of thymic stromal cell development and function. Semin. Immunopathol. 2021, 43, 85–100. [Google Scholar] [CrossRef]
- Rezzani, R.; Nardo, L.; Favero, G.; Peroni, M.; Rodella, L.F. Thymus and aging: Morphological, radiological, and functional overview. Age 2014, 36, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Yamano, T.; Nedjic, J.; Hinterberger, M.; Steinert, M.; Koser, S.; Pinto, S. Thymic B Cells Are Licensed to Present Self Antigens for Central T Cell Tolerance Induction. Immunity 2015, 42, 1048–1061. [Google Scholar] [CrossRef] [Green Version]
- Famili, F.; Wiekmeijer, A.S.; Staal, F.J. The development of T cells from stem cells in mice and humans. Future Sci. OA 2017, 3, FSO186. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sekai, M.; Matsui, T.; Fujii, Y.; Matsumoto, M.; Takeuchi, O. Hassall’s corpuscles with cellular-senescence features maintain IFNα production through neutrophils and pDC activation in the thymus. Int. Immunol. 2019, 31, 127–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, K.D.; Jenkinson, W.E.; Anderson, G. Non-Epithelial Stromal Cells in Thymus Development and Function. Front. Immunol. 2021, 12, 634367. [Google Scholar] [CrossRef] [PubMed]
- Ramieri, M.T.; Gallo, E.; Marino, M. Immunohistochemistry of Normal Thymus. In Atlas of Thymic Pathology; Jain, D., Bishop, J.A., Wick, M.R., Eds.; Springer Nature: Singapore, 2020. [Google Scholar]
- Lackey, A.E.; Muzio, M.R. DiGeorge Syndrome; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Anderson, G.; Takahama, Y. Thymic epithelial cells: Working class heroes for T cell development and repertoire selection. Trends Immunol. 2012, 33, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Lopes, N.; Sergé, A.; Ferrier, P.; Irla, M. Thymic Crosstalk Coordinates Medulla Organization and T-Cell Tolerance Induction. Front. Immunol. 2015, 6, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poliani, P.L.; Facchetti, F.; Ravanini, M.; Gennery, A.R.; Villa, A.; Roifman, C.M. Early defects in human T-cell development severely affect distribution and maturation of thymic stromal cells: Possible implications for the pathophysiology of Omenn syndrome. Blood J. Am. Soc. Hematol. 2009, 114, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, E.; Knöchel, W. Five years on the wings of fork head. Mech. Dev. 1996, 57, 3–20. [Google Scholar] [CrossRef]
- Vaidya, H.J.; Briones Leon, A.; Blackburn, C.C. FOXN1 in thymus organogenesis and development. Eur. J. Immunol. 2016, 46, 1826–1837. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.R.; Farley, A.; Blair, N.F.; Gordon, J.; Sharp, L.; Blackburn, C.C. Identification and characterization of thymic epithelial progenitor cells. Immunity 2002, 16, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.W.; Jenkinson, W.E.; Anderson, G.; Jenkinson, E.J. Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 2006, 441, 988–991. [Google Scholar] [CrossRef]
- Shprintzen, R.J. Velo-cardio-facial syndrome: 30 Years of study. Dev. Disabil. Res. Rev. 2008, 14, 3–10. [Google Scholar] [CrossRef]
- Žuklys, S.; Handel, A.; Zhanybekova, S.; Govani, F.; Keller, M.; Maio, S. Foxn1 regulates key target genes essential for T cell development in postnatal thymic epithelial cells. Nat. Immunol. 2016, 17, 1206–1215. [Google Scholar] [CrossRef]
- Lee, D.; Prowse, D.M.; Brissette, J.L. Association between mouse nude gene expression and the initiation of epithelial terminal differentiation. Dev. Biol. 1999, 208, 362–374. [Google Scholar] [CrossRef]
- Shakib, S.; Desanti, G.E.; Jenkinson, W.E.; Parnell, S.M.; Jenkinson, E.J.; Anderson, G. Checkpoints in the development of thymic cortical epithelial cells. J. Immunol. 2009, 182, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Swiggard, W.J.; Heufler, C.; Peng, M.; Mirza, A.; Steinman, R.M. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 1995, 375, 151–155. [Google Scholar] [CrossRef]
- Marcovecchio, G.E.; Bortolomai, I.; Ferrua, F.; Fontana, E.; Imberti, L.; Conforti, E. Thymic Epithelium Abnormalities in DiGeorge and Down Syndrome Patients Contribute to Dysregulation in T Cell Development. Front. Immunol. 2019, 10, 447. [Google Scholar] [CrossRef]
- Shrimpton, R.E.; Butler, M.; Morel, A.S.; Eren, E.; Hue, S.S.; Ritter, M.A. CD205 (DEC-205): A recognition receptor for apoptotic and necrotic self. Mol. Immunol. 2009, 46, 1229–1239. [Google Scholar] [CrossRef]
- Baik, S.; Jenkinson, E.J.; Lane, P.J.; Anderson, G.; Jenkinson, W.E. Generation of both cortical and Aire(+) medullary thymic epithelial compartments from CD205(+) progenitors. Eur. J. Immunol. 2013, 43, 589–594. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Roozen, P.P.; Brugman, M.H.; Staal, F.J. Differential requirements for Wnt and Notch signaling in hematopoietic versus thymic niches. Ann. N. Y. Acad. Sci. 2012, 1266, 78–93. [Google Scholar] [CrossRef]
- Goodship, J.; Cross, I.; LiLing, J.; Wren, C. A population study of chromosome 22q11 deletions in infancy. Arch. Dis. Child. 1998, 79, 348–351. [Google Scholar] [CrossRef]
- Chinen, J.; Cowan, M.J. Advances and highlights in primary immunodeficiencies in 2017. J. Allergy Clin. Immunol. 2018, 142, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Leon, M.J.; Fuentes, P.; de la Pompa, J.L.; Toribio, M.L. Dynamic regulation of NOTCH1 activation and Notch ligand expression in human thymus development. Development 2018, 145, dev165597. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, P.; Weaver, K.L.; Capobianco, A.J. Notch signalling in solid tumours: A little bit of everything but not all the time. Nat. Rev. Cancer 2011, 11, 338–351. [Google Scholar] [CrossRef]
- Laky, K.; Fowlkes, B.J. Notch signaling in CD4 and CD8 T cell development. Curr. Opin. Immunol. 2008, 20, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Shevyrev, D.; Tereshchenko, V.; Kozlov, V.; Sennikov, S. Phylogeny, Structure, Functions, and Role of AIRE in the Formation of T-Cell Subsets. Cells 2022, 11, 194. [Google Scholar] [CrossRef]
- Giardino, G.; Borzacchiello, C.; De Luca, M.; Romano, R.; Prencipe, R.; Cirillo, E. T-Cell Immunodeficiencies With Congenital Alterations of Thymic Development: Genes Implicated and Differential Immunological and Clinical Features. Front. Immunol. 2020, 11, 1837. [Google Scholar] [CrossRef]
- Bhalla, P.; Wysocki, C.A.; van Oers, N.S.C. Molecular Insights Into the Causes of Human Thymic Hypoplasia With Animal Models. Front. Immunol. 2020, 11, 830. [Google Scholar] [CrossRef]
- Hozumi, K.; Mailhos, C.; Negishi, N.; Hirano, K.; Yahata, T.; Ando, K. Delta-like 4 is indispensable in thymic environment specific for T cell development. J. Exp. Med. 2008, 205, 2507–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, N.L.; Richard-Le Goff, O.; Huntington, N.D.; Sousa, A.P.; Ribeiro, V.S.; Bordack, A. Characterization of the thymic IL-7 niche in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1512–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef] [PubMed]
- Odaka, C.; Loranger, A.; Takizawa, K.; Ouellet, M.; Tremblay, M.J.; Murata, S. Keratin 8 is required for the maintenance of architectural structure in thymus epithelium. PLoS ONE 2013, 8, e75101. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, K.E. Chromosome 22q11.2 deletion syndrome and DiGeorge syndrome. Immunol. Rev. 2019, 287, 186–201. [Google Scholar] [CrossRef]
- Zhang, L.; Zhong, T.; Wang, Y.; Jiang, Q.; Song, H.; Gui, Y. TBX1, a DiGeorge syndrome candidate gene, is inhibited by retinoic acid. Int. J. Dev. Biol. 2006, 50, 55–61. [Google Scholar] [CrossRef]
- Sekai, M.; Wang, J.; Minato, N.; Hamazaki, Y. An improved clonogenic culture method for thymic epithelial cells. J. Immunol. Methods 2019, 467, 29–36. [Google Scholar] [CrossRef]
- Alawam, A.S.; Anderson, G.; Lucas, B. Generation and Regeneration of Thymic Epithelial Cells. Front. Immunol. 2020, 11, 858. [Google Scholar] [CrossRef]
- Cavadini, P.; Vermi, W.; Facchetti, F.; Fontana, S.; Nagafuchi, S.; Mazzolari, E.; Sediva, A.; Marrella, V.; Villa, A.; Fischer, A.; et al. AIRE deficiency in thymus of 2 patients with Omenn syndrome. J Clin. Invest. 2005, 115, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Poliani, P.L.; Fontana, E.; Roifman, C.M.; Notarangelo, L.D. ζ Chain-associated protein of 70 kDa (ZAP70) deficiency in human subjects is associated with abnormalities of thymic stromal cells: Implications for T-cell tolerance. J. Allergy Clin. Immunol. 2013, 131, 597–600.e1-2. [Google Scholar] [CrossRef]
- Kreins, A.Y.; Bonfanti, P.; Davies, E.G. Current and Future Therapeutic Approaches for Thymic Stromal Cell Defects. Front. Immunol. 2021, 12, 655354. [Google Scholar] [CrossRef]
- Wong, K.; Lister, N.L.; Barsanti, M.; Lim, J.M.; Hammett, M.V.; Khong, D.M. Multilineage potential and self-renewal define an epithelial progenitor cell population in the adult thymus. Cell Rep. 2014, 8, 1198–1209. [Google Scholar] [CrossRef] [Green Version]
- Ucar, A.; Ucar, O.; Klug, P.; Matt, S.; Brunk, F.; Hofmann, T.G. Adult thymus contains FoxN1(-) epithelial stem cells that are bipotent for medullary and cortical thymic epithelial lineages. Immunity 2014, 41, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Ulyanchenko, S.; O’Neill, K.E.; Medley, T.; Farley, A.M.; Vaidya, H.J.; Cook, A.M. Identification of a Bipotent Epithelial Progenitor Population in the Adult Thymus. Cell Rep. 2016, 14, 2819–2832. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Gupta, T.; Kaur, H.; Sehgal, S.; Aggarwal, A.; Kapoor, K. Cytokeratin (CK5, CK8, CK14) expression and presence of progenitor stem cells in human fetal thymuses. Clin. Anat. 2016, 29, 711–717. [Google Scholar] [CrossRef]
- Markert, M.L.; Devlin, B.H.; Alexieff, M.J.; Li, J.; McCarthy, E.A.; Gupton, S.E. Review of 54 patients with complete DiGeorge anomaly enrolled in protocols for thymus transplantation: Outcome of 44 consecutive transplants. Blood 2007, 109, 4539–4547. [Google Scholar] [CrossRef]
- Lee, E.N.; Park, J.K.; Lee, J.R.; Oh, S.O.; Baek, S.Y.; Kim, B.S. Characterization of the expression of cytokeratins 5, 8, and 14 in mouse thymic epithelial cells during thymus regeneration following acute thymic involution. Anat. Cell Biol. 2011, 44, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Feng, Y.; Barnes, P.; Huang, F.F.; Idell, S.; Su, D.M. Deletion of FoxN1 in the thymic medullary epithelium reduces peripheral T cell responses to infection and mimics changes of aging. PLoS ONE 2012, 7, e34681. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Li, H.; Luo, H.; Zhao, Y. Thymic epithelial cell development and its dysfunction in human diseases. BioMed Res. Int. 2014, 2014, 206929. [Google Scholar] [CrossRef] [Green Version]
- Aaltonen, J.; Björses, P.; Sandkuijl, L.; Perheentupa, J.; Peltonen, L. An autosomal locus causing autoimmune disease: Autoimmune polyglandular disease type I assigned to chromosome 21. Nat. Genet. 1994, 8, 83–87. [Google Scholar] [CrossRef]
- Kyewski, B.; Derbinski, J.; Gotter, J.; Klein, L. Promiscuous gene expression and central T-cell tolerance: More than meets the eye. Trends Immunol. 2002, 23, 364–371. [Google Scholar] [CrossRef]
- Flajnik, M.F. A cold-blooded view of adaptive immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef]
- Marx, A.; Yamada, Y.; Simon-Keller, K.; Schalke, B.; Willcox, N.; Ströbel, P. Thymus and autoimmunity. Semin. Immunopathol. 2021, 43, 45–64. [Google Scholar] [CrossRef]
- Hori, S.; Normura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Anderson, M.S. Thymic tolerance as a key brake on autoimmunity. Nat. Immunol. 2018, 19, 659–664. [Google Scholar] [CrossRef]
- Pesenacker, A.M.; Cook, L.; Levings, M.K. The role of FOXP3 in autoimmunity. Curr. Opin. Immunol. 2016, 43, 16–23. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Oftedal, B.E.; Wolff, A.B.; Husebye, E.S. AIRE-mutations and autoimmune disease. Curr. Opin. Immunol. 2016, 43, 8–15. [Google Scholar] [CrossRef]
- Murata, S.; Sasaki, K.; Kishimoto, T.; Niwa, S.; Hayashi, H.; Takahama, Y. Regulation of CD8+ T cell development by thymus-specific proteasomes. Science 2007, 316, 1349–1353. [Google Scholar] [CrossRef]
- Tomaru, U.; Ishizu, A.; Murata, S.; Miyatake, Y.; Suzuki, S.; Takahashi, S. Exclusive expression of proteasome subunit β5t in the human thymic cortex. Blood 2009, 113, 5186–5191. [Google Scholar] [CrossRef] [Green Version]
- Takahama, Y.; Ohigashi, I.; Murata, S.; Tanaka, K. Thymoproteasome and peptidic self. Immunogenetics 2019, 71, 217–221. [Google Scholar] [CrossRef]
- Tomaru, U.; Konno, S.; Miyajima, S.; Kimoto, R.; Onodera, M.; Kiuchi, S. Restricted Expression of the Thymoproteasome Is Required for Thymic Selection and Peripheral Homeostasis of CD8(+) T Cells. Cell Rep. 2019, 26, 639–651.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, K.L.; Gramm, C.; Rothstein, L.; Clark, K.; Stein, R.; Dick, L. Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules. Cell 1994, 78, 761–771. [Google Scholar] [CrossRef]
- Lal-Nag, M.; Morin, P.J. The claudins. Genome Biol. 2009, 10, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekai, M.; Hamazaki, Y.; Minato, N. Medullary thymic epithelial stem cells maintain a functional thymus to ensure lifelong central T cell tolerance. Immunity 2014, 41, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Ströbel, P.; Hartmann, E.; Rosenwald, A.; Kalla, J.; Ott, G.; Friedel, G. Corticomedullary differentiation and maturational arrest in thymomas. Histopathology 2014, 64, 557–566. [Google Scholar] [CrossRef]
- Hamazaki, Y.; Sekai, M.; Minato, N. Medullary thymic epithelial stem cells: Role in thymic epithelial cell maintenance and thymic involution. Immunol. Rev. 2016, 271, 38–55. [Google Scholar] [CrossRef]
- Akiyama, T.; Shinzawa, M.; Akiyama, N. TNF receptor family signaling in the development and functions of medullary thymic epithelial cells. Front. Immunol. 2012, 3, 278. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.N.; Proekt, I.; von Moltke, J.; Wells, K.L.; Rajpurkar, A.R.; Wang, H. Thymic tuft cells promote an IL-4-enriched medulla and shape thymocyte development. Nature 2018, 559, 627–631. [Google Scholar] [CrossRef]
- Bornstein, C.; Nevo, S.; Giladi, A.; Kadouri, N.; Pouzolles, M.; Gerbe, F. Single-cell mapping of the thymic stroma identifies IL-25-producing tuft epithelial cells. Nature 2018, 559, 622–626. [Google Scholar] [CrossRef]
- Gerbe, F.; Jay, P. Intestinal tuft cells: Epithelial sentinels linking luminal cues to the immune system. Mucosal Immunol. 2016, 9, 1353–1359. [Google Scholar] [CrossRef]
- Gerbe, F.; Legraverend, C.; Jay, P. The intestinal epithelium tuft cells: Specification and function. Cell. Mol. Life Sci. 2012, 69, 2907–2917. [Google Scholar] [CrossRef] [Green Version]
- Nevo, S.; Kadouri, N.; Abramson, J. Tuft cells: From the mucosa to the thymus. Immunol. Lett. 2019, 210, 1–9. [Google Scholar] [CrossRef]
- Girard, N.; Shen, R.; Guo, T.; Zakowski, M.F.; Heguy, A.; Riely, G.J. Comprehensive genomic analysis reveals clinically relevant molecular distinctions between thymic carcinomas and thymomas. Clin. Cancer Res. 2009, 15, 6790–6799. [Google Scholar] [CrossRef] [Green Version]
- Petrini, I.; Meltzer, P.S.; Kim, I.K.; Lucchi, M.; Park, K.S.; Fontanini, G. A specific missense mutation in GTF2I occurs at high frequency in thymic epithelial tumors. Nat. Genet. 2014, 46, 844–849. [Google Scholar] [CrossRef] [Green Version]
- Rajan, A.; Girard, N.; Marx, A. State of the art of genetic alterations in thymic epithelial tumors. J. Thorac. Oncol. 2014, 9, S131–S136. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Jang, H.J.; Shah, R.; Yoon, D.; Hamaji, M.; Wald, O. Genomic Analysis of Thymic Epithelial Tumors Identifies Novel Subtypes Associated with Distinct Clinical Features. Clin. Cancer Res. 2017, 23, 4855–4864. [Google Scholar] [CrossRef] [Green Version]
- Enkner, F.; Pichlhöfer, B.; Zaharie, A.T.; Krunic, M.; Holper, T.M.; Janik, S. Molecular Profiling of Thymoma and Thymic Carcinoma: Genetic Differences and Potential Novel Therapeutic Targets. Pathol. Oncol. Res. 2017, 23, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, D.; Henley, J.D.; Chiriboga, L.; Yee, H. Diagnostic utility of thymic epithelial markers CD205 (DEC205) and Foxn1 in thymic epithelial neoplasms. Am. J. Surg. Pathol. 2007, 31, 1038–1044. [Google Scholar] [CrossRef]
- Marx, A.; Rieker, R.; Toker, A.; Länger, F.; Ströbel, P. Thymic Carcinoma: Is it a Separate Entity? From Molecular to Clinical Evidence. Thorac. Surg. Clin. 2011, 21, 25–31. [Google Scholar] [CrossRef]
- Weissferdt, A.; Moran, C.A. Thymic carcinoma, part 1: A clinicopathologic and immunohistochemical study of 65 cases. Am. J. Clin. Pathol. 2012, 138, 103–114. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Liu, Y.; Wang, Y.; Wang, H.; Lu, C. Decreased Wnt4 expression inhibits thymoma development through downregulation of FoxN1. J. Thorac. Dis. 2017, 9, 1574–1583. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. WHO Classification of Tumours of the Lung, Pleura, Thymus and Heart, 4th ed.; International Agency for Research on Cancer-IARC-Press: Lyon, France, 2015. [Google Scholar]
- Masaoka, A.; Monden, Y.; Nakahara, K.; Tanioka, T. Follow-up study of thymomas with special reference to their clinical stages. Cancer 1981, 48, 2485–2492. [Google Scholar] [CrossRef]
- Koga, K.; Matsuno, Y.; Noguchi, M.; Mukai, K.; Asamura, H.; Goya, T. A review of 79 thymomas: Modification of staging system and reappraisal of conventional division into invasive and non-invasive thymoma. Pathol. Int. 1994, 44, 359–367. [Google Scholar] [CrossRef]
- Badve, S.; Goswami, C.; Gökmen-Polar, Y.; Nelson, R.P., Jr.; Henley, J.; Miller, N. Molecular analysis of thymoma. PLoS ONE 2012, 7, e42669. [Google Scholar] [CrossRef] [Green Version]
- Riess, J.W.; West, R.; Dean, M.; Klimowicz, A.C.; Neal, J.W.; Hoang, C. GLI1, CTNNB1 and NOTCH1 protein expression in a thymic epithelial malignancy tissue microarray. Anticancer Res. 2015, 35, 669–676. [Google Scholar]
- Stransky, N.; Egloff, A.M.; Tward, A.D.; Kostic, A.D.; Cibulskis, K.; Sivachenko, A. The mutational landscape of head and neck squamous cell carcinoma. Science 2011, 333, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive genomic characterization of squamous cell lung cancers. Nature 2012, 489, 519–525. [Google Scholar] [CrossRef]
- Radovich, M.; Pickering, C.R.; Felau, I.; Ha, G.; Zhang, H.; Jo, H. The Integrated Genomic Landscape of Thymic Epithelial Tumors. Cancer Cell 2018, 33, 244–258.e10. [Google Scholar] [CrossRef] [Green Version]
- Liang, N.; Liu, L.; Huang, C.; Liu, H.; Guo, C.; Li, J. Transcriptomic and Mutational Analysis Discovering Distinct Molecular Characteristics Among Chinese Thymic Epithelial Tumor Patients. Front. Oncol. 2021, 11, 647512. [Google Scholar] [CrossRef]
- Vodicka, P.; Krskova, L.; Odintsov, I.; Krizova, L.; Sedlackova, E.; Schutzner, J. Expression of molecules of the Wnt pathway and of E-cadherin in the etiopathogenesis of human thymomas. Oncol. Lett. 2020, 19, 2413–2421. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, K.M.; Vanegas, J.R.; Brochu, S.; Shan, J.; Vainio, S.J.; Perreault, C. Wnt4 regulates thymic cellularity through the expansion of thymic epithelial cells and early thymic progenitors. Blood 2011, 118, 5163–5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Luo, H.; Li, H.; Zhao, Y. Thymic epithelial cell development and differentiation: Cellular and molecular regulation. Protein Cell 2013, 4, 342–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Fukai, I.; Masaoka, A.; Hashimoto, T.; Yamakawa, Y.; Mizuno, T.; Tanamura, O. Cytokeratins in normal thymus and thymic epithelial tumors. Cancer 1993, 71, 99–105. [Google Scholar] [CrossRef]
- Kuo, T. Cytokeratin profiles of the thymus and thymomas: Histogenetic correlations and proposal for a histological classification of thymomas. Histopathology 2000, 36, 403–414. [Google Scholar] [CrossRef]
- Rosai, J.; Sobin, L.H. Histological Typing of Tumours of the Thymus; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Ströbel, P.; Murumägi, A.; Klein, R.; Luster, M.; Lahti, M.; Krohn, K. Deficiency of the autoimmune regulator AIRE in thymomas is insufficient to elicit autoimmune polyendocrinopathy syndrome type 1 (APS-1). J. Pathol. 2007, 211, 563–571. [Google Scholar] [CrossRef]
- Marx, A.; Willcox, N.; Leite, M.I.; Chuang, W.Y.; Schalke, B.; Nix, W. Thymoma and paraneoplastic myasthenia gravis. Autoimmunity 2010, 43, 413–427. [Google Scholar] [CrossRef]
- Marx, A.; Pfister, F.; Schalke, B.; Saruhan-Direskeneli, G.; Melms, A.; Ströbel, P. The different roles of the thymus in the pathogenesis of the various myasthenia gravis subtypes. Autoimmun. Rev. 2013, 12, 875–884. [Google Scholar] [CrossRef]
- Melzer, N.; Ruck, T.; Fuhr, P.; Gold, R.; Hohlfeld, R.; Marx, A. Clinical features, pathogenesis, and treatment of myasthenia gravis: A supplement to the Guidelines of the German Neurological Society. J. Neurol. 2016, 263, 1473–1494. [Google Scholar] [CrossRef] [Green Version]
- Marx, A.; Ströbel, P.; Weis, C.A. The Pathology of the Thymus in Myasthenia Gravis. Mediastinum 2018, 2, 66. [Google Scholar] [CrossRef]
- Wolff, A.S.; Kärner, J.; Owe, J.F.; Oftedal, B.E.; Gilhus, N.E.; Erichsen, M.M. Clinical and serologic parallels to APS-I in patients with thymomas and autoantigen transcripts in their tumors. J. Immunol. 2014, 193, 3880–3890. [Google Scholar] [CrossRef] [Green Version]
- Marx, A.; Porubsky, S.; Belharazem, D.; Saruhan-Direskeneli, G.; Schalke, B.; Ströbel, P. Thymoma related myasthenia gravis in humans and potential animal models. Exp. Neurol. 2015, 270, 55–65. [Google Scholar] [CrossRef]
- Yamada, Y.; Tomaru, U.; Ishizu, A.; Kiuchi, T.; Kasahara, M.; Matsuno, Y. Expression of thymoproteasome subunit beta5t in type AB thymoma. J. Clin. Pathol. 2014, 67, 276–278. [Google Scholar] [CrossRef]
- Hayashi, A.; Fumon, T.; Miki, Y.; Sato, H.; Yoshino, T.; Takahashi, K. The evaluation of immunohistochemical markers and thymic cortical microenvironmental cells in distinguishing thymic carcinoma from type b3 thymoma or lung squamous cell carcinoma. J. Clin. Exp. Hematop. 2013, 53, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Brambilla, E.; Müller-Hermelink, H.K.; Harris, C.C. WHO Classification of Tumours of the Lung, Pleura, Thymus and Heart; Kleihues, P., Sobin, L.H., Eds.; IARC Press: Lyon, France, 2004. [Google Scholar]
- Yamada, Y.; Simon-Keller, K.; Belharazem-Vitacolonnna, D.; Bohnenberger, H.; Kriegsmann, M.; Kriegsmann, K. A Tuft Cell-Like Signature Is Highly Prevalent in Thymic Squamous Cell Carcinoma and Delineates New Molecular Subsets Among the Major Lung Cancer Histotypes. J. Thorac. Oncol. 2021, 16, 1003–1016. [Google Scholar] [CrossRef]
- Yamada, Y.; Sugimoto, A.; Hoki, M.; Yoshizawa, A.; Hamaji, M.; Date, H. POU2F3 beyond thymic carcinomas: Expression across the spectrum of thymomas hints to medullary differentiation in type A thymoma. Virchows Arch. 2022, 480, 843–851. [Google Scholar] [CrossRef]
- Marino, M.; Müller-Hermelink, H.K. Thymoma and thymic carcinoma. Relation of thymoma epithelial cells to the cortical and medullary differentiation of thymus. Virchows Arch. A Pathol. Anat. Histopathol. 1985, 407, 119–149. [Google Scholar] [CrossRef]
- Szadvari, I.; Krizanova, O.; Babula, P. Athymic nude mice as an experimental model for cancer treatment. Physiol. Res. 2016, 65 (Suppl. S4), S441–S453. [Google Scholar] [CrossRef]
- Boone, J.D.; Dobbin, Z.C.; Straughn, J.M., Jr.; Buchsbaum, D.J. Ovarian and cervical cancer patient derived xenografts: The past, present, and future. Gynecol. Oncol. 2015, 138, 486–491. [Google Scholar] [CrossRef]
- Conforti, F.; Zhang, X.; Rao, G.; De Pas, T.; Yonemori, Y.; Rodriguez, J.A. Therapeutic Effects of XPO1 Inhibition in Thymic Epithelial Tumors. Cancer Res. 2017, 77, 5614–5627. [Google Scholar] [CrossRef] [Green Version]
- Maury, J.M.; Merveilleux du Vignaux, C.; Drevet, G.; Zarza, V.; Chalabreysse, L.; Maisse, C. Activation of the mTOR/ Akt pathway in thymic epithelial cells derived from thymomas. PLoS ONE 2019, 14, e0197655. [Google Scholar] [CrossRef] [Green Version]
- Rajan, A.; Zhao, C. Deciphering the biology of thymic epithelial tumors. Mediastinum 2019, 3, 36. [Google Scholar] [CrossRef]
- Thomas, A.; Rajan, A.; Berman, A.; Tomita, Y.; Brzezniak, C.; Lee, M.J. Sunitinib in patients with chemotherapy-refractory thymoma and thymic carcinoma: An open-label phase 2 trial. Lancet Oncol. 2015, 16, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Zucali, P.A.; De Pas, T.M.; Palmieri, G.; Favaretto, A.; Chella, A.; Tiseo, M.; Caruso, M.; Simonelli, M.; Perrino, M.; De Vincenzo, F.; et al. Phase II study of everolimus in patients with thymoma and thymic carcinoma previously treated with cisplatin-based chemotherapy. J. Clin. Oncol. 2014, 36, 342–349. [Google Scholar] [CrossRef]
- Hellyer, J.A.; Ouseph, M.M.; Padda, S.K.; Wakelee, H.A. Everolimus in the treatment of metastatic thymic epithelial tumors. Lung Cancer 2020, 149, 97–102. [Google Scholar] [CrossRef]
- Lopez-Chavez, A.; Thomas, A.; Rajan, A.; Raffeld, M.; Morrow, B.; Kelly, R. Molecular profiling and targeted therapy for advanced thoracic malignancies: A biomarker-derived, multiarm, multihistology phase II basket trial. J. Clin. Oncol. 2015, 33, 1000–1007. [Google Scholar] [CrossRef]
- Wang, Y.; Thomas, A.; Lau, C.; Rajan, A.; Zhu, Y.; Killian, J.K. Mutations of epigenetic regulatory genes are common in thymic carcinomas. Sci. Rep. 2014, 4, 7336. [Google Scholar] [CrossRef] [Green Version]
- Steele, N.L.; Plumb, J.A.; Vidal, L.; Tjørnelund, J.; Knoblauch, P.; Rasmussen, A. A phase 1 pharmacokinetic and pharmacodynamic study of the histone deacetylase inhibitor belinostat in patients with advanced solid tumors. Clin. Cancer Res. 2008, 14, 804–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaccone, G.; Rajan, A.; Berman, A.; Kelly, R.J.; Szabo, E.; Lopez-Chavez, A. Phase II study of belinostat in patients with recurrent or refractory advanced thymic epithelial tumors. J. Clin. Oncol. 2011, 29, 2052–2059. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Rajan, A.; Szabo, E.; Tomita, Y.; Carter, C.A.; Scepura, B. A phase I/II trial of belinostat in combination with cisplatin, doxorubicin, and cyclophosphamide in thymic epithelial tumors: A clinical and translational study. Clin. Cancer Res. 2014, 20, 5392–5402. [Google Scholar] [CrossRef] [Green Version]
- Radovich, M.; Solzak, J.P.; Hancock, B.A.; Conces, M.L.; Atale, R.; Porter, R.F. A large microRNA cluster on chromosome 19 is a transcriptional hallmark of WHO type A and AB thymomas. Br. J. Cancer 2016, 114, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganci, F.; Vico, C.; Korita, E.; Sacconi, A.; Gallo, E.; Mori, F. MicroRNA expression profiling of thymic epithelial tumors. Lung Cancer 2014, 85, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Bellissimo, T.; Ganci, F.; Gallo, E.; Sacconi, A.; Tito, C.; De Angelis, L. Thymic Epithelial Tumors phenotype relies on miR-145-5p epigenetic regulation. Mol. Cancer 2017, 16, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tito, C.; Ganci, F.; Sacconi, A.; Masciarelli, S.; Fontemaggi, G.; Pulito, C. LINC00174 is a novel prognostic factor in thymic epithelial tumors involved in cell migration and lipid metabolism. Cell Death Dis. 2020, 11, 959. [Google Scholar] [CrossRef]
- Wang, J.; Huang, H.; Zhang, X.; Ma, H. LOXL1-AS1 promotes thymoma and thymic carcinoma progression by regulating miR-525-5p-HSPA9. Oncol. Rep. 2021, 45, 117. [Google Scholar] [CrossRef]
- Padda, S.K.; Gökmen-Polar, Y.; Hellyer, J.A.; Badve, S.S.; Singh, N.K.; Vasista, S.M. Genomic clustering analysis identifies molecular subtypes of thymic epithelial tumors independent of World Health Organization histologic type. Oncotarget 2021, 12, 1178–1186. [Google Scholar] [CrossRef]
- Psilopatis, I.; Pergaris, A.; Vrettou, K.; Theocharis, S.; Troungos, C. Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations. Int. J. Mol. Sci. 2022, 23, 4045. [Google Scholar] [CrossRef]
- Thomas, A.; Chen, Y.; Berman, A.; Schrump, D.S.; Giaccone, G.; Pastan, I. Expression of mesothelin in thymic carcinoma and its potential therapeutic significance. Lung Cancer 2016, 101, 104–110. [Google Scholar] [CrossRef]
- Hassan, R.; Thomas, A.; Alewine, C.; Le, D.T.; Jaffee, E.M.; Pastan, I. Mesothelin Immunotherapy for Cancer: Ready for Prime Time? J. Clin. Oncol. 2016, 34, 4171–4179. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Heong, V.; Peng Yong, W.; Soo, R.; Ean Chee, C.; Wong, A. A phase 1 study of the safety, pharmacokinetics and pharmacodynamics of escalating doses followed by dose expansion of the selective inhibitor of nuclear export (SINE) selinexor in Asian patients with advanced or metastatic malignancies. Ther. Adv. Med. Oncol. 2022, 14, 17588359221087555. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Klingbeil, O.; He, X.Y.; Wu, X.S.; Arun, G.; Lu, B. POU2F3 is a master regulator of a tuft cell-like variant of small cell lung cancer. Genes Dev. 2018, 32, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Hu, R.; Ding, Q.; Savage, T.K.; Huffman, K.E.; Williams, N. Subtype-specific secretomic characterization of pulmonary neuroendocrine tumor cells. Nat. Commun. 2019, 10, 3201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, K.D.; Ireland, A.S.; Oliver, T.G. Killing SCLC: Insights into how to target a shapeshifting tumor. Genes Dev. 2022, 36, 241–258. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main cellular compartments of the thymus. The figure shows the main cellular components of the thymus, epithelial and lymphoid, and their organotypic organization. Staining with CK19 outlines the thymic compartments, Cortex, Medulla and Perivascular spaces bordered by an epithelial cell lining; Hassall’s corpuscle can also be observed (A); in (B) Aire nuclear staining (white arrows) of cells around Hassall’s corpuscles (HCs) (black arrow) in the thymic Medulla. In (C) the overall thymic structure is shown in H&E: in the thymic Cortex—(C), epithelial cells (cTEC) constitute a network which is hardly seen at low magnification, where thymocytes are embedded; in the medulla (M) scattered mTEC establish a network where mature lymphocytes are embedded; HCs are also present, formed by concentric epithelial swirls (white arrows). A and B: courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany. HC: Hassall’s corpuscle; M: medulla; C: cortex; CMJ: corticomedullary junction; PVS: perivascular space.
Figure 1.
Main cellular compartments of the thymus. The figure shows the main cellular components of the thymus, epithelial and lymphoid, and their organotypic organization. Staining with CK19 outlines the thymic compartments, Cortex, Medulla and Perivascular spaces bordered by an epithelial cell lining; Hassall’s corpuscle can also be observed (A); in (B) Aire nuclear staining (white arrows) of cells around Hassall’s corpuscles (HCs) (black arrow) in the thymic Medulla. In (C) the overall thymic structure is shown in H&E: in the thymic Cortex—(C), epithelial cells (cTEC) constitute a network which is hardly seen at low magnification, where thymocytes are embedded; in the medulla (M) scattered mTEC establish a network where mature lymphocytes are embedded; HCs are also present, formed by concentric epithelial swirls (white arrows). A and B: courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany. HC: Hassall’s corpuscle; M: medulla; C: cortex; CMJ: corticomedullary junction; PVS: perivascular space.

Figure 2.
(A) shows a low magnification view of the normal thymus stained with H&E; a prominent cystic HC full of keratinic debris is clearly visible; CMJ is indicated at the interface C–M; (B) staining with CK AE1/AE3: the peripheral part of a Hassall Corpuscle (HC) is stained, as well as scattered EC in the cortex and medulla. PVS bordered by a TEC lining is shown; (C) CK10 staining of the peripheral part of HCs. C: courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany); (D) The cytokeratin antibody CK5/6 is considered a marker of a basal-layer type of CK, found in the early stages of epidermal keratinocyte differentiation. In addition to scattered TEC in the cortex and medulla, a prominent staining of subcapsular TEC bordering the cortical lobules is clearly visible. Modified from Ramieri M.T., and colleagues [26]. HC: Hassall’s corpuscle; M: Medulla; C: cortex; CMJ: corticomedullary junction; PVS: perivascular space; SubC TEC: subcapsular Thymic epithelial cells.
Figure 2.
(A) shows a low magnification view of the normal thymus stained with H&E; a prominent cystic HC full of keratinic debris is clearly visible; CMJ is indicated at the interface C–M; (B) staining with CK AE1/AE3: the peripheral part of a Hassall Corpuscle (HC) is stained, as well as scattered EC in the cortex and medulla. PVS bordered by a TEC lining is shown; (C) CK10 staining of the peripheral part of HCs. C: courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany); (D) The cytokeratin antibody CK5/6 is considered a marker of a basal-layer type of CK, found in the early stages of epidermal keratinocyte differentiation. In addition to scattered TEC in the cortex and medulla, a prominent staining of subcapsular TEC bordering the cortical lobules is clearly visible. Modified from Ramieri M.T., and colleagues [26]. HC: Hassall’s corpuscle; M: Medulla; C: cortex; CMJ: corticomedullary junction; PVS: perivascular space; SubC TEC: subcapsular Thymic epithelial cells.

Figure 3.
Corticomedullary markers in the thymus and in thymoma. (A,B) show thymoproteasome (β5t thymoproteasome, β5t) staining, respectively, in normal thymus by cTEC (a network of reactive TEC is clearly visible) (A) and by neoplastic TEC in a Type B2 thymoma (B). In (B), the dark arrow indicates neoplastic EC with strong β5t staining; (C) claudin 4-R56 stain of normal thymus: all reacting TEC are in the medulla; (D) only few EC of the type B2 thymoma are stained with claudin 4-R56, indicated by a dark arrow (courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany). C: cortex; M: medulla.
Figure 3.
Corticomedullary markers in the thymus and in thymoma. (A,B) show thymoproteasome (β5t thymoproteasome, β5t) staining, respectively, in normal thymus by cTEC (a network of reactive TEC is clearly visible) (A) and by neoplastic TEC in a Type B2 thymoma (B). In (B), the dark arrow indicates neoplastic EC with strong β5t staining; (C) claudin 4-R56 stain of normal thymus: all reacting TEC are in the medulla; (D) only few EC of the type B2 thymoma are stained with claudin 4-R56, indicated by a dark arrow (courtesy of Prof. Marx and of Prof. Ströbel, Mannheim/Göttingen, Germany). C: cortex; M: medulla.


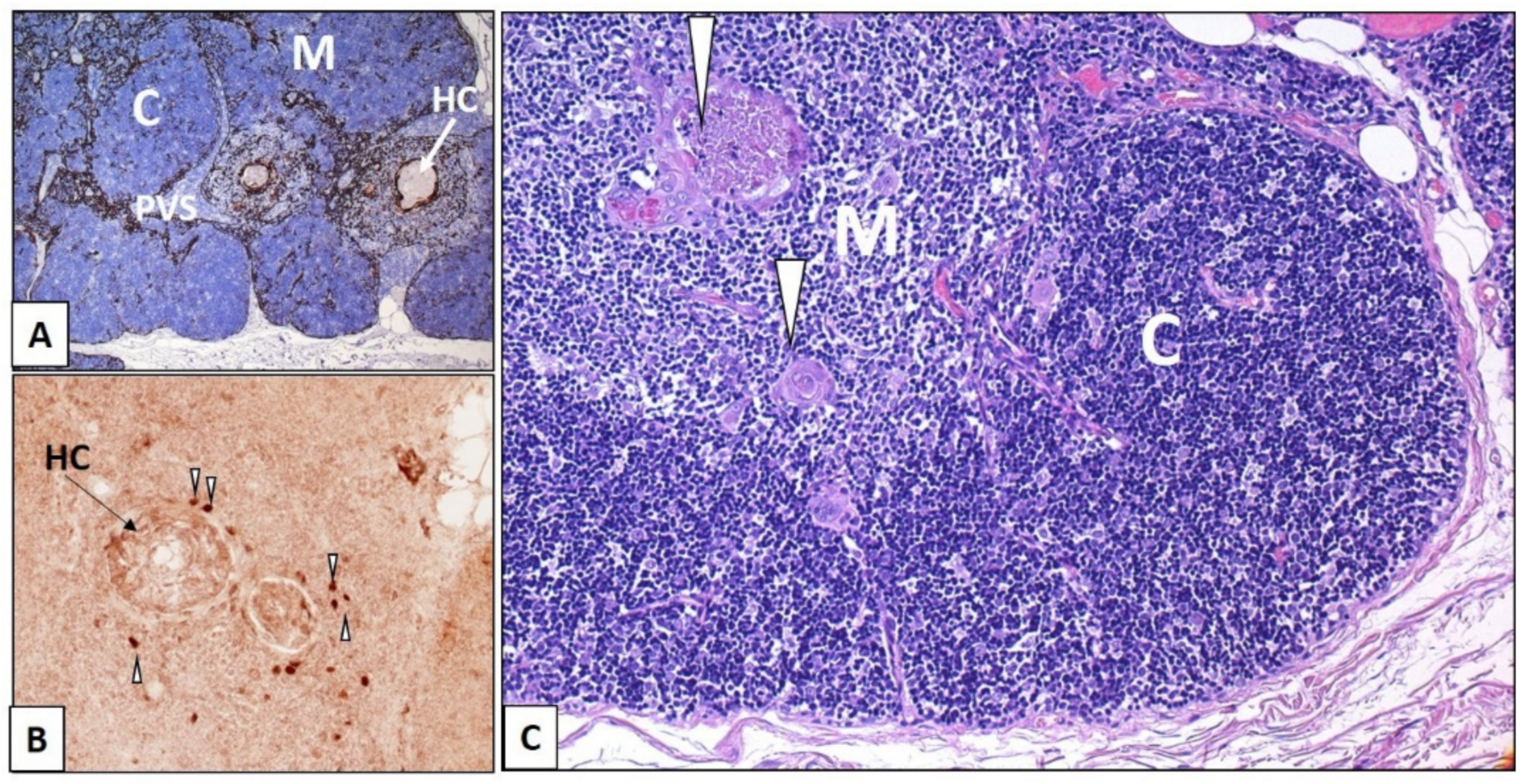{kind=link}
{kind=link}
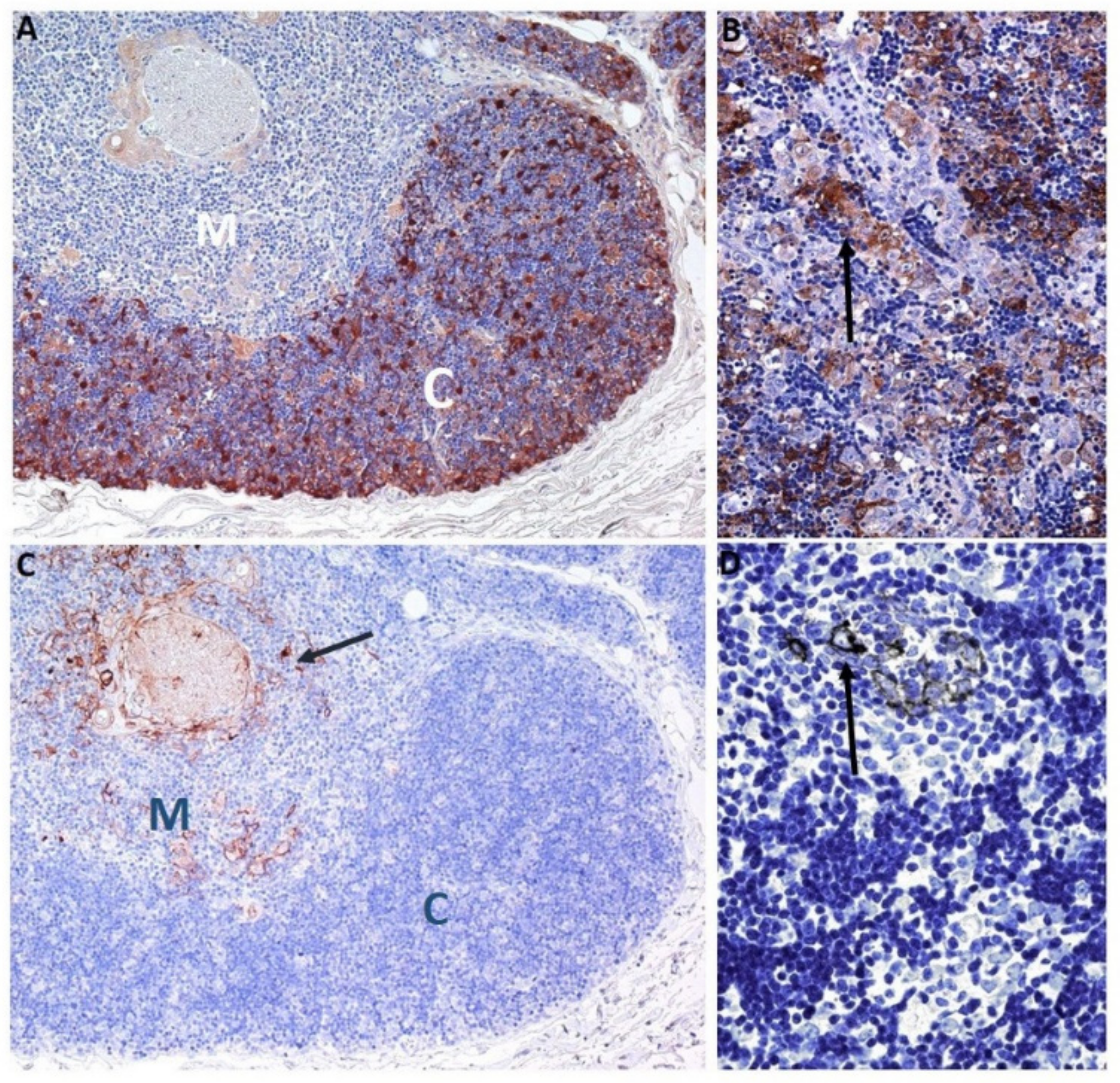{kind=link}
Table 1.
Main genetic, cellular and available clinical characteristics of selected genes involved in Thymic development, in the pathogenesis of autoimmune disease and in TET.
Table 1.
Main genetic, cellular and available clinical characteristics of selected genes involved in Thymic development, in the pathogenesis of autoimmune disease and in TET.
| Biomarker | Foxn1 Foxn1 (gene) | CD205 (Protein) Ly75 (Gene) | Notch1 | Aire Aire (Gene) | β5tProteasome Psmb11 (Gene) | Claudins (in mTEC Cld3/4) | Keratins |
|---|---|---|---|---|---|---|---|
| Chromosome involved/gene name | In humans: 17q11.2 | Humans: chromosome 2 | Humans: 9q34 | 21q22.3 | In vertebrates: Chrom. 1 and 11; in humans chrom 14 | Cld3/Cld4 on chrom 7q11; Cld5 on chrom22q11 | Different chrom.according to CK |
| Mol. biology | forkhead box gene family of TF | integral MB protein belonging to family of C-type multilecitin | TransMB receptor | TF | Multi-subunit protease complex | TransMB protein large family (more than 27 members) | CK5/14: primary CK of basal cells; CK8/18 primary CK of simple epithelium |
| Cellular localization by IHC/IF * | Nuclear expression (scattered EC) | Coarse granular cytoplasmic with membranous accentuation | Nuclear expression | Nuclear | cTEC (expression dependent on Foxn1) | TEC scattered in medulla Cld5 in endothelium | All TEC CK19+; CK14+ in subcapsular TEC, mTEC and HCs; CK8/18+ in mTEC and HCs; CK10+ only in HCs |
| Target cells/functions | Thymic cTEC and mTEC | Mature cTEC for positive selection of T lymphocytes; DC subpopulations | Regulator of TECP and mTEC fate during fetal thymus development; Notch1 signaling required for T-cell | mTEChigh; eTACs | It produces in cTEC unique Class I binding peptides supporting positive selection of CD8+ cells | Involved in Tight-Junctions(TJ); | CK8/18 and 19 stain cTEC CK5/14 stain mTEC; K5 and K8 stain TEC at the CMJ (By IF *) |
| Target genes/cells in thymus | CCL25, CXCL12, Dll4; Psmb11 encoding β5tProteasome | Involved in positive selection of thymocytes in the C; in generation of cTEC and AIRE+ mTEC | Commitment of thymus progenitors; it promotes establishment/expansion of the mTEPC pool | It acts in a multi-molecular complex | Mature cTEC | embryonic Cld3,4hi TEC maintain functional mTEC regeneration; assure lifelong central T cell tolerance | Different TEC subpop. Foxn1 always needed to maintain the 3D-thymic structure. |
| Human diseases/mice models | In AR Foxn1 deficit: Nude SCID phenotype. Several MDM with different Foxn1 alleles associated to different phenotypes. | CD205-deficient (Ly75−/−) and Ly75+/− MDM available; moreover isolated CD205 + CD40− TEC formed RTOC consisting of cortical and medullary thymic regions | loss-of-function and gain of function models (Notch loss of function results in mTEC hypoplasia, while its gain of function leads to TEPC maturation arrest) | Autoimm. polyendocrinopathy candidiasis ectodermal dysplasia (APECED; later APS1) | β5t-deficient mice displayed an altered TCR repertoire, and were functionally defective in IR; PSMB11 polymorphisms altered the CD8 T cell repertoire | Knockout models; “Cld-pathies” associated to some members of the family; some cancers | MDM: Examples: Rac1 deleted K14+ embryonic cells showed failure in thymic organogenesis; Rac1 deleted post-natal K14 + TEC showed thymic atrophy |
| Experimental and/or clinical findings | In Homoz. Mutations congenital universal alopecia, nail dystrophy; severe T-cell ID; life-threatening infections; thymic aplasia | CD205-targeted Ag administration resulted in successful vaccination against HIV gag-antigens and cancer antigens in MDM | Extrathymic disease: Multiple human disorders: developmental syndromes and adult onset diseases | Genetically engineered Aire deficient mice show severe autoimm disease manifestations | PSMB11 polymorphisms associated with a higher risk of an AID in humans (Sjögren’s disease); SNV in human β5t gene caused no severe health problems in homozygous individuals | Cld3/4 were found highly upregulated in multiple cancers and possible target of therapies; Cld3/4 are surface receptors for Clostridium perfringens enterotoxin in gut | In the K5CrexRac1flox/flox transgenic mouse, most mice showed athymia/few other showed remnants of M-C architecture |
| References | [20,27,28,29] | [30,31,32,33,34] | [35,36,37,38,39] | [40,41,42,43,44] | [45,46,47,48,49,50] | [51,52,53,54,55,56] | [41,57,58,59,60] |
Abbreviations: IHC = immunohistochemistry; IF = Immunofluorescence; MB = membrane; TransMB = transmembrane; TF = transcription factor; Autoimm. = autoimmune; ID = immunodeficiency; AID = Autoimmune Disease; IR = Immune response; AR = autosomic recessive;eTACs = Extathymic AIRE-expressing cells; TECP = Thymic epithelial progenitor cells; “Cld-pathies” = claudinopathies; subpop = subpopulations; mTEChigh = highly expressed in mTEC; RTOC = re-aggregated thymic organ cultures; MDM = murine disease models; HIV = Human Immunodeficiency Virus; Ag = Antigen; SNV: single nucleotide variants; * The immunohistochemical characteristics reported refer to the limited data available for human specimens (pediatric or hyperplastic thymuses); most experimental papers are immunofluorescence-based on embryonic/fetal or adult thymuses from mice or humans.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gallo, E.; Ramieri, M.T.; Marino, M. Structural and Functional Thymic Biomarkers Are Involved in the Pathogenesis of Thymic Epithelial Tumors: An Overview. Immuno 2022, 2, 408-429. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno2020025
AMA Style
Gallo E, Ramieri MT, Marino M. Structural and Functional Thymic Biomarkers Are Involved in the Pathogenesis of Thymic Epithelial Tumors: An Overview. Immuno. 2022; 2(2):408-429. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno2020025
Chicago/Turabian StyleGallo, Enzo, Maria Teresa Ramieri, and Mirella Marino. 2022. "Structural and Functional Thymic Biomarkers Are Involved in the Pathogenesis of Thymic Epithelial Tumors: An Overview" Immuno 2, no. 2: 408-429. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno2020025