The Effects of Live Feeding on Swimming Activity and Exhibit Use in Zoo Humboldt Penguins (Spheniscus humboldti)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
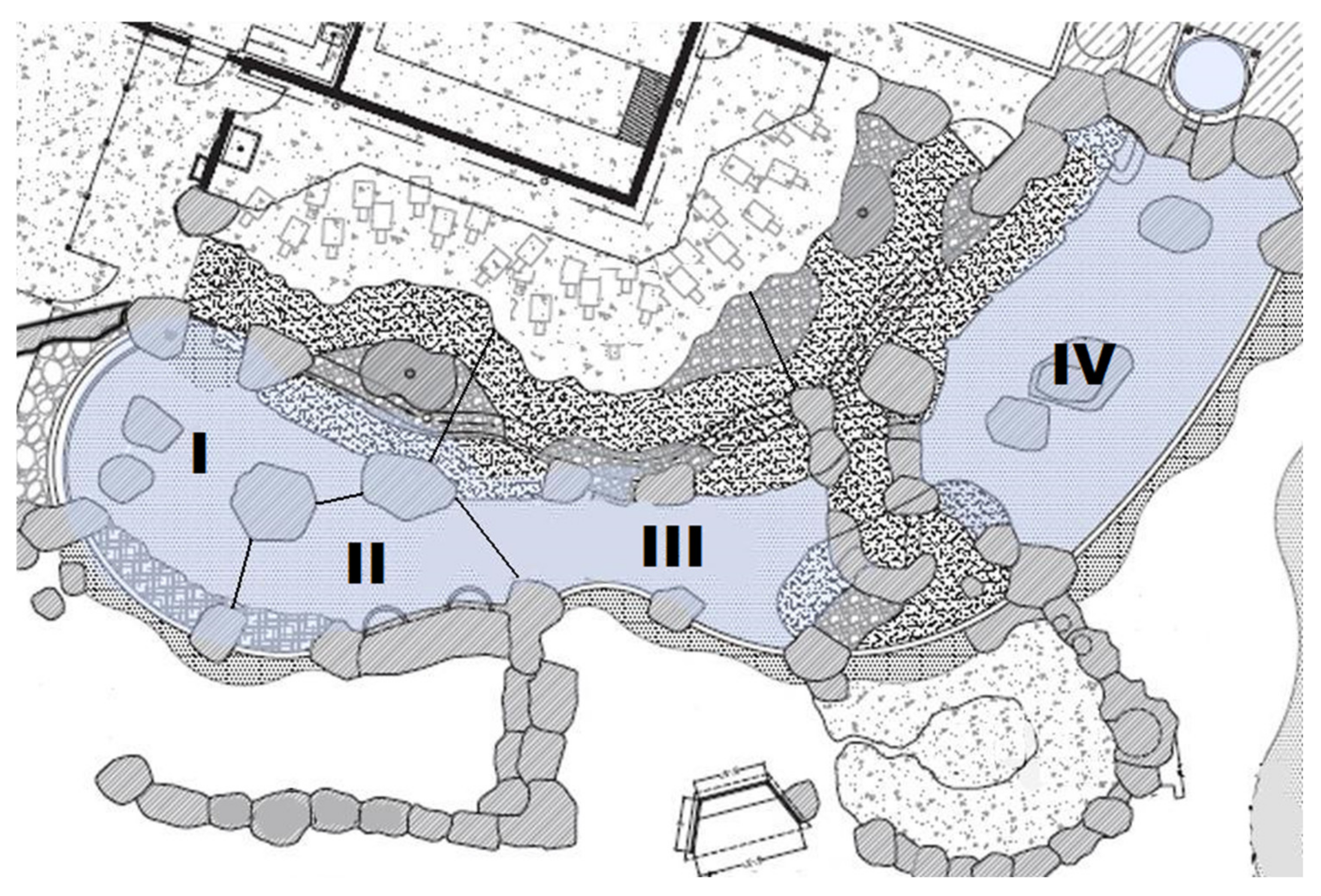
2.1. Subjects and Enclosures
2.2. Materials
2.3. Design and Procedure
2.4. Statistical Analyses
3. Results
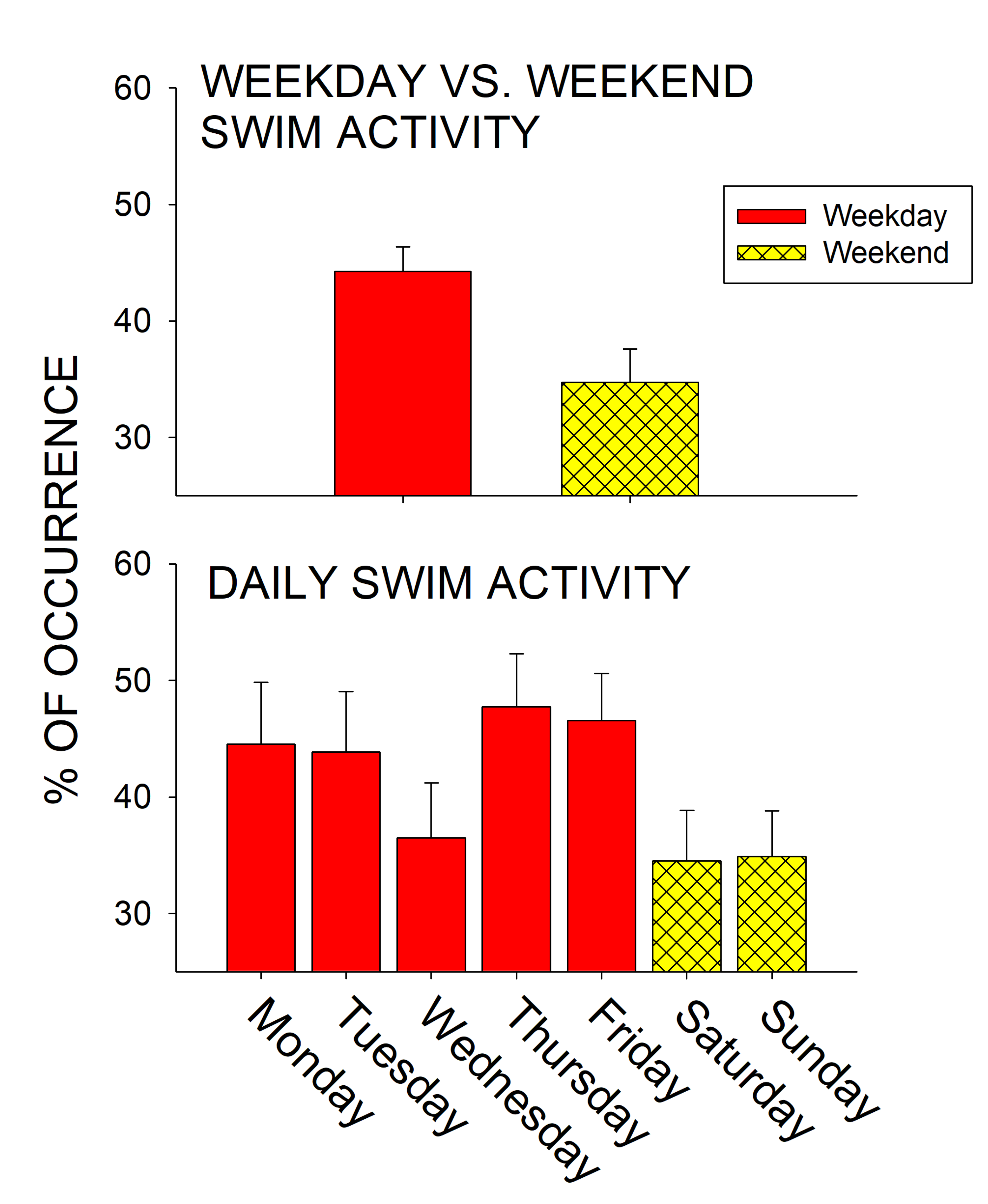
3.1. Pre-Trout Daily and Weekday Versus Weekend Swimming Activity
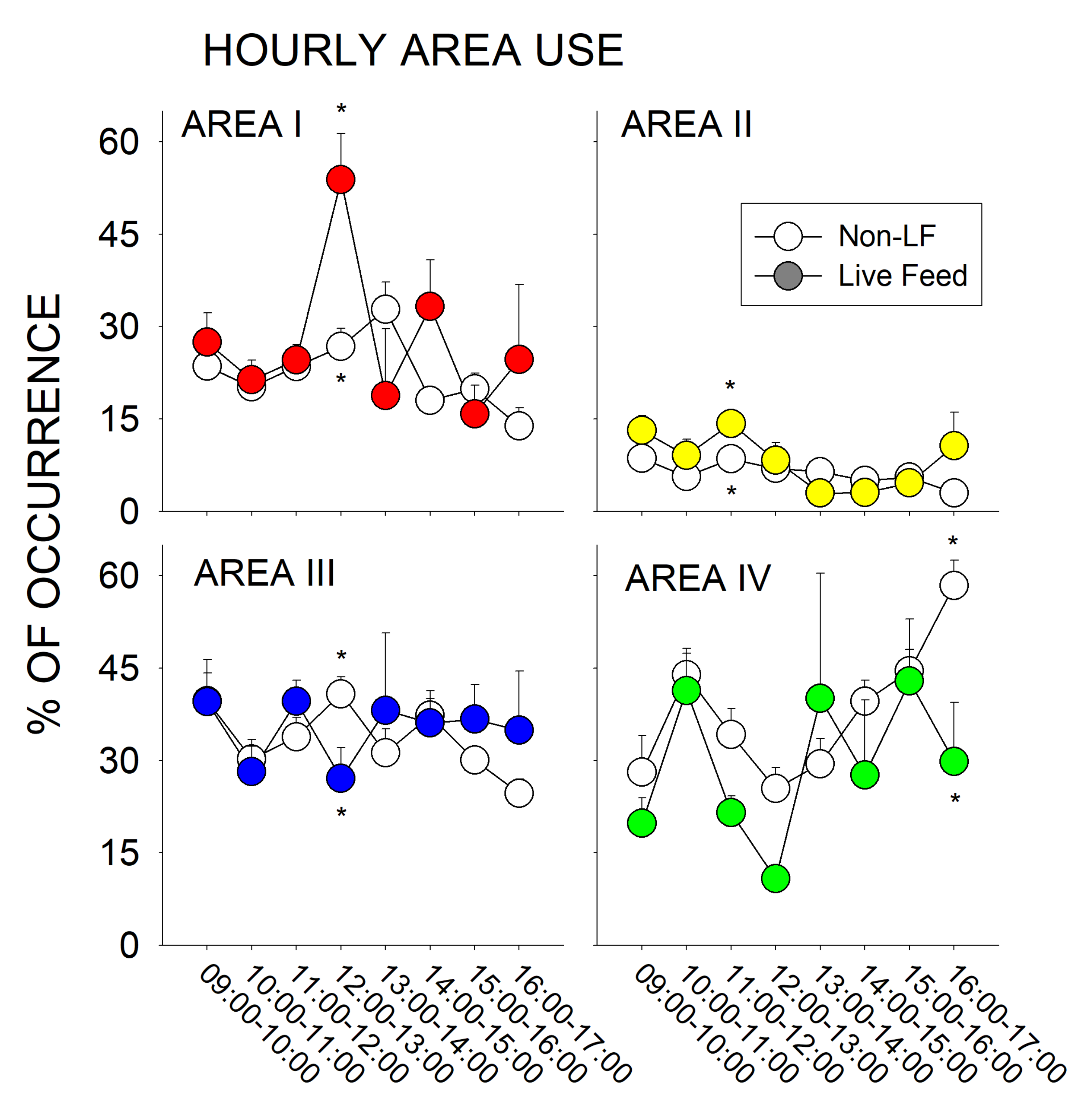
3.2. Live Feeding vs. Non-Live Feeding Days
4. Discussion
4.1. Pre-Trout Swimming Activity
4.2. Live Feeding vs. Non-Live Feeding Days
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borboroglu, P.G.; Boersma, P.D. Penguins: Natural History and Conservation; University of Washington Press: Seattle, WA, USA, 2013. [Google Scholar]
- Brown, C. Traveling speed and foraging range of macaroni and rockhopper penguins at Marion Island. J. Field Ornith. 1987, 58, 118–125. [Google Scholar]
- Lynch, W. Penguins of the World; Firefly Books Ltd.: Buffalo, NY, USA, 2007. [Google Scholar]
- Kinley, R. Enrichment training for penguins. Am. Anim. Trainer 2000, 1, 10–13. [Google Scholar]
- Fowler, G.S.; Fowler, M.E. Order Spheniciformes (penguins). In Biology, Medicine, and Surgery of South American Wild Animals; Fowler, M.E., Cubas, Z.S., Eds.; Iowa State University Press: Ames, IA, USA, 2001; Volume 547. [Google Scholar]
- Reidarson, T.; McBain, J.; Burch, L. A Novel Approach to the Treatment of Bumblefoot in Penguins. J. Avian Med. Surg. 1999, 13, 124–127. [Google Scholar]
- Maple, T.L.; Perdue, B.M. Zoo Animal Welfare; Springer: Berlin, Germany, 2013. [Google Scholar]
- Mellen, J.; Sevenich MacPhee, M. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Young, R.J. Environmental Enrichment for Captive Animals; Blackwell Publishing: Oxford, UK, 2003. [Google Scholar]
- Fernandez, E.J.; Timberlake, W. Mutual benefits of research collaborations between zoos and academic institutions. Zoo Biol. 2008, 27, 470–487. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef]
- Markowitz, H. Behavioral Enrichment in the Zoo; Van Nostrand Reinhold: New York, NY, USA, 1982. [Google Scholar]
- Shepherdson, D.J. Tracing the path of environmental enrichment in zoos. In Second Nature: Environmental Enrichment for Captive Animals; Sherpherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institutional Press: Washington, DC, USA, 1998; pp. 1–12. [Google Scholar]
- Andrews, N.L.; Ha, J.C. The effects of automated scatter feeders on captive grizzly bear activity budgets. J. Appl. Anim. Welf. Sci. 2014, 17, 148–156. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does enrichment improve reptile welfare? Leopard geckos (Eublepharis macularius) respond to five types of environmental enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar] [CrossRef]
- Carlstead, K.; Seidensticker, J.; Baldwin, R. Environmental enrichment for zoo bears. Zoo Biol. 1991, 10, 3–16. [Google Scholar] [CrossRef]
- Fernandez, E.J. Stereotypies and Foraging: Appetitive Search Behaviors and Stereotypies in Captive Animals; VDM Publishing: Saarbrucken, Germany, 2010. [Google Scholar]
- Markowitz Engineering environments for behavioral opportunities in the zoo. Behav Anal. 1978, 1, 34. [CrossRef]
- Markowitz, H.; Aday, C.; Gavazzi, A. Effectiveness of acoustic “prey”: Environmental enrichment for a captive African leopard (Panthera pardus). Zoo Biol. 1995, 14, 371–379. [Google Scholar] [CrossRef]
- Markowitz, H.; LaForse, S. Artificial prey as behavioral enrichment devices for felines. Appl. Anim. Behav. Sci. 1987, 18, 31–43. [Google Scholar] [CrossRef]
- Carlstead, K.; Brown, J.L.; Seidensticker, J. Behavioral and adrenocortical responses to environmental changes in leopard cats (Felis bengalensis). Zoo Biol. 1993, 12, 321–331. [Google Scholar] [CrossRef]
- Sherwin, C.M.; Lewis, P.D.; Perry, G.C. The effects of environmental enrichment and intermittent lighting on the behaviour and welfare of male domestic turkeys. Appl. Anim. Behav. Sci. 1999, 62, 319–333. [Google Scholar] [CrossRef]
- Carlstead, K.; Seidensticker, J. Seasonal variation in stereotypic pacing in an American black bear (Ursus americanus). Behav. Processes 1991, 25, 155–161. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Foraging devices as enrichment in captive walruses (Odobenus rosmarus). Behav. Processes 2019, 168, 103943. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.; Wells, D.L.; Hepper, P.G. The influence of olfactory stimulation on the behaviour of dogs housed in a rescue shelter. Appl. Anim. Behav. Sci. 2005, 91, 143–153. [Google Scholar] [CrossRef]
- Platt, D.M.; Novak, M.A. Videostimulation as enrichment for captive rhesus monkeys (Macaca mulatta). Appl. Anim. Behav. Sci. 1997, 52, 139–155. [Google Scholar] [CrossRef]
- Wells, D.L.; Irwin, R.M. Auditory stimulation as enrichment for zoo-housed Asian elephants (Elephas maximus). Anim. Welf. 2008, 17, 335–340. [Google Scholar]
- Fernandez, E.J.; Kinley, R.C.; Timberlake, W. Training penguins to interact with enrichment devices for lasting effects. Zoo Biol. 2019, 38, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Laule, G.; Desmond, T. Positive reinforcement training as an enrichment strategy. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 302–313. [Google Scholar]
- Altman, J.D. Animal activity and visitor learning at the zoo. Anthrozoos 1998, 11, 12–21. [Google Scholar] [CrossRef]
- Davey, G.; Henzi, P.; Higgins, L. The influence of environmental enrichment on Chinese visitor behavior. J. Appl. Anim. Welf. Sci. 2005, 8, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Godinez, A.M.; Fernandez, E.J.; Morrissey, K. Visitor behaviors and perceptions of jaguar activities. Anthrozoos 2013, 26, 613–619. [Google Scholar] [CrossRef]
- Kutska, D. Variation in visitor perceptions of a polar bear enclosure based on the presence of natural vs. un-natural enrichment items. Zoo Biol. 2009, 28, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Margulis, S.W.; Hoyos, C.; Anderson, M. Effect of felid activity on zoo visitor interest. Zoo Biol. 2003, 22, 587–599. [Google Scholar] [CrossRef]
- Stoinski, T.S.; Daniel, E.; Maple, T.L. A preliminary study of the behavioral effects of feeding enrichment on African elephants. Zoo Biol. 2000, 19, 485–493. [Google Scholar] [CrossRef]
- Law, G.; Macdonald, A.; Reid, A. Dispelling some common misconceptions about the keeping of felids in captivity. Int. Zoo Yearb. 1997, 35, 197–207. [Google Scholar] [CrossRef]
- McPhee, M.E.; Carlstead, K. The importance of maintaining natural behaviors in captive mammals. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management; Kleiman, D.G., Thompson, K.V., Kirk Baer, C., Eds.; The University of Chicago Press: Chicago, IL, USA, 2010; pp. 303–313. [Google Scholar]
- McPhee, M.E. Intact carcasses as enrichment for large felids: Effects on on-and off-Exhibit behaviors. Zoo Biol. 2002, 21, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Stark, B. The use of carcass feeding to enhance animal welfare. In Proceedings of the Seventh International Conference on Environmental Enrichment, New York, NY, USA, 31 July–5 August 2005; pp. 198–204. [Google Scholar]
- Forthman-Quick, D.L. An integrative approach to environmental engineering in zoos. Zoo Biol. 1984, 3, 65–77. [Google Scholar] [CrossRef]
- Markowitz, H.; Stevens, V.J. Behavior of Captive Wild Animals; Nelson-Hall: Chicago, IL, USA, 1978. [Google Scholar]
- Bashaw, M.J.; Bloomsmith, M.A.; Marr, M.J.; Maple, T.L. To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol. 2003, 22, 189–198. [Google Scholar] [CrossRef]
- Foster-Turley, P.; Markowitz, H. A captive behavioral enrichment study with Asian small-clawed river otters Aonyx cinerea. Zoo Biol. 1982, 1, 29–43. [Google Scholar] [CrossRef]
- Shepherdson, D.J.; Carlstead, K.; Mellen, J.D.; Seidensticker, J. The influence of food presentation on the behavior of small cats in confined environments. Zoo Biol. 1993, 12, 203–216. [Google Scholar] [CrossRef]
- Cooper, J.E.; Williams, D.L. The feeding of live food to exotic pets: Issues of welfare and ethics. J. Exot. Pet. Med. 2014, 23, 244–249. [Google Scholar] [CrossRef]
- Keller, M. Feeding live invertebrate prey in zoos and aquaria: Are there welfare concerns? Zoo Biol. 2017, 36, 316–322. [Google Scholar] [CrossRef]
- Wickins-Dražilová, D. Zoo animal welfare. J. Agric. Environ. Ethics 2006, 19, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Hosey, G.; Melfi, V.; Pankhurst, S. Feeding and nutrition. In Zoo Animals: Behaviour, Management, and Welfare; Hosey, G., Melfi, V., Pankhurst, S., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 418–460. [Google Scholar]
- Marshall, L.; McCormick, W.D.; Cooke, G.M. Perception of the ethical acceptability of live prey feeding to aquatic species kept in captivity. PLoS ONE 2019, 14, e0216777. [Google Scholar] [CrossRef] [Green Version]
- Cottle, L.; Tamir, D.; Hyseni, M.; Bühler, D.; Lindemann-Matthies, P. Feeding live prey to zoo animals: Response of zoo visitors in Switzerland. Zoo Biol. 2010, 29, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Ings, R.; Waran, N.K.; Young, R.J. Attitude of zoo visitors to the idea of feeding live prey to zoo animals. Zoo Biol. 1997, 16, 343–347. [Google Scholar] [CrossRef]
- McDole, E. Fish introduction to jaguars (Panthera onca): Response of zoo visitors and jaguars. Ph.D. Thesis, Georgia Institute of Technology, Atlanta, GA, USA, 2007. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Dickens, M. A statistical formula to quantify the “spread-of-participation” in group discussion. Speech Monogr. 1955, 22, 28–30. [Google Scholar] [CrossRef]
- Hedeen, S.E. Utilization of space by captive groups of lowland gorillas (Gorilla g. gorilla). Ohio J. Sci. 1982, 82, 27–30. [Google Scholar]
- Plowman, A.B. A note on a modification of the spread of participation index allowing for unequal zones. Appl. Anim. Behav. Sci. 2003, 83, 331–336. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Selecting and testing environmental enrichment in lemurs. Front. Psychol. 2019, 10, 2119. [Google Scholar] [CrossRef]
- Chiew, S.J.; Butler, K.L.; Sherwen, S.L.; Coleman, G.J.; Fanson, K.V.; Hemsworth, P.H. Effects of regulating visitor viewing proximity and the intensity of visitor behaviour on little penguin (Eudyptula minor) behaviour and welfare. Animals 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Chiew, S.J.; Butler, K.L.; Sherwen, S.L.; Coleman, G.J.; Melfi, V.; Burns, A.; Hemsworth, P.H. Effect of Covering a Visitor Viewing Area Window on the Behaviour of Zoo-Housed Little Penguins (Eudyptula minor). Animals 2020, 10, 1224. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.; Butler, K.L.; Hemsworth, P.H. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Davey, G. Visitors’ effects on the welfare of animals in the zoo: A review. J. Appl. Anim. Welf. Sci. 2007, 10, 169–183. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Tamborski, M.A.; Pickens, S.R.; Timberlake, W. Animal–visitor interactions in the modern zoo: Conflicts and interventions. Appl. Anim. Behav. Sci. 2009, 120, 1–8. [Google Scholar] [CrossRef]
- Hosey, G.R. Zoo animals and their human audiences: What is the visitor effect? Anim. Welf. 2000, 9, 343–357. [Google Scholar]
- Sherwen, S.L.; Hemsworth, P.H. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiew, S.J.; Hemsworth, P.H.; Melfi, V.; Sherwen, S.L.; Burns, A.; Coleman, G.J. Visitor attitudes towards little penguins (Eudyptula minor) at two Australian zoos. Front. Psychol. 2021, 12, 626185. [Google Scholar] [CrossRef] [PubMed]
- Chiew, S.J.; Hemsworth, P.H.; Sherwen, S.L.; Melfi, V.; Coleman, G.J. The effect of regulating zoo visitor-penguin interactions on zoo visitor attitudes. Front. Psychol. 2019, 10, 2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinez, A.M.; Fernandez, E.J. What is the zoo experience? How zoos impact a visitor’s behaviors, perceptions, and conservation efforts. Front. Psychol. 2019, 10, 1746. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.T.S.; Araujo, J.F. Food entrainment: Major and recent findings. Front. Behav. Neurosci. 2012, 6, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, E.J.; Timberlake, W. Superstition revisited: Sex, species, and adventitious reinforcement. Behav. Processes 2020, 170, 103979. [Google Scholar] [CrossRef]
- Gillman, A.G.; Rebec, G.V.; Pecoraro, N.C.; Kosobud, A.E. Circadian entrainment by food and drugs of abuse. Behav. Processes 2019, 165, 23–28. [Google Scholar] [CrossRef]
- Yoda, K.; Sato, K.; Niizuma, Y.; Kurita, M.; Bost, C.; Le Maho, Y.; Naito, Y. Precise monitoring of porpoising behaviour of Adélie penguins determined using acceleration data loggers. J. Exp. Biol. 1999, 202, 3121–3126. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, E.J.; Myers, M.; Hawkes, N.C. The Effects of Live Feeding on Swimming Activity and Exhibit Use in Zoo Humboldt Penguins (Spheniscus humboldti). J. Zool. Bot. Gard. 2021, 2, 88-100. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2010007
Fernandez EJ, Myers M, Hawkes NC. The Effects of Live Feeding on Swimming Activity and Exhibit Use in Zoo Humboldt Penguins (Spheniscus humboldti). Journal of Zoological and Botanical Gardens. 2021; 2(1):88-100. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2010007
Chicago/Turabian StyleFernandez, Eduardo J., Mark Myers, and Nancy C. Hawkes. 2021. "The Effects of Live Feeding on Swimming Activity and Exhibit Use in Zoo Humboldt Penguins (Spheniscus humboldti)" Journal of Zoological and Botanical Gardens 2, no. 1: 88-100. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2010007