
DNA Damage as a Potential Non-Invasive Indicator of Welfare: A Preliminary Study in Zoo-Housed Grizzly Bears (Ursus arctos horribilis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Subjects and Housing
2.3. Behavioral Observations
2.4. Sample Collection and Analysis
2.5. Additional Assay Validation Tests
2.6. Statistical Analyses
3. Results
3.1. Behavioral Comparisons among the Three Habitats
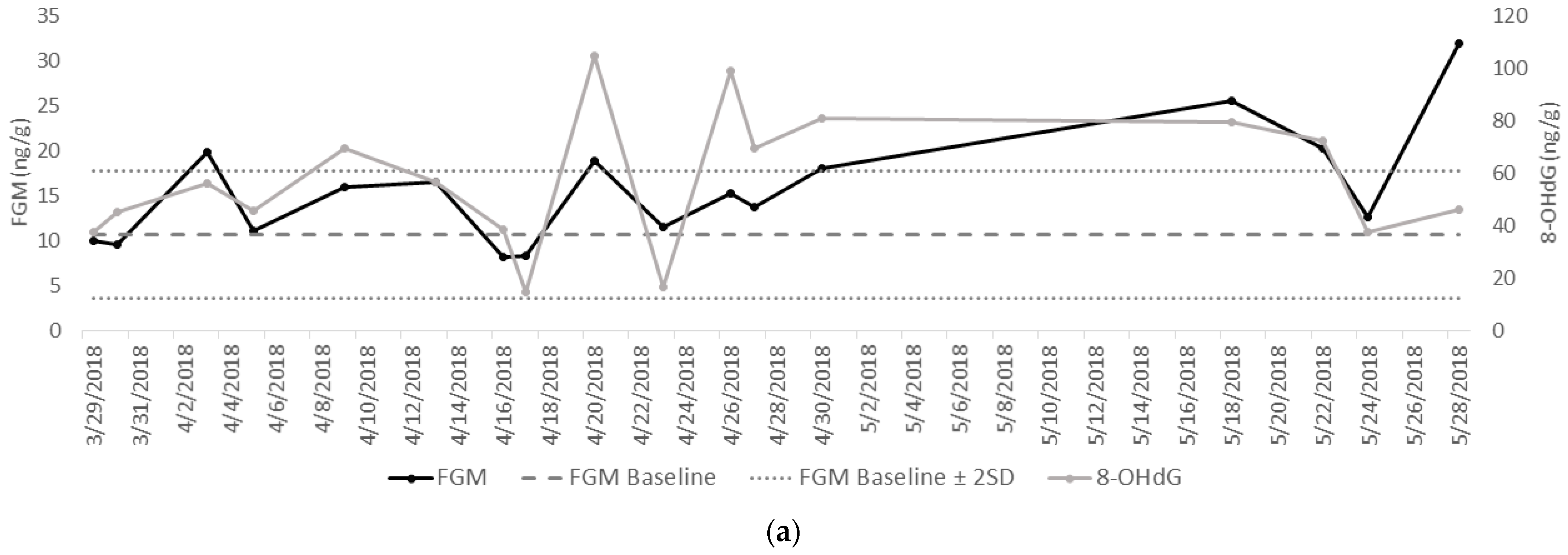
3.2. Patterns of FGMs and 8-OHdG in the Temporary, Smaller Habitat
3.3. Behavioral Comparisons with Physiological Measures
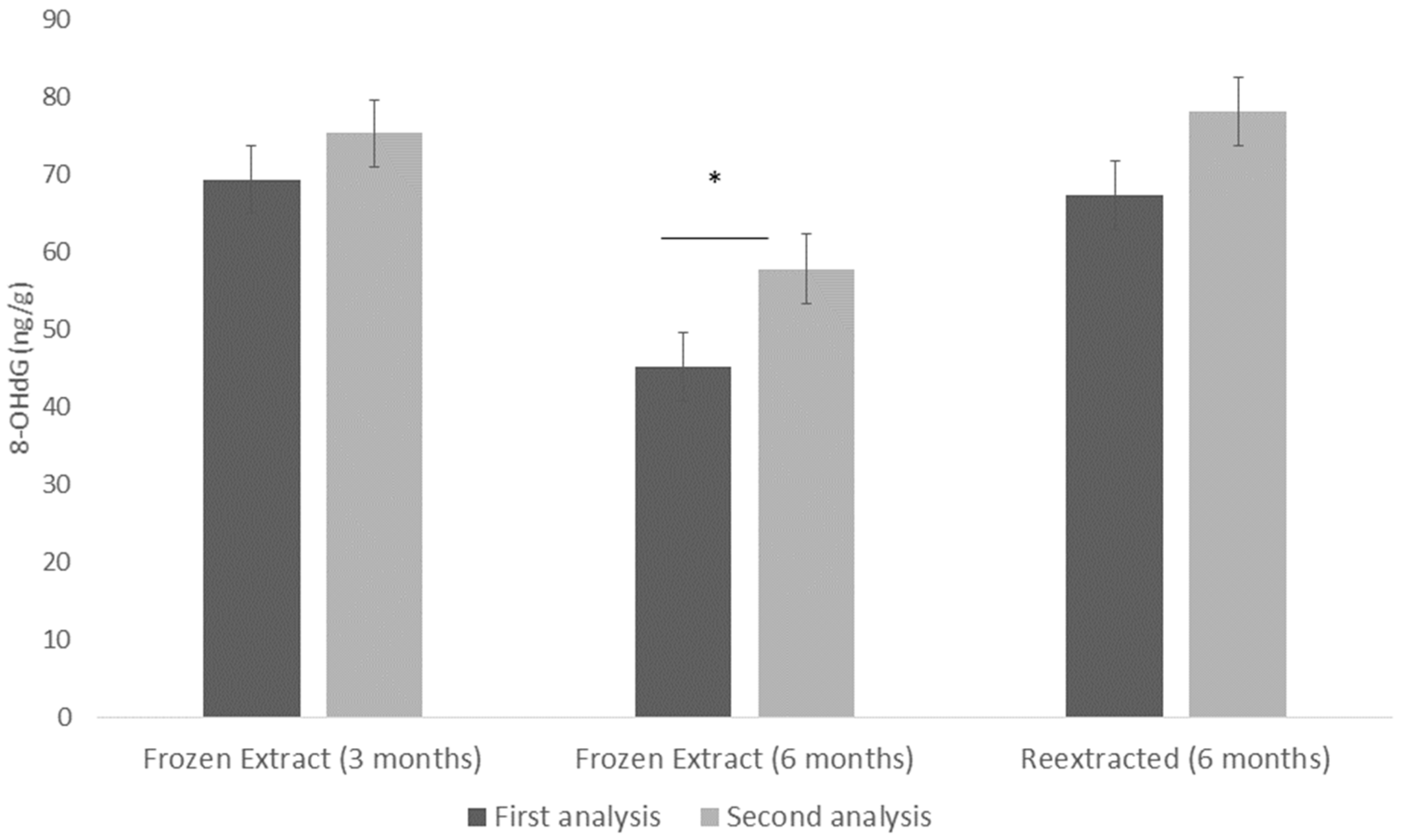
3.4. Assay Validation Tests
4. Discussion
4.1. Summary
4.2. Behavioral Patterns in the Three Habitats
4.3. Relationships between Physiological and Behavioral Indicators of Welfare
4.4. Origins and Measurement of Fecal 8-OHdG
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AZA. Association of Zoos and Aquariums, Animal Welfare Committee’s Definition of Animal Welfare. Available online: https://www.aza.org/animal_welfare_committee?locale=en (accessed on 1 February 2021).
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of Animal Welfare Research in Zoos and Aquariums: Where are we, Where to next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar]
- Mason, G.; Mendl, M. Why Is There No Simple Way of Measuring Animal Welfare? Anim. Welf. 1993, 2, 301–319. [Google Scholar]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “Stress” Are Not Synonymous. Integr. Org. Biol. 2019, 1, obz017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielebnowski, N. Stress and Distress: Evaluating Their Impact for the Well-being of Zoo Animals. J. Am. Vet. Med Assoc. 2003, 223, 973–977. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Ralph, C.; Tilbrook, A.J. Invited Review: The Usefulness of Measuring Glucocorticoids for Assessing Animal Welfare. J. Anim. Sci. 2016, 94. [Google Scholar] [CrossRef] [Green Version]
- Mormede, P.; Andanson, S.; Auperin, B.; Beerda, B.; Guemene, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the Hypothalamic-Pituitary-Adrenal Function as a Tool to Evaluate Animal Welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Bateson, M.; Poirier, C. Can biomarkers of biological age be used to assess cumulative lifetime experience? Anim. Welf. 2019, 28, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D. Oxidative Stress in Ecology and Evolution: Lessons from Avian Studies. Ecol. Lett. 2008, 11, 1238–1251. [Google Scholar] [CrossRef]
- Costantini, D.; Marasco, V.; Moller, A.P. A Meta-Analysis of Glucocorticoids as Modulators of Oxidative Stress in Vertebrates. J. Comp. Physiol. B 2011, 181, 447–456. [Google Scholar] [CrossRef]
- Olinski, R.; Gackowski, D.; Rozalski, R.; Foksinski, M.; Bialkowski, K. Oxidative DNA Damage in Cancer Patients: A Cause or a Consequence of the Disease Development? Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2003, 531, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The Interplay between Inflammation, Oxidative Stress, DNA Damage, DNA Repair and Mitochondrial Dysfunction in Depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, A.; Broedbaek, K.; Fink-Jensen, A.; Knorr, U.; Greisen Soendergaard, M.; Henriksen, T.; Weimann, A.; Jepsen, P.; Lykkesfeldt, J.; Poulsen, H.E.; et al. Increased Systemic Oxidatively Generated DNA and RNA Damage in Schizophrenia. Psychiatry Res. 2013, 209, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanolin, M.E.; Girardi, P.; Degan, P.; Rava, M.; Olivieri, M.; Di Gennaro, G.; Nicolis, M.; De Marco, R. Measurement of a Urinary Marker (8-hydroxydeoxyGuanosine, 8-OHdG) of DNA Oxidative Stress in Epidemiological Surveys: A Pilot Study. Int. J. Biol. Markers 2015, 30, 341–345. [Google Scholar] [CrossRef]
- Irie, M.; Tamae, K.; Iwamoto-Tanaka, N.; Kasai, H. Occupational and Lifestyle Factors and Urinary 8-hydroxydeoxyguanosine. Cancer Sci. 2005, 96, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, M.; Thierry, A.-M.; González-Acuña, D.; Polito, M.J. Integrating Oxidative Ecology into Conservation Physiology. Conserv. Physiol. 2013, 1, cot004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larcombe, S.D.; Tregaskes, C.A.; Coffey, J.; Stevenson, A.E.; Alexander, L.G.; Arnold, K.E. Oxidative Stress, Activity Behaviour and Body Mass in Captive Parrots. Conserv. Physiol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Gormally, B.M.G.; Estrada, R.; McVey, M.; Romero, L.M. Beyond Corticosterone: The Acute Stress Response Increases DNA Damage in House Sparrows. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2020, 1–12. [Google Scholar] [CrossRef]
- Stier, A.; Schull, Q.; Bize, P.; Lefol, E.; Haussmann, M.; Roussel, D.; Robin, J.-P.; Viblanc, V.A. Oxidative Stress and Mitochondrial Responses to Stress Exposure Suggest that King Penguins are Naturally Equipped to Resist Stress. Sci. Rep. 2019, 9, 8545. [Google Scholar] [CrossRef] [Green Version]
- Jakob-Hoff, R.; Kingan, M.; Fenemore, C.; Schmid, G.; Cockrem, J.F.; Crackle, A.; Van Bemmel, E.; Connor, R.; Descovich, K. Potential Impact of Construction Noise on Selected Zoo Animals. Animals 2019, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Waal, F.; Aureli, F.; Judge, P. Coping with Crowding. Sci. Am. 2000, 282, 76–81. [Google Scholar] [CrossRef]
- Cordoni, G.; Palagi, E. Response of Captive Lowland Gorillas (Gorilla gorilla gorilla) to Different Housing Conditions: Testing the Aggression-Density and Coping Models. J. Comp. Psychol. 2007, 121, 171–180. [Google Scholar] [CrossRef]
- Taylor, S.E. Tend and Befriend: Biobehavioral Bases of Affiliation under Stress. Curr. Dir. Psychol. Sci. 2006, 15, 273–277. [Google Scholar] [CrossRef]
- Stanic, D.; Plecas-Solarovic, B.; Petrovic, J.; Bogavac-Stanojevic, N.; Sopic, M.; Kotur-Stevuljevic, J.; Ignjatovic, S.; Pesic, V. Hydrogen Peroxide-Induced Oxidative Damage in Peripheral Blood Lymphocytes from Rats Chronically Treated with Corticosterone: The Protective Effect of Oxytocin Treatment. Chem. Biol. Interact. 2016, 256, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, R.A.; Wilson, D.E. Handbook of the Mammals of the World: Carnivores; Lynx Ediciones: Barcelona, Spain, 2009. [Google Scholar]
- Smith, T.S.; Herrero, S.; DeBruyn, T.D. Alaskan Brown Bears, Humans, and Habituation. Ursus 2005, 16, 1–10. [Google Scholar] [CrossRef]
- Gende, S.M.; Quinn, T.P. The Relative Importance of Prey Density and Social Dominance in Determining Energy Intake by Bears Feeding on Pacific Salmon. Can. J. Zool. 2004, 82, 75–85. [Google Scholar] [CrossRef]
- BearTAG. Association of Zoos and Aquariums Bear Taxon Advisory Group Regional Collection Plan. Available online: https://ams.aza.org/iweb/upload/RCP_Bear2016-0f78e49b.pdf (accessed on 6 April 2021).
- BearTAG. Association of Zoos and Aquariums Bear Taxon Advisory Group Annual Report. Available online: https://ams.aza.org/eweb/upload/Bear%20TAG%20Annual%20Report%202019-75c53476.pdf (accessed on 6 April 2021).
- Whitham, J.C.; Wielebnowski, N. New Directions for Zoo Animal Welfare Science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 48, 227–265. [Google Scholar] [CrossRef] [Green Version]
- Montaudouin, S.; Pape, G.L. Comparison between 28 Zoological Parks: Stereotypic and Social Behaviours of Captive Brown Bears (Ursus arctos). Appl. Anim. Behav. Sci. 2005, 92, 129–141. [Google Scholar] [CrossRef]
- Ross, M.R.; Niemann, T.; Wark, J.D.; Heintz, M.R.; Horrigan, A.; Cronin, K.A.; Shender, M.A.; Gillespie, K. ZooMonitor (Version 3). 2016. Available online: http://zoomonitor.org (accessed on 6 April 2021).
- Fuller, G.; Margulis, S.W.; Santymire, R. The Effectiveness of Indigestible Markers for Identifying Individual Animal Feces and Their Prevalence of Use in North American Zoos. Zoo Biol. 2011, 30, 379–398. [Google Scholar] [CrossRef]
- Palme, R.; Touma, C.; Arias, N.; Dominchin, M.F.; Lepschy, M. Steroid extraction: Get the best out of faecal samples. Wien. Tierarztl. Mon. 2013, 100, 238–246. [Google Scholar]
- Bjelakovic, G.; Beninati, S.; Pavlovic, D.; Kocic, G.; Jevtovic, T.; Kamenov, B.; Saranac, L.J.; Bjelakovic, B.; Stojanovic, I.; Basic, J. Glucocorticoids and Oxidative Stress. J. Basic Clin. Physiol. Pharmacol. 2007, 18, 115–127. [Google Scholar] [CrossRef]
- MacNicol, J.L.; Lindinger, M.I.; Pearson, W. A Time Course Evaluation of Inflammatory and Oxidative Markers Following High Intensity Exercise in Horses: A Pilot Study. J. Appl. Physiol. 2018, 124, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Tamashiro, K.L.K.; Nguyen, M.M.N.; Sakai, R.R. Social Stress: From Rodents to Primates. Front. Neuroendocrinol. 2005, 26, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Walker, S.; Lanthier, C.; Waddell, W.; Monfort, S.; Brown, J. Noninvasive Monitoring of Adrenocortical Activity in Carnivores by Fecal Glucocorticoid Analyses. Gen. Comp. Endocrinol. 2004, 137, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, M.; Mboumba, S.; Willaume, E.; Kappeler, P.M.; Charpentier, M.J.E. The Oxidative Cost of Unstable Social Dominance. J. Exp. Biol. 2014, 217, 2629. [Google Scholar] [CrossRef] [Green Version]
- Fialkowski, R.; Aufdemberge, P.; Wright, V.; Dijkstra, P. Radical change: Temporal patterns of oxidative stress during social ascent in a dominance hierarchy. Behav. Ecol. Sociobiol. 2021, 75, 43. [Google Scholar] [CrossRef]
- Kashif, S.M.; Zaidi, R.; Al-Qirim, T.M.; Hoda, N.; Banu, N. Modulation of Restraint Stress Induced Oxidative Changes in Rats by Antioxidant Vitamins. J. Nutr. Biochem. 2003, 14, 633–636. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping Styles in Animals: Current Status in Behavior and Stress Physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Costantini, D.; Ferrari, C.; Pasquaretta, C.; Cavallone, E.; Carere, C.; von Hardenberg, A.; Réale, D. Interplay Between Plasma Oxidative Status, Cortisol and Coping Styles in Wild Alpine Marmots, Marmota marmota. J. Exp. Biol. 2012, 215, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, D.; Carere, C.; Caramaschi, D.; Koolhaas, J.M. Aggressive and Non-Aggressive Personalities Differ in Oxidative Status in Selected Lines of Mice (Mus musculus). Biol. Lett. 2008, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Herzberg, D.; Strobel, P.; Chihuailaf, R.; Ramirez-Reveco, A.; Müller, H.; Werner, M.; Bustamante, H. Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows. Animals 2019, 9, 693. [Google Scholar] [CrossRef] [Green Version]
- Von Zglinicki, T. Role of Oxidative Stress in Telomere Length Regulation and Replicative Senescence. Ann. N. Y. Acad. Sci. 2000, 908, 99–110. [Google Scholar] [CrossRef]
- Whittemore, K.; Vera, E.; Martínez-Nevado, E.; Sanpera, C.; Blasco, M.A. Telomere Shortening Rate Predicts Species Life Span. Proc. Natl. Acad. Sci. USA 2019, 116, 15122–15127. [Google Scholar] [CrossRef] [Green Version]
- Haussmann, M.; Marchetto, N.M. Telomeres: Linking Stress and Survival, Ecology and Evolution. Curr. Zool. 2010, 56, 714–727. [Google Scholar] [CrossRef]
- Bateson, M. Cumulative Stress in Research Animals: Telomere Attrition as a Biomarker in a Welfare Context? Bioessays 2016, 38, 201–212. [Google Scholar] [CrossRef]
- Aydinonat, D.; Penn, D.; Smith, S.; Moodley, Y.; Hoelzl, F.; Knauer, F.; Schwarzenberger, F. Social Isolation Shortens Telomeres in African Grey Parrots (Psittacus erithacus erithacus). PLoS ONE 2014, 9, e93839. [Google Scholar] [CrossRef] [Green Version]
- Cooke, M.S.; Evans, M.D.; Dove, R.; Rozalski, R.; Gackowski, D.; Siomek, A.; Lunec, J.; Olinski, R. DNA Repair is Responsible for the Presence of Oxidatively Damaged DNA Lesions in Urine. Mutat. Res. 2005, 574, 58–66. [Google Scholar] [CrossRef]
- Gackowski, D.; Rozalski, R.; Roszkowski, K.; Jawien, A.; Foksiński, M.; Olinski, R. 8-Oxo-7,8-dihydroguanine and 8-oxo-7,8-dihydro-2′-deoxyguanosine Levels in Human Urine Do Not Depend on Diet. Free Radic. Res. 2001, 35, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Rozalski, R.; Siomek, A.; Gackowski, D.; Foksinski, M.; Gran, C.; Klungland, A.; Olinski, R. Diet Is Not Responsible for the Presence of Several Oxidatively Damaged DNA Lesions in Mouse Urine. Free Radic. Res. 2004, 38, 1201–1205. [Google Scholar] [CrossRef]
- Bogdanov, M.B.; McCabe, B.; McCabe, G.; Griffin Matson, W.R.; Matson, W.R. A Carbon Column-Based Liquid Chromatography Electrochemical Approach to Routine 8-Hydroxy-2′-Deoxyguanosine Measurements in Urine and Other Biologic Matrices: A One-Year Evaluation of Methods. Free Radic. Biol. Med. 1999, 27, 647–666. [Google Scholar] [CrossRef]
- Kato, I.; Startup, J.; Ram, J.L. Fecal Biomarkers for Research on Dietary and Lifestyle Risk Factors in Colorectal Cancer Etiology. Curr. Colorectal Cancer Rep. 2014, 10, 114–131. [Google Scholar] [CrossRef]
- Wilson, A.E.; Michaud, S.A.; Jackson, A.M.; Stenhouse, G.; Coops, N.C.; Janz, D.M. Development and Validation of Protein Biomarkers of Health in Grizzly Bears. Conserv. Physiol. 2020, 8, coaa056. [Google Scholar] [CrossRef]
- Munro, R.H.M.; Nielsen, S.E.; Price, M.H.; Stenhouse, G.B.; Boyce, M.S. Seasonal and Diel Patterns of Grizzly Bear Diet and Activity in West-Central Alberta. J. Mammal. 2006, 87, 1112–1121. [Google Scholar] [CrossRef]
- Christina, G.; Wasser, S.K.; Hunt, K.E.; Servheen, C. Factors Associated with Fecal Glucocorticoids in Alaskan Brown Bears (Ursus arctos horribilis). Physiol. Biochem. Zool. 2004, 77, 313–320. [Google Scholar]
- Loft, S.; Møller, P.; Cooke, M.S.; Rozalski, R.; Olinski, R. Antioxidant Vitamins and Cancer Risk: Is Oxidative Damage to DNA a Relevant Biomarker? Eur. J. Nutr. 2008, 47, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Stetz, J.; Hunt, K.; Kendall, K.C.; Wasser, S.K. Effects of Exposure, Diet, and Thermoregulation on Fecal Glucocorticoid Measures in Wild Bears. PLoS ONE 2013, 8, e55967. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, M.; Costantini, D. Biomarkers of oxidative status: Missing tools in conservation physiology. Conserv. Physiol. 2014, 2, cou014. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat (Relative Size) | Original (Medium) | Temporary (Smaller) | Expanded (Largest) |
|---|---|---|---|
| study period | 5 September 2017–11 November 2017 | 26 February 2018–30 May 2018 | 31 May 2018–30 September 2018 |
| outdoor habitat size | 836.1 m2 | 334.4 m2 | 1672.2 m2 |
| outdoor habitat features | gunite rockwork; dirt substrate; moat; two access points to holding; 246.0 m3 water feature with variable depths and a waterfall | gunite rockwork; dirt substrate; moat; two access points to holding; 64.3 m3 water feature | gunite rockwork; dirt, mulch, and grass substrates; several large trees; six access points to holding; large cave; same water feature as the original habitat (246.0 m3) |
| Behavior | Description |
|---|---|
| affiliative social interaction | Focal is engaged in an interaction with another bear including sniffing, licking, nuzzling, calm contact (bear initiating the contact is alert), passive recipient to such behavior, mouthing with no injuries, etc. |
| social play | Focal is engaged in a positive active interaction with another bear including wrestling, social chase, etc. Playful interactions should involve an alternating of offensive and defensive roles, and play should stop short of injury. |
| agonistic social interaction | Focal is engaged in a negative interaction with another bear including swatting, charging, biting, or attempting to bite, etc. |
| Predictor | log(FGM) | log(8-OHdG) | ||||||
|---|---|---|---|---|---|---|---|---|
| Subjects | Group | Boo | Mike | Thor | Group | Boo | Mike | Thor |
| log(FGM) | - | - | - | - | r = 0.492 p < 0.001 * n = 75 | r = 0.597 p = 0.011 * n = 17 | r = 0.472 p = 0.005 * n = 34 | r = 0.698 p < 0.001 * n = 24 |
| daily mean temperature | r = 0.134 p = 0.252 n = 75 | r = 0.646 p = 0.005 * n = 17 | r = 0.105 p = 0.554 n = 34 | r = 0.057 p = 0.790 n = 24 | r = 0.382 p = 0.001 * n = 75 | r = 0.155 p = 0.551 n = 17 | r = 0.504 p = 0.002 * n = 34 | r = 0.268 p = 0.205 n = 24 |
| log(rate affiliative) | r = 0.015 p = 0.907 n = 60 | r = −0.134 p = 0.609 n = 17 | r = 0.093 p = 0.705 n = 19 | r = 0.059 p = 0.785 n = 24 | r = 0.075 p = 0.567 n = 60 | r = −0.417 p = 0.096 n = 17 | r = 0.445 p = 0.056 n = 19 | r = −0.150 p = 0.484 n = 24 |
| log(rate social play) | r = −0.090 p = 0.496 n = 60 | r = 0.293 p = 0.253 n = 17 | r = −0.014 p = 0.956 n = 19 | r = −0.220 p = 0.302 n = 24 | r = −0.074 p = 0.574 n = 60 | r = 0.178 p = 0.495 n = 17 | r = −0.072 p = 0.768 n = 19 | r = −0.250 p = 0.239 n = 24 |
| log(rate agonistic) | r = −0.138 p = 0.292 n = 60 | r = −0.019 p = 0.942 n = 17 | r = −0.166 p = 0.498 n = 19 | r = −0.176 p = 0.410 n = 24 | r = 0.071 p = 0.589 n = 60 | r = 0.200 p = 0.442 n = 17 | r = −0.144 p = 0.556 n = 19 | r = 0.045 p = 0.836 n = 24 |
| percent active | r = 0.265 p = 0.041 * n = 60 | r = −0.238 p = 0.358 n = 17 | r = 0.442 p = 0.058 n = 19 | r = 0.363 p = 0.082 n = 24 | r = 0.091 p = 0.490 n = 60 | r = −0.170 p = 0.513 n = 17 | r = 0.137 p = 0.575 n = 19 | r = 0.376 p = 0.071 n = 24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuller, G.; Hamilton, J.; Allard, S. DNA Damage as a Potential Non-Invasive Indicator of Welfare: A Preliminary Study in Zoo-Housed Grizzly Bears (Ursus arctos horribilis). J. Zool. Bot. Gard. 2021, 2, 316-334. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030022
Fuller G, Hamilton J, Allard S. DNA Damage as a Potential Non-Invasive Indicator of Welfare: A Preliminary Study in Zoo-Housed Grizzly Bears (Ursus arctos horribilis). Journal of Zoological and Botanical Gardens. 2021; 2(3):316-334. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030022
Chicago/Turabian StyleFuller, Grace, Jennifer Hamilton, and Stephanie Allard. 2021. "DNA Damage as a Potential Non-Invasive Indicator of Welfare: A Preliminary Study in Zoo-Housed Grizzly Bears (Ursus arctos horribilis)" Journal of Zoological and Botanical Gardens 2, no. 3: 316-334. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030022