
Investigation of Environmentally Dependent Movement of Bottlenose Dolphins (Tursiops truncatus)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
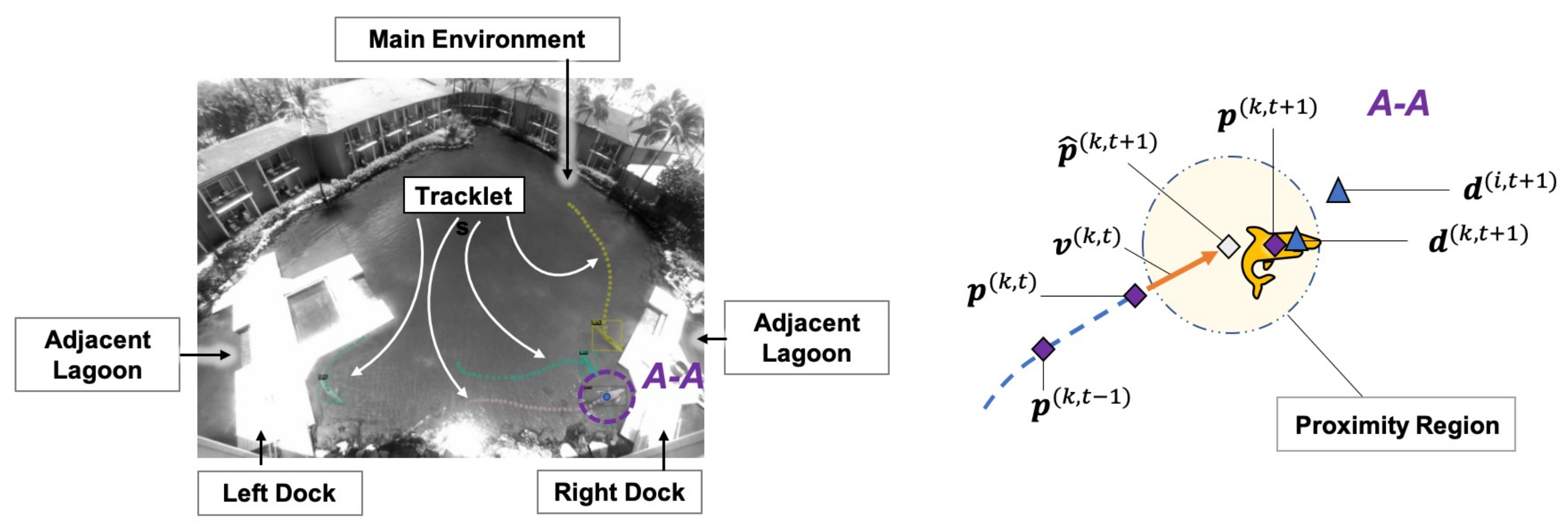
2.1. Environment Setups
2.2. Target Detection
2.3. Tracklet Formation and Kinematics
2.4. Movement in the Environment
2.4.1. Occupancy Heatmaps and Flow in the Environment
2.4.2. Probability Density
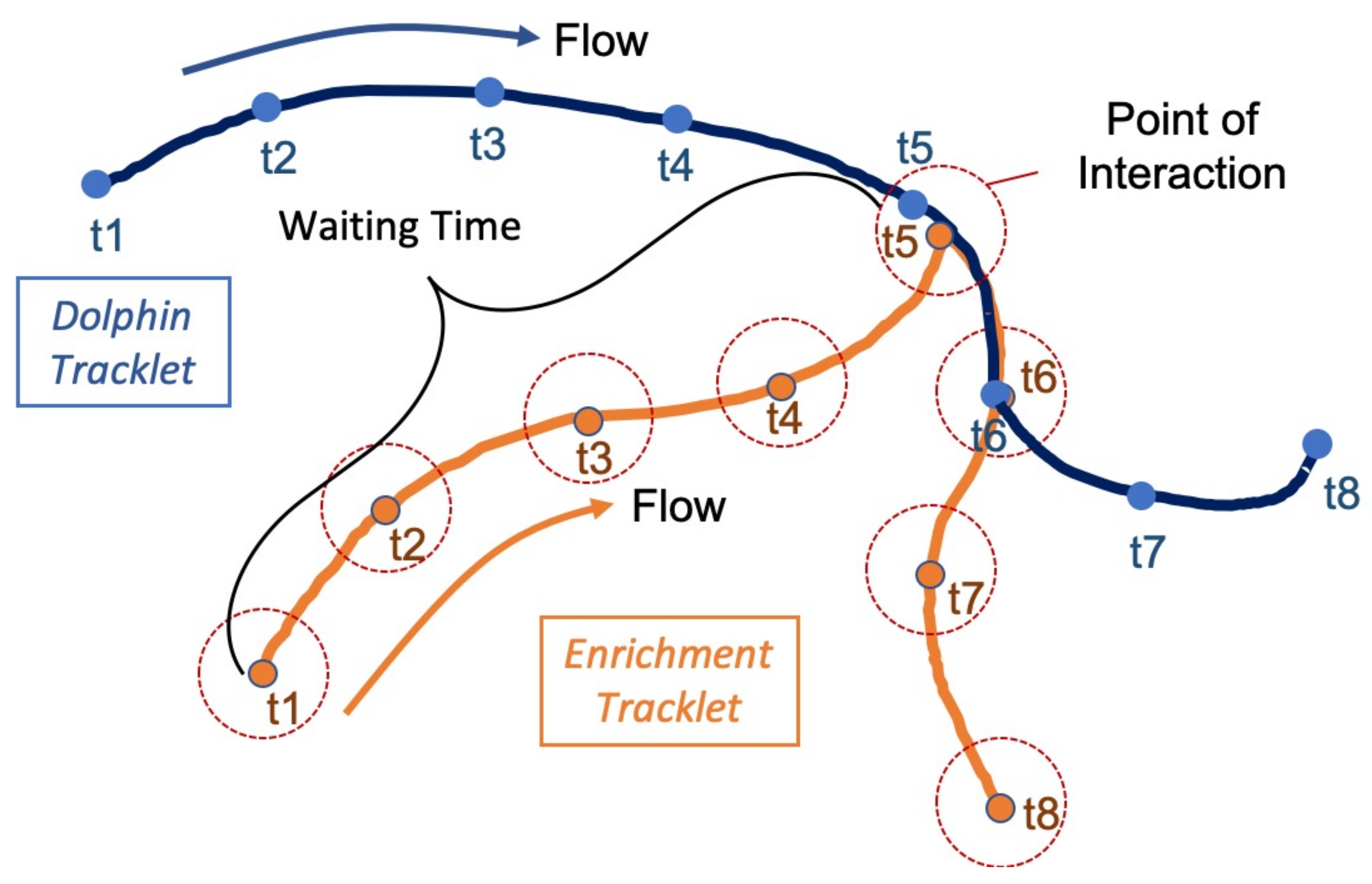
2.5. Engaging with Enrichment
3. Results
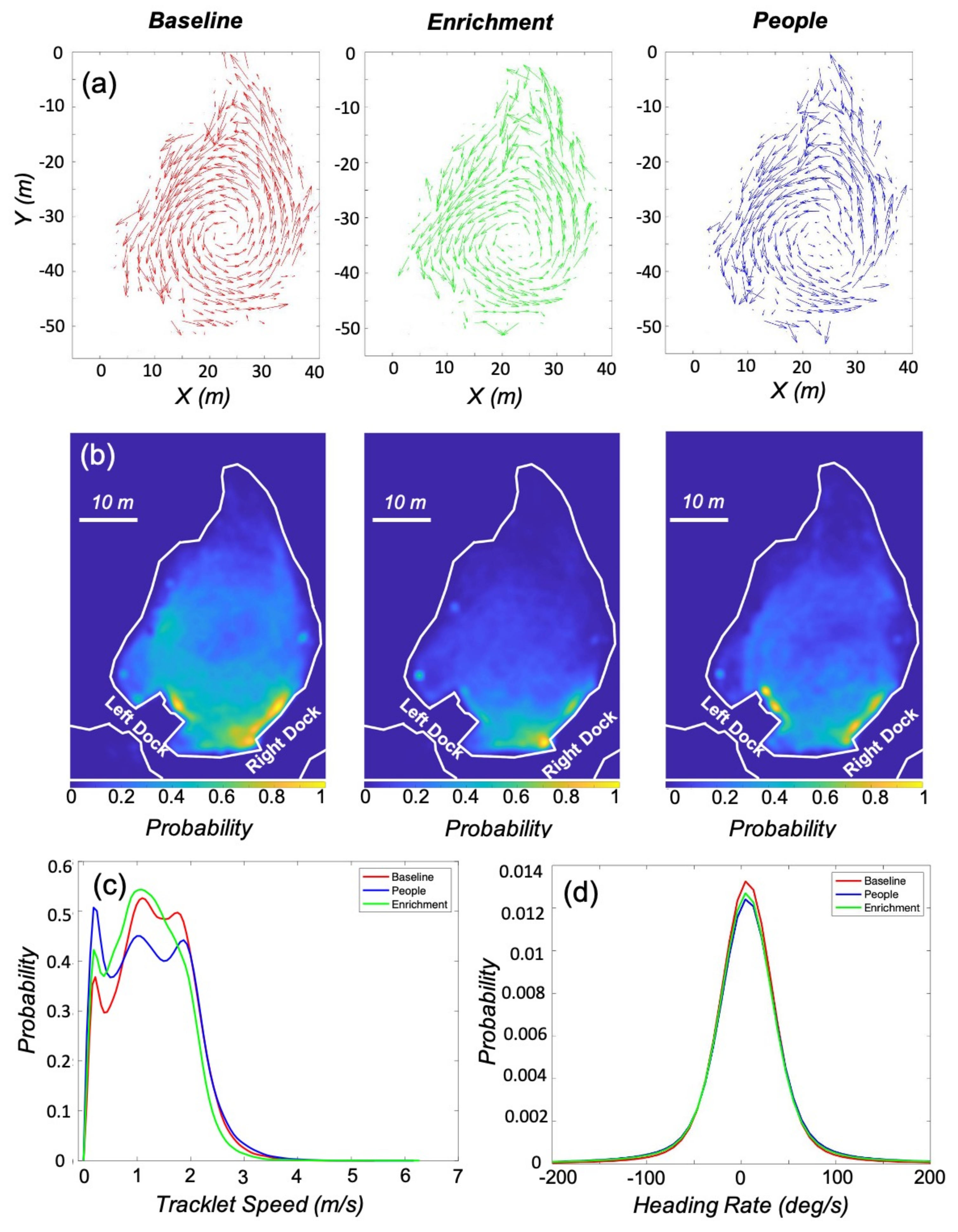
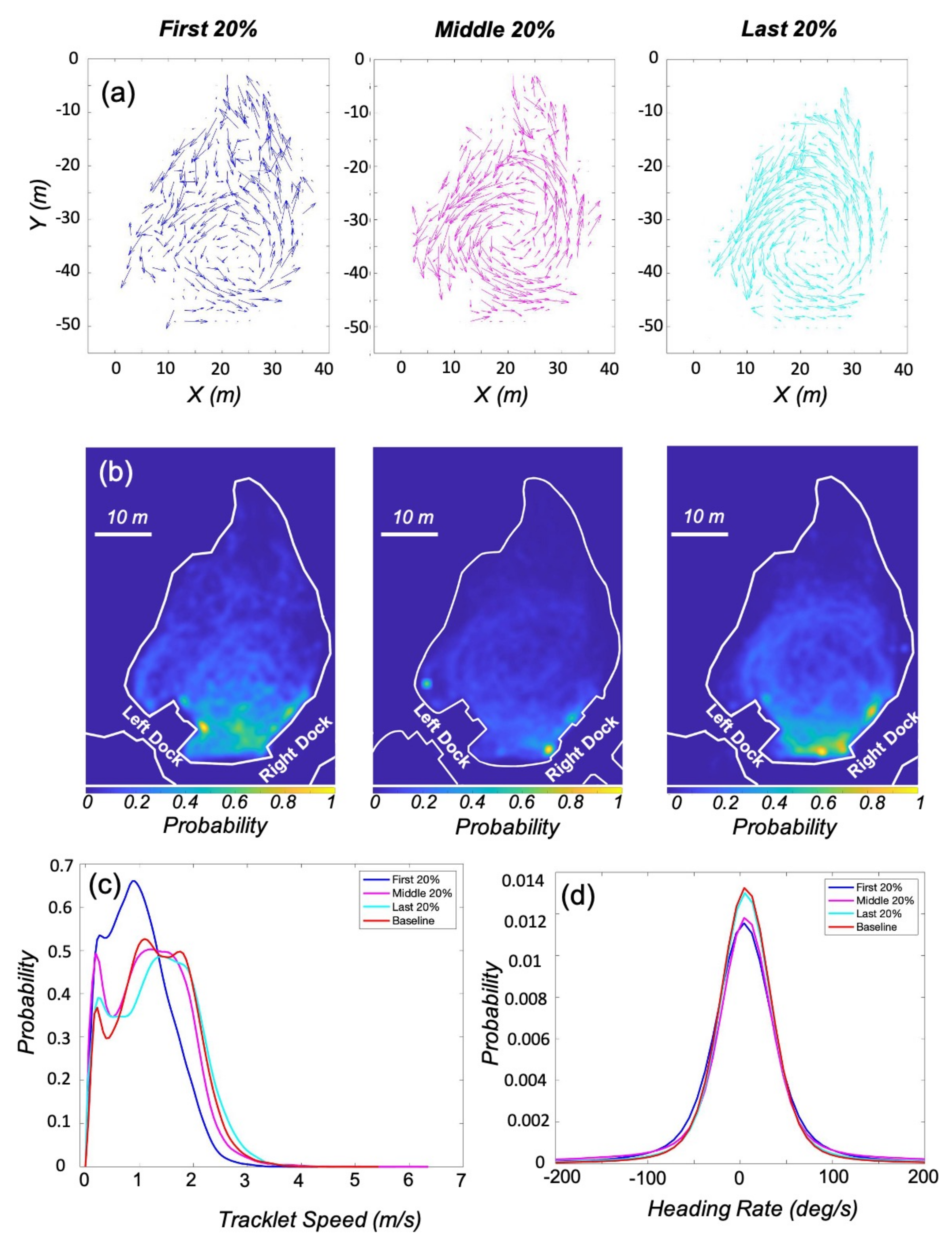
3.1. Kinematic Analysis
3.2. Enrichment
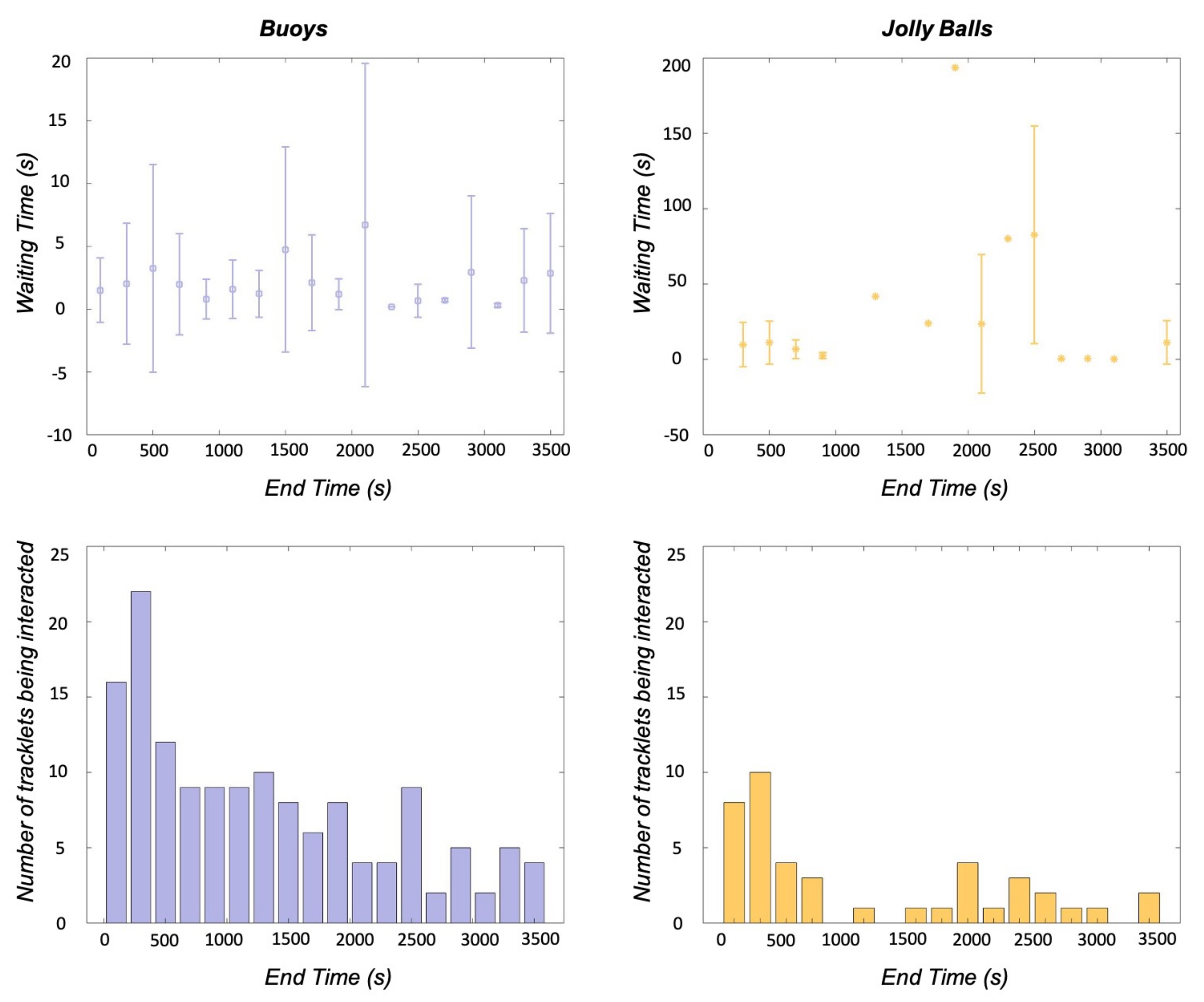
3.3. Enrichment Engagement
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kagan, R.; Carter, S.; Allard, S. A Universal Animal Welfare Framework for Zoos. J. Appl. Anim. Welf. Sci. 2015, 18 (Suppl. 1), S1–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, J.C.; Wielebnowski, N. New Directions for Zoo Animal Welfare Science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Daan, S. Adaptive Daily Strategies in Behavior. In Biological Rhythms; Springer: Boston, MA, USA, 1981; pp. 275–298. [Google Scholar]
- Stamps, J.A. Individual Differences in Behavioural Plasticities: Behavioural Plasticities. Biol. Rev. Camb. Philos. Soc. 2016, 191, 534–567. [Google Scholar] [CrossRef]
- Karnowski, J.; Hutchins, E.; Johnson, C. Dolphin Detection and Tracking. In Proceedings of the 2015 IEEE Winter Applications and Computer Vision Workshops, Waikoloa, HI, USA, 5–9 January 2015. [Google Scholar]
- Waitt, C.; Buchanan-Smith, H.M. What Time Is Feeding? Appl. Anim. Behav. Sci. 2001, 75, 75–85. [Google Scholar] [CrossRef]
- Mason, G.J. Species Differences in Responses to Captivity: Stress, Welfare and the Comparative Method. Trends Ecol. Evol. 2010, 25, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcbride, A.F.; Hebb, D.O. Behavior of the Captive Bottle-Nose Dolphin, Tursiops Truncatus. J. Comp. Physiol. Psychol. 1948, 41, 111–123. [Google Scholar] [CrossRef]
- Johnson, M.P.; Tyack, P.L. A Digital Acoustic Recording Tag for Measuring the Response of Wild Marine Mammals to Sound. IEEE J. Ocean. Eng. 2003, 28, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Shorter, K.A.; Rocho-Levine, J.; van der Hoop, J.; Moore, M.; Barton, K. Behavior Inference from Bio-Logging Sensors: A Systematic Approach for Feature Generation, Selection and State Classification. In Proceedings of the ASME 2018 Dynamic Systems and Control Conference, Atlanta, GA, USA, 30 September–3 October 2018; Volume 2. [Google Scholar]
- Van der Hoop, J.M.; Fahlman, A.; Hurst, T.; Rocho-Levine, J.; Shorter, K.A.; Petrov, V.; Moore, M.J. Bottlenose Dolphins Modify Behavior to Reduce Metabolic Effect of Tag Attachment. J. Exp. Biol. 2014, 217 Pt 23, 4229–4236. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Hoop, J.M.; Petrov, V.; Rocho-Levine, J.; Moore, M.J.; Shorter, K.A. Simulated and Experimental Estimates of Hydrodynamic Drag from Bio-logging Tags. Mar. Mamm. Sci. 2020, 36, 136–157. [Google Scholar] [CrossRef]
- Sibal, R.; Zhang, D.; Rocho-Levine, J.; Shorter, K.A.; Barton, K. Bidirectional LSTM Recurrent Neural Network plus Hidden Markov Model for Wearable Sensor-Based Dynamic State Estimation. ASME Lett. Dyn. Syst. Control 2021, 1, 1–6. [Google Scholar] [CrossRef]
- Fish, F.E.; Legac, P.; Williams, T.M.; Wei, T. Measurement of Hydrodynamic Force Generation by Swimming Dolphins Using Bubble DPIV. J. Exp. Biol. 2014, 217 Pt 2, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Gabaldon, J.; Zhang, D.; Barton, K.; Johnson-Roberson, M.; Shorter, K.A. A Framework for Enhanced Localization of Marine Mammals Using Auto-Detected Video and Wearable Sensor Data Fusion. In Proceedings of the 2017 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Vancouver, BC, Canada, 24–28 September 2017. [Google Scholar]
- Zhang, D.; Gabaldon, J.; Lauderdale, L.; Johnson-Roberson, M.; Miller, L.J.; Barton, K.; Shorter, K.A. Localization and Tracking of Uncontrollable Underwater Agents: Particle Filter Based Fusion of on-Body IMUs and Stationary Cameras. In Proceedings of the 2019 International Conference on Robotics and Automation (ICRA), Montreal, QC, Canada, 20–24 May 2019. [Google Scholar]
- Tanaka, H.; Li, G.; Uchida, Y.; Nakamura, M.; Ikeda, T.; Liu, H. Measurement of Time-Varying Kinematics of a Dolphin in Burst Accelerating Swimming. PLoS ONE 2019, 14, e0210860. [Google Scholar] [CrossRef]
- Rachinas-Lopes, P.; Ribeiro, R.; Dos Santos, M.E.; Costa, R.M. D-Track-A Semi-Automatic 3D Video-Tracking Technique to Analyse Movements and Routines of Aquatic Animals with Application to Captive Dolphins. PLoS ONE 2018, 13, e0201614. [Google Scholar] [CrossRef] [PubMed]
- van der Hoop, J.M.; Fahlman, A.; Shorter, K.A.; Gabaldon, J.; Rocho-Levine, J.; Petrov, V.; Moore, M.J. Swimming Energy Economy in Bottlenose Dolphins under Variable Drag Loading. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- He, K.; Gkioxari, G.; Dollar, P.; Girshick, R. Mask R-CNN. In Proceedings of the IEEE International Conference on Computer Vision, Venice, Italy, 22–29 October 2017. [Google Scholar]
- Ren, S.; He, K.; Girshick, R.; Sun, J. Faster R-CNN: Towards Real-Time Object Detection with Region Proposal Networks. arXiv 2015, arXiv:1506.01497. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Zisserman, A. The VIA Annotation Software for Images, Audio and Video. In Proceedings of the 27th ACM International Conference on Multimedia, Nice, France, 21–25 October 2015; ACM: New York, NY, USA, 2019. [Google Scholar]
- Gabaldon, J.; Zhang, D.; Lauderdale, L.; Miller, L.; Johnson-Roberson, M.; Barton, K.; Shorter, K.A. Vision-based monitoring and measurement of bottlenose dolphins’ daily habitat use and kinematics. bioRxiv 2021. Forthcoming. [Google Scholar]
- Berger, V.W.; Zhou, Y. Kolmogorov—Smirnov test: Overview. In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014. [Google Scholar]
- Sobel, N.; Supin, A.Y.; Myslobodsky, M.S. Rotational Swimming Tendencies in the Dolphin (Tursiops Truncatus). Behav. Brain Res. 1994, 65, 41–45. [Google Scholar] [CrossRef]
- Stafne, G.M.; Manger, P.R. Predominance of Clockwise Swimming during Rest in Southern Hemisphere Dolphins. Physiol. Behav. 2004, 82, 919–926. [Google Scholar] [CrossRef]
- Kuczaj, S.; Lacinak, T.; Fad, O.; Trone, M.; Solangi, M.; Ramos, J. Keeping environmental enrichment enriching. Int. J. Comp. Psychol. 2002, 15, 127–137. [Google Scholar]
- Harley, H.E.; Fellner, W.; Stamper, M.A. Cognitive research with dolphins (Tursiops truncatus) at Disney’s The Seas: A program for enrichment, science education, and conservation. Int. J. Comp. Psychol. 2010, 23, 331–343. [Google Scholar]
- Lauderdale, L.K.; Miller, L.J. Common Bottlenose Dolphin (Tursiops Truncatus) Problem Solving Strategies in Response to a Novel Interactive Apparatus. Behav. Process. 2019, 169, 103990. [Google Scholar] [CrossRef] [PubMed]
- Lauderdale, L.K.; Miller, L.J. Efficacy of an Interactive Apparatus as Environmental Enrichment for Common Bottlenose Dolphins (Tursiops truncatus). Anim. Welf. 2020, 29, 379–386. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter (Condition) | Mean | Standard Deviation | K-S Test Statistic | Critical Values |
|---|---|---|---|---|
| Speed (Baseline) | 1.20 m/s | 0.74 | 0 | 0 |
| Speed (Enrichment) | 1.01 m/s | 0.73 | 0.07 | 0.0036 |
| Speed (People) | 1.09 m/s | 0.81 | 0.08 | 0.0037 |
| Heading Rate (Baseline) | 5.63 deg/s | 43.01 | 0 | 0 |
| Heading Rate (Enrichment) | 5.33 deg/s | 54.65 | 0.03 | 0.0036 |
| Heading Rate (People) | 5.94 deg/s | 55.18 | 0.04 | 0.0037 |
| Parameter (Condition) | Mean | Standard Deviation | K-S Test Statistic | Critical Values |
|---|---|---|---|---|
| Speed (First 20%) | 1.00 m/s | 0.57 | 0.21 | 0.0064 |
| Speed (Middle 20%) | 1.20 m/s | 0.69 | 0.06 | 0.0058 |
| Speed (Last 20%) | 1.33 m/s | 0.72 | 0.03 | 0.0062 |
| Heading Rate (First 20%) | 4.26 deg/s | 54.89 | 0.07 | 0.0064 |
| Heading Rate (Middle 20%) | 5.56 deg/s | 74.04 | 0.05 | 0.0058 |
| Heading Rate (Last 20%) | 5.95 deg/s | 52.77 | 0.01 | 0.0062 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, D.; Gabaldon, J.; Goodbar, K.; West, N.; Barton, K.; Shorter, K.A. Investigation of Environmentally Dependent Movement of Bottlenose Dolphins (Tursiops truncatus). J. Zool. Bot. Gard. 2021, 2, 335-348. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030023
Zhang Z, Zhang D, Gabaldon J, Goodbar K, West N, Barton K, Shorter KA. Investigation of Environmentally Dependent Movement of Bottlenose Dolphins (Tursiops truncatus). Journal of Zoological and Botanical Gardens. 2021; 2(3):335-348. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030023
Chicago/Turabian StyleZhang, Zining, Ding Zhang, Joaquin Gabaldon, Kari Goodbar, Nicole West, Kira Barton, and Kenneth Alex Shorter. 2021. "Investigation of Environmentally Dependent Movement of Bottlenose Dolphins (Tursiops truncatus)" Journal of Zoological and Botanical Gardens 2, no. 3: 335-348. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg2030023