
Diet and Life-History Traits of Savannah Dwelling Waterbirds in Southern Africa: Implications for Their Conservation Status

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Clusters on Life History Traits
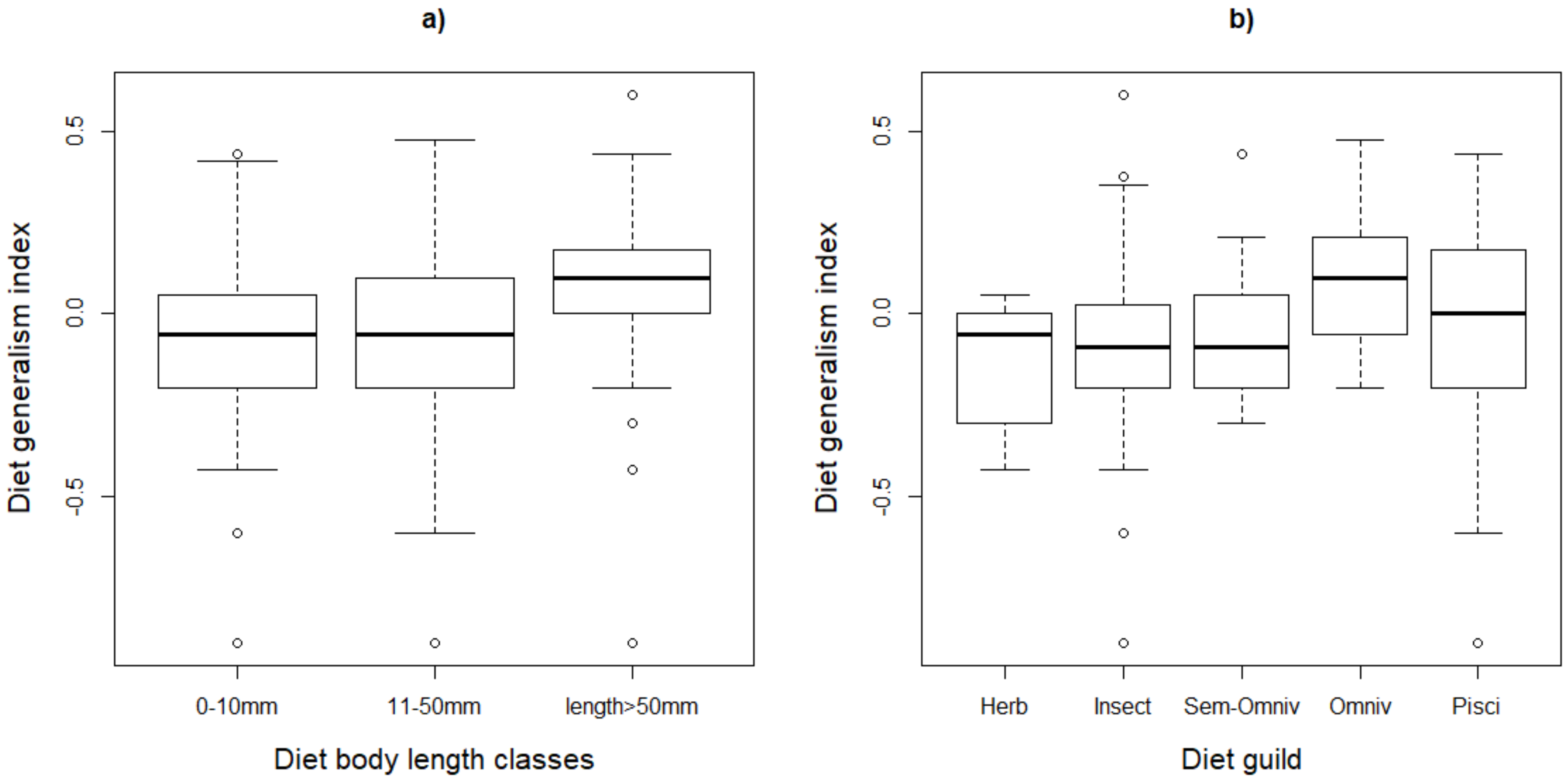
3.2. Diet and Life History Traits in Relation to Population Trends
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

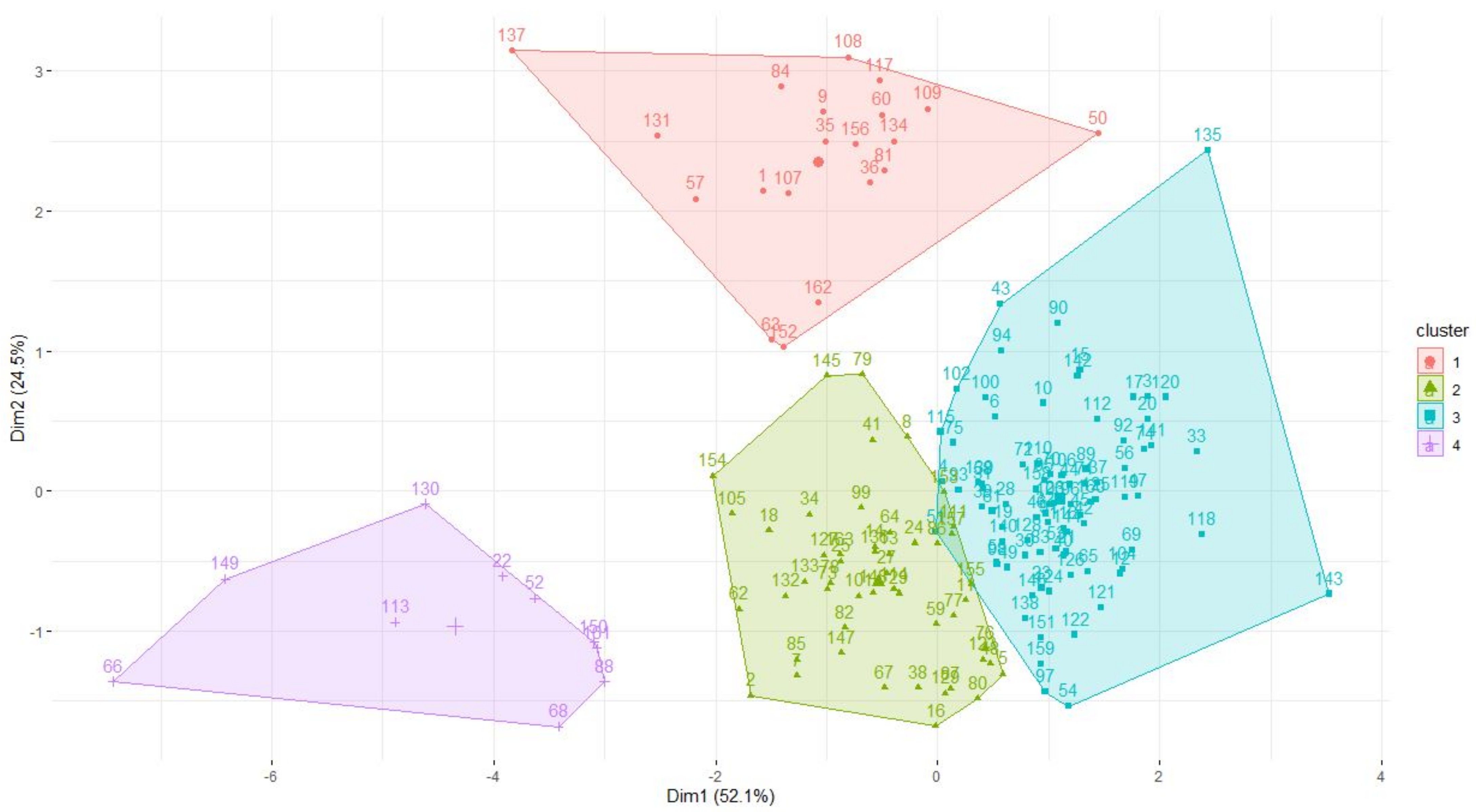{kind=link}
{kind=link}
| Cluster | Code | Local Name | Scientific Name | Diet Guild | Global Status |
|---|---|---|---|---|---|
| 1 | 3 | African Crake | Crex egregia | Semi-omn | Not Dec |
| 4 | African Darter | Anhinga rufa | Piscivores | Dec | |
| 6 | African Jacana | Actophilornis africanus | Insectivores | Not Dec | |
| 10 | African Rail | Rallus caerulescens | Omnivores | Not Dec | |
| 12 | African Snipe | Gallinago nigripennis | Insectivores | Dec | |
| 15 | Allen’s Gallinule | Porphyrio alleni | Omnivores | Dec | |
| 17 | Baillon’s Crake | Porzana pusilla | Insectivores | Dec | |
| 19 | Bar-tailed Godwit | Limosa lapponica | Omnivores | Dec | |
| 20 | Black Crake | Zapornia flavirostra | Omnivores | Dec | |
| 21 | Black Heron | Egretta ardesiaca | Piscivores | Not Dec | |
| 23 | Black Tern | Chlidonias niger | Piscivores | Dec | |
| 26 | Black-necked Grebe | Podiceps nigricollis | Insectivores | Dec | |
| 28 | Black-tailed Godwit | Limosa limosa | Omnivores | Dec | |
| 30 | Black-winged Pratincole | Glareola nordmanni | Insectivores | Dec | |
| 31 | Black-winged Stilt | Himantopus himantopus | Insectivores | Not Dec | |
| 32 | Bronze-winged Courser | Rhinoptilus chalcopterus | Insectivores | Not Dec | |
| 33 | Buff-spotted Flufftail | Sarothrura elegans | Semi-omn | Not Dec | |
| 37 | Caspian Plover | Charadrius asiaticus | Semi-omn | Dec | |
| 39 | Cattle Egret | Bubulcus ibis | Insectivores | Not Dec | |
| 40 | Chestnut-banded Plover | Charadrius pallidus | Insectivores | Not Dec | |
| 42 | Common Greenshank | Tringa nebularia | Insectivores | Not Dec | |
| 43 | Common Moorhen | Gallinula chloropus | Omnivores | Not Dec | |
| 44 | Common Redshank | Tringa totanus | Insectivores | Dec | |
| 45 | Common Ringed Plover | Charadrius hiaticula | Insectivores | Dec | |
| 46 | Common Sandpiper | Actitis hypoleucos | Omnivores | Dec | |
| 47 | Common Snipe | Gallinago gallinago | Omnivores | Dec | |
| 49 | Common Whimbrel | Numenius phaeopus | Insectivores | Dec | |
| 51 | Crowned Cormorant | Microcarbo coronatus | Piscivores | Not Dec | |
| 53 | Curlew Sandpiper | Calidris ferruginea | Insectivores | Not Dec | |
| 54 | Damara Tern | Sternula balaenarum | Piscivores | Dec | |
| 55 | Dimorphic egret | Egretta garzetta dimorpha | Piscivores | Not Dec | |
| 56 | Dwarf Bittern | Ixobrychus sturmii | Piscivores | Dec | |
| 58 | Eurasian Curlew | Numenius arquata | Omnivores | Dec | |
| 61 | Glossy Ibis | Plegadis falcinellus | Insectivores | Dec | |
| 65 | Great Snipe | Gallinago media | Insectivores | Dec | |
| 69 | Greater Painted Snipe | Rostratula benghalensis | Omnivores | Dec | |
| 70 | Greater Sandplover | Charadrius leschenaultii | Insectivores | Not Dec | |
| 71 | Green Sandpiper | Tringa ochropus | Omnivores | Not Dec | |
| 72 | Green-backed Heron | Butorides striata | Piscivores | Dec | |
| 74 | Grey Phalarope | Phalaropus fulicarius | Semi-omn | Not Dec | |
| 75 | Grey Plover | Pluvialis squatarola | Insectivores | Dec | |
| 83 | Kitlitz Plover | Charadrius pecuarius | Insectivores | Dec | |
| 89 | Lesser Jacana | Microparra capensis | Insectivores | Dec | |
| 90 | Lesser Moorhen | Gallinula angulata | Semi-omn | Dec | |
| 91 | Lesser Sandplover | Charadrius mongolus | Insectivores | Not Dec | |
| 92 | Little Bittern | Ixobrychus minutus | Piscivores | Dec | |
| 93 | Little Egret | Egretta garzetta | Piscivores | Not Dec | |
| 94 | Little Grebe | Tachybaptus ruficollis | Insectivores | Dec | |
| 95 | Little Ringed Plover | Charadrius dubius | Insectivores | Dec | |
| 96 | Little Stint | Calidris minuta | Omnivores | Dec | |
| 97 | Little Tern | Sternula albifrons | Piscivores | Dec | |
| 98 | Long-tailed Cormorant | Microcarbo africanus | Piscivores | Dec | |
| 100 | Maccoa Duck | Oxyura maccoa | Omnivores | Dec | |
| 102 | Madagascar Jacana | Actophilornis albinucha | Semi-omn | Dec | |
| 103 | Madagascar Pond Heron | Ardeola idae | Piscivores | Dec | |
| 104 | Madagascar Snipe | Gallinago macrodactyla | Semi-omn | Dec | |
| 106 | Marsh Sandpiper | Tringa stagnatilis | Insectivores | Dec | |
| 110 | Pacific Golden Plover | Pluvialis fulva | Insectivores | Dec | |
| 112 | Pied Kingfisher | Ceryle rudis | Piscivores | Dec | |
| 115 | Red Knobbed Coot | Fulica cristata | Semi-omn | Dec | |
| 116 | Red Knot | Calidris canutus | Omnivores | Dec | |
| 118 | Red-chested Flufftail | Sarothrura rufa | Omnivores | Dec | |
| 119 | Red-necked Phalarope | Phalaropus lobatus | Omnivores | Dec | |
| 120 | Red-tailed Flufftail | Sarothrura affinis | Semi-omn | Dec | |
| 121 | Red-winged Pranticole | Glareola pratincola | Insectivores | Dec | |
| 122 | Rock Pratincole | Glareola nuchalis | Insectivores | Dec | |
| 124 | Ruddy Turnstone | Arenaria interpres | Omnivores | Dec | |
| 125 | Ruff | Philomachus pugnax | Omnivores | Dec | |
| 126 | Rufous-bellied Heron | Ardeola rufiventris | Piscivores | Dec | |
| 128 | Sanderling | Calidris alba | Insectivores | Not Dec | |
| 135 | Spotted Crake | Porzana porzana | Omnivores | Not Dec | |
| 138 | Spur-winged Lapwing | Vanellus spinosus | Semi-omn | Not Dec | |
| 139 | Squacco Heron | Ardeola ralloides | Piscivores | Dec | |
| 140 | Staty Egret | Egretta vinaceigula | Piscivores | Dec | |
| 141 | Streaky-breasted Flufftail | Sarothrura boehmi | Herbivores | Dec | |
| 142 | Stripped Crake | Amaurornis marginalis | Insectivores | Dec | |
| 143 | Temminck Courser | Cursorius temminckii | Semi-omn | Not Dec | |
| 144 | Terek Sandpiper | Xenus cinereus | Omnivores | Dec | |
| 146 | Three-banded Plover | Charadrius tricollaris | Insectivores | Dec | |
| 151 | White Winged Tern | Chlidonias leucopterus | Insectivores | Not Dec | |
| 158 | White-throated Rail | Dryolimnas cuvieri | Omnivores | Not Dec | |
| 159 | Wiskered Tern | Chlidonias hybrida | Piscivores | Not Dec | |
| 160 | Wood Sandpiper | Tringa glareola | Omnivores | Not Dec | |
| 2 | 2 | African Black Oystercatcher | Haematopus moquini | Insectivores | Not Dec |
| 5 | African Finfoot | Podica senegalensis | Insectivores | Dec | |
| 7 | African Openbill Stork | Anastomus lamelligerus | Piscivores | Dec | |
| 8 | African Purple Swamphen | Porphyrio porphyrio | Omnivores | Dec | |
| 11 | African Skimmer | Rynchops flavirostris | Piscivores | Dec | |
| 13 | African Spoonbill | Platalea alba | Insectivores | Not Dec | |
| 14 | African Wattled Lapwing | Vanellus senegallus | Semi-omn | Not Dec | |
| 16 | Arctic Tern | Sterna paradisaea | Piscivores | Dec | |
| 18 | Bank Cormorant | Phalacrocorax neglectus | Piscivores | Dec | |
| 24 | Black-crowned Night Heron | Nycticorax nycticorax | Omnivores | Dec | |
| 25 | Black-headed Heron | Ardea melanocephala | Insectivores | Not Dec | |
| 27 | Blacksmith Lapwing | Vanellus armatus | Insectivores | Not Dec | |
| 29 | Black-winged Lapwing | Vanellus melanopterus | Insectivores | Dec | |
| 34 | Cape Cormorant | Phalacrocorax capensis | Piscivores | Dec | |
| 38 | Caspian Tern | Hydroprogne caspia | Piscivores | Dec | |
| 41 | Common Bittern | Botaurus stellaris | Piscivores | Dec | |
| 48 | Common Tern | Sterna hirundo | Piscivores | Not Dec | |
| 59 | Franklin’s Gull | Larus pipixcan | Insectivores | Not Dec | |
| 62 | Goliath Heron | Ardea goliath | Piscivores | Not Dec | |
| 64 | Great Egret | Ardea alba | Piscivores | Dec | |
| 67 | Greater Crested Tern | Thalasseus bergii | Piscivores | Not Dec | |
| 73 | Grey Heron | Ardea cinerea | Piscivores | Dec | |
| 76 | Grey-hooded Gull | Chroicocephalus cirrocephalus | Piscivores | Not Dec | |
| 77 | Gull-billed Tern | Gelochelidon nilotica | Insectivores | Dec | |
| 78 | Hadeda Ibis | Bostrychia hagedash | Insectivores | Not Dec | |
| 79 | Hamerkop | Scopus umbretta | Piscivores | Not Dec | |
| 80 | Hartlaub’s Gull | Chroicocephalus hartlaubii | Piscivores | Not Dec | |
| 82 | Intermediate Egret | Ardea intermedia | Piscivores | Dec | |
| 85 | Lesser Black-backed Gull | Larus fuscus | Piscivores | Not Dec | |
| 86 | Lesser Black-winged Plover | Vanellus lugubris | Semi-omn | Not Dec | |
| 87 | Lesser Crested Tern | Thalasseus bengalensis | Piscivores | Not Dec | |
| 99 | Long-toed Lapwing | Vanellus crassirostris | Semi-omn | Dec | |
| 101 | Madagascar Heron | Ardea humbloti | Piscivores | Dec | |
| 105 | Madagascar Teal | Anas bernieri | Insectivores | Dec | |
| 111 | Pied Avocet | Recurvirostra avosetta | Omnivores | Dec | |
| 114 | Purple Heron | Ardea purpurea | Piscivores | Dec | |
| 123 | Roseate Tern | Sterna dougallii | Piscivores | Not Dec | |
| 127 | Sacred Ibis | Threskiornis aethiopicus | Insectivores | Dec | |
| 129 | Sandwich Tern | Thalasseus sandvicensis | Piscivores | Not Dec | |
| 132 | Southern Bald Ibis | Geronticus calvus | Insectivores | Dec | |
| 133 | Southern Black-backed Gull | Larus dominicanus | Piscivores | Not Dec | |
| 136 | Spotted Thickknee | Burhinus capensis | Insectivores | Not Dec | |
| 145 | Three-banded Courser | Rhinoptilus cinctus | Insectivores | Not Dec | |
| 147 | Eurasian Oystercatcher | Haematopus ostralegus | Insectivores | Dec | |
| 148 | Water Thickknee | Burhinus vermiculatus | Omnivores | Dec | |
| 153 | White-backed Night Heron | Gorsachius leuconotus | Piscivores | Not Dec | |
| 154 | White-breasted Cormorant | Phalacrocorax carbo | Piscivores | Not Dec | |
| 155 | White-crowned Lapwing | Vanellus albiceps | Semi-omn | Not Dec | |
| 157 | White-fronted Plover | Charadrius marginatus | Insectivores | Dec | |
| 163 | Yellow-billed Stork | Mycteria ibis | Piscivores | Dec | |
| 3 | 22 | Black Stork | Ciconia nigra | Insectivores | Dec |
| 52 | Crowned Crane | Balearica regulorum | Semi-omn | Dec | |
| 66 | Great White Pelican | Pelecanus onocrotalus | Piscivores | Dec | |
| 68 | Greater Flamingo | Phoenicopterus roseus | Piscivores | Not Dec | |
| 88 | Lesser Flamingo | Phoeniconaias minor | Insectivores | Dec | |
| 113 | Pink-backed Pelican | Pelecanus rufescens | Piscivores | Not Dec | |
| 130 | Shoebill | Balaeniceps rex | Piscivores | Dec | |
| 149 | Wattled Crane | Grus carunculate | Omnivores | Dec | |
| 150 | White Stork | Ciconia ciconia | Piscivores | Not Dec | |
| 161 | Woolly necked Stork | Ciconia episcopus | Insectivores | Dec | |
| 4 | 1 | African Black Duck | Anas sparsa | Omnivores | Dec |
| 9 | African Pygmy Goose | Nettapus auritus | Herbivores | Dec | |
| 35 | Cape Shoveler | Spatula smithii | Omnivores | Not Dec | |
| 36 | Cape Teal | Anas capensis | Omnivores | Not Dec | |
| 50 | Corn Crake | Crex crex | Semi-omn | Not Dec | |
| 57 | Egyptian Geese | Alopochen aegyptiaca | Herbivores | Dec | |
| 60 | Fulvous Duck | Dendrocygna bicolor | Herbivores | Dec | |
| 63 | Great Crested Grebe | Podiceps cristatus | Insectivores | Not Dec | |
| 81 | Hottentot Teal | Spatula hottentota | Omnivores | Dec | |
| 84 | Knob-billed Duck | Sarkidiornis melanotos | Herbivores | Dec | |
| 107 | Meller’s Duck | Anas melleri | Herbivores | Dec | |
| 108 | Northern Mallard | Anas platyrhynchos | Omnivores | Not Dec | |
| 109 | Northern Shoveler | Spatula clypeata | Insectivores | Not Dec | |
| 117 | Red-billed Teal | Anas erythrorhyncha | Omnivores | Dec | |
| 131 | South African Shelduck | Tadorna cana | Herbivores | Not Dec | |
| 134 | Southern Pochard | Netta erythrophthalma | Herbivores | Dec | |
| 137 | Spur-winged Goose | Plectropterus gambensis | Herbivores | Not Dec | |
| 152 | White-backed Duck | Thalassornis leuconotus | Herbivores | Dec | |
| 156 | White-faced Duck | Dendrocygna viduata | Semi-omn | Not Dec | |
| 162 | Yellow-billed Duck | Anas undulata | Omnivores | Not Dec |
Appendix B
| Model | K | AICc | ΔAICc | Wi |
|---|---|---|---|---|
| Global trend~Diet body size | 3 | 208.724 | 0 | 0.788 |
| Global trend~Waterbird cluster + diet Body size | 6 | 212.534 | 3.81 | 0.117 |
| Global trend~Waterbird guild + number of prey items consumed + diet body size | 8 | 213.024 | 4.299 | 0.092 |
References
- Hockey, P.; Dean, W.; Ryan, P. Roberts Birds of Southern Africa, 7th ed.; Trustees of the John Voelcker Bird Book Fund: Cape Town, South Africa, 2005. [Google Scholar]
- Henry, D.A.; Cumming, G.S. Can waterbirds with different movement, dietary and foraging functional traits occupy similar ecological niches? Landsc. Ecol. 2017, 32, 265–278. [Google Scholar] [CrossRef]
- Végvári, Z.; Borza, S.; Juhász, K. The role of phylogeny and life history of migratory waterbirds in designing fishpond management plans. Ecol. Eng. 2015, 85, 288–295. [Google Scholar] [CrossRef]
- Thackeray, S.J.; Henrys, P.A.; Hemming, D.; Bell, J.R.; Botham, M.S.; Burthe, S.; Helaouet, P.; Johns, D.G.; Jones, I.D.; Leech, D.I. Phenological sensitivity to climate across taxa and trophic levels. Nature 2016, 535, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blueweiss, L.; Fox, H.; Kudzma, V.; Nakashima, D.; Peters, R.; Sams, S. Relationships between body size and some life history parameters. Oecologia 1978, 37, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Pianka, E.R. On r-and K-selection. Am. Nat. 1970, 104, 592–597. [Google Scholar] [CrossRef]
- Sæther, B.-E.; Lande, R.; Engen, S.; Weimerskirch, H.; Lillegård, M.; Altwegg, R.; Becker, P.H.; Bregnballe, T.; Brommer, J.E.; McCleery, R.H. Generation time and temporal scaling of bird population dynamics. Nature 2005, 436, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Dingle, H. Animal migration: Is there a common migratory syndrome? J. Ornithol. 2006, 147, 212–220. [Google Scholar] [CrossRef]
- Okes, N.C.; Hockey, P.A.R.; Cumming, G.S. Habitat use and life history as predictors of bird responses to habitat change. Conserv. Biol. 2008, 22, 151–162. [Google Scholar] [CrossRef]
- Dodman, T.; Diagana, C.H. Conservation dilemmas for intra-African migratory waterbirds. Waterbirds World 2006, 218, 223–230. [Google Scholar]
- Brochet, A.L.; Dessborn, L.; Legagneux, P.; Elmberg, J.; Gauthier-Clerc, M.; Fritz, H.; Guillemain, M. Is diet segregation between dabbling ducks due to food partitioning? A review of seasonal patterns in the Western Palearctic. J. Zool. 2012, 286, 171–178. [Google Scholar]
- Julliard, R.; Clavel, J.; Devictor, V.; Jiguet, F.; Couvet, D. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 2006, 9, 1237–1244. [Google Scholar] [CrossRef]
- Tripet, F.; Christe, P.; Møller, A.P. The importance of host spatial distribution for parasite specialization and speciation: A comparative study of bird fleas (Siphonaptera: Ceratophyllidae). J. Anim. Ecol. 2002, 71, 735–748. [Google Scholar] [CrossRef]
- Dall, S.R.; Cuthill, I.C. The information costs of generalism. Oikos 1997, 197–202. [Google Scholar] [CrossRef]
- Zanchetta, C.V.; Moore, D.J.; Weseloh, D.C.; Quinn, J.S. Population trends of colonial waterbirds nesting in Hamilton Harbour in relation to changes in habitat and management. Aquat. Ecosyst. Health Manag. 2016, 19, 192–205. [Google Scholar] [CrossRef]
- Hansen, B.D.; Menkhorst, P.; Moloney, P.; Loyn, R.H. Long-term declines in multiple waterbird species in a tidal embayment, south-east Australia. Austral Ecol. 2015, 40, 515–527. [Google Scholar] [CrossRef]
- Cheriak, L.; Barbraud, C.; Doumandji, S.; Bouguessa, S. Diet variability in the White Stork Ciconia ciconia in eastern Algeria. Ostrich 2014, 85, 201–204. [Google Scholar] [CrossRef]
- Tarakini, T.; Guerbois, S.; Wencelius, J.; Mundy, P.; Fritz, H. Integrating local ecological knowledge for waterbird conservation: Insights from Kavango-Zambezi transfrontier conservation area, Zimbabwe. Trop. Conserv. Sci. 2018, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Halse, S. Diet, body condition, and gut size of Egyptian geese. J. Wildl. Manag. 1984, 48, 569–573. [Google Scholar] [CrossRef]
- Stewart, K.; Matthiesen, D.; Leblanc, L.; West, J. Prey diversity and selectivity of the African fish eagle: Data from a roost in northern Kenya. Afr. J. Ecol. 1997, 35, 133–145. [Google Scholar] [CrossRef]
- Arzel, C.; Elmberg, J.; Guillemain, M.; Legagneux, P.; Bosca, F.; Chambouleyron, M.; Lepley, M.; Pin, C.; Arnaud, A.; Schricke, V. Average mass of seeds encountered by foraging dabbling ducks in Western Europe. Wildl. Biol. 2007, 13, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Liordos, V. Foraging guilds of waterbirds wintering in a Mediterranean coastal wetland. Zool. Stud. 2010, 49, 311–323. [Google Scholar]
- Tibshirani, R.; Walther, G.; Hastie, T. Estimating the number of clusters in a data set via the gap statistic. J. R. Stat. Soc. Ser. B Stat. Methodol. 2001, 63, 411–423. [Google Scholar] [CrossRef]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A.; Charrad, M.M. Package ‘NbClust’. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar]
- Mwedzi, T.; Zimunya, T.G.; Bere, T.; Tarakini, T.; Mangadze, T. Disentangling and ranking the influence of multiple stressors on macroinvertebrate communities in a tropical river system. Int. Rev. Hydrobiol. 2017, 102, 1–11. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing (Internet); R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bere, T.; Dalu, T.; Mwedzi, T. Detecting the impact of heavy metal contaminated sediment on benthic macroinvertebrate communities in tropical streams. Sci. Total Environ. 2016, 572, 147–156. [Google Scholar] [CrossRef]
- Guareschi, S.; Abellán, P.; Laini, A.; Green, A.; Sánchez-Zapata, J.; Velasco, J.; Millán, A. Cross-taxon congruence in wetlands: Assessing the value of waterbirds as surrogates of macroinvertebrate biodiversity in Mediterranean Ramsar sites. Ecol. Indic. 2015, 49, 204–215. [Google Scholar] [CrossRef]
- Emmerson, M.C.; Raffaelli, D. Predator–prey body size, interaction strength and the stability of a real food web. J. Anim. Ecol. 2004, 73, 399–409. [Google Scholar] [CrossRef]
- Austin, V.I.; Ribot, R.F.H.; Bennett, A.T.D. If waterbirds are nocturnal are we conserving the right habitats? Emu 2016, 116, 423–427. [Google Scholar] [CrossRef]
- Griffiths, M. Identifying Gamebird Hunting ‘Hotspots’: Implications for Conservation. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 1998. [Google Scholar]
- Ndlovu, M.; Cumming, G.S.; Hockey, P.A.; Nkosi, M.D.; Mutumi, G.L. A study of moult-site fidelity in Egyptian geese, Alopochen aegyptiaca, in South Africa. Afr. Zool. 2013, 48, 240–249. [Google Scholar] [CrossRef]
- Ramachandran, R.; Kumar, A.; Gopi Sundar, K.S.; Bhalla, R.S. Hunting or habitat? Drivers of waterbird abundance and community structure in agricultural wetlands of southern India. Ambio 2017, 46, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, R.J.; Wood, K.A.; Redpath, S.M.; Young, J.C. Conservation conflicts: Future research challenges. Curr. Trends Wildl. Res. 2016, 1, 267–282. [Google Scholar]
- Caron, A.; Abolnik, C.; Mundava, J.; Gaidet, N.; Burger, C.E.; Mochotlhoane, B.; Bruinzeel, L.; Chiweshe, N.; Garine-Wichatitsky, M.D.; Cumming, G.S. Persistence of low pathogenic avian influenza virus in waterfowl in a Southern African ecosystem. EcoHealth 2011, 8, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Haq, R.U.; Eiam-Ampai, K.; Ngoprasert, D.; Sasaki, N.; Shrestha, R.P. Changing landscapes and declining populations of resident waterbirds: A 12-year study in Bung Boraphet wetland, Thailand. Trop. Conserv. Sci. 2018, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Holmgaard, S.B.; Eythórsson, E.; Tombre, I.M. Hunter opinions on the management of migratory geese: A case of stakeholder involvement in adaptive harvest management. Hum. Dimens. Wildl. 2018, 23, 1–9. [Google Scholar] [CrossRef]
- Smith, R.V.; Stafford, J.D.; Yetter, A.P.; Horath, M.M.; Hine, C.S.; Hoover, J.P. Foraging ecology of fall-migrating shorebirds in the Illinois river valley. PLoS ONE 2012, 7, e45121. [Google Scholar] [CrossRef]
- Buelow, C.; Sheaves, M. A birds-eye view of biological connectivity in mangrove systems. Estuar. Coast. Shelf Sci. 2015, 152, 33–43. [Google Scholar] [CrossRef]
- Boere, G.C.; Galbraith, C.A.; Stroud, D.A. Waterbirds Around the World: A Global Overview of the Conservation, Management and Research of the World’s Waterbird Flyways; The Stationery Office: Edinburgh, UK, 2006; p. 960. [Google Scholar]
- Gerwing, T.G.; Kim, J.; Hamilton, D.J.; Barbeau, M.A.; Addison, J.A. Diet reconstruction using next-generation sequencing increases the known ecosystem usage by a shorebird. Auk 2016, 133, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Houlahan, J.E.; Findlay, C.S.; Schmidt, B.R.; Meyer, A.H.; Kuzmin, S.L. Quantitative evidence for global amphibian population declines. Nature 2000, 404, 752–755. [Google Scholar] [CrossRef]
- Lydeard, C.; Cowie, R.H.; Ponder, W.F.; Bogan, A.E.; Bouchet, P.; Clark, S.A.; Cummings, K.S.; Frest, T.J.; Gargominy, O.; Herbert, D.G. The global decline of nonmarine mollusks. BioScience 2004, 54, 321–330. [Google Scholar] [CrossRef]
- Ntiamoa-Baidu, Y.A.A.; Piersma, T.; Wiersma, P.; Poot, M.; Battley, P.; Gordon, C. Water depth selection, daily feeding routines and diets of waterbirds in coastal lagoons in Ghana. Ibis 1998, 140, 89–103. [Google Scholar] [CrossRef] [Green Version]


| Diet Body Length Classes | Waterbird Allocated Guilds | |||||||
|---|---|---|---|---|---|---|---|---|
| Global Status | 1 | 2 | 3 | Herb | Insect | Semi-Omniv | Omniv | Pisci |
| Declining | 55 | 20 | 24 | 8 | 31 | 8 | 24 | 28 |
| Not declining | 23 | 32 | 9 | 2 | 21 | 10 | 10 | 21 |
| Total | 78 | 52 | 33 | 10 | 52 | 18 | 34 | 49 |
| Percentage of Birds Consuming Prey Item | ||||
|---|---|---|---|---|
| Prey Item | Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 |
| Algae/plankton | 7 | 2 | - | 10 |
| Annelids | 49 | 38 | - | 20 |
| Arachnids | 25 | 6 | - | - |
| Bird eggs | 6 | 10 | - | - |
| Birds | 6 | 26 | 30 | - |
| Chilopods | 1 | - | - | - |
| Chironomids | 1 | 2 | - | 5 |
| Clinids | 1 | - | - | - |
| Coleopterans | 39 | 32 | - | 20 |
| Crustaceans | 59 | 62 | 50 | 55 |
| Culicidae | 5 | - | - | - |
| Cyanobacteria | 5 | - | 10 | - |
| Demapterans | 17 | 6 | - | 15 |
| Diatoms | 2 | - | 10 | 5 |
| Diplopods | 6 | 4 | - | 5 |
| Dipterans | 47 | 22 | 10 | 20 |
| Echinoderms | 4 | - | - | - |
| Ephemeropterans | 30 | 8 | 10 | - |
| Fruits | 7 | 6 | 20 | 90 |
| Gastropods | 20 | 12 | 20 | 05 |
| Hemipterans | 37 | 2 | - | 30 |
| Homopterans | 25 | 12 | 10 | 5 |
| Hymenoptera | 35 | 16 | - | - |
| Isopterans | 34 | 12 | - | 30 |
| Leaves | 23 | 10 | 20 | 90 |
| Lepidopterans | 35 | 16 | 20 | 15 |
| Mantids | 17 | 8 | - | 10 |
| Molluscs | 46 | 54 | 50 | 40 |
| Odonata | 25 | 18 | - | 5 |
| Orthopterans | 30 | 28 | 20 | 40 |
| Roots/tubers | 22 | 12 | 10 | 85 |
| Rotifers | 5 | 2 | 10 | - |
| Scorpiones | 8 | 8 | 10 | 20 |
| Seeds | 39 | 18 | 30 | 80 |
| Small amphibians | 30 | 42 | 60 | 20 |
| Small mammals | 7 | 24 | 20 | - |
| Small reptiles | 11 | 20 | 50 | - |
| Trichopterans | 17 | 12 | 10 | 20 |
| Tunicates | 54 | 66 | 50 | 30 |
| Variable | Estimate | Std. Error | Z Value | p Value |
|---|---|---|---|---|
| Intercept | −0.8718 | 0.2483 | −3.511 | 0.0004 |
| Diet body length 11–50 mm | 1.3418 | 0.378 | 3.55 | 0.0004 |
| Diet body length > 50 mm | −0.109 | 0.4631 | −0.235 | 0.8139 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarakini, T.; Mabika, I.; Mwedzi, T.; Mundy, P.; Fritz, H. Diet and Life-History Traits of Savannah Dwelling Waterbirds in Southern Africa: Implications for Their Conservation Status. Birds 2021, 2, 173-184. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020013
Tarakini T, Mabika I, Mwedzi T, Mundy P, Fritz H. Diet and Life-History Traits of Savannah Dwelling Waterbirds in Southern Africa: Implications for Their Conservation Status. Birds. 2021; 2(2):173-184. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020013
Chicago/Turabian StyleTarakini, Tawanda, Innocent Mabika, Tongayi Mwedzi, Peter Mundy, and Hervé Fritz. 2021. "Diet and Life-History Traits of Savannah Dwelling Waterbirds in Southern Africa: Implications for Their Conservation Status" Birds 2, no. 2: 173-184. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020013