
Searching for G-Quadruplex-Binding Proteins in Plants: New Insight into Possible G-Quadruplex Regulation


 ,
,  , and
, and 
Abstract
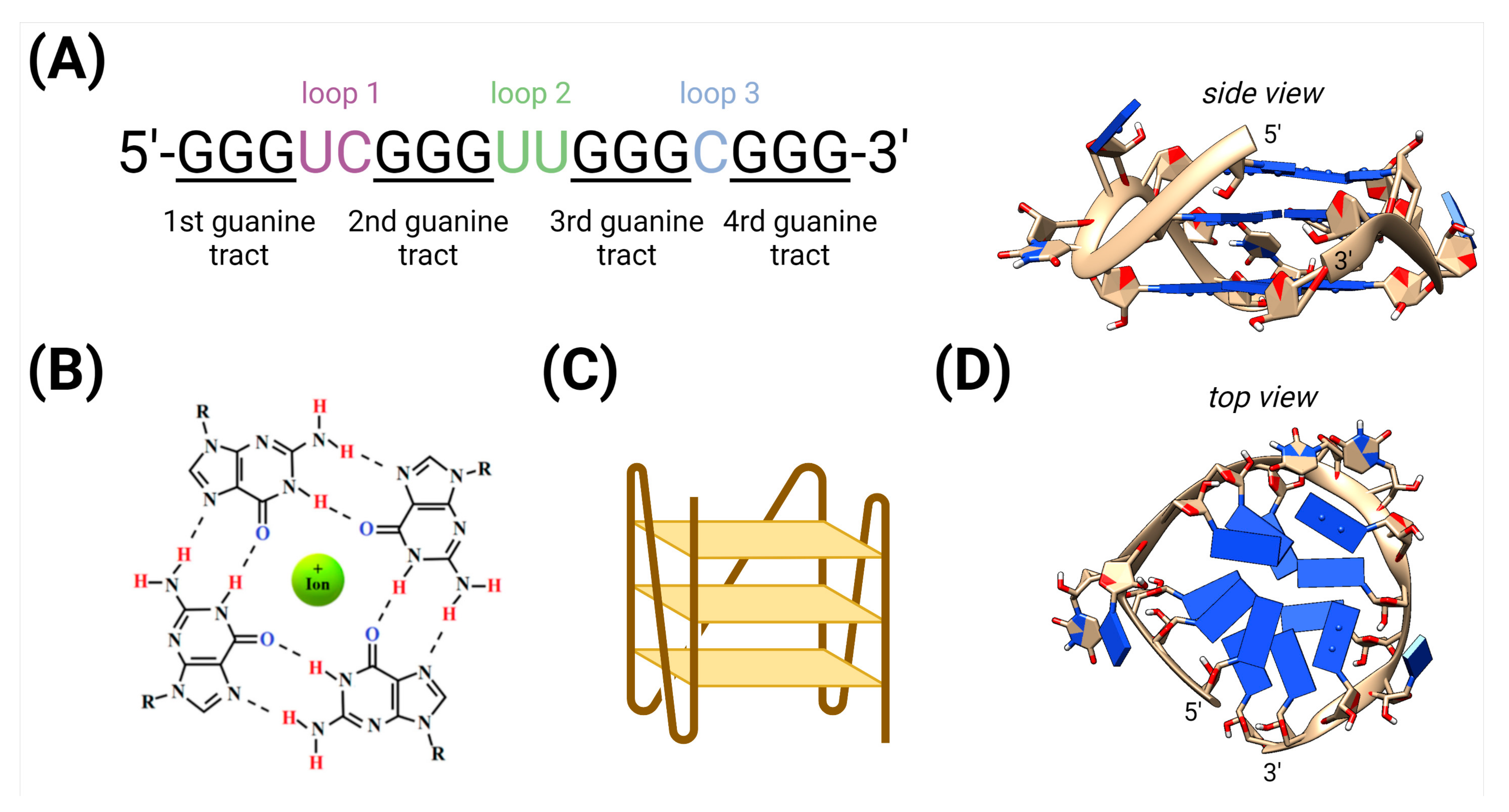
:1. Introduction
2. Materials and Methods
2.1. Identification of G4-Binding Proteins Using the FIMO Approach
2.2. Identification of G4-Binding Proteins Using BLASTp Approach
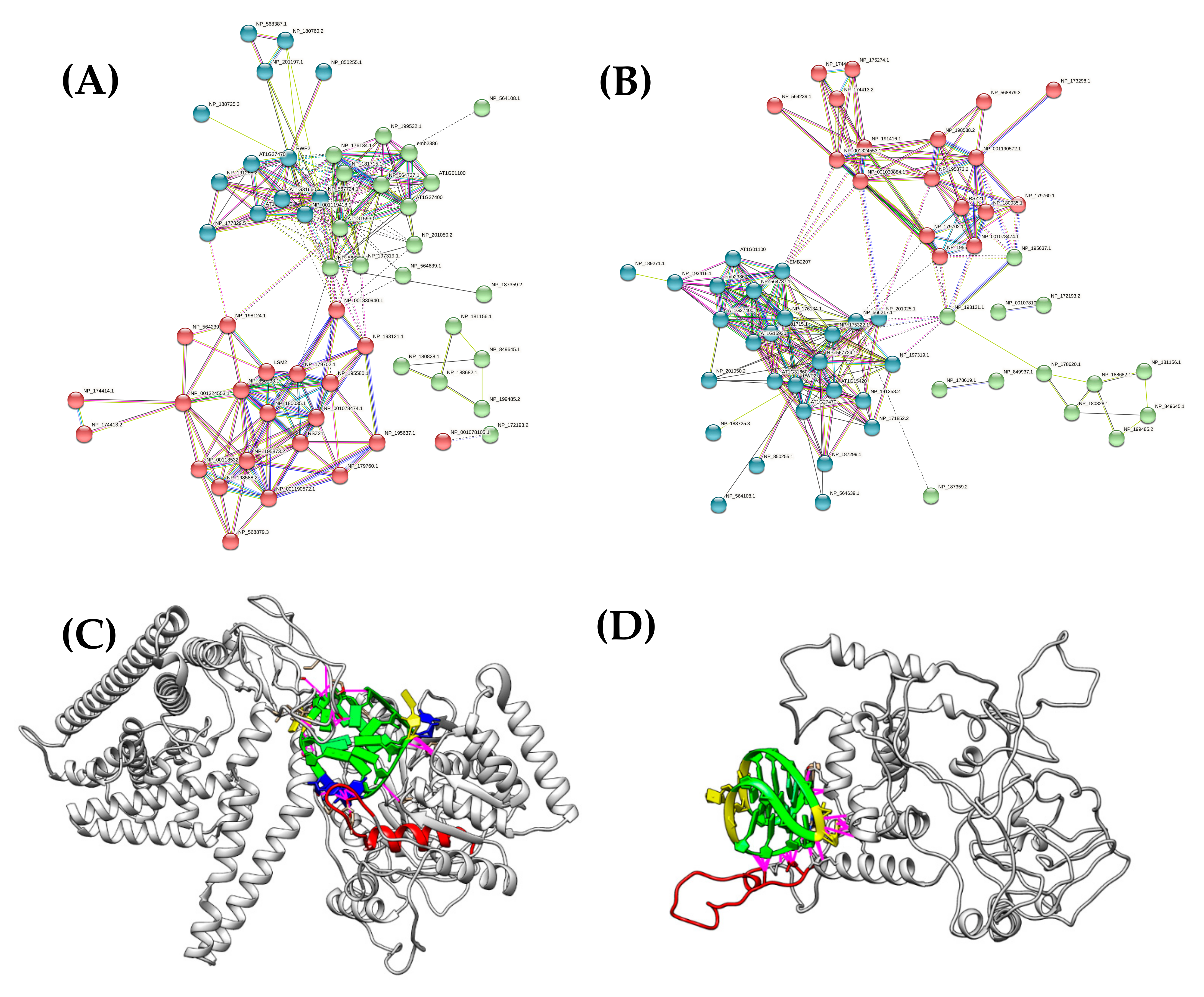
2.3. Functional Interaction Network Analysis Using STRING Approach
2.4. Process of 3D Structural Modeling and Docking
3. Results
3.1. Identification of G4-Binding Proteins in Plants Using a FIMO Approach
3.2. Identification of G4-Binding Proteins in Plants Using a BLASTp Approach
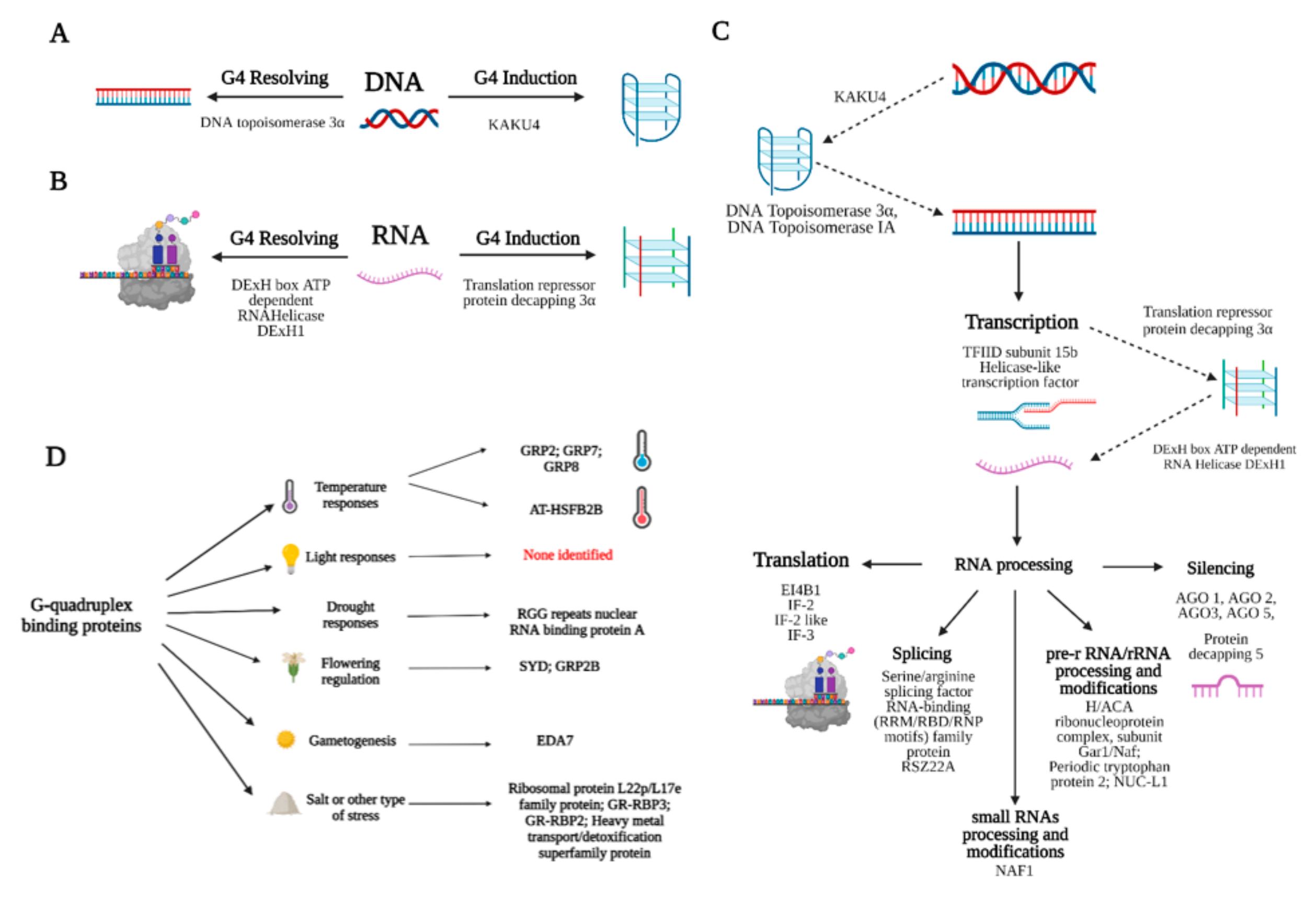
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rhodes, D.; Lipps, H.J. G-Quadruplexes and Their Regulatory Roles in Biology. Nucleic Acids Res. 2015, 43, 8627–8637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Podbevšek, P.; Plavec, J.; Sugimoto, N. Effects of Metal Ions and Cosolutes on G-Quadruplex Topology. J. Inorg. Biochem. 2017, 166, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Bartas, M.; Brázda, V.; Karlický, V.; Červeň, J.; Pečinka, P. Bioinformatics Analyses and In Vitro Evidence for Five and Six Stacked G-Quadruplex Forming Sequences. Biochimie 2018, 150, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Guedin, A.; Gros, J.; Alberti, P.; Mergny, J.-L. How Long Is Too Long? Effects of Loop Size on G-Quadruplex Stability. Nucleic Acids Res. 2010, 38, 7858–7868. [Google Scholar] [CrossRef] [Green Version]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, Topology and Structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [Green Version]
- Patro, L.P.P.; Kumar, A.; Kolimi, N.; Rathinavelan, T. 3D-NuS: A Web Server for Automated Modeling and Visualization of Non-Canonical 3-Dimensional Nucleic Acid Structures. J. Mol. Biol. 2017, 429, 2438–2448. [Google Scholar] [CrossRef]
- Hänsel-Hertsch, R.; Simeone, A.; Shea, A.; Hui, W.W.; Zyner, K.G.; Marsico, G.; Rueda, O.M.; Bruna, A.; Martin, A.; Zhang, X. Landscape of G-Quadruplex DNA Structural Regions in Breast Cancer. Nat. Genet. 2020, 52, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Simone, R.; Fratta, P.; Neidle, S.; Parkinson, G.N.; Isaacs, A.M. G-Quadruplexes: Emerging Roles in Neurodegenerative Diseases and the Non-Coding Transcriptome. FEBS Lett. 2015, 589, 1653–1668. [Google Scholar] [CrossRef]
- Wang, E.; Thombre, R.; Shah, Y.; Latanich, R.; Wang, J. G-Quadruplexes as Pathogenic Drivers in Neurodegenerative Disorders. Nucleic Acids Res. 2021, 49, 4816–4830. [Google Scholar] [CrossRef] [PubMed]
- Lavezzo, E.; Berselli, M.; Frasson, I.; Perrone, R.; Palù, G.; Brazzale, A.R.; Richter, S.N.; Toppo, S. G-Quadruplex Forming Sequences in the Genome of All Known Human Viruses: A Comprehensive Guide. PLoS Comput. Biol. 2018, 14, e1006675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartas, M.; Brázda, V.; Bohálová, N.; Cantara, A.; Volná, A.; Stachurová, T.; Malachová, K.; Jagelská, E.B.; Porubiaková, O.; Červeň, J. In-Depth Bioinformatic Analyses of Nidovirales Including Human SARS-CoV-2, SARS-CoV, MERS-CoV Viruses Suggest Important Roles of Non-Canonical Nucleic Acid Structures in Their Lifecycles. Front. Microbiol. 2020, 11, 1583. [Google Scholar] [CrossRef]
- Brázda, V.; Hároníková, L.; Liao, J.C.; Fojta, M. DNA and RNA Quadruplex-Binding Proteins. Int. J. Mol. Sci. 2014, 15, 17493–17517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.K.; Tawani, A.; Mishra, A.; Kumar, A. G4IPDB: A Database for G-Quadruplex Structure Forming Nucleic Acid Interacting Proteins. Sci. Rep. 2016, 6, 38144. [Google Scholar] [CrossRef] [Green Version]
- Cagirici, H.B.; Budak, H.; Sen, T.Z. Genome-Wide Discovery of G-Quadruplexes in Barley. Sci. Rep. 2021, 11, 7876. [Google Scholar] [CrossRef]
- Cagirici, H.B.; Sen, T.Z. Genome-Wide Discovery of G-Quadruplexes in Wheat: Distribution and Putative Functional Roles. G3 Genes Genomes Genet. 2020, 10, 2021–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefos, G.C.; Theodorou, G.; Politis, I. DNA G-Quadruplexes: Functional Significance in Plant and Farm Animal Science. Anim. Biotechnol. 2021, 32, 262–271. [Google Scholar] [CrossRef]
- Volná, A.; Bartas, M.; Karlický, V.; Nezval, J.; Kundrátová, K.; Pečinka, P.; Špunda, V.; Červeň, J. G-Quadruplex in Gene Encoding Large Subunit of Plant RNA Polymerase II: A Billion-Year-Old Story. Int. J. Mol. Sci. 2021, 22, 7381. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Duncan, S.; Yang, X.; Abdelhamid, M.A.S.; Huang, L.; Zhang, H.; Benfey, P.N.; Waller, Z.A.E.; Ding, Y. G-Quadruplex Structures Trigger RNA Phase Separation. Nucleic Acids Res. 2019, 47, 11746–11754. [Google Scholar] [CrossRef]
- Andorf, C.M.; Kopylov, M.; Dobbs, D.; Koch, K.E.; Stroupe, M.E.; Lawrence, C.J.; Bass, H.W. G-Quadruplex (G4) Motifs in the Maize (Zea mays L.) Genome Are Enriched at Specific Locations in Thousands of Genes Coupled to Energy Status, Hypoxia, Low Sugar, and Nutrient Deprivation. J. Genet. Genom. 2014, 41, 627–647. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Cho, H.S.; Nam, H.; Jo, H.; Yoon, J.; Park, C.; Dang, T.V.T.; Kim, E.; Jeong, J.; Park, S.; et al. Translational Control of Phloem Development by RNA G-Quadruplex–JULGI Determines Plant Sink Strength. Nat. Plants 2018, 4, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Sjakste, T.; Leonova, E.; Petrovs, R.; Trapina, I.; Röder, M.S.; Sjakste, N. Tight DNA-Protein Complexes Isolated from Barley Seedlings Are Rich in Potential Guanine Quadruplex Sequences. PeerJ 2020, 8, e8569. [Google Scholar] [CrossRef] [PubMed]
- Vasilyev, N.; Polonskaia, A.; Darnell, J.C.; Darnell, R.B.; Patel, D.J.; Serganov, A. Crystal Structure Reveals Specific Recognition of a G-Quadruplex RNA by a β-Turn in the RGG Motif of FMRP. Proc. Natl. Acad. Sci. USA 2015, 112, E5391–E5400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brázda, V.; Červeň, J.; Bartas, M.; Mikysková, N.; Coufal, J.; Pečinka, P. The Amino Acid Composition of Quadruplex Binding Proteins Reveals a Shared Motif and Predicts New Potential Quadruplex Interactors. Molecules 2018, 23, 2341. [Google Scholar] [CrossRef] [Green Version]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for Occurrences of a given Motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D. Reference Sequence (RefSeq) Database at NCBI: Current Status, Taxonomic Expansion, and Functional Annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gaetano, C.M.; Williams, K.R.; Bassell, G.J.; Mihailescu, M.R. FMRP Interacts with G-Quadruplex Structures in the 3′-UTR of Its Dendritic Target Shank1 MRNA. RNA Biol. 2014, 11, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P. STRING V10: Protein–Protein Interaction Networks, Integrated over the Tree of Life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-Based Protein Structure Modeling Using the RaptorX Web Server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Leonard, N.; De Bolle, X.; Depiereux, E. ESyPred3D: Prediction of Proteins 3D Structures. Bioinformatics 2002, 18, 1250–1256. [Google Scholar] [CrossRef]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Mullen, M.A.; Olson, K.J.; Dallaire, P.; Major, F.; Assmann, S.M.; Bevilacqua, P.C. RNA G-Quadruplexes in the Model Plant Species Arabidopsis thaliana: Prevalence and Possible Functional Roles. Nucleic Acids Res. 2010, 38, 8149–8163. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Silva, H.M.; Resende, B.C.; Umaki, A.C.S.; Prado, W.; da Silva, M.S.; Virgílio, S.; Macedo, A.M.; Pena, S.D.J.; Tahara, E.B.; Tosi, L.R.O.; et al. DNA Topoisomerase 3α Is Involved in Homologous Recombination Repair and Replication Stress Response in Trypanosoma Cruzi. Front. Cell Dev. Biol. 2021, 9, 849. [Google Scholar] [CrossRef]
- Goto, C.; Tamura, K.; Fukao, Y.; Shimada, T.; Hara-Nishimura, I. The Novel Nuclear Envelope Protein KAKU4 Modulates Nuclear Morphology in Arabidopsis. Plant Cell 2014, 26, 2143–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.C.; Caceres, J.F. The SR Protein Family of Splicing Factors: Master Regulators of Gene Expression. Biochem. J. 2009, 417, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Liu, Y.; Kim, M.-H.; Imai, R. An RNA Chaperone, AtCSP2, Negatively Regulates Salt Stress Tolerance. Plant Signal. Behav. 2015, 10, e1042637. [Google Scholar] [CrossRef] [PubMed]
- Köster, T.; Meyer, K.; Weinholdt, C.; Smith, L.M.; Lummer, M.; Speth, C.; Grosse, I.; Weigel, D.; Staiger, D. Regulation of Pri-MiRNA Processing by the HnRNP-like Protein AtGRP7 in Arabidopsis. Nucleic Acids Res. 2014, 42, 9925–9936. [Google Scholar] [CrossRef]
- Cao, J.-Y.; Xu, Y.-P.; Cai, X.-Z. Integrated MiRNAome and Transcriptome Analysis Reveals Argonaute 2-Mediated Defense Responses Against the Devastating Phytopathogen Sclerotinia Sclerotiorum. Front. Plant Sci. 2020, 11, 500. [Google Scholar] [CrossRef]
- Pontvianne, F.; Matía, I.; Douet, J.; Tourmente, S.; Medina, F.J.; Echeverria, M.; Sáez-Vásquez, J. Characterization of AtNUC-L1 Reveals a Central Role of Nucleolin in Nucleolus Organization and Silencing of AtNUC-L2 Gene in Arabidopsis. Mol. Biol. Cell 2007, 18, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Lange, H.; Zuber, H.; Sement, F.M.; Chicher, J.; Kuhn, L.; Hammann, P.; Brunaud, V.; Bérard, C.; Bouteiller, N.; Balzergue, S.; et al. The RNA Helicases AtMTR4 and HEN2 Target Specific Subsets of Nuclear Transcripts for Degradation by the Nuclear Exosome in Arabidopsis thaliana. PLoS Genet. 2014, 10, e1004564. [Google Scholar] [CrossRef] [Green Version]
- Western, T.L.; Cheng, Y.; Liu, J.; Chen, X. HUA ENHANCER2, a Putative DExH-Box RNA Helicase, Maintains Homeotic B and C Gene Expression in Arabidopsis. Development 2002, 129, 1569–1581. [Google Scholar] [CrossRef]
- Larochelle, M.; Lemay, J.-F.; Bachand, F. The THO Complex Cooperates with the Nuclear RNA Surveillance Machinery to Control Small Nucleolar RNA Expression. Nucleic Acids Res. 2012, 40, 10240–10253. [Google Scholar] [CrossRef] [Green Version]
- Shukla, K.; Thakur, R.S.; Ganguli, D.; Rao, D.N.; Nagaraju, G. Escherichia coli and Neisseria gonorrhoeae UvrD Helicase Unwinds G4 DNA Structures. Biochem. J. 2017, 474, 3579–3597. [Google Scholar] [CrossRef]
- Shubina, M.Y.; Musinova, Y.R.; Sheval, E.V. Nucleolar Methyltransferase Fibrillarin: Evolution of Structure and Functions. Biochem. Mosc. 2016, 81, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Fleurdépine, S.; Deragon, J.-M.; Devic, M.; Guilleminot, J.; Bousquet-Antonelli, C. A Bona Fide La Protein Is Required for Embryogenesis in Arabidopsis thaliana. Nucleic Acids Res. 2007, 35, 3306–3321. [Google Scholar] [CrossRef] [PubMed]
- Goyal, M.; Banerjee, C.; Nag, S.; Bandyopadhyay, U. The Alba Protein Family: Structure and Function. Biochim. Biophys. Acta (BBA)—Prot. Proteom. 2016, 1864, 570–583. [Google Scholar] [CrossRef]
- Li, X.; Gao, X.; Wei, Y.; Deng, L.; Ouyang, Y.; Chen, G.; Li, X.; Zhang, Q.; Wu, C. Rice APOPTOSIS INHIBITOR5 Coupled with Two DEAD-Box Adenosine 5′-Triphosphate-Dependent RNA Helicases Regulates Tapetum Degeneration. Plant Cell 2011, 23, 1416–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Ruiz, M.T.G.; McGinn, M.G.; Lowery, N.; et al. Roles and Programming of Arabidopsis ARGONAUTE Proteins during Turnip Mosaic Virus Infection. PLoS Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef]
- Kim, J.S.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Goh, C.-H.; Woo, Y.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-Rich RNA-Binding Protein7 Affects Abiotic Stress Responses by Regulating Stomata Opening and Closing in Arabidopsis thaliana. Plant J. 2008, 55, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.J.; Jang, B.; Jung, C.-H.; Ahn, S.J.; Goh, C.-H.; Cho, K.; Han, O.; Kang, H. Functional Characterization of a Glycine-Rich RNA-Binding Protein 2 in Arabidopsis thaliana under Abiotic Stress Conditions. Plant J. 2007, 50, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Ciuzan, O.; Ladomery, M.; Wilson, I.; Hancock, J.; Pamfil, D. The Arabidopsis thaliana Glycine-Rich RNA Binding Proteins AtGRP7 and AtGRP2 Are Involved in Early Development. ProEnviron. Promediu 2013, 6, 72–76. [Google Scholar]
- Chen, M.C.; Murat, P.; Abecassis, K.; Ferré-D’Amaré, A.R.; Balasubramanian, S. Insights into the Mechanism of a G-Quadruplex-Unwinding DEAH-Box Helicase. Nucleic Acids Res. 2015, 43, 2223–2231. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.C.; Tippana, R.; Demeshkina, N.A.; Murat, P.; Balasubramanian, S.; Myong, S.; Ferré-D’Amaré, A.R. Structural Basis of G-Quadruplex Unfolding by the DEAH/RHA Helicase DHX36. Nature 2018, 558, 465–469. [Google Scholar] [CrossRef]
- Tosoni, E.; Frasson, I.; Scalabrin, M.; Perrone, R.; Butovskaya, E.; Nadai, M.; Palù, G.; Fabris, D.; Richter, S.N. Nucleolin Stabilizes G-Quadruplex Structures Folded by the LTR Promoter and Silences HIV-1 Viral Transcription. Nucleic Acids Res. 2015, 43, 8884–8897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Antonio, M.; Ponjavic, A.; Radzevičius, A.; Ranasinghe, R.T.; Catalano, M.; Zhang, X.; Shen, J.; Needham, L.-M.; Lee, S.F.; Klenerman, D.; et al. Single-Molecule Visualization of DNA G-Quadruplex Formation in Live Cells. Nat. Chem. 2020, 12, 832–837. [Google Scholar] [CrossRef]
- Ravichandran, S.; Ahn, J.-H.; Kim, K.K. Unraveling the Regulatory G-Quadruplex Puzzle: Lessons from Genome and Transcriptome-Wide Studies. Front. Genet. 2019, 10, 1002. [Google Scholar] [CrossRef]
- Serikawa, T.; Spanos, C.; von Hacht, A.; Budisa, N.; Rappsilber, J.; Kurreck, J. Comprehensive Identification of Proteins Binding to RNA G-Quadruplex Motifs in the 5′ UTR of Tumor-Associated MRNAs. Biochimie 2018, 144, 169–184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Protein Full Name | AN | Length | E-Value | Query Cover |
|---|---|---|---|---|---|
| NIQI | Hypothetical protein AXX17 AT1G21190 | OAP13840.1 | 265 | 4 × 10−6 | 100 |
| Unnamed protein product | AAN60302.1 | 272 | 4 × 10−6 | 100 | |
| Unnamed protein product | CAA0224497.1 | 312 | 4 × 10−6 | 100 | |
| Alba DNA/RNA binding protein | NP_564108.1 | 315 | 4 × 10−6 | 100 | |
| Unnamed protein product | VYS46658.1 | 317 | 4 × 10−6 | 100 | |
| Contains similarity to pigpen protein from M. musculus | AAF79893.1 | 538 | 4 × 10−6 | 100 | |
| Unnamed protein product | CAD5313245.1 | 787 | 4 × 10−6 | 100 | |
| F1K23.1 | AAF24538.2 | 347 | 1 × 10−5 | 100 | |
| Hypothetical protein | AAF88129.1 | 533 | 1 × 10−5 | 100 | |
| Apoptosis inhibitory protein 5 | NP_001322605.1 | 534 | 1 × 10−5 | 100 | |
| RGG | Argonaute 3 | NP_001322307.1 | 1158 | 2 × 10−6 | 84 |
| Unnamed protein product | VYS47693.1 | 1205 | 5 × 10−6 | 84 | |
| Argonaute 3 | NP_174414.1 | 1194 | 1 × 10−5 | 84 | |
| Unnamed protein product | CAA0259614.1 | 1194 | 1 × 10−5 | 84 | |
| Argonaute 3 | NP_001322306.1 | 1120 | 3 × 10−5 | 84 | |
| AGO3 | OAP18010.1 | 1141 | 4 × 10−5 | 84 | |
| Glycine-rich protein | NP_568523.1 | 118 | 4 × 10−4 | 84 | |
| Unnamed protein product | VYS68274.1 | 118 | 4 × 10−4 | 84 | |
| Hypothetical protein AXX17 AT5G32160 | OAO90547.1 | 123 | 4 × 10−4 | 84 | |
| Hyaluronan/mRNA binding family | NP_001078399.1 | 265 | 9 × 10−4 | 80 |
| Motif | Protein Name | Organism | Simple Name | AN | Length | E-Value | Query Cover |
|---|---|---|---|---|---|---|---|
| NIQI | Uncharacterized protein | Cynara cardunculus | Globe artichoke | XP_024965254.1 | 265 | 2 × 10−7 | 100 |
| Uncharacterized protein | Asparagus officinalis | Garden asparagus | ONK65585.1 | 310 | 2 × 10−7 | 100 | |
| Keratin, type II cytoskeletal 1-like | Asparagus officinalis | Garden asparagus | XP_020272794.1 | 301 | 2 × 10−7 | 100 | |
| Hypothetical protein | Chlamydomonas incerta | - | KAG2446147.1 | 2307 | 5 × 10−7 | 100 | |
| THO complex subunit 4a | Ananas comosus | Pineapple | OAY83511.1 | 271 | 5 × 10−7 | 100 | |
| Translation initiation factor IF-2-like | Aegilops tauschii | Tausch’s goatgrass | XP_020191912.1 | 278 | 2 × 10−6 | 100 | |
| Unnamed protein product | Lactuca saligna | Willowleaf lettuce | CAB4097197.1 | 728 | 3 × 10−6 | 100 | |
| Hypothetical protein | Chlamydomonas schloesseri | - | KAG2434571.1 | 2954 | 4 × 10−6 | 100 | |
| UVRD/REP type putative DNA helicase | Klebsormidium nitens | - | GAQ80746.1 | 1395 | 4 × 10−6 | 100 | |
| Hypothetical protein | Edaphochlamys debaryana | - | KAG2484473.1 | 1151 | 4 × 10−6 | 100 | |
| RGG | Hypothetical protein | Pycnococcus provasolii | - | GHP06452.1 | 396 | 4 × 10−8 | 88 |
| Unnamed protein product | Triticum aestivum | Common wheat | CDM81338.1 | 300 | 6 × 10−6 | 84 | |
| Unnamed protein product | Microthlaspi erraticum | - | CAA7061275.1 | 1044 | 9 × 10−6 | 84 | |
| Fibrillarin 2 | Nicotiana benthamiana | Benth | CAR92137.1 | 314 | 2 × 10−5 | 96 | |
| Mediator of RNA polymerase II transcription subunit 36a-like | Nicotiana tabacum | Cultivated tobacco | NP_001311935.1 | 314 | 2 × 10−5 | 96 | |
| Predicted mediator of RNA polymerase II transcription subunit 36a-like | Nicotiana attenuata | Coyote tobacco | XP_019243309.1 | 314 | 2 × 10−5 | 96 | |
| Mediator of RNA polymerase II transcription subunit 36a-like | Nicotiana tomentosiformis | - | XP_009625367.1 | 314 | 2 × 10−5 | 96 | |
| Fibrillarin 2 | Nicotiana benthamiana | Benth | CAK32531.1 | 314 | 2 × 10−5 | 96 | |
| HEN2 | Scenedesmus sp. | - | KAF8068215.1 | 1082 | 2 × 10−5 | 84 | |
| Helicase-like transcription factor | Chlorella sorokiniana | - | PRW34024.1 | 1709 | 3 × 10−5 | 88 |
| Motif | Protein Name | Organism | AN | Length | E-Value | Query Cover |
|---|---|---|---|---|---|---|
| NIQI | Hypothetical protein | Chlamydomonas incerta | KAG2446147.1 | 2307 | 2 × 10−8 | 100 |
| Hypothetical protein | Chlamydomonas schloesseri | KAG2434571.1 | 2954 | 2 × 10−7 | 100 | |
| Hypothetical protein | Edaphochlamys debaryana | KAG2484473.1 | 1151 | 2 × 10−7 | 100 | |
| NER endonuclease | Chlamydomonas reinhardtii | ALL55223.1 | 2463 | 3 × 10−7 | 100 | |
| Hypothetical protein | Chlamydomonas schloesseri | KAG2440529.1 | 2398 | 6 × 10−7 | 100 | |
| XPO1 | Scenedesmus sp. | KAF8057112.1 | 1615 | 9 × 10−7 | 100 | |
| DNA helicase | Raphidocelis subcapitata | GBF99140.1 | 787 | 9 × 10−7 | 100 | |
| Hypothetical protein | Edaphochlamys debaryana | KAG2500053.1 | 1082 | 1 × 10−6 | 100 | |
| Hypothetical protein | Edaphochlamys debaryana | KAG2486618.1 | 1053 | 1 × 10−6 | 100 | |
| Hypothetical protein | Chlamydomonas incerta | KAG2433306.1 | 1398 | 2 × 10−6 | 100 | |
| RGG | Hypothetical protein | Pycnococcus provasolii | GHP06452.1 | 396 | 2 × 10−9 | 88 |
| HEN2 | Scenedesmus sp. | KAF8068215.1 | 1082 | 8 × 10−7 | 84 | |
| Helicase-like transcription factor | Chlorella sorokiniana | PRW34024.1 | 1709 | 1 × 10−6 | 88 | |
| Hypothetical protein | Chlamydomonas incerta | KAG2443577.1 | 671 | 3 × 10−6 | 84 | |
| Hypothetical protein | Chlamydomonas schloesseri | KAG2451206.1 | 782 | 4 × 10−6 | 88 | |
| La1 | Chlorella sorokiniana | PRW45723.1 | 398 | 1 × 10−5 | 92 | |
| DEAD-box ATP-dependent RNA helicase 10 | Chlorella sorokiniana | PRW56671.1 | 462 | 2 × 10−5 | 88 | |
| Hypothetical protein | Edaphochlamys debaryana | KAG2497970.1 | 830 | 2 × 10−5 | 96 | |
| FAD dependent oxidoreductase | Raphidocelis subcapitata | GBF89971.1 | 859 | 2 × 10−5 | 84 | |
| Translation initiation factor 3 | Ostreococcus tauri | XP_003078378.1 | 266 | 3 × 10−5 | 80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volná, A.; Bartas, M.; Nezval, J.; Špunda, V.; Pečinka, P.; Červeň, J. Searching for G-Quadruplex-Binding Proteins in Plants: New Insight into Possible G-Quadruplex Regulation. BioTech 2021, 10, 20. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech10040020
Volná A, Bartas M, Nezval J, Špunda V, Pečinka P, Červeň J. Searching for G-Quadruplex-Binding Proteins in Plants: New Insight into Possible G-Quadruplex Regulation. BioTech. 2021; 10(4):20. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech10040020
Chicago/Turabian StyleVolná, Adriana, Martin Bartas, Jakub Nezval, Vladimír Špunda, Petr Pečinka, and Jiří Červeň. 2021. "Searching for G-Quadruplex-Binding Proteins in Plants: New Insight into Possible G-Quadruplex Regulation" BioTech 10, no. 4: 20. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech10040020