Doxorubicin Enhances Procoagulant Activity of Endothelial Cells after Exposure to Tumour Microparticles on Microfluidic Devices


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
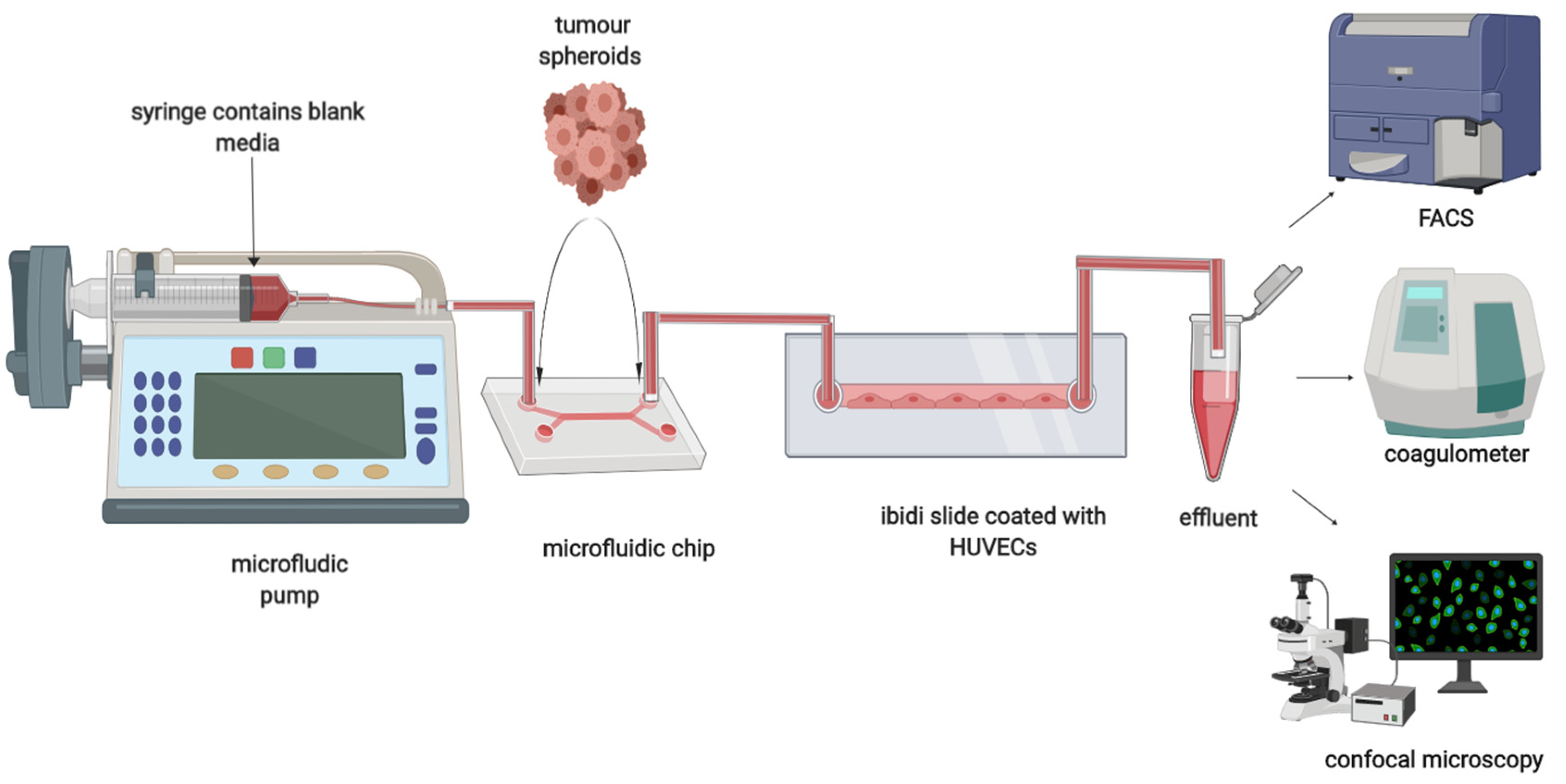
2.2. Dual Chip Microfluidic Setup
2.3. Procoagulant Activity
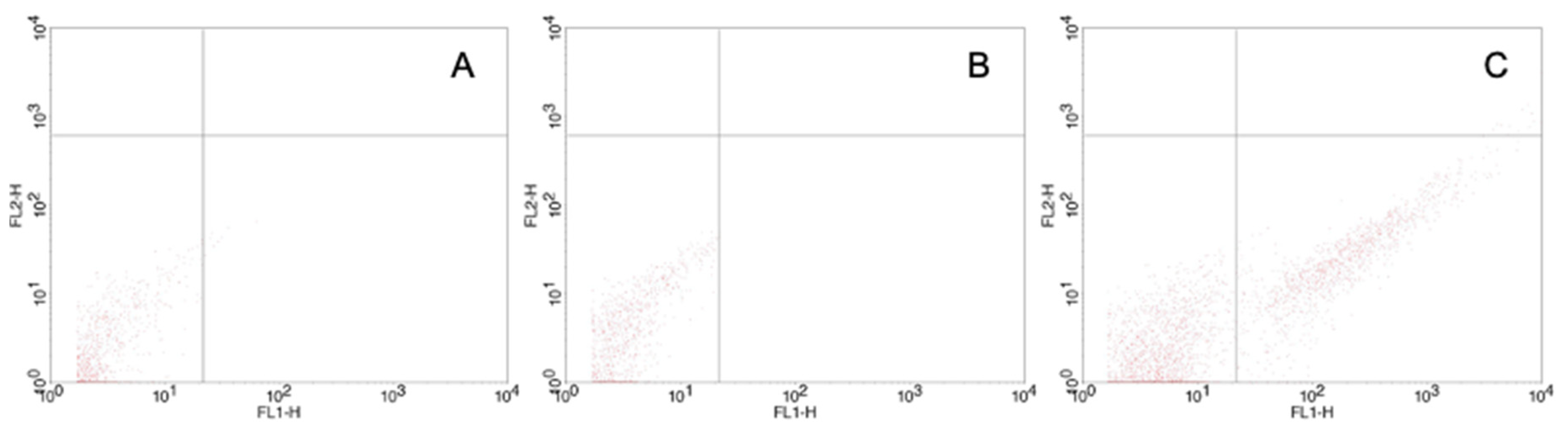
2.4. Flow Cytometry
2.5. Doxorubicin Treatment
2.6. Statistical Analysis
3. Results
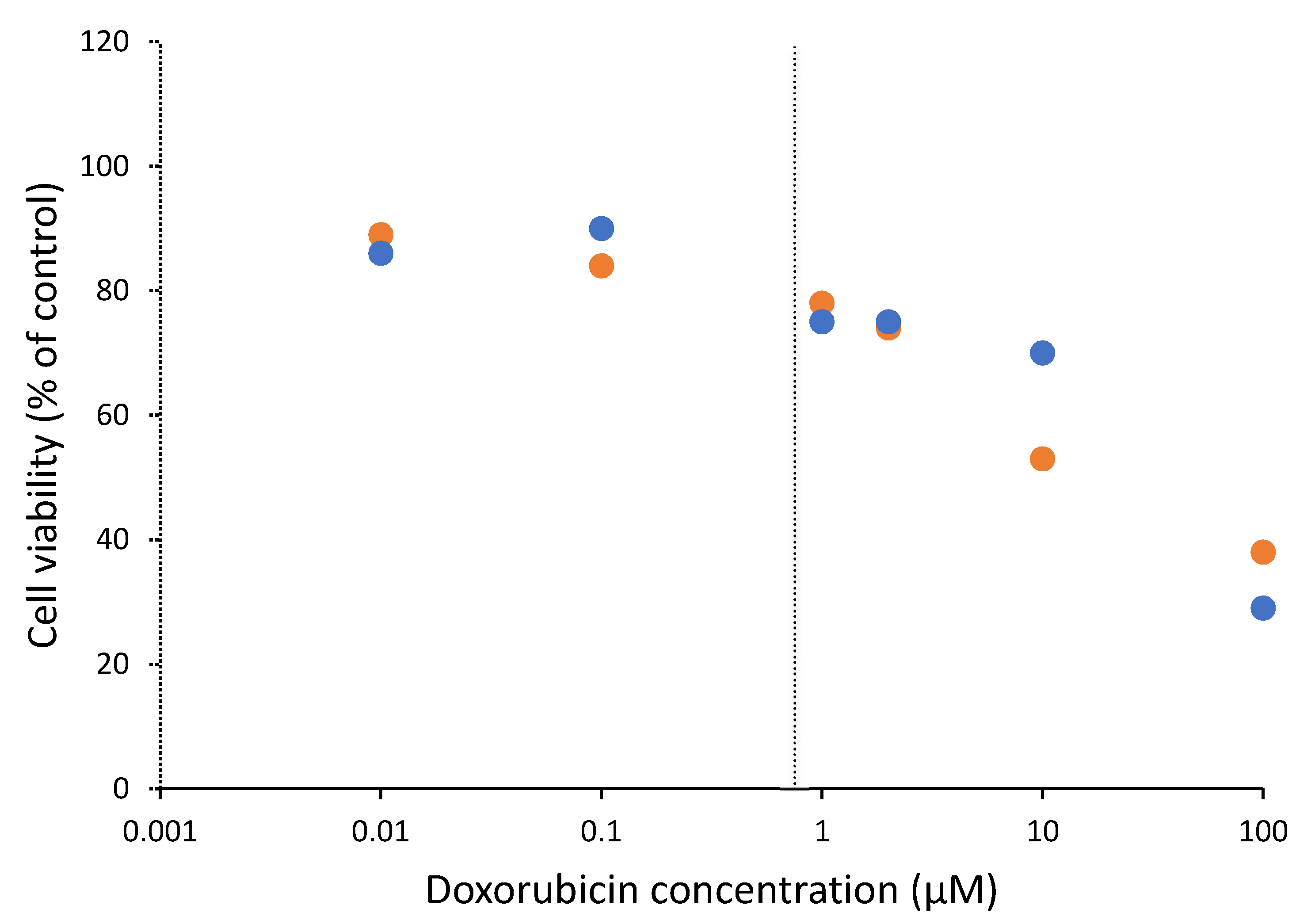
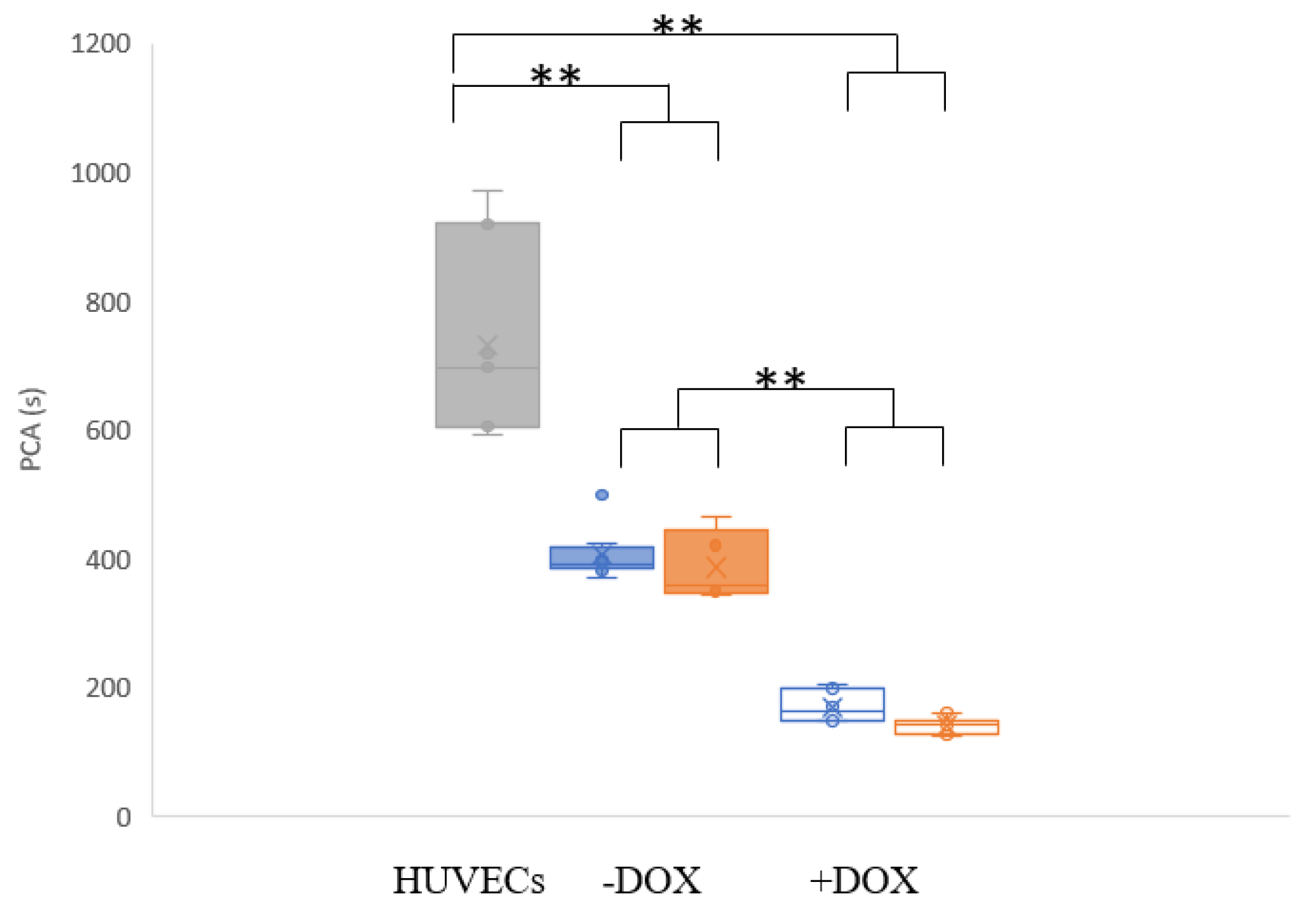
3.1. Cytotoxicity of Doxorubicin on Cells
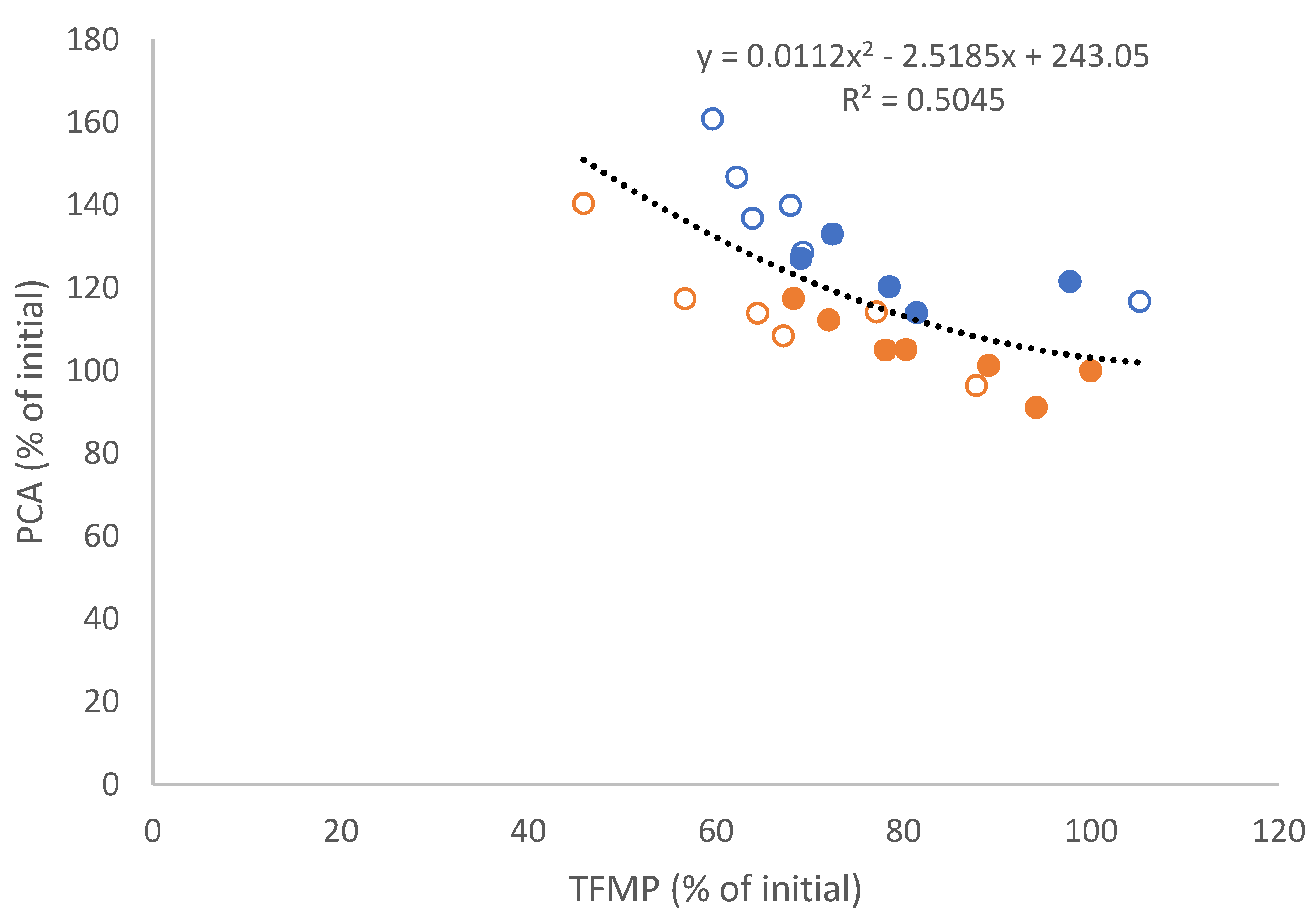
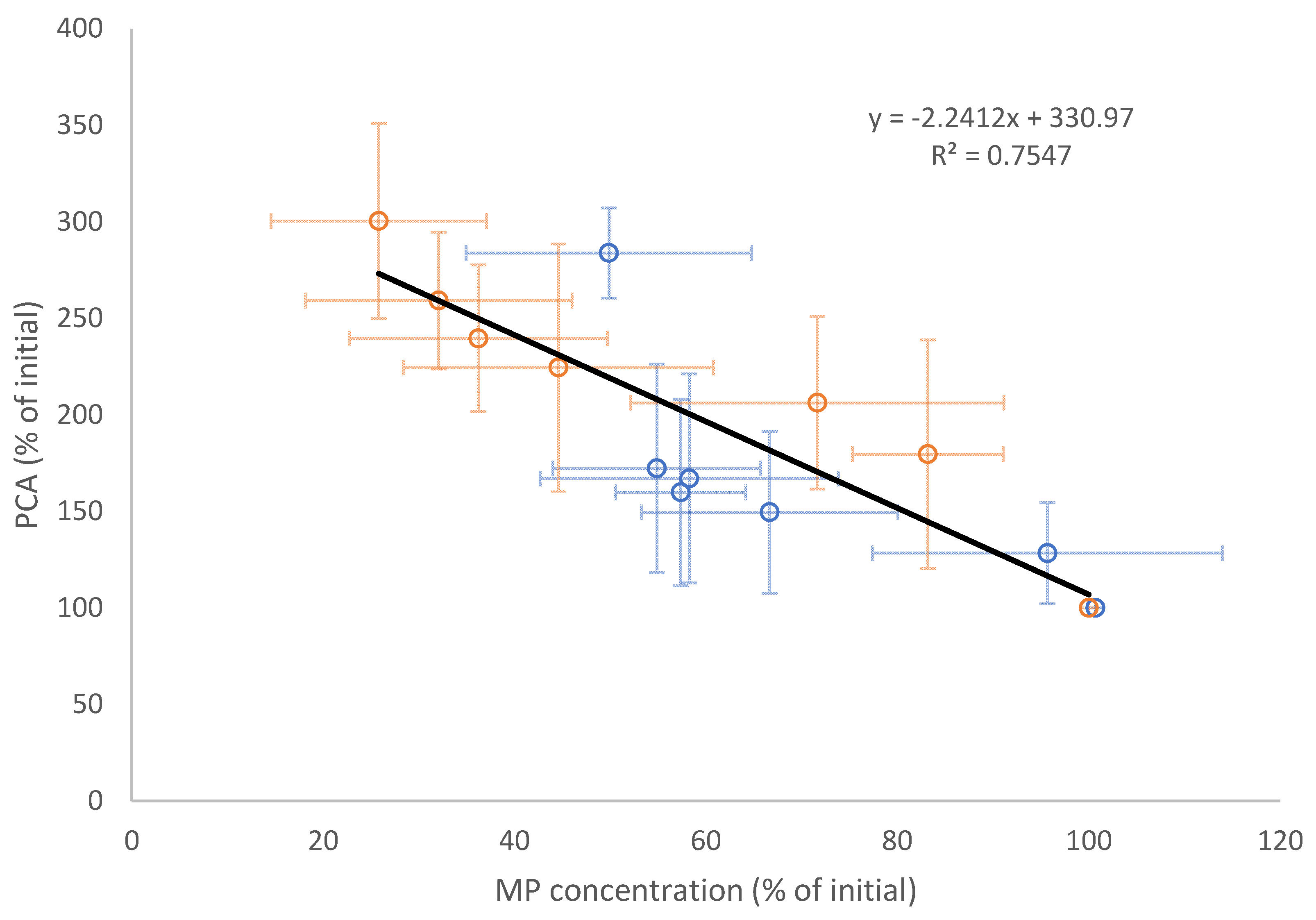
3.2. Tumour Spheroid Flow Experiments
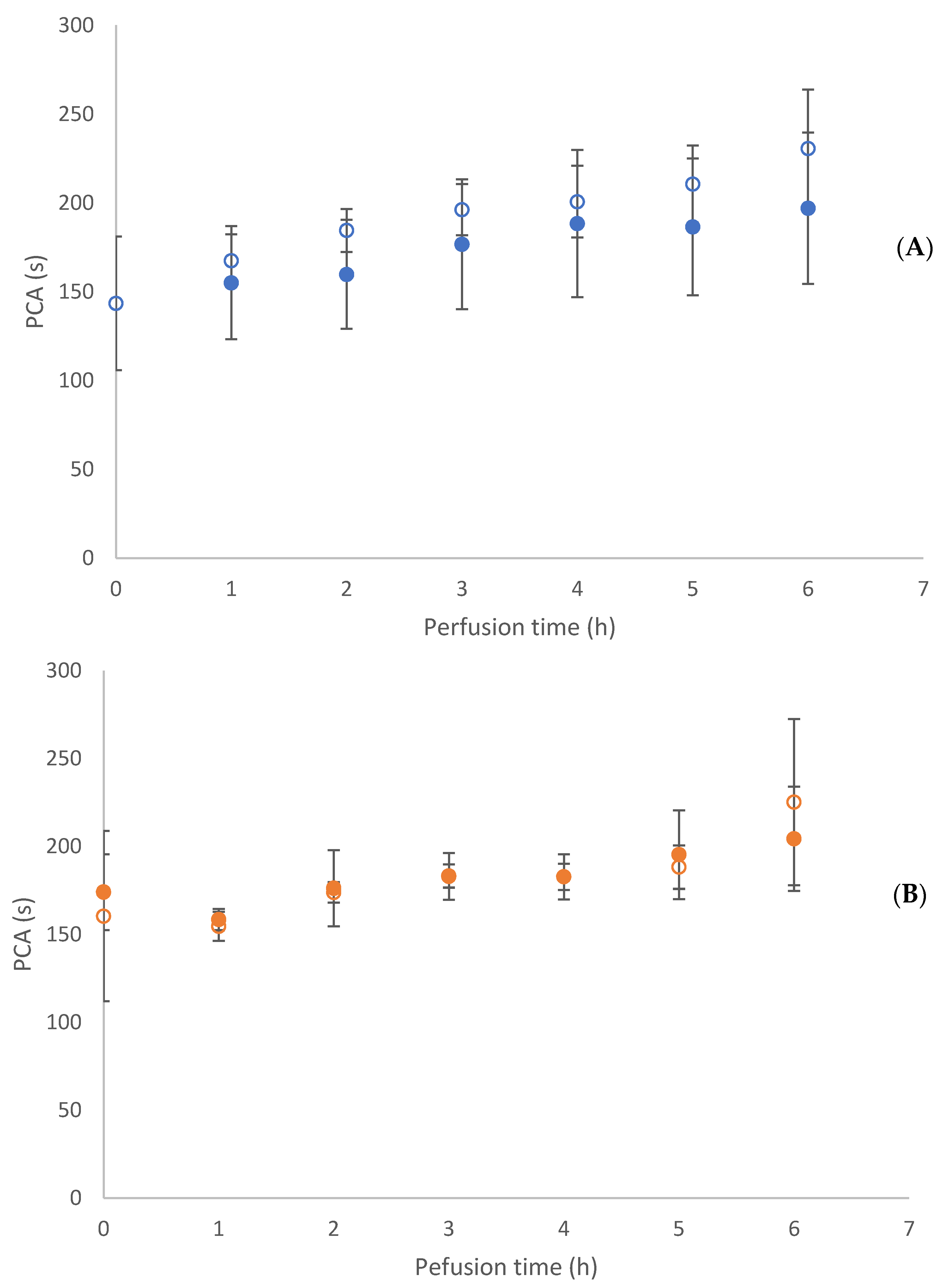
3.3. Flow Experiments Analysing HUVECs’ Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Seaman, S.; Nelson, A.; Noble, I. Cancer-associated thrombosis, low-molecular-weight heparin, and the patient experience: A qualitative study. Patient Prefer. Adherence 2014, 8, 453–461. [Google Scholar]
- Echrish, H.; Xiao, Y.; Madden, L.; Allgar, V.; Cooke, J.; Wedgwood, K.; Dasgupta, D.; Greenman, J.; Maraveyas, A. Effect of resection of localized pancreaticobiliary adenocarcinoma on angiogenic markers and tissue factor related pro-thrombotic and pro-angiogenic activity. Thromb. Res. 2014, 134, 479–487. [Google Scholar] [CrossRef]
- Aleman, M.M.; Gardiner, C.; Harrison, P.; Wolberg, A.S. Differential contributions of monocyte- and platelet-derived microparticles towards thrombin generation and fibrin formation and stability. J. Thromb. Haemost. 2011, 9, 2251–2261. [Google Scholar] [CrossRef] [Green Version]
- Shai, E.; Varon, D. Development, cell differentiation, angiogenesis—Microparticles and their roles in angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 10–14. [Google Scholar] [CrossRef]
- Campello, E.; Spiezia, L.; Radu, C.M.; Bulato, C.; Castelli, M.; Gavasso, S.; Simioni, P. Endothelial, platelet, and tissue factor-bearing microparticles in cancer patients with and without venous thromboembolism. Thromb. Res. 2011, 127, 473–477. [Google Scholar] [CrossRef]
- Brodsky, S.V.; Zhang, F.; Nasjletti, A.; Goligorsky, M.S. Endothelium-derived microparticles impair endothelial function in vitro. Am. J. Physiol. -Heart Circ. Physiol. 2004, 286, H1910–H1915. [Google Scholar] [CrossRef] [Green Version]
- Barteneva, N.S.; Fasler-Kan, E.; Bernimoulin, M.; Stern, J.N.H.; Ponomarev, E.D.; Duckett, L.; Vorobjev, I.A. Circulating microparticles: Square the circle. BMC Cell Biol. 2013, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Nomura, S.; Shimizu, M. Clinical significance of procoagulant microparticles. J. Intensive Care 2015, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Gold, B.; Cankovic, M.; Furtado, L.V.; Meier, F.; Gocke, C.D. Do Circulating Tumor Cells, Exosomes, and Circulating Tumor Nucleic Acids Have Clinical Utility? J. Mol. Diagn. 2015, 17, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Manly, D.A.; Boles, J.; Mackman, N. Role of tissue factor in venous thrombosis. Annu. Rev. Physiol. 2011, 73, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Yates, K.R.; Welsh, J.; Echrish, H.H.; Greenman, J.; Maraveyas, A.; Madden, L.A. Pancreatic cancer cell and microparticle procoagulant surface characterization: Involvement of membrane-expressed tissue factor, phosphatidylserine and phosphatidylethanolamine. Blood Coagul. Fibrinolysis 2011, 22, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Date, K.; Hall, J.; Greenman, J.; Maraveyas, A.; Madden, L.A. Tumour and microparticle tissue factor expression and cancer thrombosis. Thromb. Res. 2013, 131, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Haddad, T.C.; Greeno, E.W. Chemotherapy-induced thrombosis. Thromb. Res. 2006, 118, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Swystun, L.L.; Shin, L.Y.Y.; Beaudin, S.; Liaw, P.C. Chemotherapeutic agents doxorubicin and epirubicin induce a procoagulant phenotype on endothelial cells and blood monocytes. J. Thromb. Haemost. 2009, 7, 619–626. [Google Scholar] [CrossRef]
- Furie, B.C.; Furie, B. Cancer-associated thrombosis. Blood Cellsmolecules Dis. 2006, 36, 177–181. [Google Scholar] [CrossRef]
- Falanga, A.; Russo, L. Epidemiology, risk and outcomes of venous thromboembolism in cancer. Hamostaseologie 2012, 32, 115–125. [Google Scholar]
- Barni, S.; Labianca, R.; Agnelli, G.; Bonizzoni, E.; Verso, M.; Mandalà, M.; Brighenti, M.; Petrelli, F.; Bianchini, C.; Perrone, T. Chemotherapy-associated thromboembolic risk in cancer outpatients and effect of nadroparin thromboprophylaxis: Results of a retrospective analysis of the PROTECHT study. J. Transl. Med. 2011, 9, 179. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.A.; Adel, N.; Riedel, E.; Bhutani, M.; Feldman, D.R.; Tabbara, N.E.; Soff, G.; Parameswaran, R.; Hassoun, H. High incidence of thromboembolic events in patients treated with cisplatin-based chemotherapy: A large retrospective analysis. J. Clin. Oncol. 2011, 29, 3466–3473. [Google Scholar] [CrossRef]
- Nuver, J.; Smit, A.; Sleijfer, D.T.; Van Gessel, A.; Van Roon, A.; Van Der Meer, J.; van den Berg, M.; Burgerhof, J.; Hoekstra, H.; Sluiter, W. Microalbuminuria, decreased fibrinolysis, and inflammation as early signs of atherosclerosis in long-term survivors of disseminated testicular cancer. Eur. J. Cancer 2004, 40, 701–706. [Google Scholar] [CrossRef]
- Soultati, A.; Mountzios, G.; Avgerinou, C.; Papaxoinis, G.; Pectasides, D.; Dimopoulos, M.-A.; Papadimitriou, C. Endothelial vascular toxicity from chemotherapeutic agents: Preclinical evidence and clinical implications. Cancer Treat. Rev. 2012, 38, 473–483. [Google Scholar] [CrossRef]
- Hoshi, A.; Matsumoto, A.; Chung, J.; Isozumi, Y.; Koyama, T. Activation of coagulation by a thalidomide-based regimen. Blood Coagul. Fibrinolysis 2011, 22, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.; Wheeler, H.R.; Geczy, C.L. Modulation of tissue factor on human monocytes by cisplatin and adriamycin. Br. J. Haematol. 1992, 81, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Lim, K.M.; Kim, K.Y.; Bae, O.N.; Noh, J.Y.; Chung, S.M.; Shin, S.; Yun, Y.P.; Chung, J.H. Doxorubicin-induced platelet cytotoxicity: A new contributory factor for doxorubicin-mediated thrombocytopenia. J. Thromb. Haemost. 2009, 7, 1172–1183. [Google Scholar] [CrossRef]
- Ben Aharon, I.; Bar Joseph, H.; Tzabari, M.; Shenkman, B.; Farzam, N.; Levi, M.; Shalgi, R.; Stemmer, S.M.; Savion, N. Doxorubicin-Induced Vascular Toxicity—Targeting Potential Pathways May Reduce Procoagulant Activity. PLoS ONE 2013, 8, e75157. [Google Scholar] [CrossRef]
- Sontheimer-Phelps, A.; Hassell, B.A.; Ingber, D.E. Modelling cancer in microfluidic human organs-on-chips. Nat. Rev. Cancer 2019, 19, 65–81. [Google Scholar] [CrossRef]
- Garcia-Cordero, J.L.; Maerkl, S.J. Microfluidic systems for cancer diagnostics. Curr. Opin. Biotechnol. 2020, 65, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Adesanya, M.A.; Maraveyas, A.; Madden, L.A. Cancer microvesicles induce tissue factor-related procoagulant activity in endothelial cells in vitro. Blood Coagul. Fibrinolysis 2017, 28, 365–372. [Google Scholar] [CrossRef]
- Algarni, A.; Greenman, J.; Madden, L.A. Procoagulant tumor microvesicles attach to endothelial cells on biochips under microfluidic flow. Biomicrofluidics 2019, 13, 064124. [Google Scholar] [CrossRef]
- Cointe, S.; Judicone, C.; Robert, S.; Mooberry, M.; Poncelet, P.; Wauben, M.; Nieuwland, R.; Key, N.; Dignat-George, F.; Lacroix, R. Standardization of microparticle enumeration across different flow cytometry platforms: Results of a multicenter collaborative workshop. J. Thromb. Haemost. 2017, 15, 187–193. [Google Scholar] [CrossRef]
- Benameur, T.; Osman, A.; Parray, A.; Ait Hssain, A.; Munusamy, S.; Agouni, A. Molecular mechanisms underpinning microparticle-mediated cellular injury in cardiovascular complications associated with diabetes. Oxidative Med. Cell. Longev. 2019, 6475187. [Google Scholar] [CrossRef] [Green Version]
- Camussi, G.; Deregibus, M.C.; Bruno, S.; Cantaluppi, V.; Biancone, L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int. 2010, 78, 838–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, L.G.; Alqarni, S.; Maraveyas, A.; Madden, L.A. Isolated tumour microparticles induce endothelial microparticle release in vitro. Blood Coagul. Fibrinolysis 2020, 31, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AgouTii, A.; Ahmed Mostefai, H.; Porro, C.; Carusio, N.; Favre, J.; Richard, V.; Henrion, D.; Carmen Martinez, M.; Andriantsitohaina, R. Sonic hedgehog carried by microparticles corrects endothelial injury through nitric oxide release. FASEB J. 2007, 21, 2735–2741. [Google Scholar] [CrossRef] [Green Version]
- Benameur, T.; Soleti, R.; Porro, C.; Andriantsitohaina, R.; Martínez, M.C. Microparticles carrying Sonic hedgehog favor neovascularization through the activation of nitric oxide pathway in mice. PLoS ONE 2010, 5, e12688. [Google Scholar] [CrossRef] [Green Version]
- Barry, O.P.; Praticò, D.; Lawson, J.A.; FitzGerald, G.A. Transcellular activation of platelets and endothelial cells by bioactive lipids in platelet microparticles. J. Clin. Investig. 1997, 99, 2118–2127. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algarni, A.; Greenman, J.; Madden, L.A. Doxorubicin Enhances Procoagulant Activity of Endothelial Cells after Exposure to Tumour Microparticles on Microfluidic Devices. Hemato 2020, 1, 23-34. https://0-doi-org.brum.beds.ac.uk/10.3390/bloods1010006
Algarni A, Greenman J, Madden LA. Doxorubicin Enhances Procoagulant Activity of Endothelial Cells after Exposure to Tumour Microparticles on Microfluidic Devices. Hemato. 2020; 1(1):23-34. https://0-doi-org.brum.beds.ac.uk/10.3390/bloods1010006
Chicago/Turabian StyleAlgarni, Abdulrahman, John Greenman, and Leigh A. Madden. 2020. "Doxorubicin Enhances Procoagulant Activity of Endothelial Cells after Exposure to Tumour Microparticles on Microfluidic Devices" Hemato 1, no. 1: 23-34. https://0-doi-org.brum.beds.ac.uk/10.3390/bloods1010006