
Proteomic Characterization of Spontaneous Stress-Induced In Vitro Apoptosis of Human Acute Myeloid Leukemia Cells; Focus on Patient Heterogeneity and Endoplasmic Reticulum Stress

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Preparation
2.2. Cell Culture and Proteomic Cell Preparation
2.3. Analysis of AML Cell Viability and Proliferation
2.4. Liquid Chromatography (LC) Tandem Mass Spectrometry (MS) Analysis
2.5. Statistical and Bioinformatical Analyses
3. Results
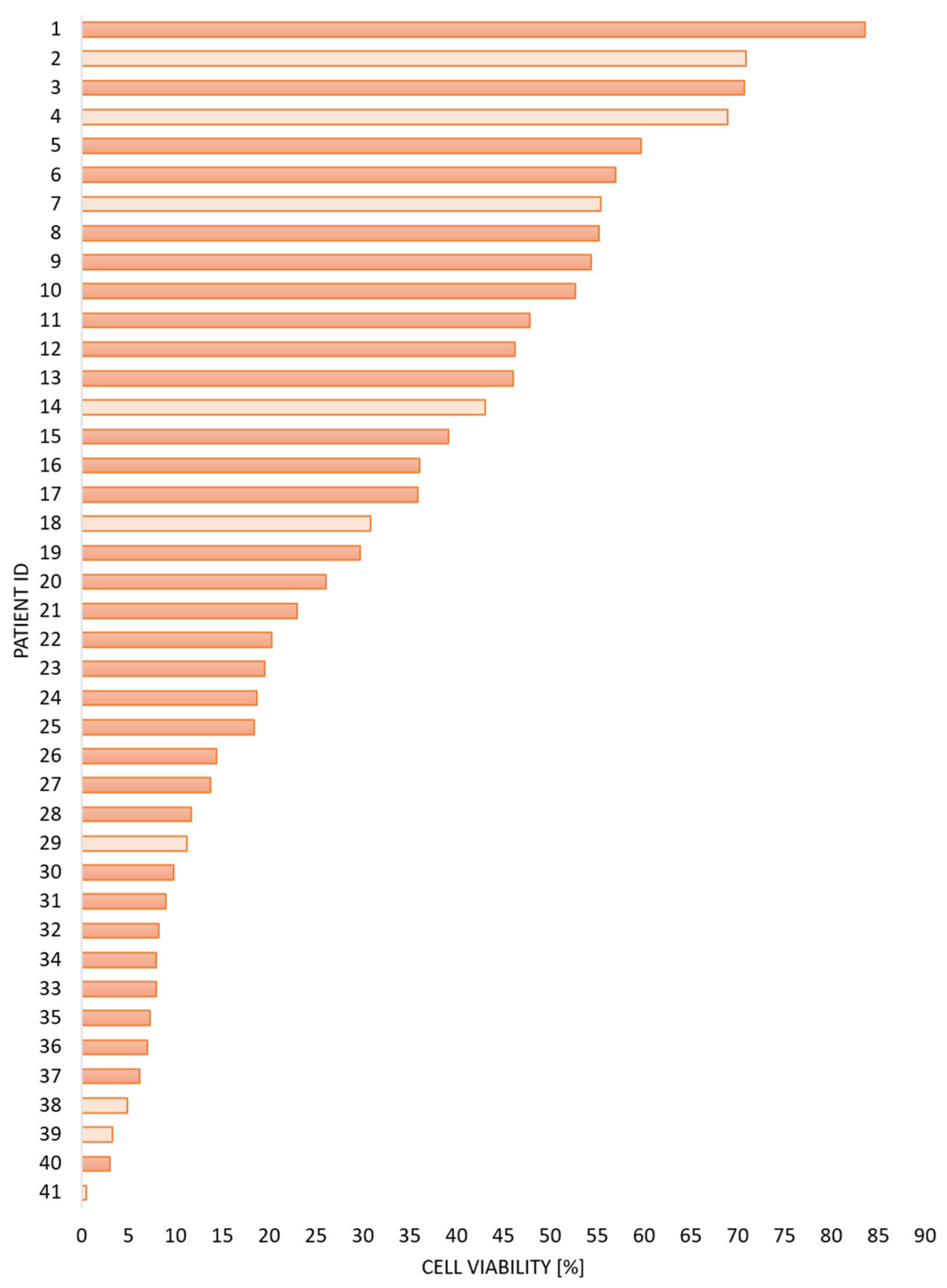
3.1. AML Patients Are Heterogeneous with Regard to Spondtaneous Apoptosis during In Vitro Culture of Their AML Cells
3.2. The Proteomic Analysis of AML Cells with High and Low Viability after In Vitro Culture
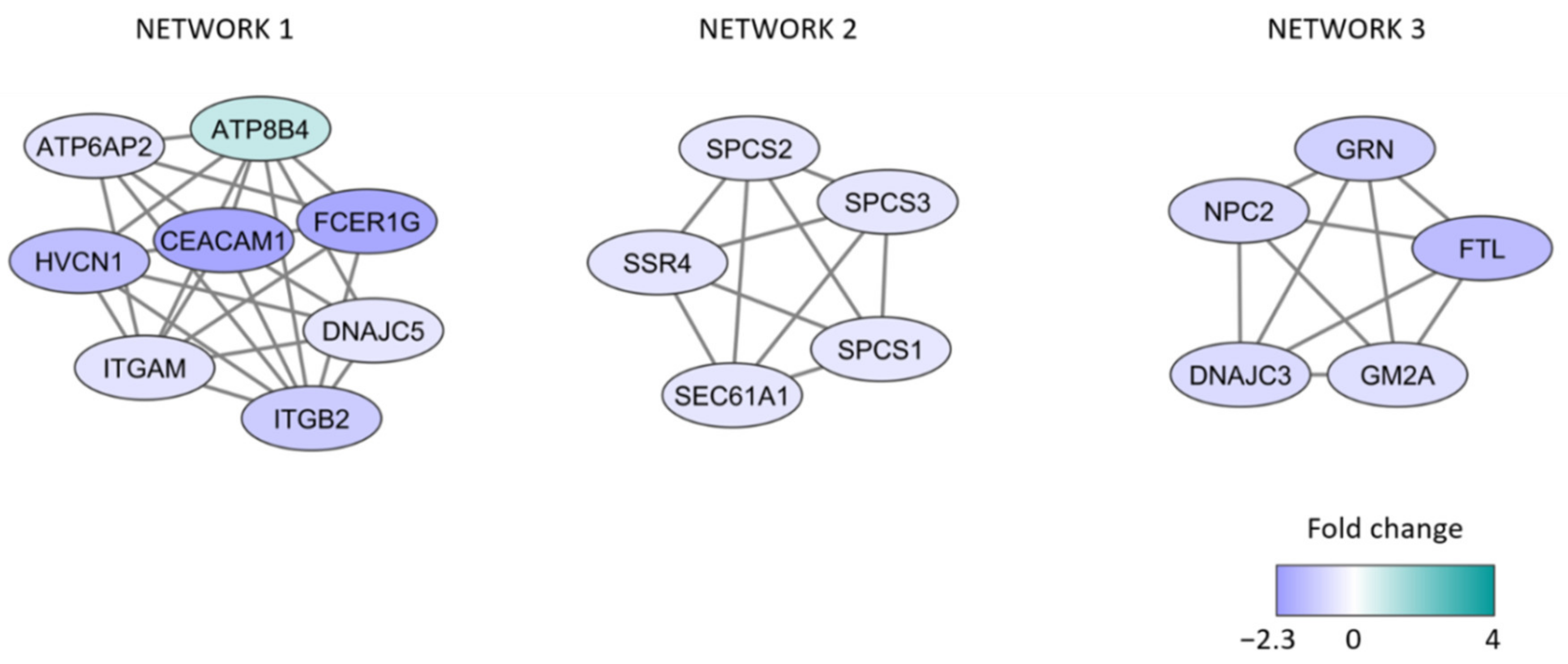
3.3. Identification of Proteins with Different Expression When Comparing Patients with High and Low AML Cell Viability after In Vitro Culture
- A majority of the proteins are endoplasmic reticulum proteins (ATP6AP2, ATP8B4, SSR4, DNAJC3, ITGM, ITGB2, SPCS1/2/3, SEC61A1, GRN, NPC2, GM2A).
- Several proteins are also involved in the endoplasmic reticulum stress response/unfolded protein response (ATP6AP, GRN, DNAJC3, possibly also ITGAM/ITGB2), function as chaperon (DNAJC5), or are increased during cellular stress (FTL).
- Many of the proteins are also involved in the regulation of apoptosis (HVCN1, ATP6AP, CEACAM1, GRN, FTL, DNAJC3).
- Finally, GM2A and ATP6AP2 are lysosomal proteins.
3.4. The BCL-2 Family
- BCL2, MCL1, and BID could be quantified for all 32 patients, whereas BAX was detected for all 17 low viability patients and all except one high viability patient. The levels did not differ between the two patient groups.
- BAK1 was expressed for 16 low and for 13 high viability patients. The levels did not differ between patients with high and low viability.
- BMF was quantified for a minority of both high viability (six patients) and low viability patients (five patients); the levels did not differ significantly between the groups.
- NOXA, BIK, HRK, BCLXL, BFL1, BIM, and PUMA could not be quantified for any patient.
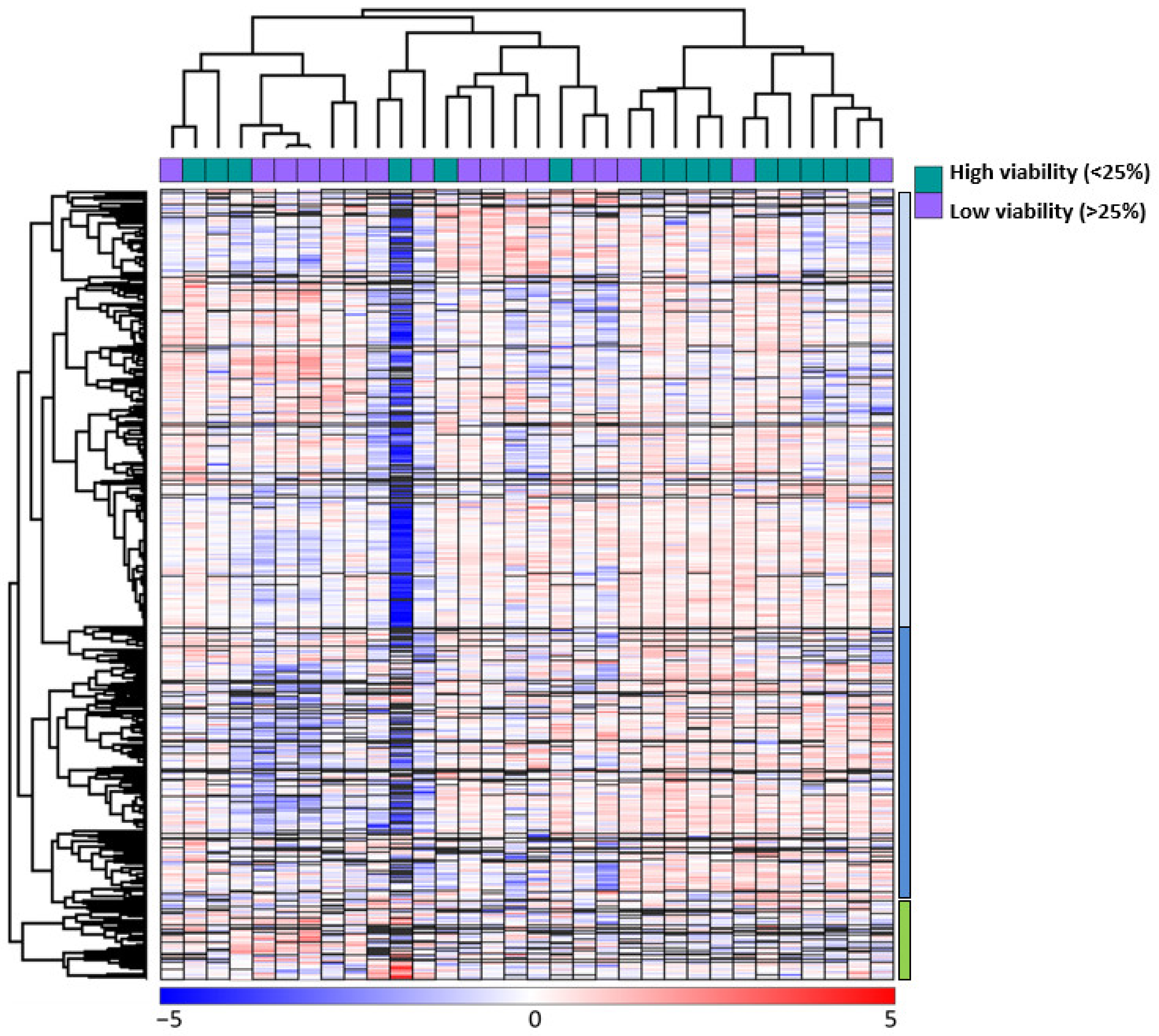
3.5. Unuspervised Hierarchical Clustering Analysis Based on the Global AML Cell Proteomic Profile during Ongoing Spontaneous Stress-Induced Apoptosis
3.6. Hierarchical Clustering Analysis Based on the Overall Proteomic Profiles of the AML Cells; the Associations between Bax:Bcl2 Balance and Patient Subclassification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef] [Green Version]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.A.; Fenaux, P.; Tallman, M.S.; Estey, E.H.; Löwenberg, B.; Naoe, T.; Lengfelder, E.; Döhner, H.; Burnett, A.K.; Chen, S.J.; et al. Management of acute promyelocytic leukemia: Updated recommendations from an expert panel of the European LeukemiaNet. Blood 2019, 133, 1630–1643. [Google Scholar] [CrossRef] [Green Version]
- Gjertsen, B.T.; Øyan, A.M.; Marzolf, B.; Hovland, R.; Gausdal, G.; Døskeland, S.O.; Dimitrov, K.; Golden, A.; Kalland, K.H.; Hood, L.; et al. Analysis of acute myelogenous leukemia: Preparation of samples for genomic and proteomic analyses. J. Hematother. Stem Cell Res. 2002, 11, 469–481. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Gjertsen, B.T.; Foss, B.; Huang, T.S. New strategies in the treatment of acute myelogenous leukemia (AML): In vitro culture of aml cells—The present use in experimental studies and the possible importance for future therapeutic approaches. Stem Cells 2001, 19, 1–11. [Google Scholar] [CrossRef]
- Griffiths, M.; Sundaram, H. Drug design and testing: Profiling of antiproliferative agents for cancer therapy using a cell-based methyl-[3H]-thymidine incorporation assay. Methods Mol. Biol. 2011, 731, 451–465. [Google Scholar] [PubMed]
- Cavanagh, B.L.; Walker, T.; Norazit, A.; Meedeniya, A.C. Thymidine analogues for tracking DNA synthesis. Molecules 2011, 16, 7980–7993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryningen, A.; Ersvaer, E.; Øyan, A.M.; Kalland, K.H.; Vintermyr, O.K.; Gjertsen, B.T.; Bruserud, Ø. Stress-induced in vitro apoptosis of native human acute myelogenous leukemia (AML) cells shows a wide variation between patients and is associated with low BCL-2:Bax ratio and low levels of heat shock protein 70 and 90. Leuk. Res. 2006, 30, 1531–1540. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Gjertsen, B.T.; von Volkman, H.L. In vitro culture of human acute myelogenous leukemia (AML) cells in serum-free media: Studies of native AML blasts and AML cell lines. J. Hematother. Stem Cell Res. 2000, 9, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Bruserud, Ø.; Frostad, S.; Foss, B. In vitro culture of acute myelogenous leukemia blasts: A comparison of four different culture media. J. Hematother. 1999, 8, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Brenner, A.K.; Aasebø, E.; Hernandez-Valladares, M.; Selheim, F.; Berven, F.; Grønningsæter, I.S.; Bartaula-Brevik, S.; Bruserud, Ø. The capacity of long-term in vitro proliferation of acute myeloid leukemia cells supported only by exogenous cytokines is associated with a patient subset with adverse outcome. Cancers 2019, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aasebø, E.; Berven, F.S.; Bartaula-Brevik, S.; Stokowy, T.; Hovland, R.; Vaudel, M.; Døskeland, S.O.; McCormack, E.; Batth, T.S.; Olsen, J.V.; et al. Proteome and phosphoproteome changes associated with prognosis in acute myeloid leukemia. Cancers 2020, 12, 709. [Google Scholar] [CrossRef] [Green Version]
- Bruserud, Ø.; Aasebø, E.; Hernandez-Valladares, M.; Tsykunova, G.; Reikvam, H. Therapeutic targeting of leukemic stem cells in acute myeloid leukemia—The biological background for possible strategies. Expert Opin. Drug Discov. 2017, 12, 1053–1065. [Google Scholar] [CrossRef]
- Hernandez-Valladares, M.; Bruserud, Ø.; Selheim, F. The implementation of mass spectrometry-based proteomics workflows in clinical routines of acute myeloid leukemia: Applicability and perspectives. Int. J. Mol. Sci. 2020, 21, 6830. [Google Scholar] [CrossRef]
- Aasebø, E.; Berven, F.S.; Hovland, R.; Døskeland, S.O.; Bruserud, Ø.; Selheim, F.; Hernandez-Valladares, M. The progression of acute myeloid leukemia from first diagnosis to chemoresistant relapse: A comparison of proteomic and phosphoproteomic profiles. Cancers 2020, 12, 1466. [Google Scholar] [CrossRef] [PubMed]
- Mer, A.S.; Lindberg, J.; Nilsson, C.; Klevebring, D.; Wang, M.; Grönberg, H.; Lehmann, S.; Rantalainen, M. Expression levels of long non-coding RNAs are prognostic for AML outcome. J. Hematol. Oncol. 2018, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stäubert, C.; Bhuiyan, H.; Lindahl, A.; Broom, O.J.; Zhu, Y.; Islam, S.; Linnarsson, S.; Lehtiö, J.; Nordström, A. Rewired metabolism in drug-resistant leukemia cells: A metabolic switch hallmarked by reduced dependence on exogenous glutamine. J. Biol. Chem. 2015, 290, 8348–8359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarevic, V.; Hörstedt, A.S.; Johansson, B.; Antunovic, P.; Billström, R.; Derolf, Å.; Lehmann, S.; Möllgård, L.; Peterson, S.; Stockelberg, D.; et al. Failure matters: Unsuccessful cytogenetics and unperformed cytogenetics are associated with a poor prognosis in a population-based series of acute myeloid leukaemia. Eur. J. Haematol. 2015, 94, 419–423. [Google Scholar] [CrossRef]
- Grønbæk, K.; Müller-Tidow, C.; Perini, G.; Lehmann, S.; Bach Treppendahl, M.; Mills, K.; Plass, C.; Schlegelberger, B. European Genomics and Epigenomics Study on MDS and AML (EuGESMA), COST Action BM0801. A critical appraisal of tools available for monitoring epigenetic changes in clinical samples from patients with myeloid malignancies. Haematologica 2012, 97, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Eppert, K.; Takenaka, K.; Lechman, E.R.; Waldron, L.; Nilsson, B.; van Galen, P.; Metzeler, K.H.; Poeppl, A.; Ling, V.; Beyene, J.; et al. Stem cell gene expression programs influence clinical outcome in human leukemia. Nat. Med. 2011, 17, 1086–1093. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Stapnes, C.; Doskeland, A.P.; Hatfield, K.; Ersvaer, E.; Ryningen, A.; Lorens, J.B.; Gjertsen, B.T.; Bruserud, Ø. The proteasome inhibitors bortezomib and PR-171 have antiproliferative and proapoptotic effects on primary human acute myeloid leukaemia cells. Br. J. Haematol. 2007, 136, 814–828. [Google Scholar] [CrossRef]
- Grønningsæter, I.S.; Reikvam, H.; Aasebø, E.; Bartaula-Brevik, S.; Tvedt, T.H.; Bruserud, Ø.; Hatfield, K.J. Targeting cellular metabolism in acute myeloid leukemia and the role of patient heterogeneity. Cells 2020, 9, 1155. [Google Scholar] [CrossRef] [PubMed]
- Aasebø, E.; Birkeland, E.; Selheim, F.; Berven, F.; Brenner, A.K.; Bruserud, Ø. The extracellular bone marrow microenvironment—A proteomic comparison of constitutive protein release by in vitro cultured osteoblasts and mesenchymal stem cells. Cancers 2020, 13, 62. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Arntzen, M.Ø.; Koehler, C.J.; Barsnes, H.; Berven, F.S.; Treumann, A.; Thiede, B.; Isobari, Q. Software for isobaric quantitative proteomics using IPTL, iTRAQ, and TMT. J. Proteome Res. 2011, 10, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Scholz, C.; Lyon, D.; Refsgaard, J.C.; Jensen, L.J.; Choudhary, C.; Weinert, B.T. Avoiding abundance bias in the functional annotation of post-translationally modified proteins. Nat. Methods 2015, 12, 1003–1004. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetz, C.; Papa, F.R. The unfolded protein response and cell fate control. Mol. Cell 2018, 69, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chipurupalli, S.; Kannan, E.; Tergaonkar, V.; D’Andrea, R.; Robinson, N. Hypoxia induced ER stress response as an adaptive mechanism in cancer. Int. J. Mol. Sci. 2019, 20, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef]
- Leibowitz, B.; Yu, J. Mitochondrial signaling in cell death via the Bcl-2 family. Cancer Biol. Ther. 2010, 9, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Hata, A.N.; Engelman, J.A.; Faber, A.C. The BCL2 family: Key mediators of the apoptotic response to targeted anticancer therapeutics. Cancer Discov. 2015, 5, 475–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Abrams, J.M.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; Dawson, T.M.; Dawson, V.L.; El-Deiry, W.S.; Fulda, S.; et al. Molecular definitions of cell death subroutines: Recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ. 2012, 19, 107–120. [Google Scholar] [CrossRef]
- Pearce, D.J.; Taussig, D.; Zibara, K.; Smith, L.L.; Ridler, C.M.; Preudhomme, C.; Young, B.D.; Rohatiner, A.Z.; Lister, T.A.; Bonnet, D. AML engraftment in the NOD/SCID assay reflects the outcome of AML: Implications for our understanding of the heterogeneity of AML. Blood 2006, 107, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Sharawat, S.K.; Bakhshi, R.; Vishnubhatla, S.; Gupta, R.; Bakhshi, S. BAX/BCL2 RMFI ratio predicts better induction response in pediatric patients with acute myeloid leukemia. Pediatr. Blood Cancer 2013, 60, E63–E66. [Google Scholar] [CrossRef]
- Kornblau, S.M.; Vu, H.T.; Ruvolo, P.; Estrov, Z.; O’Brien, S.; Cortes, J.; Kantarjian, H.; Andreeff, M.; May, W.S. BAX and PKCalpha modulate the prognostic impact of BCL2 expression in acute myelogenous leukemia. Clin. Cancer Res. 2000, 6, 1401–1409. [Google Scholar]
- Del Poeta, G.; Venditti, A.; Del Principe, M.I.; Maurillo, L.; Buccisano, F.; Tamburini, A.; Cox, M.C.; Franchi, A.; Bruno, A.; Mazzone, C.; et al. Amount of spontaneous apoptosis detected by Bax/Bcl-2 ratio predicts outcome in acute myeloid leukemia (AML). Blood 2003, 101, 2125–2131. [Google Scholar] [CrossRef]
- Deng, G.; Lane, C.; Kornblau, S.; Goodacre, A.; Snell, V.; Andreeff, M.; Deisseroth, A.B. Ratio of bcl-xshort to bcl-xlong is different in good- and poor-prognosis subsets of acute myeloid leukemia. Mol. Med. 1998, 4, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ong, Y.L.; McMullin, M.F.; Bailie, K.E.; Lappin, T.R.; Jones, F.G.; Irvine, A.E. High bax expression is a good prognostic indicator in acute myeloid leukaemia. Br. J. Haematol. 2000, 111, 182–189. [Google Scholar] [PubMed]
- Reichenbach, F.; Wiedenmann, C.; Schalk, E.; Becker, D.; Funk, K.; Scholz-Kreisel, P.; Todt, F.; Wolleschak, D.; Döhner, K.; Marquardt, J.U.; et al. Mitochondrial BAX determines the predisposition to apoptosis in human AML. Clin. Cancer Res. 2017, 23, 4805–4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Poeta, G.; Ammatuna, E.; Lavorgna, S.; Capelli, G.; Zaza, S.; Luciano, F.; Ottone, T.; Del Principe, M.I.; Buccisano, F.; Maurillo, L.; et al. The genotype nucleophosmin mutated and FLT3-ITD negative is characterized by high bax/bcl-2 ratio and favourable outcome in acute myeloid leukaemia. Br. J. Haematol. 2010, 149, 383–387. [Google Scholar] [CrossRef]
- Schaich, M.; Illmer, T.; Seitz, G.; Mohr, B.; Schäkel, U.; Beck, J.F.; Ehninger, G. The prognostic value of Bcl-XL gene expression for remission induction is influenced by cytogenetics in adult acute myeloid leukemia. Haematologica 2001, 86, 470–477. [Google Scholar]
- Duval, A.; Olaru, D.; Campos, L.; Flandrin, P.; Nadal, N.; Guyotat, D. Expression and prognostic significance of heat-shock proteins in myelodysplastic syndromes. Haematologica 2006, 91, 713–714. [Google Scholar]
- Thomas, X.; Campos, L.; Mounier, C.; Cornillon, J.; Flandrin, P.; Le, Q.H.; Piselli, S.; Guyotat, D. Expression of heat-shock proteins is associated with major adverse prognostic factors in acute myeloid leukemia. Leuk. Res. 2005, 29, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Löwenberg, B.; van Putten, W.L.; Touw, I.P.; Delwel, R.; Santini, V. Autonomous proliferation of leukemic cells in vitro as a determinant of prognosis in adult acute myeloid leukemia. N. Engl. J. Med. 1993, 328, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, W.J.; Löwenberg, B.; van Putten, W.L.; Ploemacher, R.E. Improved prognostic significance of cytokine-induced proliferation in vitro in patients with de novo acute myeloid leukemia of intermediate risk: Impact of internal tandem duplications in the Flt3 gene. Leukemia 2001, 15, 1046–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griessinger, E.; Anjos-Afonso, F.; Pizzitola, I.; Rouault-Pierre, K.; Vargaftig, J.; Taussig, D.; Gribben, J.; Lassailly, F.; Bonnet, D. A niche-like culture system allowing the maintenance of primary human acute myeloid leukemia-initiating cells: A new tool to decipher their chemoresistance and self-renewal mechanisms. Stem Cells Transl. Med. 2014, 3, 520–529. [Google Scholar] [CrossRef]
- Buteyn, N.J.; Santhanam, R.; Merchand-Reyes, G.; Murugesan, R.A.; Dettorre, G.M.; Byrd, J.C.; Sarkar, A.; Vasu, S.; Mundy-Bosse, B.L.; Butchar, J.P.; et al. Activation of the intracellular pattern recognition receptor NOD2 promotes acute myeloid leukemia (AML) cell apoptosis and provides a survival advantage in an animal model of AML. J. Immunol. 2020, 204, 1988–1997. [Google Scholar] [CrossRef] [PubMed]
- Karakas, T.; Miething, C.C.; Maurer, U.; Weidmann, E.; Ackermann, H.; Hoelzer, D.; Bergmann, L. The coexpression of the apoptosis-related genes bcl-2 and wt1 in predicting survival in adult acute myeloid leukemia. Leukemia 2002, 16, 846–854. [Google Scholar] [CrossRef] [Green Version]
- Haes, I.; Dendooven, A.; Mercier, M.L.; Puylaert, P.; Vermeulen, K.; Kockx, M.; Deiteren, K.; Maes, M.B.; Berneman, Z.; Anguille, S. Absence of BCL-2 expression identifies a subgroup of AML with distinct phenotypic, molecular, and clinical characteristics. J. Clin. Med. 2020, 9, 3090. [Google Scholar] [CrossRef]
- Lee, M.G.; Liu, Y.C.; Lee, Y.L.; El-Shazly, M.; Lai, K.H.; Shih, S.P.; Ke, S.; Hong, M.C.; Du, Y.C.; Yang, J.C.; et al. Heteronemin, a marine sesterterpenoid-type metabolite, induces apoptosis in prostate LNcap cells via oxidative and ER stress combined with the inhibition of topoisomerase II and Hsp90. Mar. Drugs 2018, 16, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. Cytotoxic effect of celastrol alone or in combination with paclitaxel on anaplastic thyroid carcinoma cells. Tumor Biol. 2017, 39, 1010428317698369. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Xu, H.; Yang, Y.; Zhao, D.; Wen, Y.; Lv, C.; Qiu, H.; Wang, C. The regulation of miR-320a/XBP1 axis through LINC00963 for endoplasmic reticulum stress and autophagy in diffuse large B-cell lymphoma. Cancer Cell Int. 2021, 21, 305. [Google Scholar] [CrossRef]
- Rahmani, M.; Mayo, M.; Dash, R.; Sokhi, U.K.; Dmitriev, I.P.; Sarkar, D.; Dent, P.; Curiel, D.T.; Fisher, P.B.; Grant, S. Melanoma differentiation associated gene-7/interleukin-24 potently induces apoptosis in human myeloid leukemia cells through a process regulated by endoplasmic reticulum stress. Mol. Pharmacol. 2010, 78, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Pae, H.O.; Jeong, S.O.; Jeong, G.S.; Kim, K.M.; Kim, H.S.; Kim, S.A.; Kim, Y.C.; Kang, S.D.; Kim, B.N.; Chung, H.T. Curcumin induces pro-apoptotic endoplasmic reticulum stress in human leukemia HL-60 cells. Biochem. Biophys. Res. Commun. 2007, 353, 1040–1045. [Google Scholar] [CrossRef]
- De Sá Bacelar, T.; da Silva, A.J.; Costa, P.R.; Rumjanek, V.M. The pterocarpanquinone LQB 118 induces apoptosis in tumor cells through the intrinsic pathway and the endoplasmic reticulum stress pathway. Anticancer Drugs 2013, 24, 73–83. [Google Scholar] [CrossRef]
- Navid, F.; Colbert, R.A. Causes and consequences of endoplasmic reticulum stress in rheumatic disease. Nat. Rev. Rheumatol. 2017, 13, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, E.; Logue, S.E.; Healy, S.J.; Manie, S.; Samali, A. The role of the unfolded protein response in cancer progression: From oncogenesis to chemoresistance. Biol. Cell 2019, 111, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Kim, D.S.; Kim, H.T.; Lee, J.W.; Chung, C.H.; Ahn, T.; Lim, J.M.; Kim, I.K.; Chae, H.J.; Kim, H.R. Enhanced lysosomal activity is involved in Bax inhibitor-1-induced regulation of the endoplasmic reticulum (ER) stress response and cell death against ER stress: Involvement of vacuolar H+-ATPase (V-ATPase). J. Biol. Chem. 2011, 286, 24743–24753. [Google Scholar] [CrossRef] [Green Version]
- Scherer, O.; Steinmetz, H.; Kaether, C.; Weinigel, C.; Barz, D.; Kleinert, H.; Menche, D.; Müller, R.; Pergola, C.; Werz, O. Targeting V-ATPase in primary human monocytes by archazolid potently represses the classical secretion of cytokines due to accumulation at the endoplasmic reticulum. Biochem. Pharmacol. 2014, 91, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Iurlaro, R.; Muñoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [Green Version]
- Casado, P.; Wilkes, E.H.; Miraki-Moud, F.; Hadi, M.M.; Rio-Machin, A.; Rajeeve, V.; Pike, R.; Iqbal, S.; Marfa, S.; Lea, N.; et al. Proteomic and genomic integration identifies kinase and differentiation determinants of kinase inhibitor sensitivity in leukemia cells. Leukemia 2018, 32, 1818–1822. [Google Scholar] [CrossRef]
- Schardt, J.A.; Mueller, B.U.; Pabst, T. Activation of the unfolded protein response in human acute myeloid leukemia. Methods Enzymol. 2011, 489, 227–243. [Google Scholar]
- Schardt, J.A.; Weber, D.; Eyholzer, M.; Mueller, B.U.; Pabst, T. Activation of the unfolded protein response is associated with favorable prognosis in acute myeloid leukemia. Clin. Cancer Res. 2009, 15, 3834–3841. [Google Scholar] [CrossRef] [Green Version]
- Rouault-Pierre, K.; Lopez-Onieva, L.; Foster, K.; Anjos-Afonso, F.; Lamrissi-Garcia, I.; Serrano-Sanchez, M.; Mitter, R.; Ivanovic, Z.; de Verneuil, H.; Gribben, J.; et al. HIF-2α protects human hematopoietic stem/progenitors and acute myeloid leukemic cells from apoptosis induced by endoplasmic reticulum stress. Cell Stem Cell 2013, 13, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Martinez, E.; Di Marcantonio, D.; Solanki-Patel, N.; Aghayev, T.; Peri, S.; Ferraro, F.; Skorski, T.; Scholl, C.; Fröhling, S.; et al. JUN is a key transcriptional regulator of the unfolded protein response in acute myeloid leukemia. Leukemia 2017, 31, 1196–1205. [Google Scholar] [CrossRef] [Green Version]
- Akkök, C.A.; Liseth, K.; Nesthus, I.; Løkeland, T.; Tefre, K.; Bruserud, O.; Abrahamsen, J.F. Autologous peripheral blood progenitor cells cryopreserved with 5 and 10 percent dimethyl sulfoxide alone give comparable hematopoietic reconstitution after transplantation. Transfusion 2008, 48, 877–883. [Google Scholar] [CrossRef]
- Abrahamsen, J.F.; Rusten, L.; Bakken, A.M.; Bruserud, Ø. Better preservation of early hematopoietic progenitor cells when human peripheral blood progenitor cells are cryopreserved with 5 percent dimethylsulfoxide instead of 10 percent dimethylsulfoxide. Transfusion 2004, 44, 785–789. [Google Scholar] [CrossRef]
- Sasnoor, L.M.; Kale, V.P.; Limaye, L.S. Prevention of apoptosis as a possible mechanism behind improved cryoprotection of hematopoietic cells by catalase and trehalose. Transplantation 2005, 80, 1251–1260. [Google Scholar] [CrossRef]
- Sasnoor, L.M.; Kale, V.P.; Limaye, L.S. Supplementation of conventional freezing medium with a combination of catalase and trehalose results in better protection of surface molecules and functionality of hematopoietic cells. J. Hematother. Stem Cell Res. 2003, 12, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Khetan, S.; Corey, O. Maintenance of stem cell viability and differentiation potential following cryopreservation within 3-dimensional hyaluronic acid hydrogels. Cryobiology 2019, 90, 83–88. [Google Scholar] [CrossRef]
- Miyazaki, T.; Suemori, H. Slow cooling cryopreservation optimized to human pluripotent stem cells. Adv. Exp. Med. Biol. 2016, 951, 57–65. [Google Scholar] [PubMed]
- Demirci, S.; Doğan, A.; Şişli, B.; Sahin, F. Boron increases the cell viability of mesenchymal stem cells after long-term cryopreservation. Cryobiology 2014, 68, 139–146. [Google Scholar] [CrossRef]
- Bissoyi, A.; Nayak, B.; Pramanik, K.; Sarangi, S.K. Targeting cryopreservation-induced cell death: A review. Biopreserv. Biobank. 2014, 12, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Shibamiya, A.; Schulze, E.; Krauß, D.; Augustin, C.; Reinsch, M.; Schulze, M.L.; Steuck, S.; Mearini, G.; Mannhardt, I.; Schulze, T.; et al. Cell banking of hiPSCs: A practical guide to cryopreservation and quality control in basic research. Curr. Protoc. Stem Cell Biol. 2020, 55, e127. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Pelosi, E.; Castelli, G. Precision medicine treatment in acute myeloid leukemia is not a dream. Hemato 2021, 2, 8. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sex and Age (n = 41) | Cytogenetic Abnormalities | ||
|---|---|---|---|
| Males/females | 22/19 | Normal karyotype | 21 |
| Age (years; median/range) | 70/18–87 | Favorable | 4 |
| Intermediate | 9 | ||
| Predisposition/previous disease | Adverse | 4 | |
| Previous chronic myeloid neoplasia | 1 | Not tested | 3 |
| Myelodysplastic syndrome | 8 | ||
| Relapsed AML | 3 | FLT3 status | |
| Chemotherapy | 0 | ITD | 14 |
| Normal | 19 | ||
| Morphology/FAB classification | Not tested | 8 | |
| M0/M1 | 17 | ||
| M2 | 8 | NPM1 status | |
| M4/M5 | 16 | Insertion | 14 |
| M6/M7 | 0 | Insertion + Flt3-ITD | 8 |
| Normal | 20 | ||
| CD34 expression | 22 | Not tested | 7 |
| Term | Description | FDR | Category | Foreground Count | Background Count | p-Value | s-Value |
|---|---|---|---|---|---|---|---|
| High expression in patients with high viability/low spontaneous apoptosis | |||||||
| KW-0539 | Nucleus | 0.0017 | UniProt keywords | 115 | 2597 | 2.13 × 10−6 | 1.32 |
| KW-0832 | Ubl conjugation | 0.0017 | UniProt keywords | 72 | 1443 | 2.11 × 10−6 | 0.98 |
| KW-0805 | Transcription regulation | 0.0017 | UniProt keywords | 52 | 829 | 1.50 × 10−6 | 0.90 |
| KW-0804 | Transcription | 0.0017 | UniProt keywords | 53 | 877 | 1.46 × 10−6 | 0.89 |
| KW-1017 | Isopeptide bond | 0.0017 | UniProt keywords | 59 | 1135 | 2.54 × 10−6 | 0.83 |
| KW-0863 | Zinc-finger | 0.0018 | UniProt keywords | 38 | 586 | 2.37 × 10−6 | 0.65 |
| High expression in patients with low viability/high spontaneous apoptosis | |||||||
| KW-0675 | Receptor | 0.00032 | UniProt keywords | 14 | 174 | 2.77 × 10−7 | 0.92 |
| KW-0256 | Endoplasmic reticulum | 0.00033 | UniProt keywords | 28 | 644 | 5.61 × 10−7 | 1.49 |
| KW-0472 | Membrane | 0.00041 | UniProt keywords | 65 | 2341 | 1.25 × 10−6 | 2.55 |
| KW-1133 | Transmembrane helix | 0.00041 | UniProt keywords | 50 | 1217 | 2.13 × 10−6 | 2.36 |
| KW-0732 | Signal | 0.00041 | UniProt keywords | 37 | 658 | 1.04 × 10−6 | 2.05 |
| KW-1015 | Disulfide bond | 0.00041 | UniProt keywords | 35 | 665 | 1.18 × 10−6 | 1.88 |
| Term | Description | FDR | Category | Foreground Count | Background Count | p-Value | s-Value |
|---|---|---|---|---|---|---|---|
| Network 1 | |||||||
| KW-1003 | Cell membrane | 4.28 × 10−5 | UniProt | 8 | 824 | 3.7 × 10−8 | 6.57 |
| GOCC:0030141 | Secretory granule | 0.00017 | CC-TM | 8 | 468 | 4.6 × 10−8 | 6.85 |
| GOCC:0098588 | Bounding membrane of organelle | 0.00017 | CC-TM | 8 | 844 | 4.4 × 10−8 | 6.47 |
| GOCC:0030659 | Cytoplasmic vesicle membrane | 0.00017 | CC-TM | 8 | 331 | 3.1 × 10−7 | 6.21 |
| GOCC:0030667 | Secretory granule membrane | 0.00017 | CC-TM | 8 | 175 | 5.9 × 10−7 | 6.08 |
| GOCC:0098805 | Whole membrane | 0.00017 | CC-TM | 8 | 954 | 1.2 × 10−7 | 6.00 |
| Network 2 | CC-TM | ||||||
| HSA-72766 | Translation | 0.00038 | Reactome | 5 | 279 | 1.0 × 10−7 | 6.71 |
| HSA-1799339 | SRP-dependent cotranslational protein targeting to membrane | 0.00080 | Reactome | 5 | 103 | 2.6 × 10−7 | 6.49 |
| GOCC:0005787 | Signal peptidase complex | 3.76 × 10−5 | CC-TM | 3 | 5 | 9.6 × 10−9 | 4.81 |
| map03060 | Protein export | 0.00013 | KEGG | 4 | 19 | 3.0 × 10−7 | 5.20 |
| GOCC:0005789 | Endoplasmic reticulum membrane | 0.00024 | CC-TM | 5 | 290 | 1.2 × 10−7 | 6.62 |
| HSA-400511 | Synthesis, secretion, and inactivation of GPI | 0.00026 | Reactome | 3 | 6 | 1.4 × 10−8 | 4.70 |
| Network 3 | |||||||
| GOCC:0034774 | Secretory granule lumen | 3.2 × 10−5 | CC-TM | 5 | 167 | 8.2 × 10−9 | 7.90 |
| GOCC:0035578 | Azurophil granule lumen | 9.68 × 10−5 | CC-TM | 5 | 80 | 9.8 × 10−8 | 6.93 |
| GO:0005764 | Lysosome | 0.0036 | CC | 5 | 465 | 1.3 × 10−6 | 5.50 |
| Term | Description | FDR | Category | Foreground Count | Background Count | p-Value | s-Value |
|---|---|---|---|---|---|---|---|
| Upregulated in the right patient cluster B (n = 12) | |||||||
| KW-0832 | Ubl conjugation | 0.0019 | UniProt | 53 | 1465 | 1.77 × 10−6 | 1.16 |
| KW-1017 | Isopeptide bond | 0.0019 | UniProt | 44 | 1148 | 3.27 × 10−6 | 0.97 |
| KW-0805 | Transcription regulation | 0.0019 | UniProt | 37 | 842 | 1.84 × 10−6 | 0.95 |
| KW-0804 | Transcription | 0.0019 | UniProt | 37 | 891 | 1.62 × 10−6 | 0.92 |
| KW-0863 | Zinc-finger | 0.0053 | UniProt | 26 | 597 | 4.10 × 10−5 | 0.51 |
| GO:0003700 | DNA-binding transcription factor activity | 0.0095 | GO MF | 19 | 223 | 1.6 × 10−6 | 068 |
| Downregulated in the right patient cluster (n = 12) | |||||||
| GO:0005886 | plasma membrane | 0.00084 | GO CC | 60 | 1625 | 9.48 × 10−7 | 2.83 |
| GO:0005576 | extracellular region | 0.00084 | OG CC | 52 | 1624 | 9.05 × 10−7 | 2.28 |
| GO:0005615 | extracellular space | 0.00095 | GO CC | 49 | 1414 | 2.06 × 10−6 | 2.11 |
| GO:0031410 | cytoplasmic vesicle | 0.00085 | GO CC | 40 | 1286 | 1.54 × 10−6 | 1.65 |
| GO:0030312 | external encapsulating structure | 0.00079 | GO CC | 11 | 117 | 2.83 × 10−7 | 0.73 |
| GO:0031012 | extracellular matrix | 0.00079 | GO CC | 11 | 117 | 2.83 × 10−7 | 0.73 |
| Term | Description | FDR | Category | Foreground Count | Background Count | p-Value | s-Value |
|---|---|---|---|---|---|---|---|
| Protein cluster 1A | |||||||
| KW-0472 | Membrane | 0.018 | UniProt | 1721 | 2530 | 1.42 × 10−5 | 0,20 |
| KW-0256 | Endoplasmic reticulum | 0.017 | UniProt | 520 | 671 | 1.82 × 10−5 | 0.12 |
| KW-0813 | Transport | 0.033 | UniProt | 718 | 991 | 8.39 × 10−5 | 0.11 |
| GOCC:0012505 | Endomembrane system | 0.036 | GO CC-TM | 1404 | 2001 | 1.08 × 10−5 | 0.21 |
| GOCC:0016020 | Membrane | 0.036 | GO CC-TM | 1736 | 2551 | 1.35 × 10−5 | 0.20 |
| GOCC:0031090 | Organelle membrane | 0.036 | GO CC-TM | 1165 | 1619 | 9.22 × 10−6 | 0.20 |
| Protein cluster 1B | |||||||
| KW-0539 | Nucleus | 0.0044 | UniProt | 1078 | 2737 | 8.27 × 10−6 | 0.44 |
| KW-0804 | Transcription | 0.0044 | UniProt | 438 | 929 | 5.85 × 10−6 | 0.31 |
| KW-0805 | Transcription regulation | 0.0044 | UniProt | 415 | 879 | 5.79 × 10−6 | 0.30 |
| KW-0238 | DNA-binding | 0.0044 | UniProt | 305 | 625 | 5.22 × 10−6 | 0.23 |
| KW-0832 | Ubl conjugation | 0.0044 | UniProt | 579 | 1507 | 2.31 × 10−5 | 0.19 |
| KW-0863 | Zinc-finger | 0.0044 | UniProt | 275 | 626 | 9.99 × 10−6 | 0.16 |
| Protein cluster 2 | |||||||
| KW-0325 | Glycoprotein | 0.0012 | UniProt | 172 | 1034 | 3.60 × 10−6 | 0.69 |
| KW-1015 | Disulfide bond | 0.0012 | UniProt | 142 | 791 | 3.25 × 10−6 | 0.62 |
| KW-1003 | Cell membrane | 0.0012 | UniProt | 153 | 929 | 3.01 × 10−6 | 0.62 |
| KW-0732 | Signal | 0.0012 | UniProt | 131 | 771 | 3.16 × 10−6 | 0.54 |
| KW-0472 | Membrane | 0.0012 | UniProt | 284 | 2530 | 4.91 × 10−6 | 0.53 |
| KW-0964 | Secreted | 0.0012 | UniProt | 76 | 372 | 2.37 × 10−6 | 0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aasebø, E.; Brenner, A.K.; Hernandez-Valladares, M.; Birkeland, E.; Reikvam, H.; Selheim, F.; Berven, F.S.; Bruserud, Ø. Proteomic Characterization of Spontaneous Stress-Induced In Vitro Apoptosis of Human Acute Myeloid Leukemia Cells; Focus on Patient Heterogeneity and Endoplasmic Reticulum Stress. Hemato 2021, 2, 607-627. https://0-doi-org.brum.beds.ac.uk/10.3390/hemato2030039
Aasebø E, Brenner AK, Hernandez-Valladares M, Birkeland E, Reikvam H, Selheim F, Berven FS, Bruserud Ø. Proteomic Characterization of Spontaneous Stress-Induced In Vitro Apoptosis of Human Acute Myeloid Leukemia Cells; Focus on Patient Heterogeneity and Endoplasmic Reticulum Stress. Hemato. 2021; 2(3):607-627. https://0-doi-org.brum.beds.ac.uk/10.3390/hemato2030039
Chicago/Turabian StyleAasebø, Elise, Annette K. Brenner, Maria Hernandez-Valladares, Even Birkeland, Håkon Reikvam, Frode Selheim, Frode S. Berven, and Øystein Bruserud. 2021. "Proteomic Characterization of Spontaneous Stress-Induced In Vitro Apoptosis of Human Acute Myeloid Leukemia Cells; Focus on Patient Heterogeneity and Endoplasmic Reticulum Stress" Hemato 2, no. 3: 607-627. https://0-doi-org.brum.beds.ac.uk/10.3390/hemato2030039