
Glucocorticoid-Induced Osteocytic Cell Death in a Hypoxic Environment Is Associated with Necroptosis

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Assay
2.3. Immunocytochemistry
2.4. Western Blotting
2.5. Statistical Analysis
3. Results
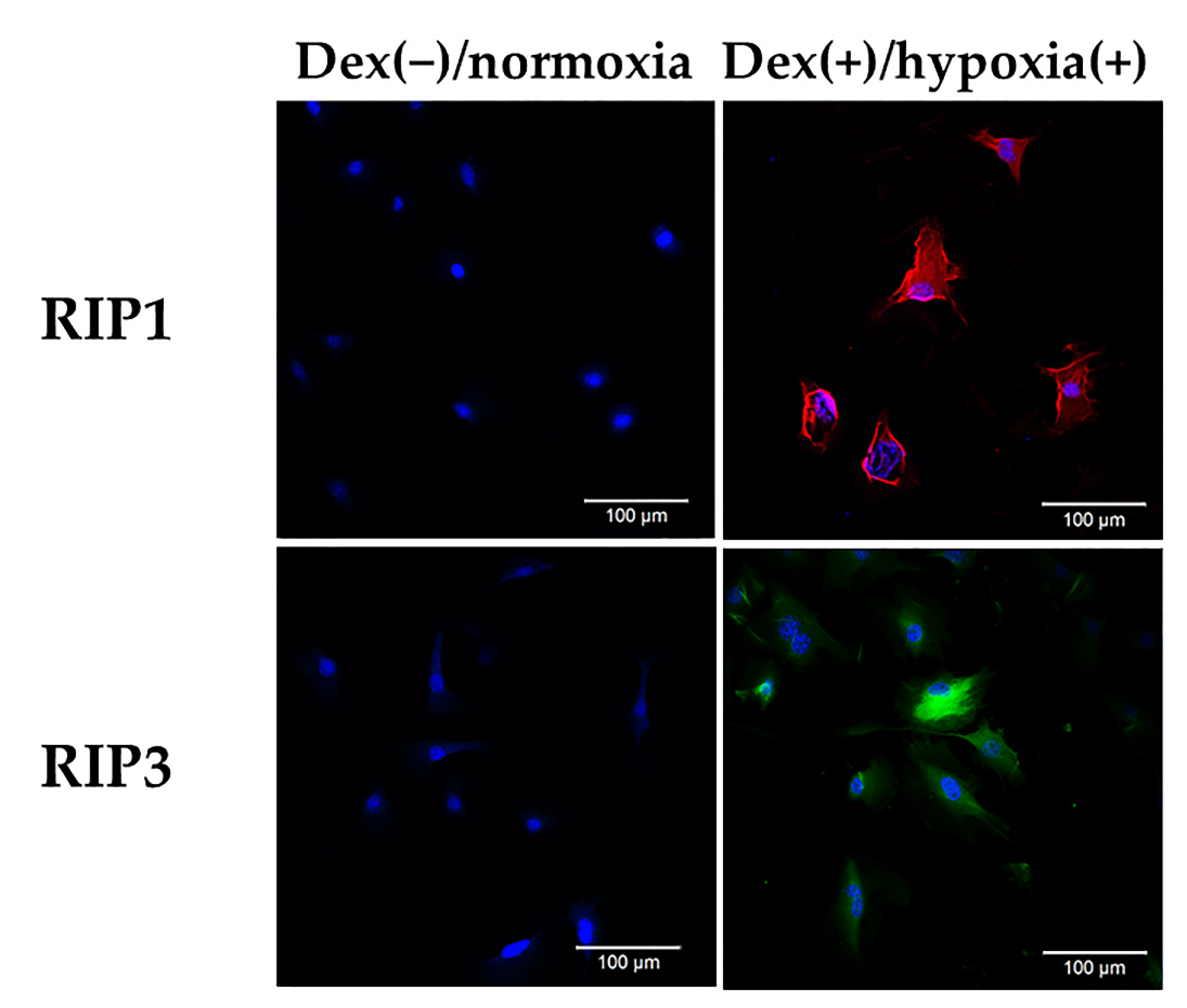
3.1. Expression of RIP1 and RIP3 in Osteocytic Cells under an H-D Stress Environment
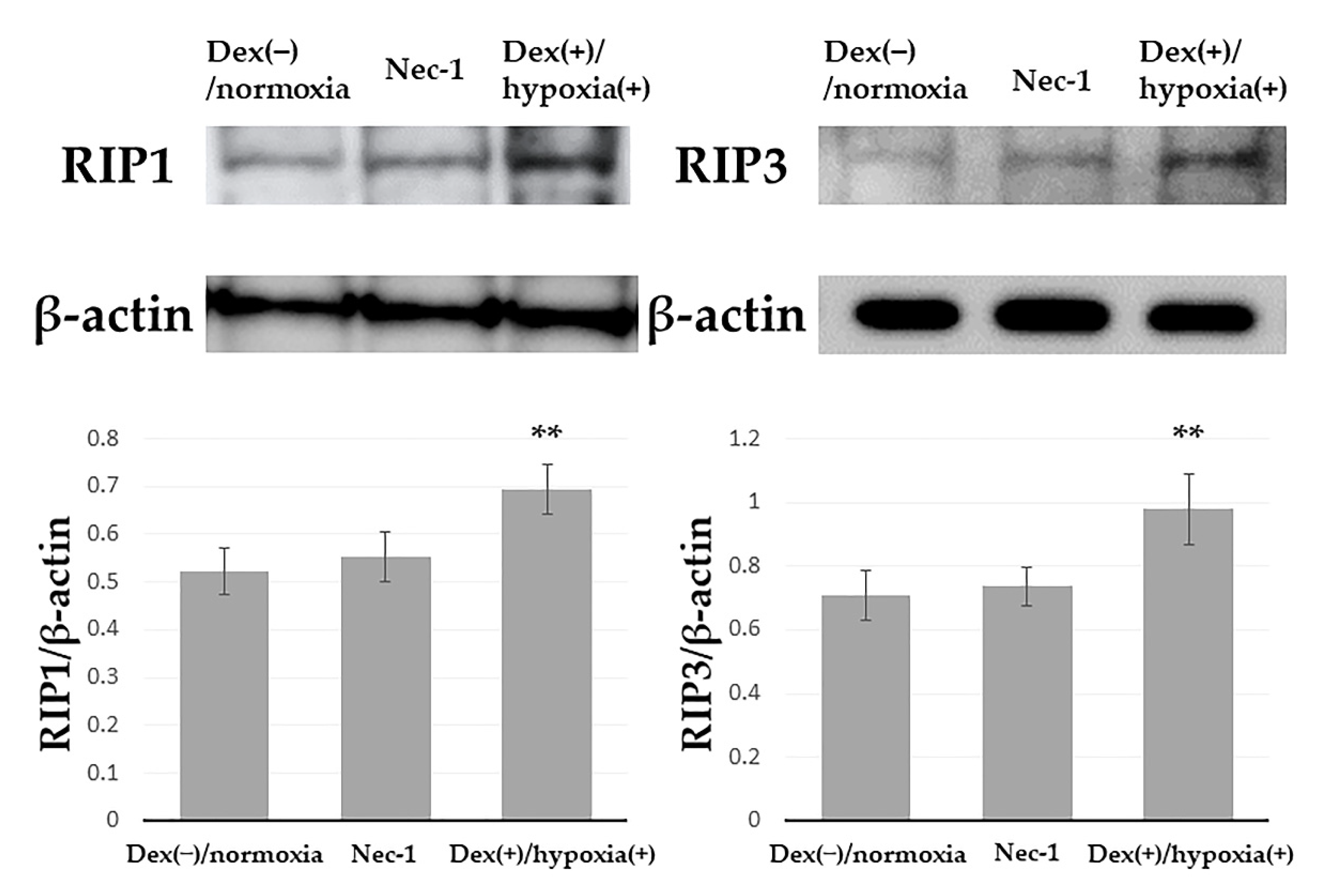
3.2. Inhibition of Expression of RIP1, RIP3 by Necrostation-1
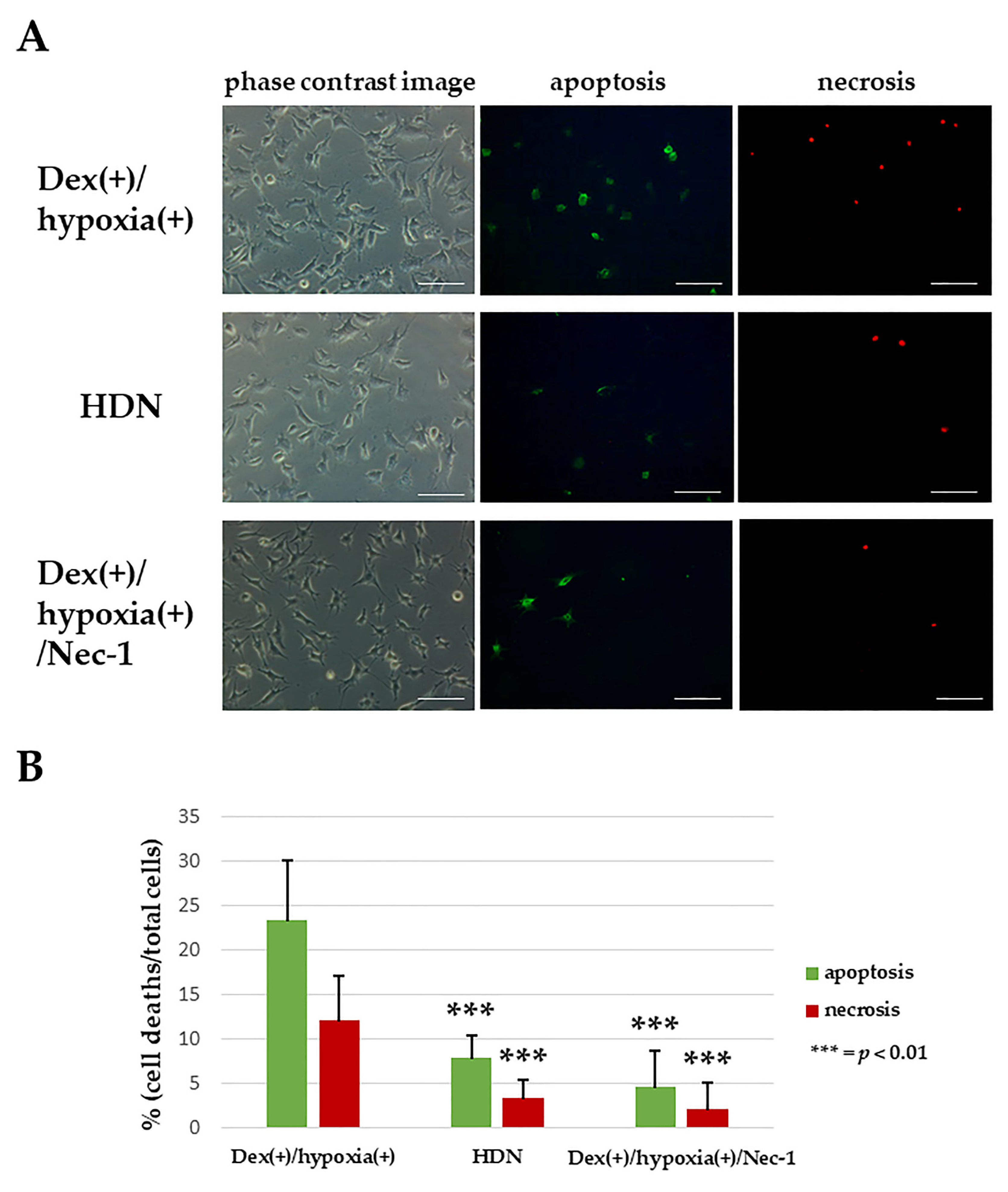
3.3. Inhibition of Osteocytic Cell Death in an H-D Stress Environment by Necrostatin-1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ichiseki, T.; Matsumoto, T.; Nishino, M.; Kaneuji, A.; Katsuda, S. Oxidative stress and vascular permeability in ster-oid-induced osteonecrosis model. J. Orthop. Sci. 2004, 9, 509–515. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Ichiseki, T.; Ueda, S.; Ueda, Y.; Shimazaki, M.; Kaneuji, A.; Kawahara, N. Mitochondrial stress and redox fail-ure in steroid-associated osteonecrosis. Int. J. Med. Sci. 2018, 15, 205–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Ueda, S.; Ichiseki, T.; Shimasaki, M.; Ueda, Y.; Kaneuji, A.; Kawahara, N. Taurine Inhibits Glucocorti-coid-Induced Bone Mitochondrial Injury, Preventing Osteonecrosis in Rabbits and Cultured Osteocytes. Int. J. Mol. Sci. 2020, 21, 6892. [Google Scholar] [CrossRef] [PubMed]
- Laster, S.M.; Wood, J.G.; Gooding, L.R. Tumor necrosis factor can induce both apoptic and necrotic forms of cell lysis. J. Immunol. 1988, 141, 2629–2634. [Google Scholar]
- Lu, J.V.; Chen, H.C.; Walsh, C.M. Necroptotic signaling in adaptive and innate immunity. Semin. Cell Dev. Biol. 2014, 35, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christofferson, D.E.; Yuan, J. Necroptosis as an alternative form of programmed cell death. Curr. Opin. Cell Biol. 2010, 22, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Khoury, M.K.; Gupta, K.; Franco, S.R.; Liu, B. Necroptosis in the Pathophysiology of Disease. Am. J. Pathol. 2019, 190, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Moriwaki, K.; Chan, F.K.-M. RIP3: A molecular switch for necrosis and inflammation. Genes Dev. 2013, 27, 1640–1649. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dugger, D.L.; Wickliffe, K.E.; Kapoor, N.; de Almagro, M.C.; Vucic, D.; Komuves, L.; Ferrando, R.E.; French, D.M.; Webster, J.; et al. Activity of Protein Kinase RIPK3 Determines Whether Cells Die by Necroptosis or Apoptosis. Science 2014, 343, 1357–1360. [Google Scholar] [CrossRef]
- He, S.; Huang, S.; Shen, Z. Biomarkers for the detection of necroptosis. Cell. Mol. Life Sci. 2016, 73, 2177–2181. [Google Scholar] [CrossRef]
- Lin, Q.S.; Chen, P.; Wang, W.X.; Lin, C.C.; Zhou, Y.; Yu, L.H.; Lin, Y.X.; Xu, Y.F.; Kang, D.Z. RIP1/RIP3/MLKL mediates dopa-minergic neuron necroptosis in a mouse model of Parkinson disease. Lab. Investig. 2020, 100, 503–511. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, Y.; Cui, M.; Jin, L.; Wang, Y.; Lv, F.; Liu, Y.; Zheng, W.; Shang, H.; Zhang, J.; et al. CaMKII is a RIP3 substrate mediating ischemia- and oxidative stress-induced myo-cardial necroptosis. Nat. Med. 2016, 22, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Thornton, C.; Leaw, B.; Mallard, C.; Nair, S.; Jinnai, M.; Hagberg, H. Cell Death in the Developing Brain after Hypox-ia-Ischemia. Front Cell Neurosci. 2017, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Ichiseki, T.; Ueda, S.; Ueda, Y.; Tuchiya, M.; Kaneuji, A.; Kawahara, N. Involvement of necroptosis, a newly recognized cell death type, in steroid-induced osteonecrosis in a rabbit model. Int. J. Med. Sci. 2017, 14, 110–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, M.; Ikeda, H.; Ishizu, A.; Miyatake, Y.; Hayase, H.; Yoshiki, T. Altered expression of apoptosis-related genes in osteo-cytes exposed to high-dose steroid hormones and hypoxic stress. Pathobiology 2006, 73, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Ichiseki, T.; Yoshitomi, Y.; Yonekura, H.; Ueda, Y.; Kaneuji, A.; Matsumoto, T. Osteocytic cell necrosis is caused by a combination of glucocorticoid-induced Dickkopf-1 and hypoxia. Med. Mol. Morphol. 2014, 48, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Saito, M.; Goto, T.; Ueshima, K.; Ishida, M.; Hayashi, S.; Ikoma, K.; Mazda, O.; Kubo, T. Heme oxygenase-1 prevents glucocorticoid and hypoxia-induced apoptosis and necrosis of osteocyte-like cells. Med. Mol. Morphol. 2019, 52, 173–180. [Google Scholar] [CrossRef]
- Ueda, S.; Ichiseki, T.; Ueda, Y.; Kaneuji, A.; Kawahara, N.; Matsumoto, T. The timing of glucocorticoid-induced osteocytic cell necrosis under hypoxia. Int. J. Clin. Exp. Pathol. 2016, 9, 7470–7475. [Google Scholar]
- Kato, Y.; Windle, J.; Koop, B.A.; Mundy, G.R.; Bonewald, L.F. Establishment of an Osteocyte-like Cell Line, MLO-Y4. J. Bone Miner. Res. 2010, 12, 2014–2023. [Google Scholar] [CrossRef]
- Rasband, W.S. ImagJ; U. S. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: https://imagej.nih.gov/ij/ (accessed on 24 May 2021).
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Zhe-Wei, S.; Li-Sha, G.; Yue-Chun, L. The Role of Necroptosis in Cardiovascular Disease. Front. Pharmacol. 2018, 9, 721. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Nam, H.S.; Cho, M.K.; Lee, S.H. Arctigenin induces necroptosis through mitochondrial dysfunction with CCN1 up-regulation in prostate cancer cells under lactic acidosis. Mol. Cell Biochem. 2020, 467, 45–56. [Google Scholar] [CrossRef]
- Lyu, A.-R.; Kim, T.H.; Park, S.J.; Shin, S.-A.; Jeong, S.-H.; Yu, Y.; Huh, Y.H.; Je, A.R.; Park, M.J.; Park, Y.-H. Mitochondrial Damage and Necroptosis in Aging Cochlea. Int. J. Mol. Sci. 2020, 21, 2505. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Shimasaki, M.; Ichiseki, T.; Hirata, H.; Kawahara, N.; Ueda, Y. Mitochondrial Transcription Factor A added to Os-teocytes in a Stressed Environment has a Cytoprotective Effect. Int. J. Med. Sci. 2020, 17, 1293–1299. [Google Scholar] [CrossRef]
- Northington, F.J.; Chavez-Valdez, R.; Graham, E.M.; Razdan, S.; Gauda, E.B.; Martin, L.J. Necrostatin Decreases Oxidative Damage, Inflammation, and Injury after Neonatal HI. J. Cereb. Blood. Flow. Metab. 2010, 31, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Valdez, R.; Martin, L.; Flock, D.; Northington, F. Necrostatin-1 attenuates mitochondrial dysfunction in neurons and astrocytes following neonatal hypoxia–ischemia. Neuroscience 2012, 219, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.X.; Li, S.S.; Sun, F.Y. Necrostatin-1 Prevents Necroptosis in Brains after Ischemic Stroke via Inhibition of RIPK1-Mediated RIPK3/MLKL Signaling. Aging Dis. 2019, 10, 807–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmitriev, Y.V.; Minasian, S.M.; Demchenko, E.; Galagudza, M. Study of cardioprotective effects of necroptosis inhibitors on isolated rat heart subjected to global ischemia-reperfusion. Bull. Exp. Biol. Med. 2013, 155, 245–248. [Google Scholar] [CrossRef]
- Szobi, A.; Rajtik, T.; Adameova, A. Effects of Necrostatin-1, an Inhibitor of Necroptosis, and its Inactive Analogue Nec-1i on Basal Cardiovascular Function. Physiol. Res. 2016, 65, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Degterev, A.; Huang, Z.; Boyce, M.; Li, Y.; Jagtap, P.; Mizushima, N.; Cuny, G.D.; Mitchison, T.J.; Moskowitz, M.A.; Yuan, J. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nat. Chem. Biol. 2005, 1, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.; Shen, B.B.; Pu, Y.M.; Zhao, H.W.; Liu, H.; Wu, X.F. TRPM-7 inhibits necroptosis via the PARP-1/RIP-1 signaling pathway and attenuates renal ischemic reperfusion injury in renal tubular epithelial cells. Int. J. Clin. Exp. Pathol. 2017, 10, 88–96. [Google Scholar]
- Cui, H.; Zhu, Y.; Yang, Q.; Zhao, W.; Zhang, S.; Zhou, A.; Jiang, D. Necrostatin-1 treatment inhibits osteocyte necroptosis and trabecular deterioration in ovariectomized rats. Sci. Rep. 2016, 6, 33803. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, S.; Ichiseki, T.; Shimasaki, M.; Hirata, H.; Kawahara, N.; Ueda, Y. Glucocorticoid-Induced Osteocytic Cell Death in a Hypoxic Environment Is Associated with Necroptosis. BioChem 2021, 1, 98-106. https://0-doi-org.brum.beds.ac.uk/10.3390/biochem1020009
Ueda S, Ichiseki T, Shimasaki M, Hirata H, Kawahara N, Ueda Y. Glucocorticoid-Induced Osteocytic Cell Death in a Hypoxic Environment Is Associated with Necroptosis. BioChem. 2021; 1(2):98-106. https://0-doi-org.brum.beds.ac.uk/10.3390/biochem1020009
Chicago/Turabian StyleUeda, Shusuke, Toru Ichiseki, Miyako Shimasaki, Hiroaki Hirata, Norio Kawahara, and Yoshimichi Ueda. 2021. "Glucocorticoid-Induced Osteocytic Cell Death in a Hypoxic Environment Is Associated with Necroptosis" BioChem 1, no. 2: 98-106. https://0-doi-org.brum.beds.ac.uk/10.3390/biochem1020009