
Molecular Detection and Genotyping of Enteric Protists in Asymptomatic Schoolchildren and Their Legal Guardians in Madrid, Spain

 and
and
Abstract
:1. Introduction
2. Results
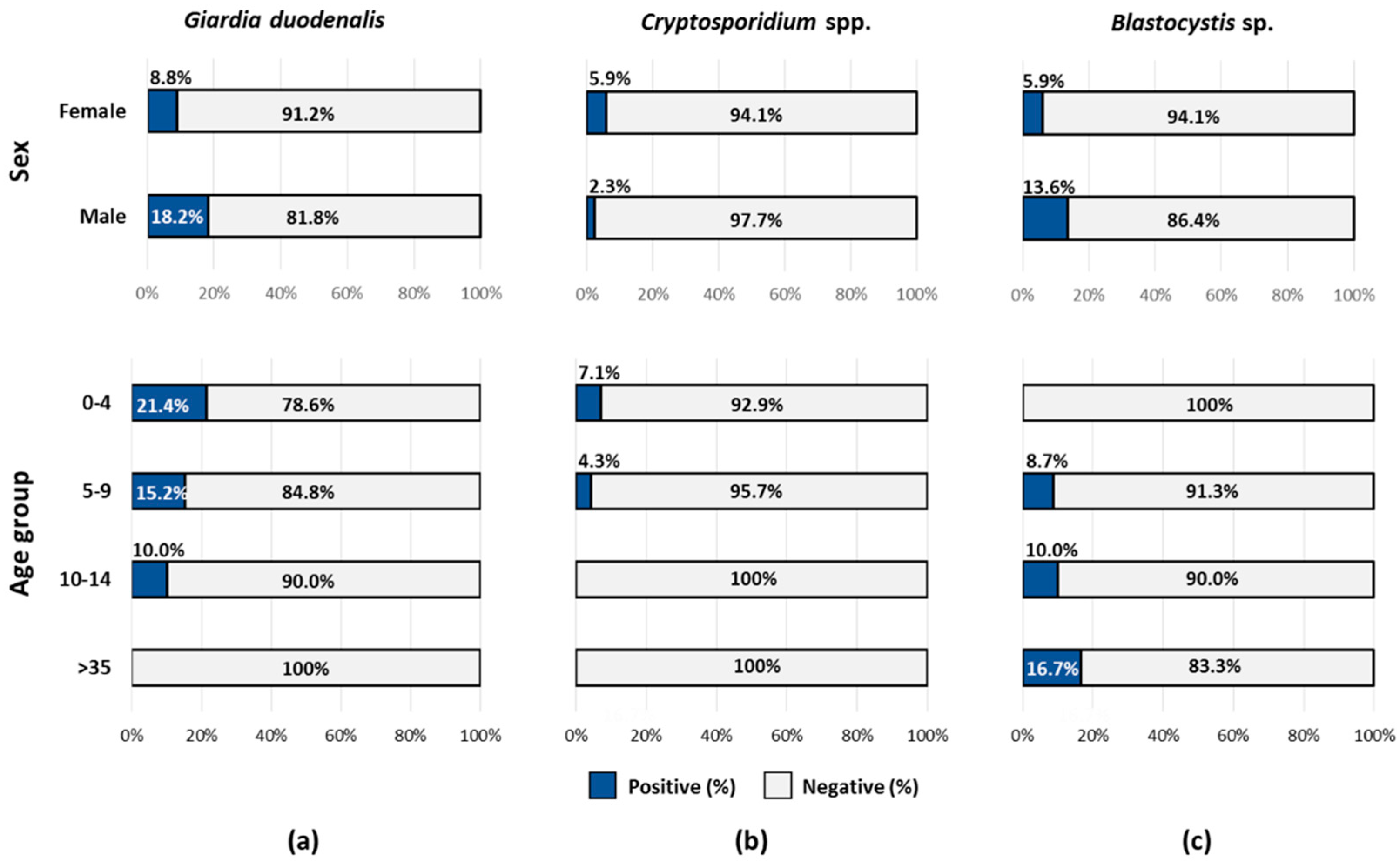
2.1. Occurrence of Protist Enteroparasites
2.2. Molecular Characterization of G. duodenalis Isolates
2.3. Molecular Characterization of Cryptosporidium spp. Isolates
2.4. Molecular Characterization of Blastocystis sp. Isolates
2.5. Risk Association Analysis
3. Discussion
4. Materials and Methods
4.1. Study Area and Stool Sample Collection
4.2. DNA Extraction and Purification
4.3. Molecular Detection and Characterization of Giardia duodenalis
4.4. Molecular Detection and Characterization of Cryptosporidium spp.
4.5. Molecular Detection and Characterization of Blastocystis sp.
4.6. Molecular Detection and Characterization of Enterocytozoon bieneusi
4.7. Sequence Analyses
4.8. Epidemiological Questionnaire
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Carrero, J.C.; Reyes-López, M.; Serrano-Luna, J.; Shibayama, M.; Unzueta, J.; León-Sicairos, N.; de la Garza, M. Intestinal amoebiasis: 160 years of its first detection and still remains as a health problem in developing countries. Int. J. Med. Microbiol. 2020, 310, 151358. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Becnel, J.; Weiss, L.M.; Keeling, P.J.; Didier, E.S.; Williams, B.P.; Bjornson, S.; Kent, M.L.; Freeman, M.A.; Brown, M.J.F.; et al. Microsporidia-emergent pathogens in the global food chain. Trends Parasitol. 2016, 32, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Giangaspero, A.; Gasser, R.B. Human cyclosporiasis. Lancet Infect. Dis. 2019, 19, e226–e236. [Google Scholar] [CrossRef]
- Black, R.E.; Morris, S.S.; Bryce, J. Where and why are 10 million children dying every year? Lancet 2003, 361, 2226–2234. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef]
- de Boer, M.D.; Schuurs, T.A.; Vermeer, M.; Ruijs, G.J.H.M.; van der Zanden, A.G.M.; Weel, J.F.; Bruijnesteijn van Coppenraet, L.E.S. Distribution and relevance of Dientamoeba fragilis and Blastocystis species in gastroenteritis: Results from a case-control study. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 197–203. [Google Scholar] [CrossRef]
- Fletcher, S.M.; Stark, D.; Harkness, J.; Ellis, J. Enteric protozoa in the developed world: A public health perspective. Clin. Microbiol. Rev. 2012, 25, 420–449. [Google Scholar] [CrossRef] [Green Version]
- Efstratiou, A.; Ongerth, J.E.; Karanis, P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks-An update 2011–2016. Water Res. 2017, 114, 14–22. [Google Scholar] [CrossRef]
- Zahedi, A.; Ryan, U. Cryptosporidium-An update with an emphasis on foodborne and waterborne transmission. Res. Vet. Sci. 2020, 132, 500–512. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Macpherson, C.N.L. The socioeconomic burden of parasitic zoonoses: Global trends. Vet. Parasitol. 2011, 182, 79–95. [Google Scholar] [CrossRef]
- Azcona-Gutiérrez, J.M.; de Lucio, A.; Hernández-de-Mingo, M.; García-García, C.; Soria-Blanco, L.M.; Morales, L.; Aguilera, M.; Fuentes, I.; Carmena, D. Molecular diversity and frequency of the diarrheagenic enteric protozoan Giardia duodenalis and Cryptosporidium spp. in a hospital setting in Northern Spain. PLoS ONE 2017, 12, e0178575. [Google Scholar] [CrossRef] [Green Version]
- Hanevik, K.; Wensaas, K.A.; Rortveit, G.; Eide, G.E.; Mørch, K.; Langeland, N. Irritable bowel syndrome and chronic fatigue 6 years after Giardia infection: A controlled prospective cohort study. Clin. Infect. Dis. 2014, 59, 1394–1400. [Google Scholar] [CrossRef]
- Ryan, U.; Cacciò, S.M. Zoonotic potential of Giardia. Int. J. Parasitol. 2013, 43, 943–956. [Google Scholar] [CrossRef]
- Yang, X.; Guo, Y.; Xiao, L.; Feng, Y. Molecular epidemiology of human cryptosporidiosis in low- and middle-income countries. Clin. Microbiol. Rev. 2021, 34, e00087–e00119. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Santin, M. Host specificity of Enterocytozoon bieneusi and public health implications. Trends Parasitol. 2019, 35, 436–451. [Google Scholar] [CrossRef]
- Mateo, M.; Mateo, M.; Montoya, A.; Bailo, B.; Saugar, J.M.; Aguilera, M.; Fuentes, I.; Carmena, D. Detection and molecular characterization of Giardia duodenalis in children attending day care centers in Majadahonda, Madrid, Central Spain. Medicine 2014, 93, e75. [Google Scholar] [CrossRef]
- de Lucio, A.; Martínez-Ruiz, R.; Merino, F.J.; Bailo, B.; Aguilera, M.; Fuentes, I.; Carmena, D. Molecular genotyping of Giardia duodenalis isolates from symptomatic individuals attending two major public hospitals in Madrid, Spain. PLoS ONE 2015, 10, e0143981. [Google Scholar] [CrossRef] [Green Version]
- de Lucio, A.; Merino, F.J.; Martínez-Ruiz, R.; Bailo, B.; Aguilera, M.; Fuentes, I.; Carmena, D. Molecular genotyping and sub-genotyping of Cryptosporidium spp. isolates from symptomatic individuals attending two major public hospitals in Madrid, Spain. Infect. Genet. Evol. 2016, 37, 49–56. [Google Scholar] [CrossRef]
- Reh, L.; Muadica, A.S.; Köster, P.C.; Balasegaram, S.; Verlander, N.Q.; Chércoles, E.R.; Carmena, D. Substantial prevalence of enteroparasites Cryptosporidium spp., Giardia duodenalis and Blastocystis sp. in asymptomatic schoolchildren in Madrid, Spain, November 2017 to June 2018. Eur. Surveill. 2019, 24, 1900241. [Google Scholar] [CrossRef]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernández-de-Mingo, M.; Reh, L.; Balasegaram, S.; Verlander, N.Q.; Ruiz Chércoles, E.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp. in asymptomatic school children in Leganés, Madrid (Spain). Microorganisms 2020, 8, 466. [Google Scholar] [CrossRef] [Green Version]
- Carmena, D.; Cardona, G.A.; Sánchez-Serrano, L.P. Current situation of Giardia infection in Spain: Implications for public health. World J. Clin. Infect. Dis. 2012, 2, 1–12. [Google Scholar] [CrossRef]
- Cardona, G.A.; Carabin, H.; Goñi, P.; Arriola, L.; Robinson, G.; Fernández-Crespo, J.C.; Clavel, A.; Chalmers, R.M.; Carmena, D. Identification and molecular characterization of Cryptosporidium and Giardia in children and cattle populations from the province of Álava, North of Spain. Sci. Total Environ. 2011, 412–413, 101–108. [Google Scholar] [CrossRef]
- de Lucio, A.; Bailo, B.; Aguilera, M.; Cardona, G.A.; Fernández-Crespo, J.C.; Carmena, D. No molecular epidemiological evidence supporting household transmission of zoonotic Giardia duodenalis and Cryptosporidium spp. from pet dogs and cats in the province of Álava, Northern Spain. Acta Trop. 2017, 170, 48–56. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, J.; Canut-Blasco, A.; Martín-Sánchez, A.M. Seasonal prevalences of Cryptosporidium and Giardia infections in children attending day care centres in Salamanca (Spain) studied for a period of 15 months. Eur. J. Epidemiol. 1996, 12, 291–295. [Google Scholar] [CrossRef]
- Segura, R.; Prim, N.; Montemayor, M.; Valls, M.E.; Muñoz, C. Predominant virulent IbA10G2 subtype of Cryptosporidium hominis in human isolates in Barcelona: A five-year study. PLoS ONE 2015, 10, e0121753. [Google Scholar] [CrossRef] [Green Version]
- Abal-Fabeiro, J.L.; Maside, X.; Llovo, J.; Bartolomé, C. Aetiology and epidemiology of human cryptosporidiosis cases in Galicia (NW Spain), 2000–2008. Epidemiol. Infect. 2015, 143, 3022–3035. [Google Scholar] [CrossRef] [Green Version]
- Martín-Sánchez, A.M.; Canut-Blasco, A.; Rodríguez-Hernández, J.; Montes-Martínez, I.; García-Rodríguez, J.A. Epidemiology and clinical significance of Blastocystis hominis in different population groups in Salamanca (Spain). Eur. J. Epidemiol. 1992, 8, 553–559. [Google Scholar]
- González-Moreno, O.; Domingo, L.; Teixidor, J.; Gracenea, M. Prevalence and associated factors of intestinal parasitisation: A cross-sectional study among outpatients with gastrointestinal symptoms in Catalonia, Spain. Parasitol. Res. 2011, 108, 87–93. [Google Scholar] [CrossRef]
- del Águila, C.; Navajas, R.; Gurbindo, D.; Ramos, J.T.; Mellado, M.J.; Fenoy, S.; Muñoz Fernandez, M.A.; Subirats, M.; Ruiz, J.; Pieniazek, N.J. Microsporidiosis in HIV-positive children in Madrid (Spain). J. Eukaryot. Microbiol. 1997, 44, 84S–85S. [Google Scholar] [CrossRef] [PubMed]
- Paulos, S.; Köster, P.C.; de Lucio, A.; Hernández-de-Mingo, M.; Cardona, G.A.; Fernández-Crespo, J.C.; Stensvold, C.R.; Carmena, D. Occurrence and subtype distribution of Blastocystis sp. in humans, dogs and cats sharing household in northern Spain and assessment of zoonotic transmission risk. Zoonoses Public Health 2018, 65, 993–1002. [Google Scholar] [CrossRef]
- Pipatsatitpong, D.; Rangsin, R.; Leelayoova, S.; Naaglor, T.; Mungthin, M. Incidence and risk factors of Blastocystis infection in an orphanage in Bangkok, Thailand. Parasites Vectors 2012, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muadica, A.S.; Balasegaram, S.; Beebeejaun, K.; Köster, P.C.; Bailo, B.; Hernández-de-Mingo, M.; Dashti, A.; Dacal, E.; Saugar, J.M.; Fuentes, I.; et al. Risk associations for intestinal parasites in symptomatic and asymptomatic schoolchildren in central Mozambique. Clin. Microbiol. Infect. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- del Águila, C.; López-Velez, R.; Fenoy, S.; Turrientes, C.; Cobo, J.; Navajas, R.; Visvesvara, G.S.; Croppo, G.P.; Da Silva, A.J.; Pieniazek, N.J. Identification of Enterocytozoon bieneusi spores in respiratory samples from an AIDS patient with a 2-year history of intestinal microsporidiosis. J. Clin. Microbiol. 1997, 35, 1862–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galván, A.L.; Sánchez, A.M.; Valentín, M.A.; Henriques-Gil, N.; Izquierdo, F.; Fenoy, S.; del Aguila, C. First cases of microsporidiosis in transplant recipients in Spain and review of the literature. J. Clin. Microbiol. 2011, 49, 1301–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lores, B.; López-Miragaya, I.; Arias, C.; Fenoy, S.; Torres, J.; del Aguila, C. Intestinal microsporidiosis due to Enterocytozoon bieneusi in elderly human immunodeficiency virus–negative patients from Vigo, Spain. Clin. Infect. Dis. 2002, 34, 918–921. [Google Scholar] [CrossRef] [Green Version]
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2021. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Alfellani, M.A.; Nørskov-Lauritsen, S.; Prip, K.; Victory, E.L.; Maddox, C.; Nielsen, H.V.; Clark, C.G. Subtype distribution of Blastocystis isolates from synanthropic and zoo animals and identification of a new subtype. Int. J. Parasitol. 2009, 39, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; van Rooyen, M.A.; van Lieshout, L.; Polderman, A.M. Real-time PCR for the detection of Giardia lamblia. Mol. Cell. Probes 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Read, C.M.; Monis, P.T.; Thompson, R.C. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic heterogeneity at the beta-giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triose phosphate isomerase gene characterization and potential zoonotic transmission of Giardia duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thailand. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feltus, D.C.; Giddings, C.W.; Schneck, B.L.; Monson, T.; Warshauer, D.; McEvoy, J.M. Evidence supporting zoonotic transmission of Cryptosporidium spp. in Wisconsin. J. Clin. Microbiol. 2006, 44, 4303–4308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA barcoding of Blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef]
- Buckholt, M.A.; Lee, J.H.; Tzipori, S. Prevalence of Enterocytozoon bieneusi in swine: An 18-month survey at a slaughterhouse in Massachusetts. Appl. Environ. Microbiol. 2002, 68, 2595–2599. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Locus | Assemblage | Sub-Assemblage | Isolates | Reference Sequence | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|---|---|
| gdh | B | BIV | 1 | L40508 | 76–491 | None | MW810324 |
| bg | B | ‒ | 1 | AY072727 | 100–593 | G159A, C165T, C309T, C324T, C393T, T471C | MW810325 |
| tpi | B | BIV | 1 | AF069560 | 1–479 | None | MW810326 |
| Subtype | Allele | Isolates | Reference Sequence | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|---|
| ST2 | 12 | 2 | AB070997 | 8–599 | G185A, T267A, C324T, A477G | MW789153 |
| 11+12 | 1 | AB070997 | 19–607 | G185R, T267A, C324T, A477C, T478C | MW789154 | |
| ST3 | 34 | 2 | AB107965 | 13–602 | None | MW789155 |
| ST4 | 42 | 2 | AB071000 | 14–606 | G78A, G221T, A234G | MW789156 |
| ST8 | 95 | 1 | AB107970 | 55–614 | C128A, A177C, G179A, G190C, T228C, A256T | MW789157 |
| Variable | Category | n | Giardia duodenalis | Cryptosporidium spp. | Blastocystis sp. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cases | OR | 95% CI | p | Cases | OR | 95% CI | p | Cases | OR | 95% CI | p | |||
| Gender | Male | 44 | 8 | — | — | 1 | — | — | 6 | — | — | |||
| Female | 34 | 3 | 2.3 | 0.56–9.4 | 0.33 | 2 | 0.37 | 0.0–4.3 | 0.60 | 2 | 2.5 | 0.48–13 | 0.50 | |
| Age group | ≤7 years | 40 | 7 | — | — | 2 | — | — | 1 | — | — | |||
| >7 years | 36 | 4 | 1.7 | 0.45–6.3 | 0.43 | 1 | 1.8 | 0.16–21 | >0.99 | 5 | 0.16 | 0.02–1.4 | 0.10 | |
| Contact with infants | Exposed | 9 | 0 | — | — | 0 | — | — | 1 | — | — | |||
| Unexposed | 61 | 10 | 0.0 | NA | 0.33 | 3 | 0.0 | 0.0–NA | >0.99 | 5 | 1.4 | 0.14–14 | 0.60 | |
| Diarrhoea in family | Exposed | 16 | 2 | — | — | 3 | — | — | 1 | — | — | |||
| Unexposed | 60 | 8 | 0.92 | 0.09–5.4 | >0.99 | 0 | Inf | 1.7–Inf | 0.01 | 6 | 0.60 | 0.07–5.4 | >0.99 | |
| Diarrhoea in schoolmates | Exposed | 14 | 5 | — | — | 0 | — | — | 0 | — | — | |||
| Unexposed | 3 | 0 | Inf | 0.16–Inf | 0.51 | 0 | NA | NA | >0.99 | 1 | 0.0 | 0.0–8.4 | 0.20 | |
| Recent travel abroad | Exposed | 25 | 2 | — | — | 1 | — | — | 1 | — | — | |||
| (EU countries) | Unexposed | 51 | 8 | 0.47 | 0.09–2.4 | 0.50 | 2 | 1.5 | 0.13–18 | >0.99 | 6 | 0.31 | 0.04–2.7 | 0.40 |
| Recent travel abroad | Exposed | 8 | 1 | — | — | 0 | — | — | 1 | — | — | |||
| (non-EU countries) | Unexposed | 68 | 9 | 0.93 | 0.10–8.5 | >0.99 | 3 | 0.0 | NA | >0.99 | 6 | 1.5 | 0.15–14 | 0.60 |
| Contact with dogs | Exposed | 5 | 0 | — | — | 0 | — | — | 0 | — | — | |||
| Unexposed | 72 | 10 | 0.0 | NA | 0.40 | 3 | 0.0 | NA | >0.99 | 8 | 0.0 | 0.0–NA | >0.99 | |
| Contact with cats | Exposed | 8 | 0 | — | — | 0 | — | — | 2 | — | — | |||
| Unexposed | 69 | 10 | 0.0 | NA | 0.59 | 3 | 0.0 | NA | >0.99 | 6 | 3.5 | 0.57–21.3 | 0.20 | |
| Source of drinking water | Tap | 77 | 11 | — | — | 3 | — | — | 8 | |||||
| Bottled | 25 | 2 | 1.9 | 0.40–9.3 | 0.50 | 1 | 0.97 | 0.10–9.8 | >0.99 | 3 | 0.85 | 0.21–3.5 | >0.99 | |
| Springs | 6 | 1 | 0.83 | 0.09–7.8 | >0.99 | 0 | Inf | 0.03–Inf | >0.99 | 2 | 0.23 | 0.04–1.5 | 0.20 | |
| Others | 4 | 1 | 0.50 | 0.05–5.3 | 0.50 | 0 | Inf | 0.02–Inf | >0.99 | 1 | 0.35 | 0.03–3.8 | 0.40 | |
| Swimming in pools | Exposed | 33 | 2 | — | — | 2 | — | — | 2 | — | — | |||
| Unexposed | 41 | 7 | 0.31 | 0.06–1.6 | 0.30 | 1 | 2.6 | 0.22–30 | 0.60 | 6 | 0.38 | 0.07–2.0 | 0.30 | |
| Handwashing | Always | 19 | 2 | — | — | 1 | — | — | 2 | — | — | |||
| Habitually | 49 | 5 | 1.0 | 0.18–5.9 | >0.99 | 2 | 1.3 | 0.11–15 | >0.99 | 6 | 0.84 | 0.15–4.6 | >0.99 | |
| Never | 9 | 3 | 0.24 | 0.03–1.8 | 0.30 | 0 | Inf | 0.01–Inf | >0.99 | 0 | Inf | 0.09–Inf | 0.30 | |
| Vegetable washing | Always | 60 | 9 | — | — | 3 | — | — | 5 | — | — | |||
| Habitually | 17 | 1 | 2.8 | 0.33–24 | 0.44 | 0 | Inf | 0.10–Inf | >0.99 | 3 | 0.42 | 0.09–1.9 | 0.40 | |
| Never | 0 | 0 | 0.0 | NA | >0.99 | 0 | 0.0 | NA | >0.99 | 0 | 0.0 | NA | >0.99 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köster, P.C.; Dashti, A.; Reh, L.; Bailo, B.; Hernández-de-Mingo, M.; Muadica, A.S.; Carmena, D. Molecular Detection and Genotyping of Enteric Protists in Asymptomatic Schoolchildren and Their Legal Guardians in Madrid, Spain. Parasitologia 2021, 1, 83-94. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1020010
Köster PC, Dashti A, Reh L, Bailo B, Hernández-de-Mingo M, Muadica AS, Carmena D. Molecular Detection and Genotyping of Enteric Protists in Asymptomatic Schoolchildren and Their Legal Guardians in Madrid, Spain. Parasitologia. 2021; 1(2):83-94. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1020010
Chicago/Turabian StyleKöster, Pamela Carolina, Alejandro Dashti, Lucia Reh, Begoña Bailo, Marta Hernández-de-Mingo, Aly Salimo Muadica, and David Carmena. 2021. "Molecular Detection and Genotyping of Enteric Protists in Asymptomatic Schoolchildren and Their Legal Guardians in Madrid, Spain" Parasitologia 1, no. 2: 83-94. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1020010