1. Introduction
Urban areas are increasingly becoming recognized as novel ecosystems that have no natural analogue [
1]. Urban green and blue space (UGBS) supports a variety of species, many of which are of conservation concern, with recent research highlighting the potential for UGBS to support biodiversity in light of the increased habitat loss and fragmentation observed in natural environments [
2,
3,
4,
5,
6,
7,
8]. The recent Intergovernmental Science Policy Platform on Biodiversity and Ecosystem Services (IPBES) report [
9] identified that over 1 million species are now threatened with extinction, many within decades. This deterioration is directly linked to human activity, with urban areas ranked as one of the primary drivers of this loss, and the driver with the largest global impact [
9]. Of note within this report is the implementation of nature-based solutions, including increasing ecological connectivity within urban areas [
9].
Due to the patchy nature of UGBS, the availability of corridors can be seen as an essential connective link for many species. For example, Lepczyk et al. [
6] note that networks of urban green spaces provide passage through the urban matrix, and when such habitats are found in high frequency and proximity to one another, they have the potential to lessen the risk of sink habitats and ecological traps in urban areas. Landscape metrics are the predominant method of quantifying the composition of a landscape, allowing for the description of spatial patterns and ecological processes over time and space [
10]. Several landscape metrics have been developed and applied within the discipline [
11,
12,
13,
14,
15,
16], effectively capturing the influence of landscape configuration, including connectivity, on wider ecological processes that supports effective and targeted management strategies.
In urban environments, studies using landscape metrics have identified a low connectivity of green and blue spaces, partially due to the dominance of the built environment in cities [
17]. Such dominance of the built environment has a detrimental effect on biodiversity due to a lack of habitat [
18,
19]. Initiatives to increase UGBS (particularly green space) have increased bird diversity in global cities [
20] with recent research identifying that the amount of forest within an urban ecosystem has the largest independent effect on forest bird diversity [
21]. An increase in area of suitable habitat (i.e., forest) within a largely inhospitable matrix (i.e., built) is in line with key biogeographic patterns such as species-area relationship [
22], but investigations have been less conclusive with regard to the impact that landscape configurations have on bird diversity [
23,
24,
25].
A recent study by Soifer et al. [
25] found that the landscape configuration in the southern USA greatly influenced bird diversity, in part due to the need to consider multiple scales of analysis. The selection of the spatial scale when studying biodiversity and landscape metrics is particularly pertinent [
26], as factors and processes that are found to be important at one scale are not always found to be at other scales, which renders interpretation and prediction difficult [
27]. Several studies have adopted a multiscale approach for studying the ecological processes responsible for spatial patterns of biodiversity [
15,
28,
29,
30,
31]. For example, Croci et al. [
32] explored the role of landscape metrics in quantifying the configuration of urban woodlands in Rennes, France on biodiversity. They explored the habitat surrounding all woodlands along the rural–urban gradient at two spatial scales (100 and 600 m), with birds more sensitive to variations at wider spatial scales.
Another challenge in understanding the role of UGBS in supporting biodiversity is identifying exactly where these land covers are. Many formal sources of land cover are reported at coarse resolutions. For example, CORINE land cover data in Europe have a (relatively) fine resolution through its vector conceptualization, but has a coarse thematic resolution, considering the built environment as all habitats within the urban area and subsequently overlooking unique habitats such as gardens, hedgerows, and ponds. These habitats have been found to be vitally important for urban biodiversity [
33], and when considered together can form corridors and networks through the often-inhospitable matrix. To overcome this within urban ecosystems, remote sensing is the predominant method used for the identification and classification of UGBS [
34,
35,
36,
37,
38,
39,
40,
41] and has been identified as a powerful method for uncovering these ‘invisible’ green spaces in the urban framework. Sentinel-2 is the primary sensor utilized in Europe in recent years due to its wide swatch, frequent revisit time, fine spatial resolution (10 m) and zero cost [
42,
43,
44]. Such products have been found to identify previously unmapped green space when compared to formal government sources [
34].
Subsequently, with a lack of consensus on the importance of the overall landscape configuration on bird biodiversity and the potentially confounding impact of scale on such results, research is still needed to understand the impact of UGBS configuration and spatial scale on biodiversity. Moreover, with the compounding use of coarse representations of UGBS that ignore ‘invisible’ spaces, there persists a need to quantify species–landscape relationships in urban areas using high-resolution products. Subsequently, the aim of this research is to explore the influence of landscape configuration on bird biodiversity patterns. This study will explore three main research questions: (1) What is the extent of UGBS in Cork City, Ireland? (2) What are the important landscape configurations that influence bird biodiversity patterns? Additionally, (3) how does spatial scale impact these species–landscape relationships?
4. Discussion
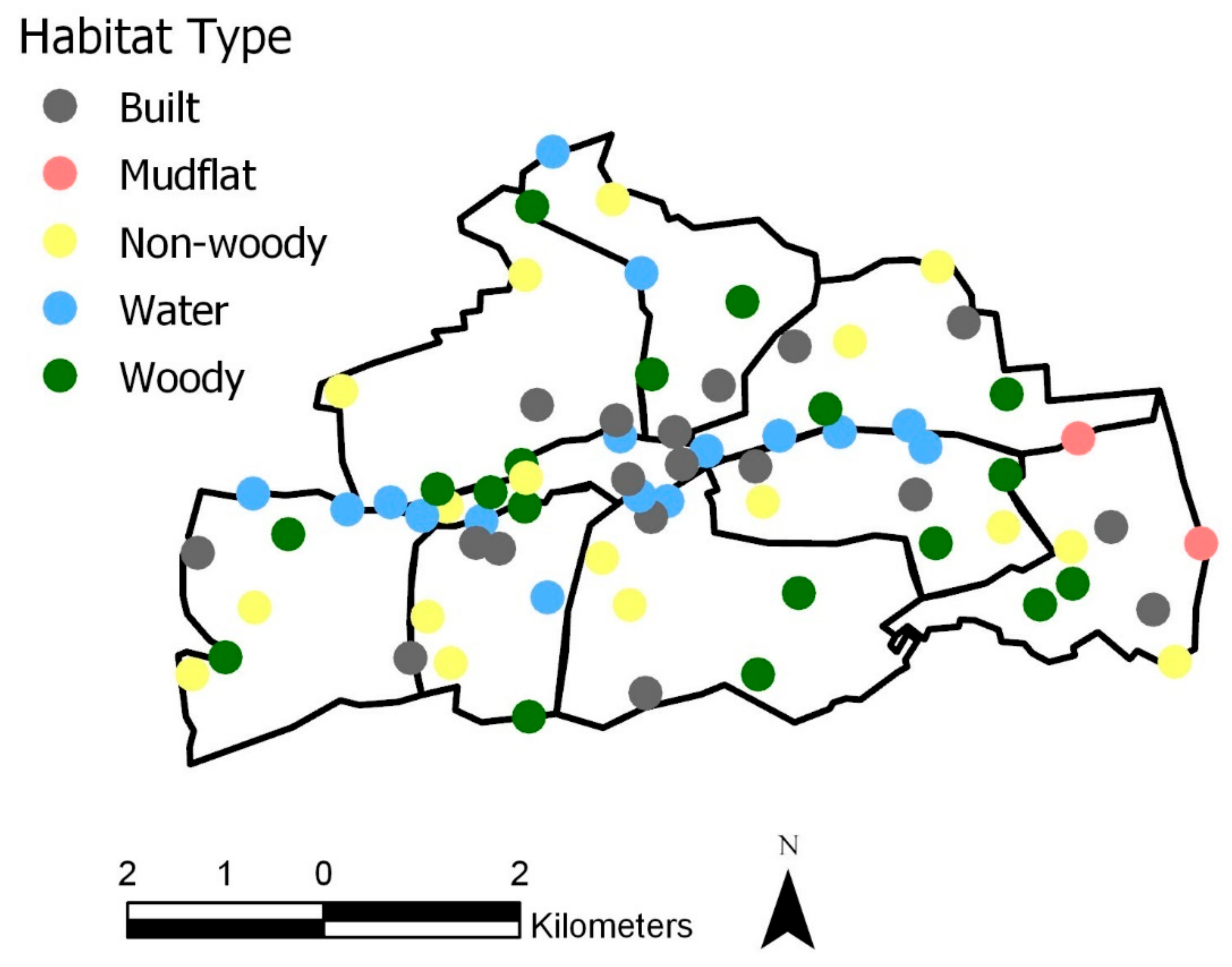
The main aim of this study was to explore the role of UGBS on biodiversity patterns for bird species in Cork City, Ireland. A new representation of UGBS using Sentinel-2 and a supervised land cover classification was created (
Figure 2,
Supplementary Information S3), recording high accuracy for most land cover classes, with the exception of water bodies (
Table 2 and
Table 3). It was identified that the configuration of UGBS plays an important role in determining biodiversity patterns, and moreover these relationships do vary across scale (
Table 5 and
Table 6).
The results of the UGBS land cover classification (
Figure 2,
Supplementary Information S3) highlight the ability of Sentinel-2 to capture the ‘invisible’ green space of Cork City. The invisible green space in this study is primarily urban gardens, but we also noted several brownfield sites in the north of the city. Our results support previous research that similarly identified the capability for satellite imagery to capture these invisible spaces [
34]. This means UGBS classifications can be generated to provide a reliably accurate visualization of the city’s green spaces to support initiatives and policy strategies, such as the previously mentioned Cork City Heritage Plan 2015–2020 and the Draft Cork City Heritage and Biodiversity Plan (2021–2026). The UGBS classification can also aid in identifying corridors in the city, which previous research [
2,
64] has identified as important to support endangered species in urban regions.
Despite the high accuracy when modelling the green spaces, our results suggest that there is a limitation to using Sentinel-2 imagery for capturing water, which contradicts previous research in urban areas [
44]. We posit the reason for such a finding is the scale and configuration of the city. When compared to global cities that have generated classified land cover maps using Sentinel-2, Cork City and the River Lee are relatively small [
65,
66]. Our classification has been impacted where the river width is less than 10 m, or where overhanging tree canopy, floating and riparian vegetation or shadows prevent the full river width from being viewed within a single pixel. The mixed-pixel challenge in the classification process occurs when the spectral signature averaged over a pixel is more representative of the land cover classes with the higher reflectance values [
42], in this case woody and built, compared to the water class which has very low reflectance values at all wavelengths. The spatial resolution also compounds issues in the poor producer accuracy of the water class (
Table 3). When multiple surfaces congregate in a tightly-knit urban environment, the 10 m × 10 m pixel sizes of Sentinel-2 data lead to further mixed-pixel contamination, particularly along river edges. However, the combination of a high producer and user accuracy for the other classes provides reassurance that those classes are correctly classified. The other classes, being more generic in their nature, typically extend over multiple pixels in a non-linear form, and thus while there may be some edge effects of mixed pixel misclassification, the internal pixels for each region can be confidently interpreted. Moreover, by using the landscape configuration metrics implemented in FRAGSTATS and including interactions in the regression analysis, it is possible to see what is immediately surrounding the rivers. Blue space was expected to have a mixed impact on bird diversity, and this is displayed in the positive blue-green and negative blue-built interactions found in the regression models (
Table 6).
We identified mixed relationships with water and biodiversity, with water forming a negative relationship with richness at 100 m and a positive relationship at 200 m when parameterized using class metrics (
Table 5). These results are likely dependent on the blue space type and the habitats that surround it. The edge effect is one such phenomenon that we posit can explain these inverted relationships across scale, as there is a tendency for an increase in species richness and abundance arising from a mixing between two communities [
67]. Richness generally increased when blue spaces were flanked by woody riparian corridors, compared to low levels of richness where the river flowed through the city center itself and was surrounded by the built environment (
Table 7). Future avenues of research could explore the role of mixed pixels and their inference in the edge effect phenomena, developing a spatially explicit accuracy assessment that accounts for location within the landscape configuration. As such, it is important that individual variables are not considered in isolation in any statistical models, but rather the overall landscape configuration is accounted for. However, our regression models suggest that the presence of water surrounded by built environments should not be entirely discounted.
The interaction between water and CONNECT is a possible indication of how blue space can positively affect bird diversity from a connective standpoint, testing the potential for river systems as connective functions for bird diversities in urban areas [
68,
69]. In other studies, species richness was found to decline as rivers entered the urban core [
68], corroborating our results when land cover was considered as a main effect in the statistical models (
Table 5 and
Table 6). However, the positive interaction between water and CONNECT corroborates other studies [
12,
70], where the incorporation of a connectivity metric was found to increase biodiversity. In our study area, the River Lee flows through the city center, where the built environment almost represents an “urban wall” where there is little to no green space, restricting movement of bird diversity (
Figure 2). Due to the Lee splitting in two on the north and south side of the city center, it offers two connective networks for birds to freely move between either side of the city and greener pastures, and as such highlights the important of considering the ecology of the city as a whole (supporting [
25]), as the unique geography can result in novel ecological patterns that may only be captured at multiple spatial scales [
26].
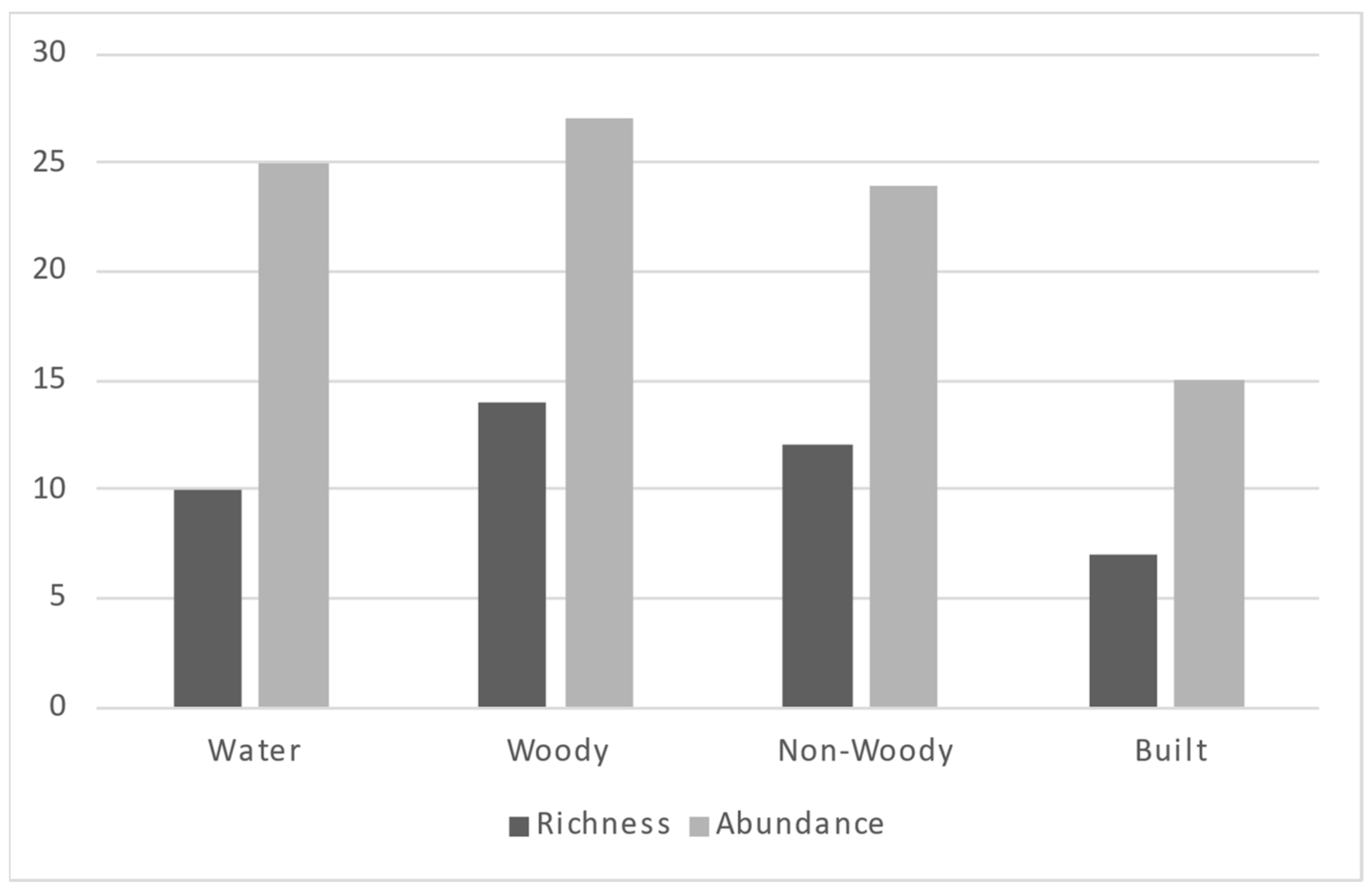
The relationships with green and built spaces were less surprising (
Table 5 and
Table 6). Green space was shown to have a positive relationship with bird diversity, and by contrast, the built class exhibited negative relationships with bird diversity. These relationships were expected as green space was projected to have a positive impact on bird diversity, particularly woody habitat, while built environment was projected to have a negative impact, as has been found in other studies [
2,
35,
71,
72]. For example, our research corroborates the findings of Keten et al. [
71] who developed a riparian quality index to quantify the impact on such habitats on avian biodiversity, identifying that as the proportion of urban cover decreases in riparian habitat and is replaced with more natural green coverage, biodiversity increases. However, our results suggest that the built class should not be totally discounted as it can aid bird diversity in the form of providing suitable nesting habitats and alternative food sources [
73,
74,
75]. This is particularly pertinent for Cork City, where a large amount of the green space returned in the UGBS classification was private gardens. The combination of built and ‘invisible’ green space within the wider landscape of Cork is clear in supporting an increased biodiversity. Moreover, the observation of a whitethroat at a brownfield site associated with scrubland habitat further demonstrates the role of invisible green spaces to biodiversity.
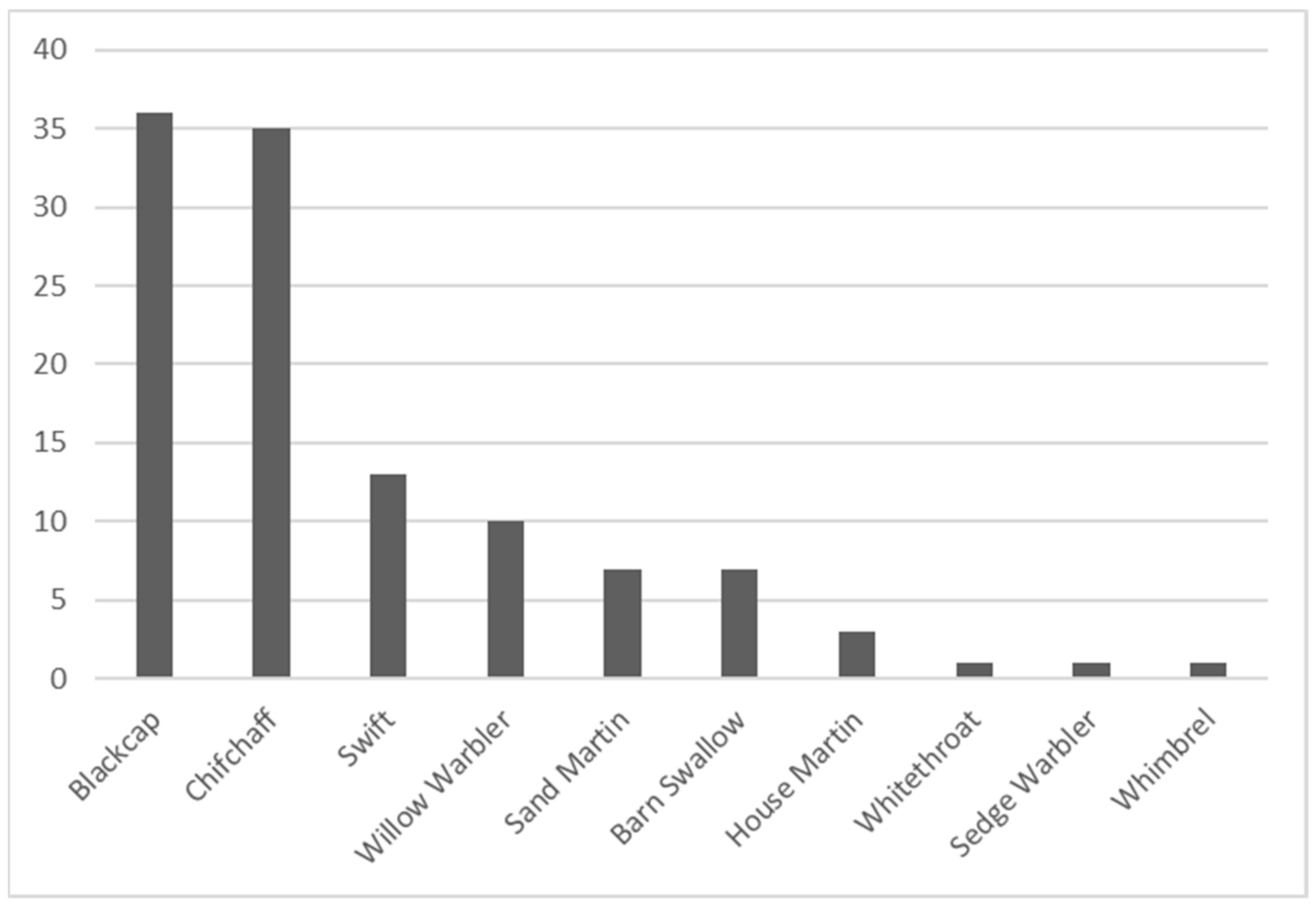
One of the main results emanating from these field surveys is that 38% of the species recorded in Cork City are species of conservation concern in Ireland [
76]. Of note were swifts, grey wagtails (
Motacliia cinerea), meadow pipits (
Anthus pratensis) and black-tailed godwits (wintering) on the Red list, while there were 20 species on the Amber list, including barn swallows, sand martins, willow warblers, European starlings (
Sturnus vulgaris), and herring gulls (
Larus argentatus) (wintering). This information demonstrates how important urban areas such as Cork City can be for species monitoring and management. For example, urban landscapes have become important for many threatened gull species that use buildings for nesting sites [
74]; however, building renovations have resulted in a loss of breeding habitat for many other endangered species such as swifts [
63]. Urban nest-boxes have been reported as a compensatory measure [
77], yet in Cork City the high prevalence of swifts (
Figure 4) may be due to the many vacant and derelict buildings [
78], although further research at a city scale is needed to quantify how many swifts are supported within the city and specifically where they are nesting. As such, it is important to consider species-specific responses to landscape configurations in urban areas, as important relationships may be overlooked when species are aggregated into an overall biodiversity metric.
It is important to note here that we present our results with some caveats, including the removal of three field surveys in water locations and the potential for more validation points to provide confidence intervals to land cover classifications. Moreover, the issue of detectability in bird surveys is well recognized [
55,
79,
80,
81,
82], with various considerations required when implementing surveys. Incomplete detectability can be considered less of a concern if a rigorous sampling design is implemented, which reduces the variation in detection probabilities to less than any variation in the population size [
83]. We implemented several of the protocols outlined by Hedblom and Soderström [
83], such as a consistent calendar period, surveying during peak vocal activity, avoiding adverse weather conditions, only counting birds within our radius, discounting birds that flew high above the site, and waiting a short period upon arrival to the site to begin surveying. The exception to this was that we could not conduct repeat surveys due to time and resource constraints affiliated with this study, although this is not out of line with other studies that have found the efficacy of one count studies to be sufficient [
84,
85]. However, it should be noted that there will always be uncertainty surrounding detectability even when the number of surveys increases [
86].
We decided not to deconstruct our biodiversity metrics into species groups based on detectability or traits as has been suggested for birds and other taxa [
79,
87]. Instead, to explore any variation and uncertainty associated with slight differences in surveys, we undertook further analysis and grouped the results by zone, taking the average of point counts and landscape variables within a zone, following [
7]. For example, the richness values of 16 at site 2 and 10 at site 6 (both woody) were averaged to become 13. We completed this for all classes within each zone, averaging both the response variables and the landscape metrics. We then re-ran our statistical models (
Supplementary Information S7). Across both class and landscape metrics, we observed slight differences in the variables returned by the stepAIC process as would be expected when using an information criterion. However, we only reported seven instances where the coefficient inverted, out of all possible combinations of variables (including two-way interactions), which is 36 for the landscape metric model and 10 for the class metric model. Of these seven inversions, only two were significant. The main difference between the original model (
Table 6) and the aggregated model (
Supplementary Information S7) is that for abundance, water now returns a positive relationship at all scales as opposed to a negative one when considered as a main effect. The aggregation of sites as opposed to repeat counts may be one such artefact for returning different relationships, as we are combining results from different water environments as opposed to repeating counts at the exact same location. However, such findings do not contradict the main conclusions drawn from our results, as our original interpretation that land cover classes should not be considered in isolation from the surrounding landscape configuration remain due to positive water and connectivity interactions. As the results of this aggregation largely concur with our original models, and due to the fact that such an aggregation reduces our
n substantially (
n = 35), as well as the uncertainty from treating different locations as repeat surveys, we prefer to report the results of our original model, with the caveat that such abundance metrics should be treated with caution due to lack of repeat surveys.
Another perspective that warrants future research is to classify different tree cover types such as coniferous and deciduous trees, or applying a land use classification to different green spaces such as parks, cemeteries, and gardens. The coniferous and deciduous types of wooded areas would make a difference when it comes to bird diversity [
79]. This is because certain species or habitat specialists prefer coniferous woodland, such as coal tits (
Periparus ater), crossbills (
Loxia curvirosta) and goldcrests (
Regulus regulus) compared to deciduous woodland which would have a larger assortment of species [
88]. A classification of the various green space types in Cork City coupled with survey results for each green space type would also provide key information on bird diversity that could help the necessary stakeholders develop and distribute information on biodiversity and heritage for the Draft Cork City Heritage and Biodiversity Plan 2021–2026. It was not possible to discriminate between coniferous and deciduous woodland, however, as the spatial resolution was not adequate or fine enough to allow for such identifications. The lack of cloud free imagery for the winter months also played a part in this as it may have been possible to notice the lack of leaves on deciduous, compared to coniferous trees. Satellite imagery with finer resolution could possibly be more effective in classifying the vegetation types such as deciduous and coniferous vegetation and produce a more accurate result for water [
41,
89,
90].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}