1. Introduction
All living forms are compartmentalized. Membranes surround the cells of all organisms and intracellular organelles, which act as a barrier to control the transfer of substances between the inside and outside of these structures. The lipophilic nature of the membranes prevents the passage of charged particles, such as ions. Therefore, the transport of ions must be regulated by specific proteins that are an integral part of the membranes, acting as channels allowing the entry and exit of ions [
1]. These proteins maintain asymmetric concentrations of ions and facilitate their selective transport, which leads to the formation of energy that is eventually stored in the form of ATP [
2]. Therefore, the control of ion transport and the homeostatic regulation of ion concentration levels across the membrane plays a critical role in sustaining life.
Zinc plays an important role in protein function, being a structural element of many macromolecules. Its importance has been recognized since 1869 when it was found to be necessary for the growth of
Aspergillus niger [
3] and later was found that it also affected the growth of plants and animals. In humans, Ananda S Prasad first suspected zinc deficiency in 1958 after he, at the request of James A. Halsted, evaluated a patient with iron deficiency. In addition In addition to iron deficiency, the patient appeared ~10 years older and was severely stunted and prepubertal age [
4].
As a micronutrient, the importance of zinc has been reflected through the numerous diseases that are associated with its dietary deficiency. For example, zinc deficiency led to growth disorder, cognitive disorder and compromised immune function. Zinc deficiency occurs because of nutritional factors, but also in various disease states (malabsorption, Crohn’s disease, alcoholism, liver cirrhosis, chronic renal disease and other chronically debilitating diseases) [
5,
6,
7]. Indeed, zinc supplementation improved these syndromes, as well as testicular and ovarian dysfunction or growth retardation [
8].
Labile zinc is a small fraction of total intracellular zinc that is loosely linked to proteins and easily interchangeable. It is the pool sensitive to chelation and capable of detection by a variety of chemical and genetic sensors, participates in signal transduction pathways. Both free and labile zinc function as a second messenger molecule, modulating the activity of several enzymes and, as a result, signaling and metabolic pathways, as well as cellular activities. When cellular zinc levels are elevated, zinc transporter proteins (ZnT) contribute to modulating the cytoplasmic zinc balance by exporting zinc out to extracellular space or by sequestrating cytoplasmic zinc into intracellular compartments. On the contrary, when cellular zinc is depleted, zinc transporter (Zrt) and Irt-like proteins, known as ZIP proteins, work to increase cytoplasmic zinc concentrations [
9,
10,
11].
At the cellular level, several natural molecules can bind labile zinc and promote its passage across lipophilic membranes. Such molecules are known as ionophores, from the ancient Greek, meaning “ion carriers”. Ionophores are membrane-active compounds that do not depend on active metabolism to exert their biological activity. They are chemical entities that bind reversibly with the ion, forming a supramolecular complex that can diffuse across hydrophobic membranes [
12]. This capacity of ionophores to alter the normal ion balance of membranes can be used as a mechanism for inducing cell death, or to eliminate harmful microorganisms or cells with abnormal growth, like tumor cells [
13].
Zinc chelators are able to transport zinc cations across the plasma membrane, independently of zinc transporters [
14]. In chemistry, zinc is available in the form of inorganic and organic salts. Some inorganic zinc salts are zinc sulfate, zinc chloride, zinc oxide and zinc carbonate, while organic salts include zinc citrate, zinc lactate, zinc acetate, zinc gluconate, zinc stearate, zinc picolinate and zinc ascorbate [
15,
16]. The ion selectivity of ionophores is a combined function of the energy required for desolvation of the ion and the liganding energy obtained on complexation. For metal binding, thermodynamic stability of complexes is the measure of a metal ion’s tendency to selectively form a certain metal complex and is directly connected to metal-ligand bond energies, which would result in a decrease in the system’s free energy [
17,
18]. Several molecular characteristics can be correctly represented as a sum of individual bond properties, without the need to explicitly solve any wave equations. The molecular mechanics method refers to the study of such molecular characteristics in terms of non-quantum mechanical models (MM). In a nutshell, MM posits that the energy of a molecular system is made up of five additive, non-interacting components [
19,
20].
Numerous investigations have demonstrated that flavonoids influence the metabolism of zinc [
21]. For example, rats fed with baicalin and rutin for long periods of time had lower liver levels of total zinc, iron and copper, suggesting that certain flavonoids, contain metal-binding motifs, like phytate, may retain these metals and render them inaccessible for absorption. On the other hand, polyphenols can transport zinc across the plasma membrane without relying on cell transport systems like zinc transporters or endocytosis. Dabbagh-Bazarbachi et al. investigated the interactions between a variety of polyphenols and polyphenol metabolites and zinc in mouse hepatocarcinoma Hepa 1–6 cells under physiological conditions [
22]. After the addition of quercetin and epigallocatechin-gallate, the zinc-polyphenol chelation complex was formed and transported across the lipid bilayer. When given to cells, these complexes rapidly increase the intracellular zinc pool, which may be detected using various fluorophores such as Zinquin or FluoZin-3 [
23]. They concluded that flavonoids could generate water-insoluble membrane-permeant zinc complexes that penetrate the plasma membrane and serve as zinc ionophores.
In order to ascertain whether other natural compounds, such as flavonoids, glutathione and plant amino acids, can serve as new zinc ionophores, we here provide a computational energetic study of the forming zinc complexes. The study is based on obtaining a molecular conformational structure of the zinc complexes with the lowest possible energy content. We also discuss the relevance and biological implications of the formation of such complexes. We hope that this preliminary study could offer new opportunities to prevent zinc deficiency by using nutraceutics, such as amino acids, glutathione or flavonoids, together with zinc supplementation.
2. Results
For initial considerations and validation of our study, we computed the energy and favorable spatial orientation of some well-known zinc ionophores when complexed with zinc cations. The differences in their energetic level, derived from molecular mechanics, is summarized in
Table 1. For each complex, we studied different three-dimensional structural conformations, considering those with the lowest possible energy and favorable geometry respect to the internal position of the zinc cation.
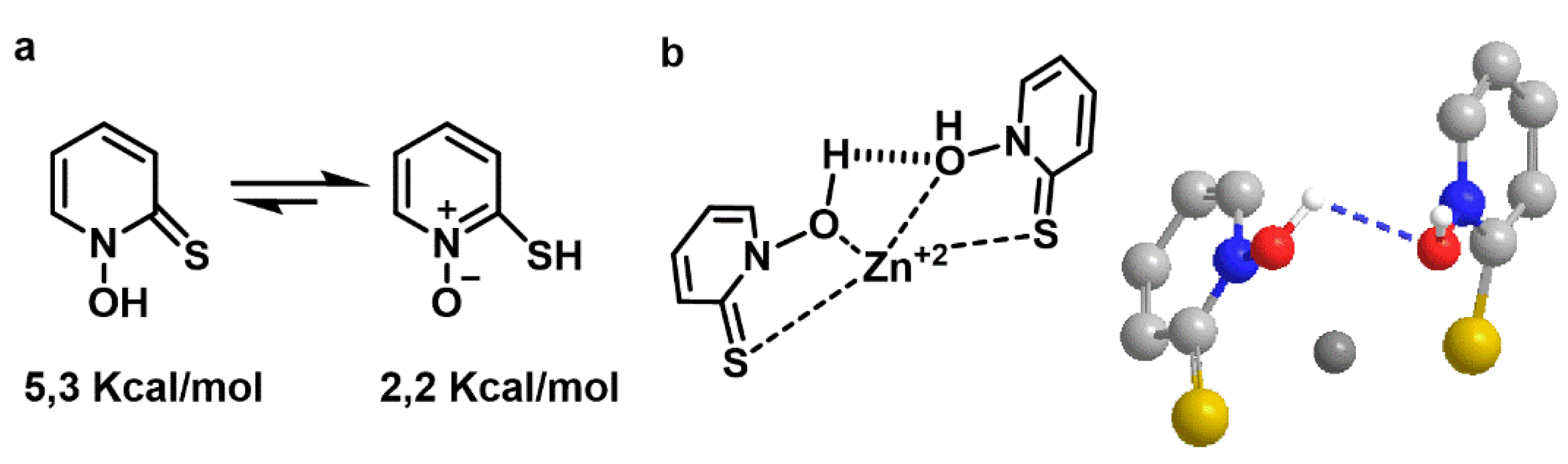
Free pyrithione has two structures with different energies. The left one has an energy of E = 5.3 kcal/mol and the right one is E = 2.2 kcal/mol (the lower the energy, the higher the stability, so the balance is shifted to the right). The zinc-pyrithione complex has an energy of E = −37.2 kcal/mol, which is considerably more stable than free pyrithione (
Figure 1). The favorable geometry of the complex and spatial orientation of the molecules, showed that the zinc cation was in an internal position between two adjacent pyrithione molecules, with the polar groups orienting towards the inside.
The efficient shielding of its polar core, which delocalizes the cation charge, may explain the resultant complex’s lipid solubility, consistent with the role of pyrithione as a zinc ionophore (
Figure 1).
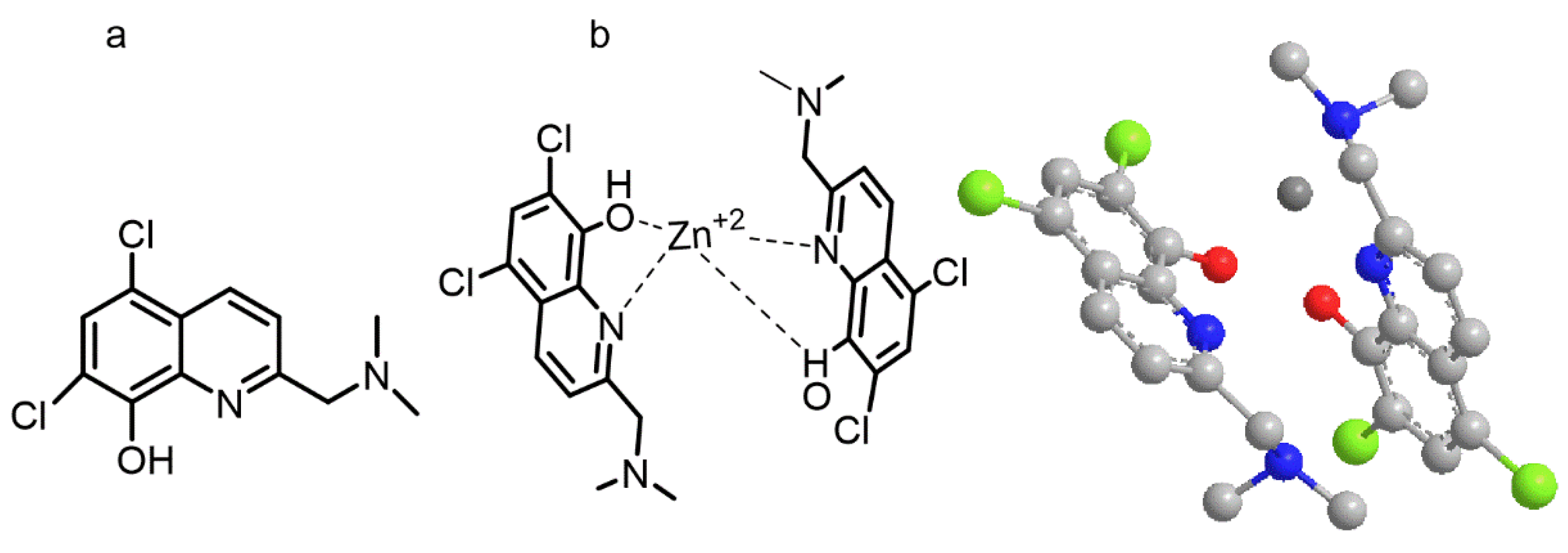
Similarly, the zinc ionophore adopted a disposition around the zinc cation with two molecules of 5,7-Dichloro-2-((dimethylamino) methyl) quinolin-8-ol (PBT2) in opposite orientations (
Figure 2). The computed energy for the free form is E = 11.7 kcal/mol whereas for the zinc/PBT2 complex depicted in
Figure 2b was E = −110.5 kcal/mol.
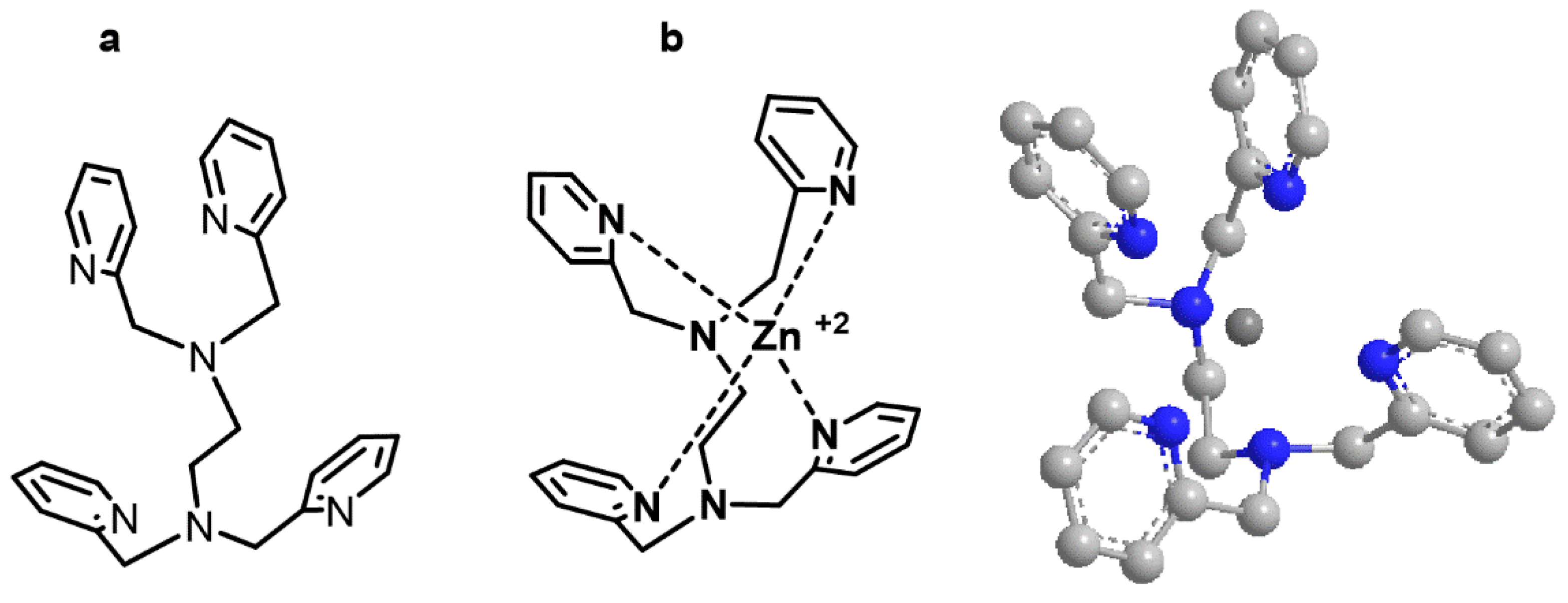
TPEN (N,N,N′,N′-tetrakis(2-pyridinylmethyl)-1,2-ethanediamine), a zinc ionophore without oxygen atoms, has an E = 38.9 kcal/mol for the free form and of E = −126.6 kcal/mol for the complex zinc/TPEN. The spatial orientation of the most favorable complex also showed an internal zinc cation surrounded by four pyridine rings (
Figure 3).
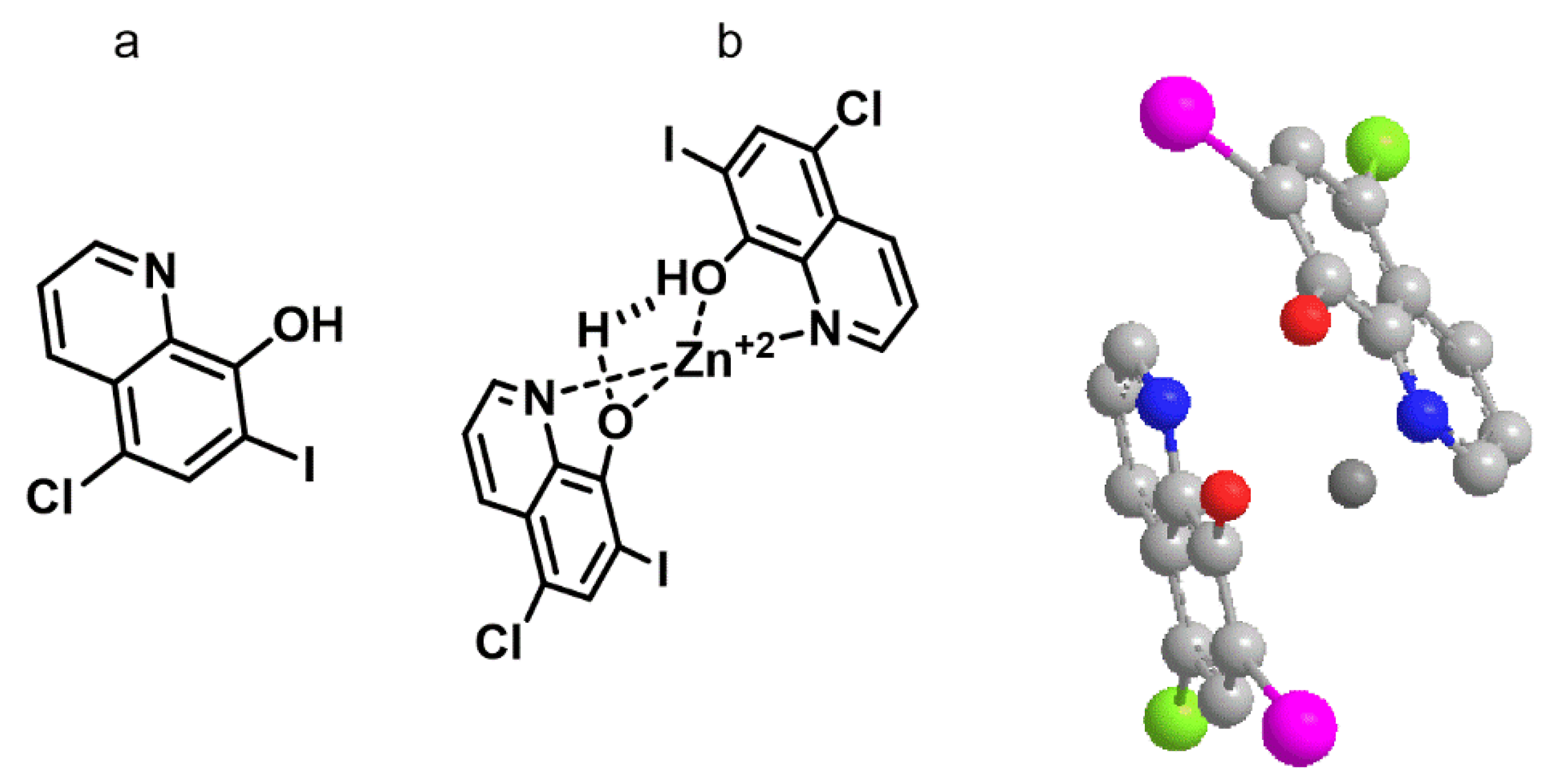
Clioquinol (5-Chloro-7-iodo-8-quinolinol) had an energy of E = 6.9 kcal/mol, whereas for the complex with zinc the energy was of E = −78.4 kcal/mol, indicating the favorable formation of the zinc. The geometry and spatial orientation of the complex showed the internal zinc cation between two adjacent hydroxyquinoline groups in opposite directions and the zinc cation coordinated with oxygen and nitrogen atoms (
Figure 4).
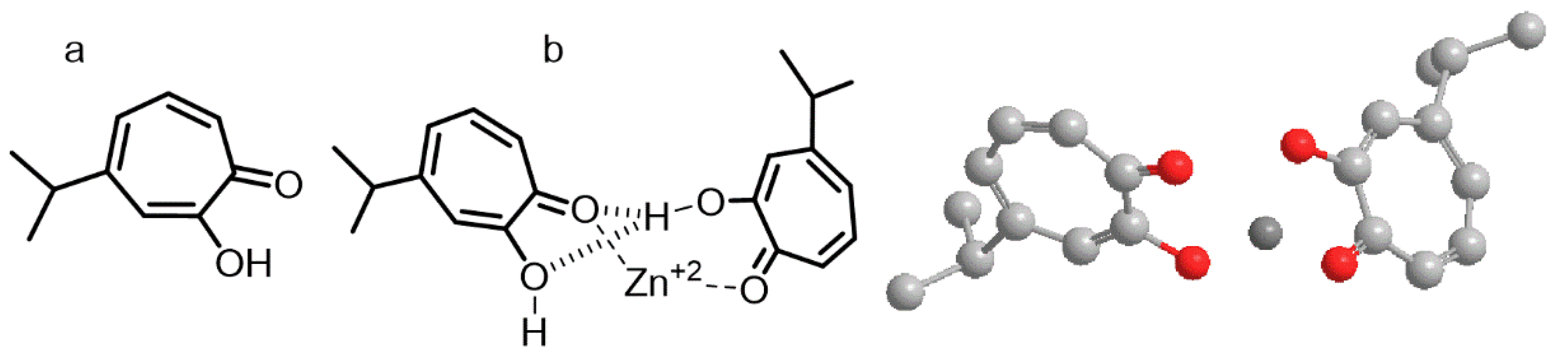
Hinokitiol (2-hydroxycyclohepta-2,4,6-trien-1-one) is structurally related to tropolone (a well-known chelating agents). In our computational analysis, free hinokitiol had an energy of E = 18.1 kcal/mol, the dimer form had an energy of E = 29.5 kcal/mol, whereas the complex hinokitiol/zinc had an energy content of E = −125.6 kcal/mol. The spatial orientation for the lowest energetic form of hinokitiol/zinc showed an internal zinc cation coordinated with oxygen atoms from the hydroxyl and carbonyl groups of the aromatic rings that were orienting their hydrophobic isobutyl groups at opposite ends (
Figure 5).
The greatest difference in energy was found for the TPEN molecules, in free form was of 38.9 kcal/mol and complexed with zinc was of E = −126.6 kcal/mol. The lowest shift in the computed molecular mechanical energy was found for pyrithione, which in free form was of E = 2.2 kcal/mol and and in combination with zinc cation, was of E = −37.2 kcal/mol (
Table 1).
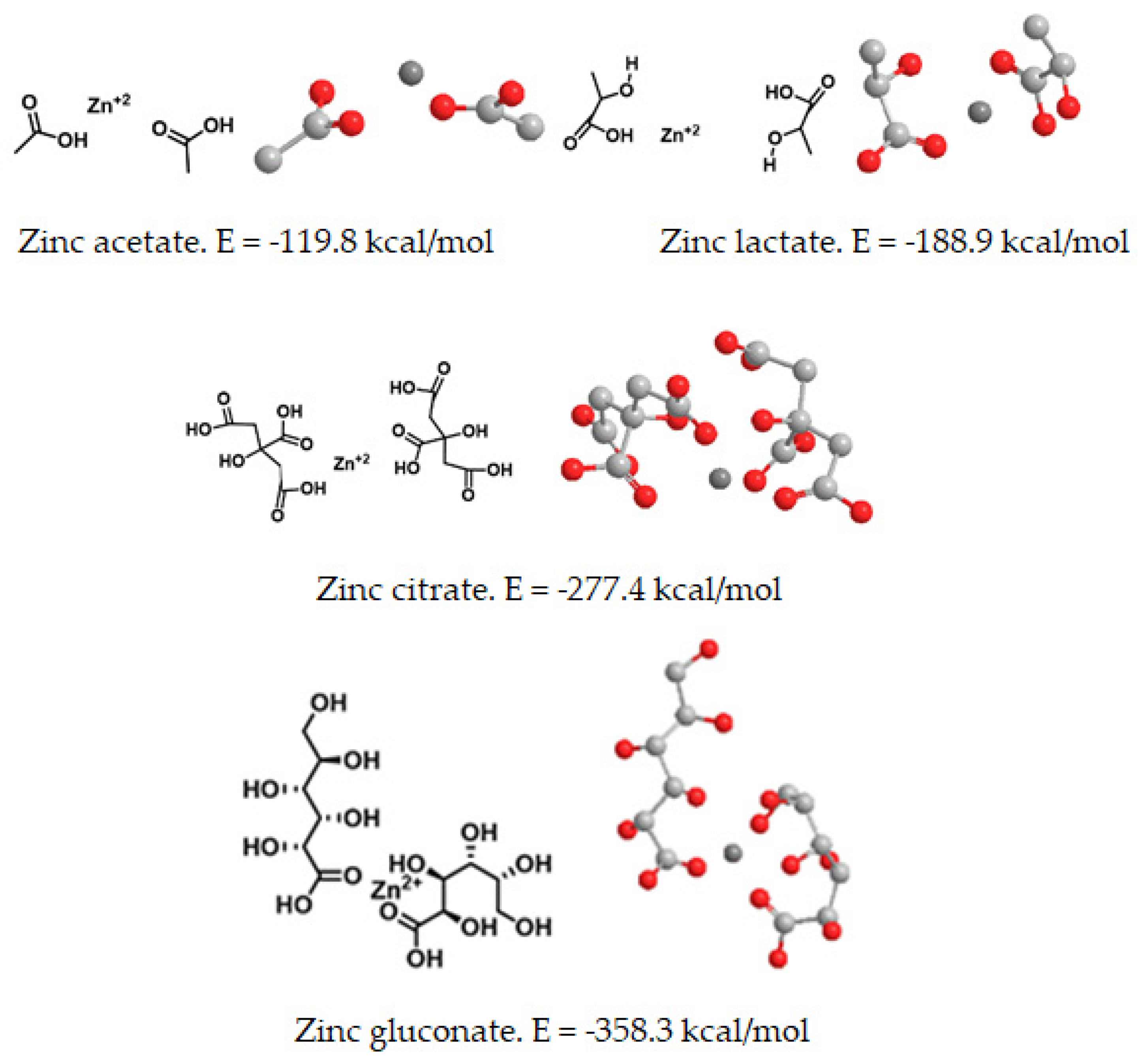
Next, an energetic study was carried out to analyze whether there may be energetically optimized ionophores that are able to chelate zinc, independently of their previous commercial forms (acetate, citrate, gluconate and zinc lactate), which are already on the market. In free forms, the lowest energy (E = −358.3 kcal/mol) was found for zinc gluconate, whereas the highest energy for a free form of zinc salts of organic acids (E = −119.8 kcal/mol) corresponded to zinc acetate. In all organic forms studied, the three-dimensional analysis of the complexes showed an internal zinc cation surrounded by two organic molecules in coordination with oxygen atoms from hydroxyl and carbonyl groups of the organic molecules. (
Figure 6).
When combined with an amino acid (tryptophan) or a polyphenol (silybin), the corresponding complexes with zinc showed different energy. The lowest energy was found for the zinc gluconate, whereas the highest energy obtained corresponded to the amino acid and flavonoid complexes with zinc citrate (
Table 1,
Figure 6). Similar values were obtained for amino acid and flavonoid complexes formed with zinc acetate and zinc lactate (
Table 1). Therefore, we decided to use zinc gluconate for further theoretical studies, as it was the most stable form of dietary zinc salts of organic acids, according to our molecular mechanics calculations (
Figure 6). The complexes with zinc gluconate showed the lowest energy values, so the formation of these complexes with zinc gluconate were more stable than those formed with other forms of zinc salts of organic acids (
Table 2).
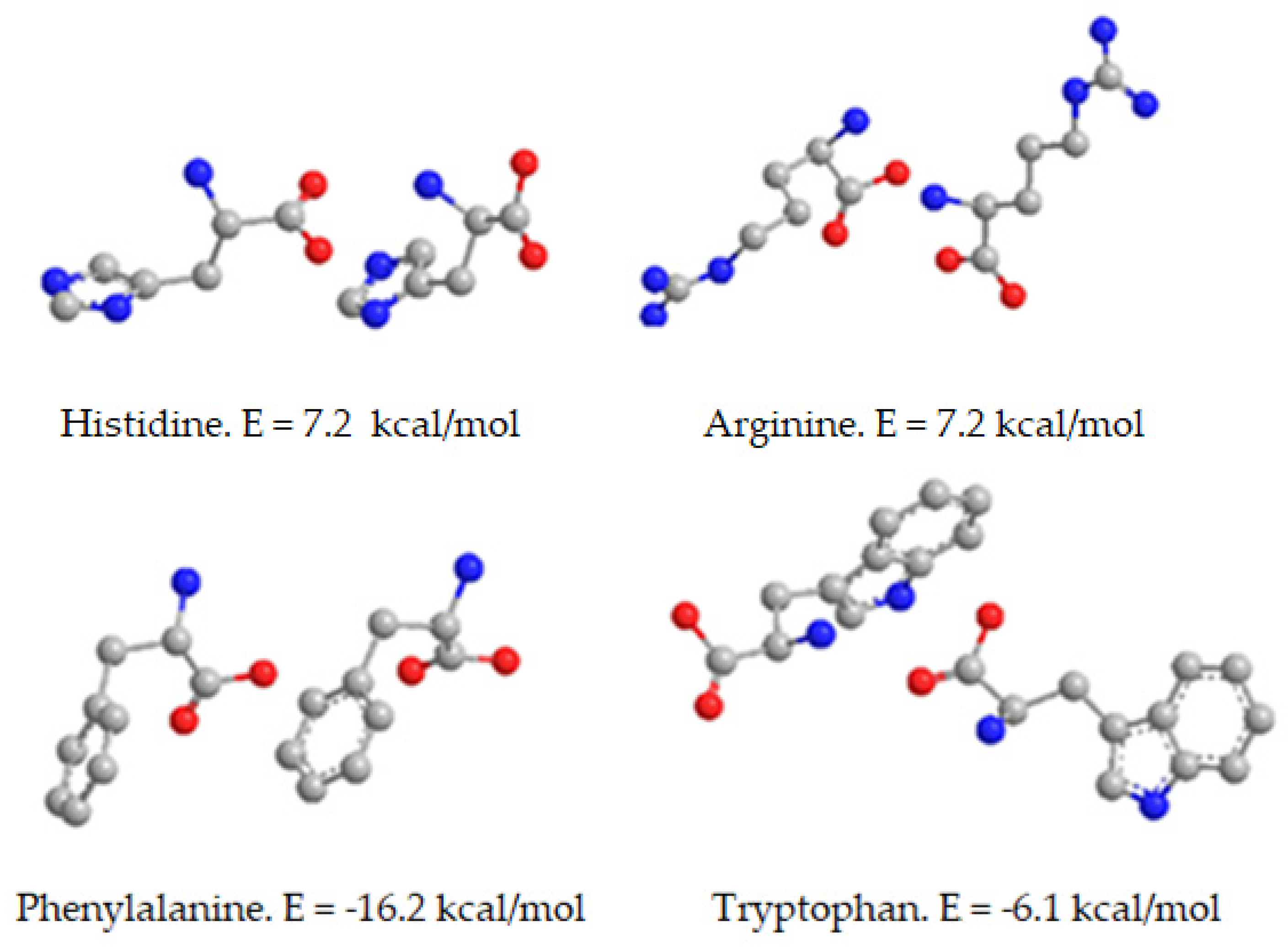
For this reason, we computed the energy and favorable spatial orientation of zinc gluconate when complexed with natural compounds, such as amino acids (phenylalanine, tryptophan, histidine and arginine). The most favorable forms of free amino acids were as dimers with different spatial orientation, according to our energetic calculations (
Figure 7). The dimer of amino acid with lowest energy level was arginine (E = −16.7 kcal/mol) whereas the least stable and most energetic dimer of the amino acids studied corresponded to tryptophan (E = −6.1 kcal/mol). The spatial orientations are shown in
Figure 7.
When complexes with zinc gluconate, the computed energy and spatial orientation of the studied amino acids indicated the favorable formation of the corresponding ionophores of zinc cation surrounded by two molecules of amino acids and two molecules of gluconate, as shown in
Figure 8.
The calculated energy of free zinc gluconate was of E = −358.3 Kcal/mol (
Table 1). However, when complexed with the amino acids, the computed energy was lower in all cases studied, indicating that there are favorable mechanical forces that make these complexes more stable and less energetic than in their corresponding free forms. For the amino acids studied, the lowest value of computed mechanical energy was obtained for the complex of tryptophan with zinc gluconate (
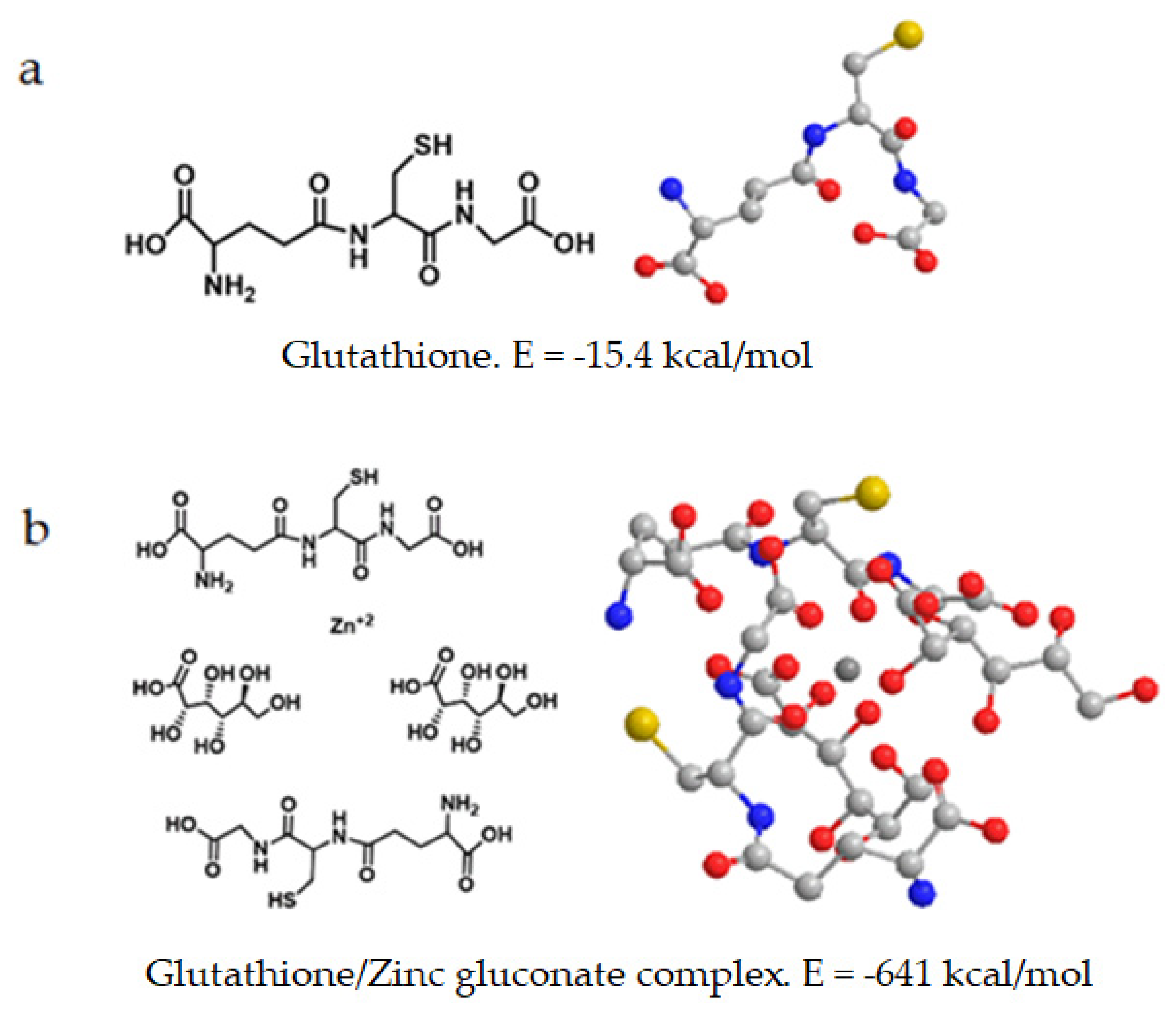
Table 3). The lowest energy of all compounds is obtained for the glutathione/zinc gluconate complex, with −641 kcal/mol.
The computed energy value of glutathione and the spatial orientation of both free form and when complexed with zinc gluconate is shown in
Figure 9.
The complex of glutathione with zinc gluconate showed the least energetic value (E = −641 kcal/mol) obtained from the compounds studied (
Table 2). The most favorable spatial orientation corresponded to the zinc cation surrounded by four molecules: two outer antiparallel mirrored images of GSH molecules at opposite ends and two inner gluconate molecules with same orientation (
Figure 9b).
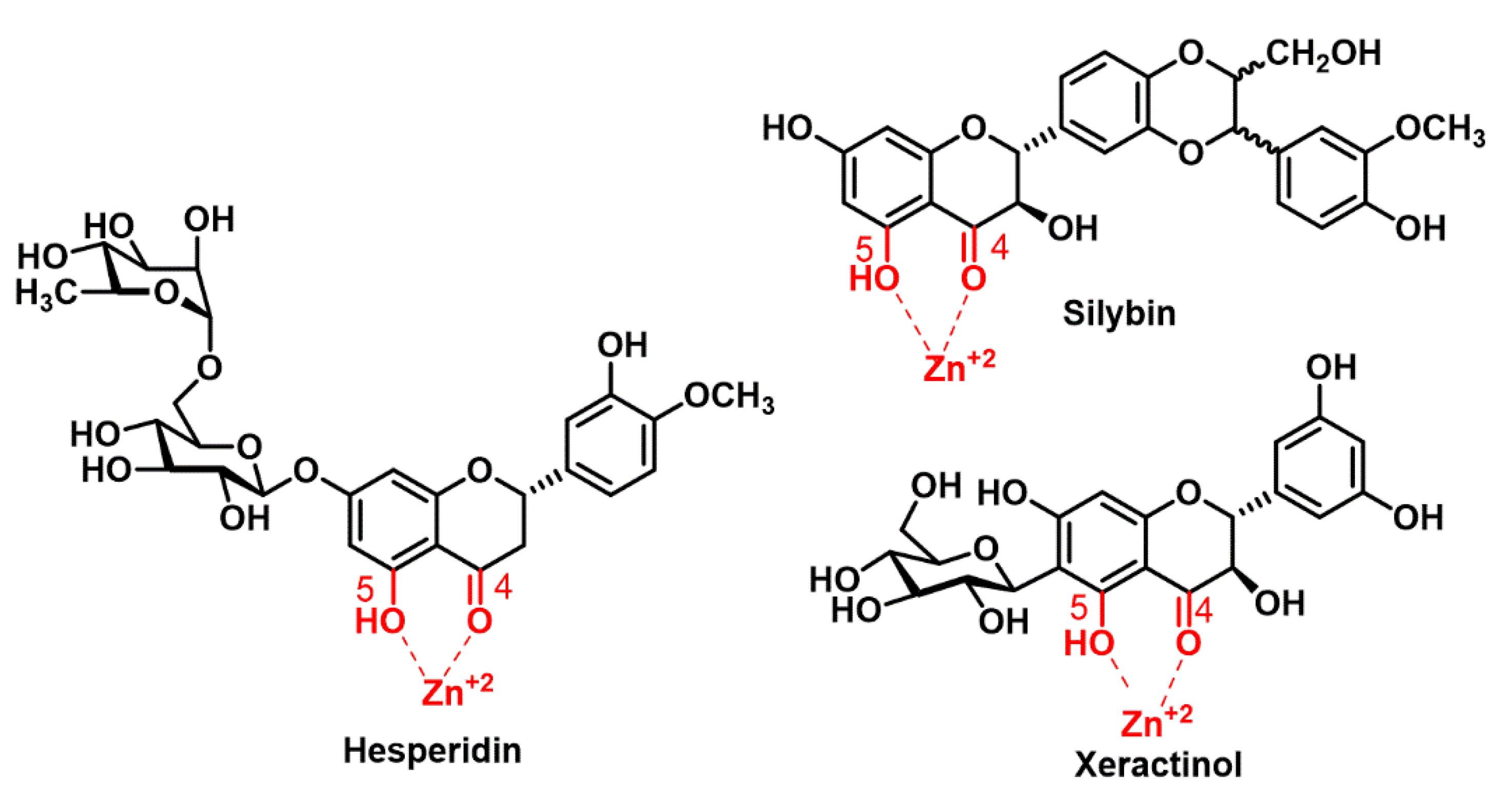
Finally, we performed a computational analysis of the complex of three well known polyphenols (hesperidin, silybin and xeractinol) with zinc gluconate, with the assumption that the high stability of the complexes was due to the formation of a greater number of intramolecular hydrogen bonds between the oxygenated functions of the polyphenol and the zinc cation. The zinc binding sites can be formed through the hydroxyl and carbonyl groups of the polyphenols (
Figure 10).
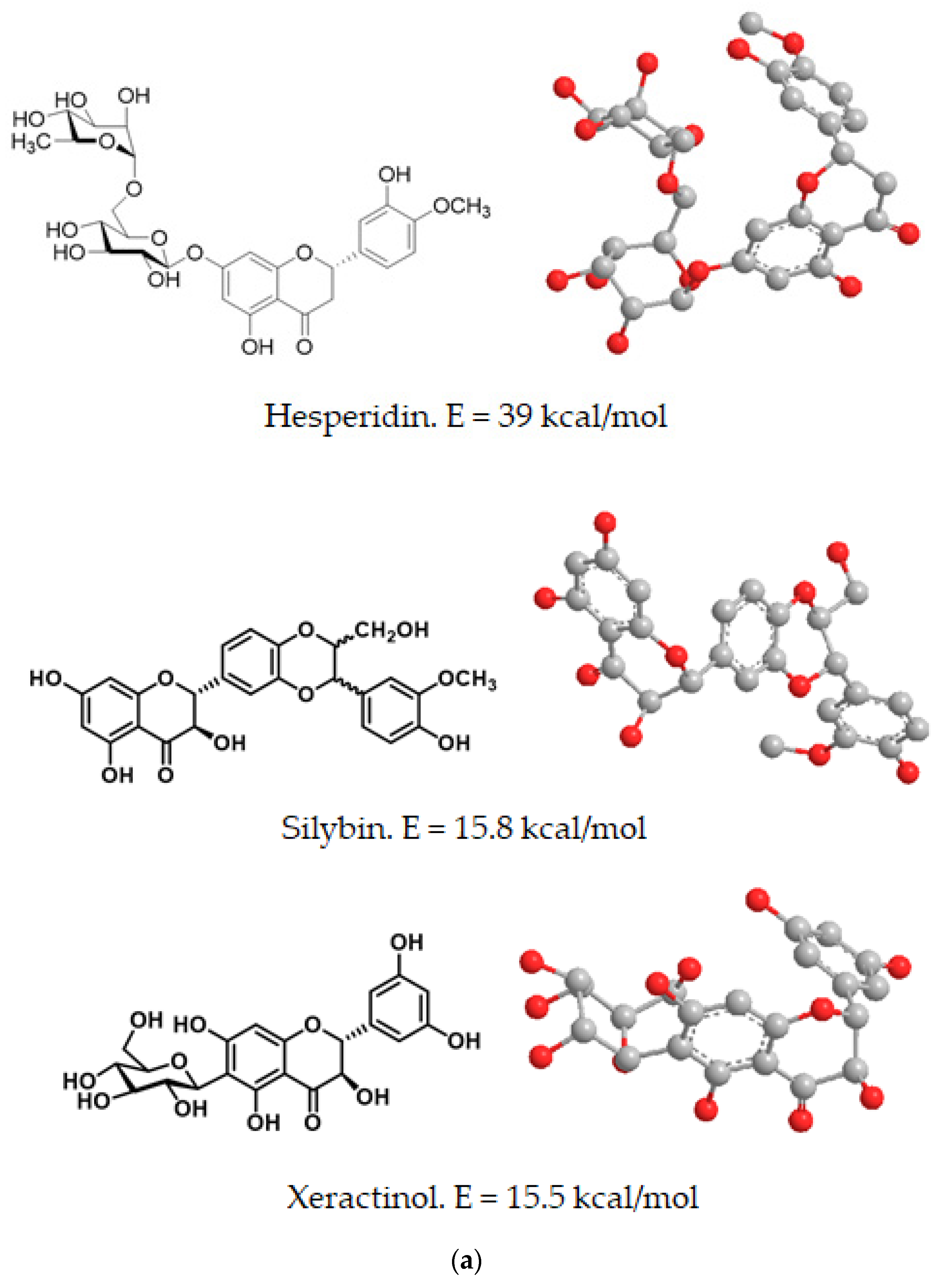
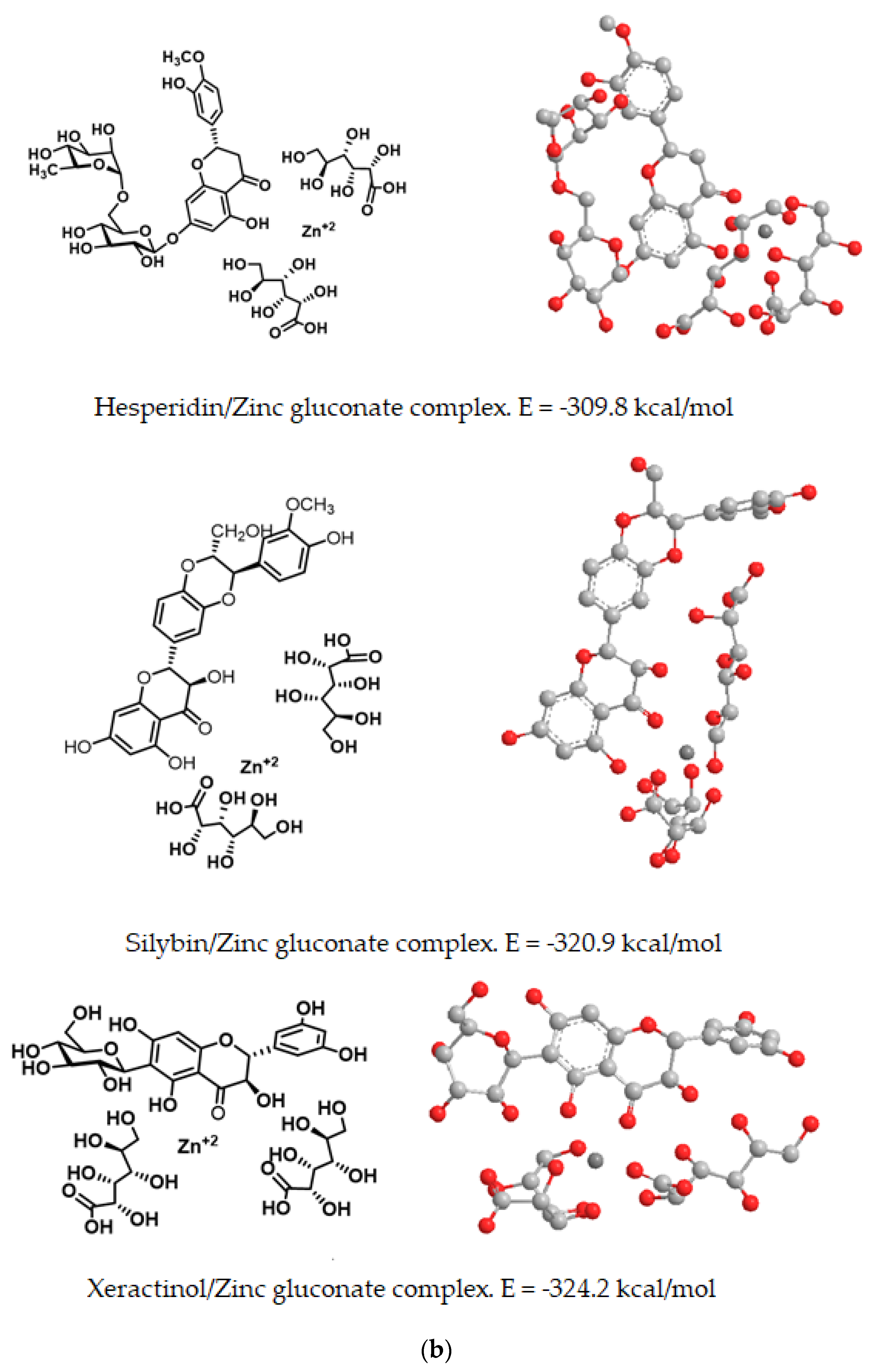
Both in free form and in combination with zinc gluconate, the computed energy content and spatial orientation of dietary polyphenols (hesperidin, silybin and xeractinol), as representatives of the flavonoid family, is shown in in
Table 2 and
Figure 11.
The lowest energy value, as computed by molecular mechanics and thus most stable form of these complexes was found for the xeractinol/zinc gluconate (E = −324.2 kcal/mol) and the higher value of computed energy (E = −309.81 kcal/mol) was fond for the hesperidin/zinc gluconate complex (
Figure 11). In all flavonoid studies, the complexes with zinc gluconate consisted in an internal zinc cation surrounded by two molecules of gluconate and one molecules of flavonoid with their carbonyl or hydroxyl groups facing internally to facilitate the coordination with the internal zinc (
Figure 11).
3. Discussion
We here provide a computational energetic analysis of the forming zinc complexes to determine whether natural substances, such as flavonoids, glutathione and plant amino acids, can act as novel zinc ionophores. Our research is focused on the study of molecular conformational structure of the zinc complexes with the lowest amount of energy. This favorable energy content indicates that polyphenols, glutathione and amino acids may form zinc complexes, which might function as novel ionophores. These natural compounds might be attractive candidates for the treatment of diseases caused by zinc deficiency.
The chemistry of ionophore molecules is an important area of supramolecular chemistry [
24]. The notion of a metal ionophore consist of a ligand attached to a metal cation in a reversible manner. In the bonded form, the metal complex is generally charge neutral as well as sufficiently hydrophobic to facilitate membrane passage across lipid membranes. Metal cation release can then be activated by either a low local concentration of the metal cation or the presence of a stimulus that favors metal cation release. Due to the reversibility of the metal binding process, the ionophore may repeatedly transport metal cations across membranes [
25].
Experimental determinations of the stability constants of ionophore metal-complexes and the corresponding free energies of complexation reactions are time-consuming and expensive tasks. Stable and favorable complexation with metals may improve ionophore bioavailability to cells, being a major critical step in the process of a chemical acting as zinc ionophores [
26]. As a result, a theoretical quantitative assessment of complex stability, together with the analysis of the geometry and spatial orientation around the metal cation, may prove to be a valuable complement to experimental investigations, allowing researchers to decrease the number of tests performed and to identify the strategy of “optimization” of known metal binders, among other advantages.
The zinc ion is stereo-chemically inactive. Zinc conformational structure is not important in our study, only as a chelating agent. The zinc ion is highly flexible in terms of coordination numbers, exhibiting coordination dynamics that do not conform to the stereochemical restrictions common to transition metal ions. Additionally, it is distinguished from magnesium (Mg
2+) and calcium (Ca
2+) because it forms considerably stronger complexes with water and a range of anions and ligands [
26]. These features are critical for the proper functioning of its biological activities.
To validate our molecular mechanics approach, we first computed the energy content of several known zinc ionophores (pyrithione, PBT2, TPEN, clioquinol and hinokitiol) both as free molecules and when forming complexes with zinc cation. In all cases studied, the values obtained for the free molecules had positive values and negative values when complexed with zinc cation. In some complex up to 100 kcal/mol lower. This reduction in conformational energy implies the favorable formation of the zinc complexes, as corresponding for zinc ionophores.
Another aspect of potential ionophore molecules is the geometry of the most stable form of the ionophore-metal cation complex [
25]. Ionophores are flexible, the final spatial orientation of the complexes includes configurational or conformational remodeling of the ionophore to accommodate the internalization of the metal ion. Ionophores, in general, are molecules having backbones of a range of different topologies that include oxygen or nitrogen atoms that are strategically spaced [
25]. The liganding atoms include a variety of functional groups, including ether, alcohol, carboxyl and amide, among others. It is through ion-dipole interaction that the neutral oxygen or nitrogen function as a ligand of cations, which is comparable to the solvation of ions in highly dielectric fluids. For the permeation through the lipophilic environment of the membrane, the polar groups of ionophore complexes must face inward, while the outside of the complex is covered with different hydrocarbon hydrophobic groupings. In our analysis, the most favorable geometry of the complex zinc pyrithione showed the zinc cation in an internal position between two adjacent pyrithione molecules, with their polar groups orienting towards the inside [
27]. Similarly, for the zinc ionophore TPEN, the spatial orientation of the most favorable complex with zinc showed an internal zinc cation surrounded by four pyridine rings. The resultant complex’s lipid solubility of ionophores can be explained in part by the efficient shielding of its polar core, which delocalizes the cation charge and in part by the complex’s external compatibility with low-dielectric solvents [
28]. Several articles have recently been published that report the utility of theoretical calculations on these kind of chemical interactions [
29,
30].
In human nutrition, zinc deficiency is a global nutritional problem. Zinc supplementation has been used for the treatment of several diseases, but mainly for its potential ability to attenuate the accumulation of free radicals [
31]. The total zinc content of eukaryotic cells is quite high, approximately 200 µM. However, the majority of cellular zinc is firmly bound to proteins [
32]. A weak-binding fraction, capable of rapidly triggering ligand exchanges, is represented by the free (bioavailable, rapidly exchangeable, or labile) zinc ion. This labile zinc comprises the major mobile reactive pool of cellular zinc. This pool is not a viable source for incorporation into proteins, as it is loosely bound to low molecular ligands (such as amino acids and citrate). When labile zinc combines with a low-molecular-weight ligands or chelators to be absorbed, solubility is increased [
33]. As a result, the chemical form in which this labile zinc is made available is of critical consideration. Amino acids (e.g., histidine, tryptophan) and organic acids (e.g., citrate, gluconate) have been used to improve zinc absorption and have been utilized in zinc supplements to improve zinc bioavailability. In most of these studies usually only one type of Zn
2+salt was used rising the question whether different counterions and ligands of Zn
2+ may influence its uptake and antioxidative function.
Zinc supplementation has been proposed as an efficacious chemo-preventive agent for some cancers [
34]. Zinc can restrict the growth of cancer cells via cell cycle arrest, as well as induction of apoptosis and necrosis [
35]. In clinical studies oral supplementation with zinc gluconate resulted in a decreased frequency of infections, production of tumor necrosis factor alpha (TNF-) and oxidative stress in elderly subjects, compared to placebo [
36]. Therefore, any strategies that can increase zinc levels in the cancer cells are likely to be effective cancer treatment strategies, having minimal toxicity to normal cells. Accordingly, various complexes between zinc and bioactive molecules have been prepared and explored in the development of new antitumor drugs [
25]. Although zinc sulfate is the most widely utilized zinc form, zinc gluconate stabilized with glycine has the same bioavailability and metabolic behavior as zinc sulphate in aqueous solution with the advantages of good solubility, mild flavor and low cost of production [
37]. Our theoretical results suggest that zinc gluconate forms the most favorable complex of zinc among four zinc salts of organic acids: zinc gluconate, zinc citrate, zinc acetate and zinc lactate. These organic salts are already available on the market as zinc food supplements. All salt studied have negative energy values of up to E = −358.3 kcal/mol, as in the case of zinc gluconate, indicating that they were acting as zinc complexes. Therefore, zinc gluconate was used as a starting point for its complexation with amino acids, glutathione and flavonoids.
Tryptophan and histidine-rich sites exposed to the solution could be the most likely targets for the first phase of the metallization process by zinc [
38]. Zinc complexes with proteinogenic amino acids have been recently proposed as potential candidates for dermatological treatments in the cosmetic and pharmaceutical industries. Theoretically, these derivatives should be better absorbed and safer due to their non-ionic structure and weak influence on the skin pH, which will make them excellent ingredients for dermal products [
39]. Our theoretical tree-dimensional study shows that histidine and tryptophan formed stable complexes with zinc gluconate. The complexes obtained with tryptophan or histidine have lower energy values, up to 300 kcal/mol less than in the case of zinc salts of organic acids, indicating the formation of a more stable basal complex than the individual compounds. We observed that more ligands are formed between the zinc cation of the primary ionophores with the amino acids and polyphenols. The decrease in the total energy of the complex might be due to the formation of hydrogen bonds between the hydroxyl and amino groups present in final complex, which results in greater energy stability of the final complex. This is consistent with the finding that, in clinical studies, orally administered zinc-histidine displayed a 3-fold bioavailability, as compared to zinc sulphate [
40]. Zinc salts of amino acids may exert neuroprotection. In the brain, zinc-histidine was revealed to be the preferred substrate for membrane transport, suggesting that amino acids play a role in biological zinc complexation [
26]. Zinc-histidine has been studied as the most effective yet least toxic form of zinc for the treatment of central nervous system disorders associated with zinc deficiency. The neuroprotective role of zinc supplementation with histidine is related to the attenuation of oxidative stress [
41].
Glutathione is a key antioxidant peptide found in plants, animals, fungi and some bacteria and archaea, capable of preventing damage to important cellular components caused by reactive oxygen species (ROS), such as free radicals, peroxides, lipid peroxides, as well as heavy metals [
42]. Reduced glutathione GSH is considered a chelating ligand for zinc, as they form various complexes [
26]. It is a peptide formed between the carboxyl group of the glutamate side chain and cysteine, where the carboxyl group of this residue is attached by peptide linkage to glycine. Our theoretical result also showed that the complexes obtained with glutathione have much lower energy contents than zinc gluconate.
Flavonoids are antioxidants present in food and play an important role in the defense against oxidative stress [
43], which is present in various physiological and pathophysiological processes. The antioxidant activity of phenolic compounds results from a combination of their transition metal chelating and free radical scavenging properties. In addition, they act by inhibiting enzymes such as oxidases, lipoxygenase, myeloperoxidase, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and xanthine oxidase, preventing the generation of ROS in vivo, as well as organic hydroperoxides [
44,
45]. For example, zinc and polyphenols play an important antioxidant role in the immune response against SARS-CoV-2 [
46]. We first studied the potential interaction of the flavonoids silybin, with zinc salts of organic acids. The energy of silybin, when forming complexes with zinc salts of organic acids showed lower values for zinc lactate and zinc acetate, but slightly higher for zinc gluconate and zinc citrate. This is consistent with previous reports showing that the flavones morin and quercetin interfered with zinc accumulation, which resulted in growth improvement. However, supplemental zinc turned the initially benefic action of morin and quercetin into potential toxicity, which may increase the anticancer potency of these drugs [
47]. The complexation with Zn
2+ also improved the biological activity of the free rutin. Rutin–zinc has shown cytotoxicity against leukemia, multiple myeloma and melanoma cell lines, but did not show any cytotoxicity against normal cells in vitro. One explanation is that Rutin–zinc complex is more efficient as an antioxidant agent than free rutin [
48].
In humans, diabetic polyneuropathy has been documented to improve by taking supplements of zinc/polyphenol [
49]. When we consume zinc without any food, iron inhibits the absorption of zinc in our body, with a molar ratio of iron:zinc of 25:1 [
16]. The formation of poorly soluble complexes with natural chelating agents, such as inositol hexaphosphate and pentaphosphates (phytic acid), leads to reduced assimilation of zinc. Phytates, present in foods such as cereals, corn and rice, has a very negative effect on zinc absorption [
50]. These metal-polyphenol complexes are reported to exhibit better pharmaceutical properties. Some studies indicate that certain individuals should anticipate their zinc status if they routinely eat procyanidin-containing food [
51].
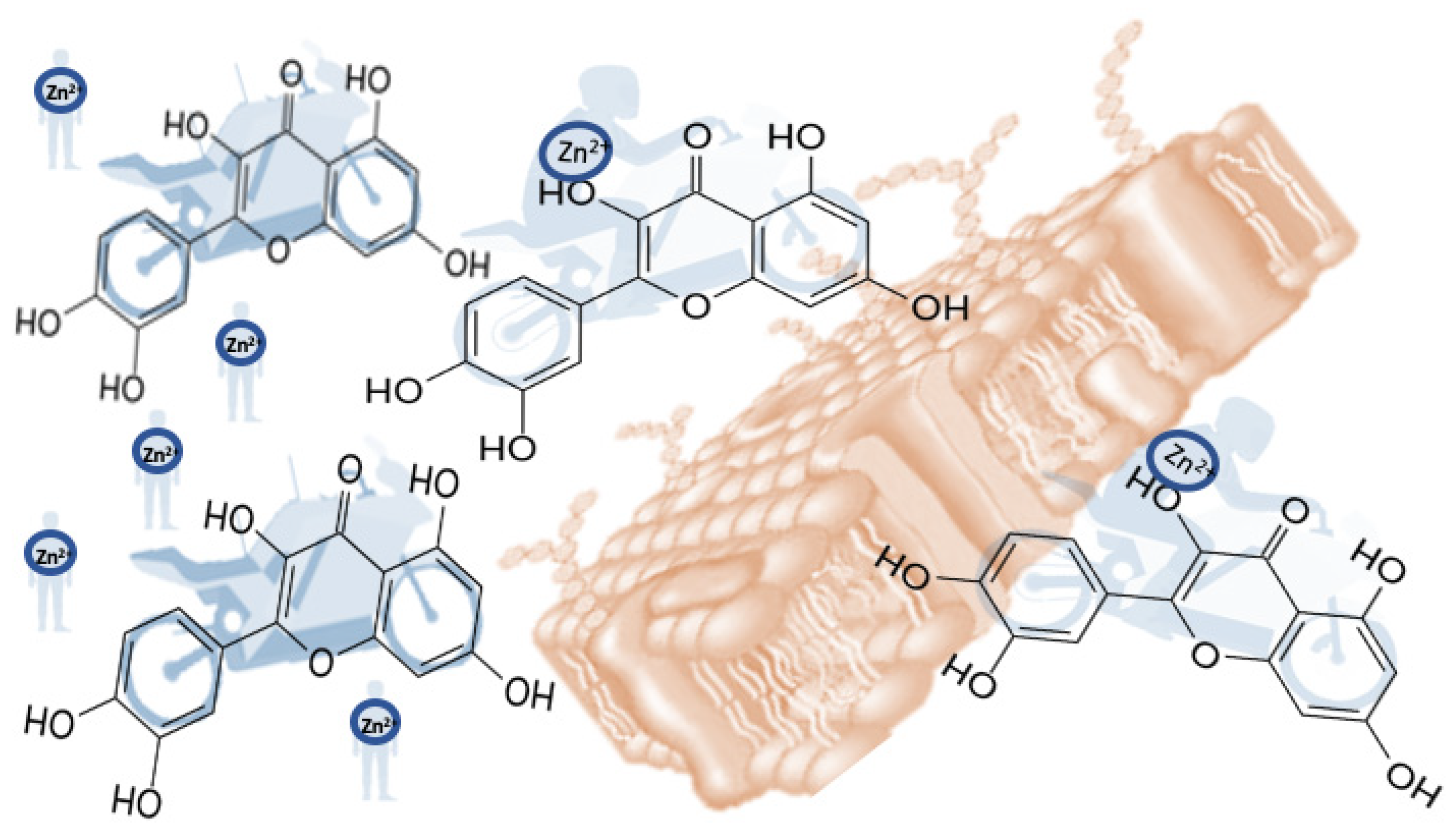
Ionophores are molecules that alter the permeability of biological membranes in response to certain metal ions for which they have an affinity and selectivity. Ionophores form a strong bond with a specific metal ion, shielding it from the effects of the surrounding environment. As a result, the metal ion is more easily transported into the hydrophobic core of the lipid membrane. Thus, ionophores function as a mobile carrier, transporting ions through the hydrophobic environment of cell membranes. We can illustrate the capacity of flavonoids, for example, as zinc ionophores, with a motorcyclist/motorbike interaction model. The biker would represent the zinc cation, which by itself cannot cross the membrane without the help of the motorbike, which, in this case, would be the flavonoid acting as an ionophore (
Figure 12).


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}