
How Threatened Is Scincella huanrenensis? An Update on Threats and Trends
by
 , ,
, ,
Yucheol Shin
1,2,† ,
,
Kevin R. Messenger
3,†,
Kyo Soung Koo
4,
Sang Cheol Lee
5,
Mian Hou
6 and
Amaël Borzée
1,*
1
Laboratory of Animal Behaviour and Conservation, College of Biology and the Environment, Nanjing Forestry University, 159 Longpan Rd, Nanjing 210037, China
2
Department of Biological Sciences, College of Natural Science, Kangwon National University, Chuncheon 24341, Korea
3
Herpetology and Applied Conservation Lab, College of Biology and the Environment, Nanjing Forestry University, 159 Longpan Rd, Nanjing 210037, China
4
Research Institute of EcoScience, Ewha Womans University, Seoul 03760, Korea
5
Bioresource and Environmental Center, Incheon National University, Incheon 22012, Korea
6
College of Continuing (Online) Education, Sichuan Normal University, Chengdu 610068, China
*
Author to whom correspondence should be addressed.
†
The authors contributed equally.
Conservation 2021, 1(1), 58-72; https://0-doi-org.brum.beds.ac.uk/10.3390/conservation1010005
Submission received: 9 March 2021
/
Accepted: 22 March 2021
/
Published: 25 March 2021
Abstract
:It is important to understand the dynamics of population size to accurately assess threats and implement conservation activities when required. However, inaccurate estimates are harming both the threat estimation process, and the resulting conservation actions. Here, we address the extinction threats to Scincella huanrenensis, a species described in the People’s Republic of China, but also occurring on the Korean peninsula. Estimating the threats to the species is not an easy task due to its unknown population status in the Democratic People’s Republic of Korea. Here we analysed the literature to acquire the known presence point for the species, along with datapoints originating from opportunistic field surveys, and employed habitat suitability models to estimate the range of the species. We then followed the categories and criteria of the IUCN Red List of Threatened Species to assess the extinction risk of the species. We found the species not to be fitting the threatened category at the global scale based on the range size, the only category for which enough data was available. We recommend the status of the species on the IUCN Red List of Threatened Species to be updated as it is now listed as critically endangered (CR), a listing fitting a national assessment for the People’s Republic (PR) of China. While this species is possibly less threatened than currently listed, this is not a genuine improvement, and specific conservation aspects should not be neglected due to its specialisation to medium to high elevation habitat.
1. Introduction
Determining the threat to a species is of principal importance to establish corresponding conservation plans and prevent the species from going extinct [1]. The International Union for Conservation of Nature (IUCN) Red List of Threatened Species is the principal global agency taking care of updating such statuses, and the only organisation that has a general reach [2,3]. These lists are regularly updated when resources are available [4], and conservation resources are generally dependent on these lists, with good practices having resulted in positive conservation results [5]. For instance, the adequate assessment of the Oaxaca treefrog (Sarcohyla celata) has resulted in conservation activities from research and local communities, and the species has consequently been moved from Critically Endangered (CR) to Near Threatened (NT) [6,7].
In opposition, inadequate assessment can result in inaccurate listing in the IUCN Red List of Threatened Species, resulting in the diversion of already limited resources away from the most needing taxa. Similarly, threats to a species or a sub-population may be overlooked, resulting in increased risk to the species. Another negative outcome from inadequate assessment is the resulting loss in public trust from mistakes, leading to a devaluation of the work conducted by the IUCN Red List of Threatened Species since its establishment [8,9]. It is however important to consider the difference between the absence of data, where threats in a region can be overlooked because the species is not known to be occurring, and the omission of existing data. Here we address the case of the Huanren ground skink, Scincella huanrenensis, a species present in the north-eastern part of the People’s Republic of China (hereafter PR China) and across the Korean peninsula. The species has been assessed as Critically Endangered (CR) by the IUCN Red List of Threatened Species [10] based on a subset of its range, and Near Threatened (NT) in the Republic of Korea (hereafter R Korea; [11]).
As the species ranges through the Democratic People’s Republic of Korea (hereafter DPR Korea), an area where collecting data is comparatively difficult, we modelled the distribution of the species, and established a proxy of the extent of occurrence (EOO) based on the model results. We then followed the categories and criteria of the IUCN Red List of Threatened Species, and recommended an update in the listing of the species based on the category and criteria we were able to apply [12].
2. Materials and Methods
2.1. Species Introduction
Scincella huanrenensis Zhao and Huang 1982 was described based on an adult male (holotype DMNH 810045), an adult female (paratype DMNH 810025) and 64 paratypes from Huanren County, Liaoning Province, PR China and deposited in the Dalian Museum of Natural History and Chengdu Institute of Biology (Figure 1; [13]). The species has been recorded at mid-elevation on the Korean peninsula and the Liaoning province in China [14,15]. In these regions, the species inhabits valleys, rocky outcrops, small agricultural fields adjacent to forests, mountain trails, and mid-elevation basins, where it generally uses rocks, fallen logs and the leaf litter as a principal habitat [16,17]. Scincella huanrenensis is similar to S. tsinlingensis and S. vandenburghi but differs in scalation from both species [13,18]. The species is ovoviviparous, giving birth to two to five juveniles in China [19], four to five juveniles in DPR Korea [20,21], and five to ten juveniles in R Korea [15].
2.2. Data Collection
To determine the range of S. huanrenensis, we relied on three types of data, resulting in a total of 40 occurrences (Table 1). First, we used datapoints collected by the authors during field surveys. None of the surveys were conducted with the aim of detecting this species, but all surveys had a herpetological focus. There is therefore no unified protocol, and all observations can be defined as opportunistic.
The second type of data included in the dataset originates from the literature. We used Google Scholar with the key word “Scincella huanrenensis” and analysed all 54 articles resulting from the search. Most articles did not provide any geographical reference, or were already including the datapoints originating from the field surveys [15,18]. Only some articles were precise enough for us to be able to extract coordinates to be used for the analyses [13,14,22,23] (with [23] linked to specimens deposited in the natural history museum of Ewha Woman’s University). In the case of the holotype description, we determined the approximate GPS coordinates based on the explanation provided in the text. In addition to the literature searched online, we consulted the works of Won [20] and Kim and Han [21], which together provide the most comprehensive overview of herpetofauna in DPR Korea. In their species accounts, the only scincid species is presented as “Lygosoma laterale”. However, we were able to confidently identify this name to be S. huanrenensis based on the combination of morphological characteristics and ovoviviparity of these skinks, as reported by both authors. The latter characteristic is especially conclusive in identifying S. huanrenensis because the sympatric species S. vandenburghi is oviparous [15,24]. As the reported localities largely overlapped between the two studies, we extracted five unique datapoints from both Won [20] and Kim and Han [21]. There are no available vouchered specimens of S. huanrenensis collected from DPR Korea. Therefore, these datapoints represent the only available evidence of the presence of this species in the country.
The third type of data collected was extracted from citizen science databases, including Global Biodiversity Information Facility (GBIF.org, https://0-doi-org.brum.beds.ac.uk/10.15468/dl.sh22cc; accessed on 30 October 2020) and iNaturalist (inaturalist.org; https://www.inaturalist.org/observations?taxon_id=37666; accessed on 30 October 2020). As some of the datapoints are transferred from iNaturalist to GBIF, we excluded all duplicate datapoints before further analyses (Table 1). We also included survey records of the 4th National Ecosystem Survey conducted between years 2014 and 2018 by the National Institute of Ecology in R Korea (accessed through the EcoBank platform; [25]).
In addition, we added the type locality of the species, which was the basis for the IUCN assessment published in 2019. Furthermore, based on the known presence of the species in Jilin and DPR Korea but the absence or insufficiency of GPS coordinates, we also determined one putative GPS coordinate for each region in order to create a bias layer to correct for variations in sampling efforts between regions. Both putative sites were selected to match the habitat of the species within the given area, and elevation at which it generally occurs [10]. The first putative point was extracted from the data provided by Bobrov [26] and located at 42.160443° N and 125.662098° E. The second datapoint was communicated by a herpetological researcher in DPR Korea at a workshop in Pyongyang in 2019—38.369705° N and 128.161001° E. The datapoint itself may not be perfectly accurate but the species is known to be present in the area. These two putative datapoints were only used to weight the model against sampling bias (see Section 2.3. Range modelling), and not included in the analysis itself. Their putative nature does not create a caveat in our analyses.
2.3. Range Modeling
We obtained 19 bioclimatic and elevation layers from WorldClim 2.1 [27]. In addition, we considered three vegetation layers (broadleaf, needleleaf, and mixed forest covers) and two topographical variables (slope and terrain ruggedness) downloaded from EarthEnv (http://www.earthenv.org/; accessed on 25 January 2021; [28]) as the limited ecological information on the species suggest preference for mountainous forest habitat [13,18]. All layers followed a 1 km spatial resolution and were cropped to a combined extent in the Korean Peninsula, Jilin province, and Liaoning province to fully encompass the known occurrences as well as the broader area surrounding them. Next, we created a model calibration area to define the area that has been accessible to dispersal by the species during its evolutionary and biogeographic history [29]. We created the calibration area by applying a 100 km (0.9 dd) radius circular buffer around all occurrence points and four random points generated within DPR Korea, and we then cropped all environmental layers to this area (Figure 2). We generated four random points in the “gap” area between georeferenced and putative datapoints in DPR Korea, Jilin, and Liaoning to create a non-disjunct calibration area covering the total range of the species. While the dispersal ability of this species is unknown, it is likely to be limited by the presence of certain environmental features (e.g., low elevation plains and basins) [30] and the presence of unsampled populations is highly likely. Therefore, we considered the buffer radius defined here to be sufficient to take unsampled populations into account without overestimating the dispersal potential. Next, we spatially thinned our occurrence dataset to reduce spatial clustering using the R package spThin [31]. We applied a thinning distance parameter of 1 km to match the spatial resolution of our environmental layers. The thinning process reduced the number of datapoints from 40 to 31. We used these spatially thinned occurrences for all downstream analyses.
To build accurate habitat suitability models, it is important to select environmental variables that have low collinearity but are also relevant to the species’ ecology [32]. Thus, we followed a two-step approach to select the environmental variables used in our analyses. First, we ran five replicated models in Maxent (software version 3.4.1; https://biodiversityinformatics.amnh.org/open_source/maxent/; accessed on 5 February 2021) using the default settings, and all 25 environmental variables as inputs, and removed the variables with contributions lower than 5%. Then, we conducted a Spearman’s rank correlation test among the retained variables and removed the ones that were highly correlated (coefficient > 0.7). This process resulted in the selection of the following variables—bio 3 (isothermality), bio 12 (annual precipitation), bio 17 (precipitation of driest quarter), and elevation. While the three forest cover layers were not selected due to low contribution scores in the initial Maxent run, we included them independently in subsequent models considering the known ecology of the species [15,20,21]. Therefore, a total of seven environmental variables were used for all downstream modelling. Layer cropping and Spearman’s test were conducted in R version 3.6.3 [33] using packages raster [34], rgdal [35], rgeos [36], and ntbox [37].
As our occurrence data were heavily biased towards R Korea, we adjusted this issue by creating a bias layer. First, we sampled 10,000 random points in proportion to the number of occurrences in each section of our study area (R Korea, DPR Korea, Jilin and Liaoning). As a result, we randomly sampled 68.8% of the random points in R Korea (known presence point n = 22), 18.7% from DPR Korea (known presence point n = 6; including one putative datapoint), 3.1% from Jilin (known presence point n = 1; one putative datapoint), and 9.4% from Liaoning (known presence point n = 3). Next, we converted these points into a density raster reflecting an overall sampling effort per region [28]. We implemented this process using R packages dismo [38] and MASS [39].
To find optimal Maxent model parameters for our analyses, we ran a series of models in the ENMeval package [40] in R. From a total of 48 candidate models, we selected a combination of Linear and Quadratic feature classes and a regularization multiplier of 0.5 based on the AICc value (Supplementary Table S1). In total, we generated 15 subsample replicated models with random seed enabled, using 5000 iterations and 20% of data randomly selected for model testing. In addition, the density raster generated from proportionately sampled random points was used as an input to modify the selection of background points. We took the average of 15 replicated runs and evaluated the model performance based on the area under the receiver operating characteristic curve (AUC), for which a value closer to 1 indicates good model performance [29], the True Skill Statistic (TSS), which ranges from −1 to 1 [41], and our knowledge on the species. We used the model averaged across 15 replicates trained on the calibration area for downstream analyses on the extent of occurrence (EOO) and threat status assessment. We also projected this model to the entire extent of R Korea, DPR Korea, Jilin, and Liaoning to visualize potentially suitable but unsampled areas to aid future survey efforts.
In a separate modelling attempt, we used the biomod2 package [42] in R to build an ensemble species distribution model. The advantage of this approach is that it combines predictions of multiple modelling algorithms and thereby reduces predictive uncertainties of a single modelling method [43]. We used the following six algorithms for modelling—Classification Tree Analysis (CTA), Generalized Boosting Models (GBM), Generalized Linear Models (GLM), Random Forests (RF), Surface Range Envelope (SRE), and Maximum Entropy (Maxent). We did not use other modelling algorithms available in biomod2 due to the continued failure of the models generated from these algorithms to converge. The environmental variables used were the same as for the Maxent modelling described above, and we randomly sampled three sets of 10,000 pseudo-absence points. While model parameters for other algorithms were set to default values, we specifically tuned Maxent parameters to the values determined by ENMeval (Linear Quadratic features, regularization multiplier = 0.5). We ran five replicates per pseudo-absence set per modelling method, resulting in a total of 90 individual models. We evaluated the performance of individual models using AUC and TSS. To construct a final ensemble model, we filtered and selected the models with TSS values greater than 0.7 and implemented committee averaging. We projected the ensemble model to the entire study area and evaluated the performance of committee-averaged model based on AUC, TSS, and our knowledge on the distribution of the species.
In addition to the three evaluation criteria (AUC, TSS, expert knowledge), we compared both Maxent and ensemble models to five null models as an additional measure of model performance. We generated each null model in the Maxent software using a set of 31 random points (equal number to the spatially thinned species occurrence data), auto features, default regularization multiplier (=1), 5000 iterations, random seed enabled, 10,000 randomly selected background points, and in 15 subsample replicates with 20% of data used for model testing. We evaluated the performance of null models using the AUC and the plotted distribution.
2.4. Estimating the Extent of Occurence
We then estimated the extent of occurrence (EOO) of the species following the criteria of the IUCN Red List of Threatened Species. We followed two complementary methods, the first one relying on the minimum convex polygon (MCP), computed in R using the package ConR [44], and confined to the land area. The second method relied on the calculation of the area of presence, here matching with habitat suitability, by converting the continuous probability surface originating from the Maxent model output into a binary presence/absence map. To do so, we reclassified the output model raster using Maximum Training Sensitivity plus the specificity threshold value (=0.58). Following this process, cells with a suitability value greater than the threshold received a value of 1 (presence) whereas cells with a suitability value lower than the threshold received a value of 0 (absence). We implemented the raster reclassification and area calculation in R version 3.6.3 [45] using the package raster [34].
2.5. Assessment
The threat assessment conducted here follows the categories and criteria of the IUCN Red List of Threatened Species as they are the most robust [46] and they adequately represent threats to a species at the global scale [47]. The IUCN Red List is divided into three threatened categories—Vulnerable, Endangered, and Critically Endangered, and assessments are conducted against quantitative thresholds for the five criteria that determine whether a species is at risk of extinction—A, population size reduction; B, geographic range size; C, small population size and decline; D, very small population and/or restricted distribution; and/or E, quantitative analysis of extinction risk. A species that does not meet any of these criteria is placed into one of the other non-threatened IUCN categories (Least Concern and Near Threatened). Another advantage of the IUCN Red List of Threatened Species is that it can be applied to numerous types of datasets, including those with partial data [48,49].
2.6. General Threats to the Species
Here, we refer to all threats, habitats, uses and trades following the IUCN Red List criteria and categories (www.iucnredlist.org/resources/redlistguidelines; accessed on 12 December 2020). In terms of general threats in all three nations, the species is impacted by residential and commercial developments in parts of its range [50], and it is heavily harvested for traditional medicine in PR China [10], resulting in the depletion of some population and potential local extirpation. It is however not known to be harvested for the pet trade.
The species is however impacted by climate change due to its ectothermic nature [30,51]. The current call to limit global warming to 1.5 °C above preindustrial levels [52] is unlikely to be backed by efficient regulation despite the increasing number of requests [53]; and, even if efficient regulations are implemented, the environment is still estimated to be severely impacted [54,55], with a 2.3 °C air temperature and 4.4% humidity increase in Asia, a 2.7 °C and 3.3% increase in North East Asia, and a peak increase of 7.0 °C (13% humidity) if the Paris Climate Agreements are not held [56].
Scincella huanrenensis is currently known to be from forested areas only, and it is not known to occur in orchard or other tree plantations. The degradation of forests for development is likely to be the main threat to this species throughout its range [22,57,58,59]. The species is rare and observed infrequently, with a population size estimated to be small, fragmented [60], and decreasing [10]. The species is currently listed as NT in R Korea, noting the need for additional studies [11,50]. Based on the international database Protected Planet (https://www.protectedplanet.net/ accessed on 12 December 2020), the species occurs in several protected areas, including Odesan National Park in R Korea, Mount Kumgang National Park in DPR Korea and Dayahe Drifting Scenic Area in PR China. In addition, the species is listed in the “List of Beneficial or of Important Economic or Scientific Value Terrestrial Wild Animals under States Protection”, under the protection of the “Law of the People’s Republic of China on the Protection of Wildlife” [10].
2.7. Species Assessment
2.7.1. Population Size Reduction
The population size for the species is not known but it is decreasing in some areas, due to its association with old growth forests, which are likely to have strongly declined in DPR Korea [58,59], and were strongly restricted in R Korea in the mid 1900 but are now being restored [50]. The species is also known to be strongly declining in PR China [10] and it is likely under threat because of deforestation and development in R Korea [11]. The data are however insufficient for this category to be applied.
2.7.2. Geographic Range
To assess the category to which S. huanrenensis should be assigned to in view of its geographic range, we compared the values obtained from the MCP and the binary map based on habitat suitability against the thresholds used by the IUCN Red List. The criteria specify that a species should be listed as vulnerable if its EOO is < 20,000 km2 and it fulfils at least two of the three following conditions—(a) “severely fragmented”, (b) “continuing decline observed, estimated, inferred or projected in any of—(i) extent of occurrence; (ii) area of occupancy; (iii) area, extent and/or quality of habitat; (iv) number of locations or subpopulations; (v) number of mature individuals” and (c) extreme fluctuations in any of the points i to iv listed above.
2.7.3. Small Population Size and Decline
The population size of the species has not been estimated, and surveys to determine population size and abundance are not available. Thus, this category could not be applied.
2.7.4. Very Small or Restricted Population
The number of mature individuals is unknown, thus this category could not be applied.
2.7.5. Quantitative Analysis
There are no data available for a quantitative analysis, thus this category could not be applied.
3. Results
3.1. Habitat Suitability Modeling
The models generated from the actual species occurrences showed significantly better performance (AUC > 0.8; TSS > 0.6) than the null models generated from randomly sampled points (all null models—AUC < 0.5). The scores of the two evaluation metrics indicated good predictive performance for both the ensemble and Maxent models, with the ensemble model (AUC = 0.96; TSS = 0.80) performing better than the Maxent model (AUC = 0.85; TSS = 0.63). However, when we plotted and compared these two models, it was evident that the ensemble model failed to adjust sampling bias towards R Korea, while potential distribution of the species in DPR Korea was well represented by the Maxent model. In addition, the Maxent model was more consistent with our knowledge on the species’ ecology and known distribution. Therefore, we selected the Maxent model for further analyses and all models mentioned hereafter refer to this model. Plots of the ensemble and null models are available in Supplementary Figures S1 and S2.
Among the environmental variables used, elevation had the highest contribution in terms of percentage, followed by isothermality (bio3), precipitation of the driest quarter (bio17), deciduous broadleaf forest cover, annual precipitation (bio12), mixed forest cover, and needleleaf forest cover (Table 2). Elevation in highly suitable areas (suitability > 0.58, as per binarization threshold) ranged from 400–1500 m a.s.l. This is largely consistent with a previous study on the ecology of the species, which found an average elevational distribution of 619.50 ± 180.21 m a.s.1 [15].
All occurrence points fell within the climatically suitable area (Figure 3), which encompasses the Korean peninsula, and Liaoning and Jilin provinces of PR China. The area of high suitability (>0.58) was located mostly on the east coast of the Korean peninsula, along the mid- to high-elevation mountains of the Baekdudaegan Mountain Range (Figure 3A). The model projected on to the entire study extent highlighted areas of high habitat suitability (>0.58) outside the calibration area where unknown populations may be found in the future (Figure 3B).
3.2. Geographic Range Assessment
Our estimation of the EOO of the species through the MCP resulted in an area of 87,648.1 km2, and the model predicted a surface of 26,721.9 km2 based on habitat suitability (Figure 4). The species range estimated from both measures is crossing the threshold for any of the threatened categories (i.e., VU threshold = 20,000 km2). In addition, the range of the species is likely to be naturally fragmented following the landscape, but unlikely to be severely fragmented with the exception of the sub-populations in PR China. The continued decline inferred from the quality of habitat is likely to result in a decrease in the number of locations or subpopulations and in the number of mature individuals. The decline is however too low to trigger the category A based on a projected or suspected population reduction.
4. Discussion
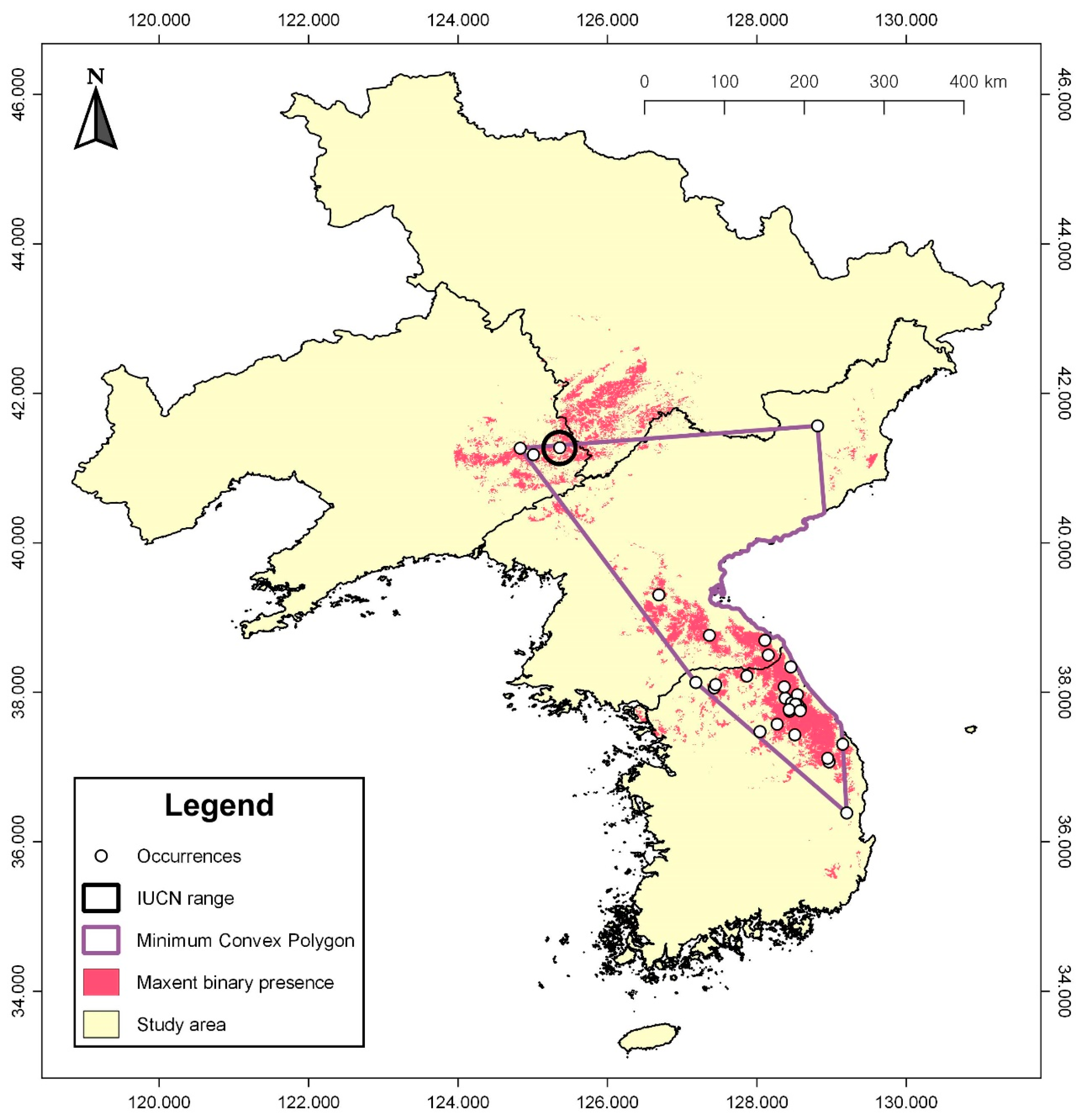
The main conclusion of our analysis is that despite a marked decline due to habitat loss, Scincella huanrenensis at the global scale is not crossing the threshold to be listed as threatened on any of the five criteria of the IUCN Red List of Threatened Species. While the results of models are always open to scrutiny [61], the surface of the MCP follows the description of the criteria. This conclusion however goes against the latest update for the species on the IUCN Red List of Threatened species, which specified an EOO between 50 and 99 km2. This published EOO is matching with the area around the species’ eponymous city, and it is likely to generally fit a national assessment for the species in PR China, although the published EOO only partially overlaps with the area of the type locality for the species (Figure 4). The discrepancy between our results and the published assessment highlights the need for international teams to assess species, especially when the species is likely to be distributed over trans-boundary ranges [62]. The update recommended here may therefore not be accurate either as it does not include researchers from the DPR Korea and it is therefore biased towards the regions where the authors are based, namely PR China and R Korea. In addition, the rather large discrepancy between the two independent measures of EOO is likely to reflect the general paucity and/or insufficiency of validated datapoints from DPR Korea and Jilin. Therefore, the disjunction of EOO estimated from Maxent binary presence (Figure 4) is likely to change with additional sampling of the species.
If our analyses were used to update the species account on the IUCN Red List, this would not be a genuine improvement in terms of the threats to the species, despite a much broader range than earlier described, as the threats have not changed, nor stabilized, and deforestation is still a primary driver of population decline in the species. In addition, forest cover is likely to keep on decreasing in all three nations (https://www.globalforestwatch.org/; 24 December 2020), noting the strong renewed policies for forest conservation in DPR Korea [63]. It is however important to update the status of the species on a flagship database such as the IUCN Red List of Threatened Species, as erroneous assessments are likely to trigger mistrust from users, and create a bias against its use by decision makers [64]. We recommend the use of the 2019 assessment as a national assessment for PR China.
Finally, our results highlight that the principal need for the species is to be understood through detailed ecological studies, as its cryptic nature may make it more threatened than estimated. For instance, the model projected over the entire study area estimated a broad area of high suitability (>0.58) across Jilin province although there is no GPS coordinate to indicate the presence of S. huanrenensis in the province. Similarly, the species is likely to be widespread along mountainous regions of DPR Korea. This likely indicates the potential for the discovery of unknown populations, pending additional survey efforts. Areas of high habitat suitability were also predicted for the southwestern part of R Korea where several major mountains are located, such as Jiri Mountain (>1000 m a.s.l). However, the species has not been recorded in this region so far, and our model predicted a “gap” of habitat suitability between the southwestern regions of R Korea with no verified occurrences and the eastern regions of R Korea with several verified occurrences (Figure 3B). Further field validation of our models will be needed to investigate the potential presence of the species in the southwestern regions of R Korea, or the presence of geographic barriers that might have created an environmentally suitable, but unreachable, habitat for the species.
5. Conclusions
In conclusion, we estimate that Scincella huanrenensis is not threatened at the global scale, falling in the Least Concern category following the guidelines of the IUCN Red List of Threatened Species, despite an observed or expected decline at most sites where the species is present. We however bring attention to the populations in PR China, which are threatened with extinction, and for which the 2019 IUCN Red List assessment is fitting. In addition, we highlight that the exact range of the species is not known and investigation in the form of field surveys is needed to clarify the exact distribution of the species. With the current data available, the protection of the species would best be implemented through the protection of forests at the adequate elevation (400–1500 m a.s.l.).
Supplementary Materials
See the Supplementary Figures and Table available https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/conservation1010005/s1 in a single file accompanying this article.
Author Contributions
Conceptualization, Y.S. and A.B.; Data curation, Y.S., K.R.M. and A.B.; Funding acquisition, A.B.; Investigation, A.B.; Methodology, Y.S., K.R.M. and A.B.; Project administration, A.B.; Resources, Y.S., K.R.M., K.S.K., S.C.L., M.H. and A.B.; Software, Y.S., K.R.M. and A.B.; Supervision, A.B.; Validation, Y.S., K.R.M., K.S.K., S.C.L., M.H. and A.B.; Visualization, Y.S., K.R.M., K.S.K., S.C.L., M.H. and A.B.; Writing—original draft, Y.S. and A.B.; Writing—review and editing, Y.S., K.R.M., K.S.K., S.C.L., M.H. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data and/or data sources used for this study can be found in the text, tables and Supplementary Materials.
Acknowledgments
This work is in no way a critic of the assessment provided for Scincella huanrenensis on The IUCN Red List of Species, but a praise to the integrity of the system and only aims at providing the data needed for an update.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wren, S.; Angulo, A.; Meredith, H.; Kielgast, J.; Dos Santos, M.; Bishop, P.; IUCN SSC Amphibian Specialist Group. Amphibian Conservation Action Plan. 2015. Available online: http://www.amphibians.org/acap/ (accessed on 12 December 2020).
- Rodrigues, A.S.; Pilgrim, J.D.; Lamoreux, J.F.; Hoffmann, M.; Brooks, T.M. The value of the IUCN Red List for conservation. Trends Ecol. Evol. 2006, 21, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Al Dhaheri, S.; Bennett, E.L.; Biggs, D.; Bignell, A.; Byers, O.; Cooney, R.; Donaldson, J.; Dublin, H.T.; Eggermont, H.; et al. IUCN’s encounter with 007: Safeguarding consensus for conservation. Oryx 2019, 53, 741–747. [Google Scholar] [CrossRef]
- Rondinini, C.; Di Marco, M.; Visconti, P.; Butchart, S.H.; Boitani, L. Update or Outdate: Long-Term Viability of the IUCN Red List. Conserv. Lett. 2013, 7, 126–130. [Google Scholar] [CrossRef]
- Betts, J.; Young, R.P.; Hilton-Taylor, C.; Hoffmann, M.; Rodríguez, J.P.; Stuart, S.N.; Milner-Gulland, E. A framework for evaluating the impact of the IUCN Red List of threatened species. Conserv. Biol. 2019, 34, 632–643. [Google Scholar] [CrossRef]
- Canseco-Márquez, L.; Ramírez-González, C.G.; González-Bernal, E. Discovery of another new species of Charadrahyla (Anura, Hylidae) from the cloud forest of northern Oaxaca, México. Zootaxa 2017, 4329, 64–72. [Google Scholar] [CrossRef] [PubMed]
- IUCN SSC Amphibian Specialist Group. Sarcohyla celata. In The IUCN Red List of Threatened Species; IUCN SSC Amphibian Specialist Group: Gland, Switzerland, 2020; Volume T55438A53953583. [Google Scholar]
- Broderick, A.C.; Frauenstein, R.; Glen, F.; Hays, G.C.; Jackson, A.L.; Pelembe, T.; Ruxton, G.D.; Godley, B.J. Are green turtles globally endangered? Glob. Ecol. Biogeogr. 2006, 15, 21–26. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Shanker, K. Marine turtles and IUCN Red Listing: A review of the process, the pitfalls, and novel assessment approaches. J. Exp. Mar. Biol. Ecol. 2008, 356, 52–68. [Google Scholar] [CrossRef]
- Zhao, W.; Li, P. Scincella huanrenensis. In The IUCN Red List of Threatened Species; IUCN SSC Amphibian Specialist Group: Gland, Switzerland, 2019. [Google Scholar]
- NIBR. Red Data Book of Republic of Korea. In Volume 2: Amphibians and Reptiles; National Institute of Biological Resources, Ministry of Environment: Incheon, Korea, 2019. [Google Scholar]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 14. Prepared by the Standards and Petitions Committee. 2019. Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 15 February 2021).
- Zhao, E.; Huang, K. A survey of amphibians and reptiles in Liaoning province. Acta Herpetol. Sin. 1982, 1, 1–23. [Google Scholar]
- Dong, B.-J.; Wang, J.-Q.; Zhou, Z.-Y.; Lu, Y.-Y.; Li, P.-P. Research on reproductive behavior and strategy of Scincella huanrensis. Sichuan J. Zool. 2007, 26, 302–304. [Google Scholar]
- Lee, S.-C. Systematic and Ecological Studies of the Suborder Sauria (Reptilia, Squamata) in Korea; University of Incheon: Incheon, Korea, 2010. [Google Scholar]
- Chang, M.; Song, J.; Lee, J.; Oh, H. The current status of Korean lizards (Reptilia: Squamata). Korean J. Environ. Ecol. 2006, 20, 352–358. [Google Scholar]
- Song, J.Y. Current status and distribution of reptiles in the Republic of Korea. Korean J. Environ. Biol. 2007, 25, 124–138. [Google Scholar]
- Koo, K.S.; Kwon, S.-R.; Chang, M.-H.; Song, J.-Y. New geographic distribution record of the rare skink Scincella huanrenensis Zhao and Huang, 1982 (Squamata, Scincidae). Herpetol. Notes 2020, 13, 199–201. [Google Scholar]
- Zhao, E.-M.; Zhao, K.-T.; Zhou, K.-Y. Fauna Sinica, Reptilia Vol 2, Squamata, Lacertilia; Science Press: Beijing, China, 1999. [Google Scholar]
- Won, H.K. Amphibian and Reptilian fauna of Korea; Academy of Sciences: Pyongyang, Democratic People’s Republic of Korea, 1971; p. 170. [Google Scholar]
- Kim, L.; Han, G. Chosun Animal Encyclopedia, Herpetology Volume; Roh, H., Ed.; Science and Technology Publisher: Pyongyang, Democratic People’s Republic of Korea, 2009; p. 138. [Google Scholar]
- Dong, B.-J. Current status and protection of Scincella huanrenensis. Sichuan J. Zool. 2005, 24, 279–281. [Google Scholar]
- Shin, Y.; Jang, Y.; Allain, S.J.R.; Borzée, A. Catalogue of herpetological specimens of the Ewha Womans University Natural History Museum (EWNHM), Republic of Korea. ZooKeys 2020, 965, 103–139. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Koo, K.-S.; Kim, I.-H.; Park, D. Complete mitochondrial genomes of Scincella vandenburghi and S. huanrenensis (Squamata: Scincidae). Mitochondrial DNA Part B 2016, 1, 237–238. [Google Scholar] [CrossRef]
- Kim, H.W.; Yoon, S.; Kim, M.; Shin, M.; Yoon, H.; Kim, K. EcoBank: A flexible database platform for sharing ecological data. Biodivers. Data J. 2021, 9, e61866. [Google Scholar] [CrossRef]
- Bobrov, V.V. North, west, and east limits of the Sahara-Gobi faunistic region in Eurasia (according to distribution of lizards (reptilia, sauria). Biol. Bull. 1999, 26, 469–478. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Tuanmu, M.-N.; Jetz, W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Glob. Ecol. Biogeogr. 2014, 23, 1031–1045. [Google Scholar] [CrossRef]
- Soberon, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Ofori, B.Y.; Stow, A.J.; Baumgartner, J.B.; Beaumont, L.J. Combining dispersal, landscape connectivity and habitat suitability to assess climate-induced changes in the distribution of Cunningham’s skink, Egernia cunninghami. PLoS ONE 2017, 12, e0184193. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R foundation for statistical computing: Vienna, Austria, 2020. [Google Scholar]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.0-7. 2019. Available online: https://CRAN.R-project.org/package=raster (accessed on 23 February 2021).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ’Geospatial’ Data Abstraction Library. R Package Version 1.4-8. 2019. Available online: https://CRAN.R-project.org/package=rgdal (accessed on 23 February 2021).
- Bivand, R.; Rundell, C. Rgeos: Interface to Geometry Engine—Open Source (’GEOS’). R Package Version 0.5-2. 2019. Available online: https://CRAN.R-project.org/package=rgeos (accessed on 23 February 2021).
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Peterson, A.T.; Falconi, M.; Contreras-Díaz, R.G.; Martínez-Meyer, E.; Barve, V.; Barve, N. Ntbox: An R package with graphical user interface for modelling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. R Package Version 1.1-4. 2017. Available online: https://CRAN.R-project.org/package=dismo (accessed on 23 February 2021).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Biomod2: Ensemble Platform for Species Distribution Modeling. R Package Version 3.4.6. 2020. Available online: https://CRAN.R-project.org/package=biomod2 (accessed on 23 February 2021).
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Dauby, G. ConR: Computation of Parameters Used in Preliminary Assessment of Conservation Status. R Package Version 1.3.0. 2020. Available online: https://CRAN.R-project.org/package=ConR (accessed on 23 February 2021).
- R Core Team. R Version 3.6.3; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 23 February 2021).
- Maes, D.; Isaac, N.J.B.; Harrower, C.A.; Collen, B.; Van Strien, A.J.; Roy, D.B. The use of opportunistic data for IUCN Red List assessments. Biol. J. Linn. Soc. 2015, 115, 690–706. [Google Scholar] [CrossRef]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akcakaya, H.T.; Leader-Williams, N.; Miller-Gulland, E.J.; Stuartt, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef]
- Mace, G.M.; Kunin, W. Classifying threatened species: Means and ends. Philos. Trans. R. Soc. B Biol. Sci. 1994, 344, 91–97. [Google Scholar] [CrossRef]
- Hermoso, V.; Kennard, M.J.; Linke, S. Evaluating the costs and benefits of systematic data acquisition for conservation assessments. Ecography 2014, 38, 283–292. [Google Scholar] [CrossRef]
- Macias, D.; Shin, Y.; Borzée, A. An update on the conservation status and ecology of Korean terrestrial squamates. J. Nat. Conserv. 2021, 60, 125971. [Google Scholar] [CrossRef]
- Yang, C.; Tang, S.; Luo, Z. Distribution changes of Chinese skink (Eumeces chinensis) in China: The impacts of global climate change. Asian Herpetol. Res. 2020, 11, 132–139. [Google Scholar]
- COP21. Paris Climate Agreement; United Nations: Paris, France, 2015; Chapter 27. [Google Scholar]
- Mundaca, L.; Ürge-Vorsatz, D.; Wilson, C. Demand-side approaches for limiting global warming to 1.5 °C. Energy Effic. 2019, 12, 343–362. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Moss, R.H.; Babkier, M.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.A.; Elgizouli, I.; Emori, S.; Lin, E.; Hibbbard, L.; et al. Towards New Scenarios for the Analysis of Emissions: Climate Change, Impacts and Response Strategies; Intergovernmental Panel on Climate Change, Ed.; Intergovernmental Panel on Climate Change Secretariat (IPCC): Noordwijkerhout, The Netherlands, 2008. [Google Scholar]
- Xu, Y.; Zhou, B.T.; Wu, J.; Han, Z.Y.; Zhang, Y.X.; Wu, J. Asian climate change under 1.5–4 C warming targets. Adv. Clim. Chang. Res. 2017, 8, 99–107. [Google Scholar] [CrossRef]
- Kim, I.H.; Jang, H.J.; Roh, N.H.; Park, D.S. Endangered risk analysis of Korean three rare reptile species. In Proceedings of the 8th Annual Meeting of the Society of Korean Herpetologists, Wonju, Korea, 10 July 2015. [Google Scholar]
- Stone, R. Seeking Cures for North Korea’s Environmental Ills. Science 2012, 335, 1425–1426. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-D.; Miller-Rushing, A.J. Degradation, urbanization, and restoration: A review of the challenges and future of conservation on the Korean Peninsula. Biol. Conserv. 2014, 176, 262–276. [Google Scholar] [CrossRef]
- Jang, H.J.; Kim, D.I.; Jang, M.H. Distribution of reptiles in South Korea—Based on the 3rd National Ecosystem Survey. Korean J. Herpetol. 2016, 7, 30–35. [Google Scholar]
- Aubry, K.B.; Raley, C.M.; McKelvey, K.S. The importance of data quality for generating reliable distribution models for rare, elusive, and cryptic species. PLoS ONE 2017, 12, e0179152. [Google Scholar] [CrossRef]
- Arrondo, E.; Moleón, M.; Cortés-Avizanda, A.; Jiménez, J.; Beja, P.; Sánchez-Zapata, J.A.; Donázar, J.A. Invisible barriers: Differential sanitary regulations constrain vulture movements across country borders. Biol. Conserv. 2018, 219, 46–52. [Google Scholar] [CrossRef]
- National Biodiversity Strategy and Action Plan. In National Biodiversity Strategy and Action Plan of DPR Korea; Gef, U. (Ed.) UNEP & GEF: Pyongyang, Democratic People’s Republic of Korea, 2017; Available online: http://extwprlegs1.fao.org/docs/pdf/prk163532.pdf (accessed on 12 February 2021).
- Cook, C.N.; Mascia, M.B.; Schwartz, M.W.; Possingham, H.P.; Fuller, R.A. Achieving conservation science that bridges the knowledge–action boundary. Conserv. Biol. 2013, 27, 669–678. [Google Scholar] [CrossRef]
Figure 1.
Paratype and live Scincella huanrenensis. The plate for the paratype (DMNH810014, female) is extracted from the literature [9] and the live individual was photographed in Odaesan National Park in September 2019.
Figure 1.
Paratype and live Scincella huanrenensis. The plate for the paratype (DMNH810014, female) is extracted from the literature [9] and the live individual was photographed in Odaesan National Park in September 2019.

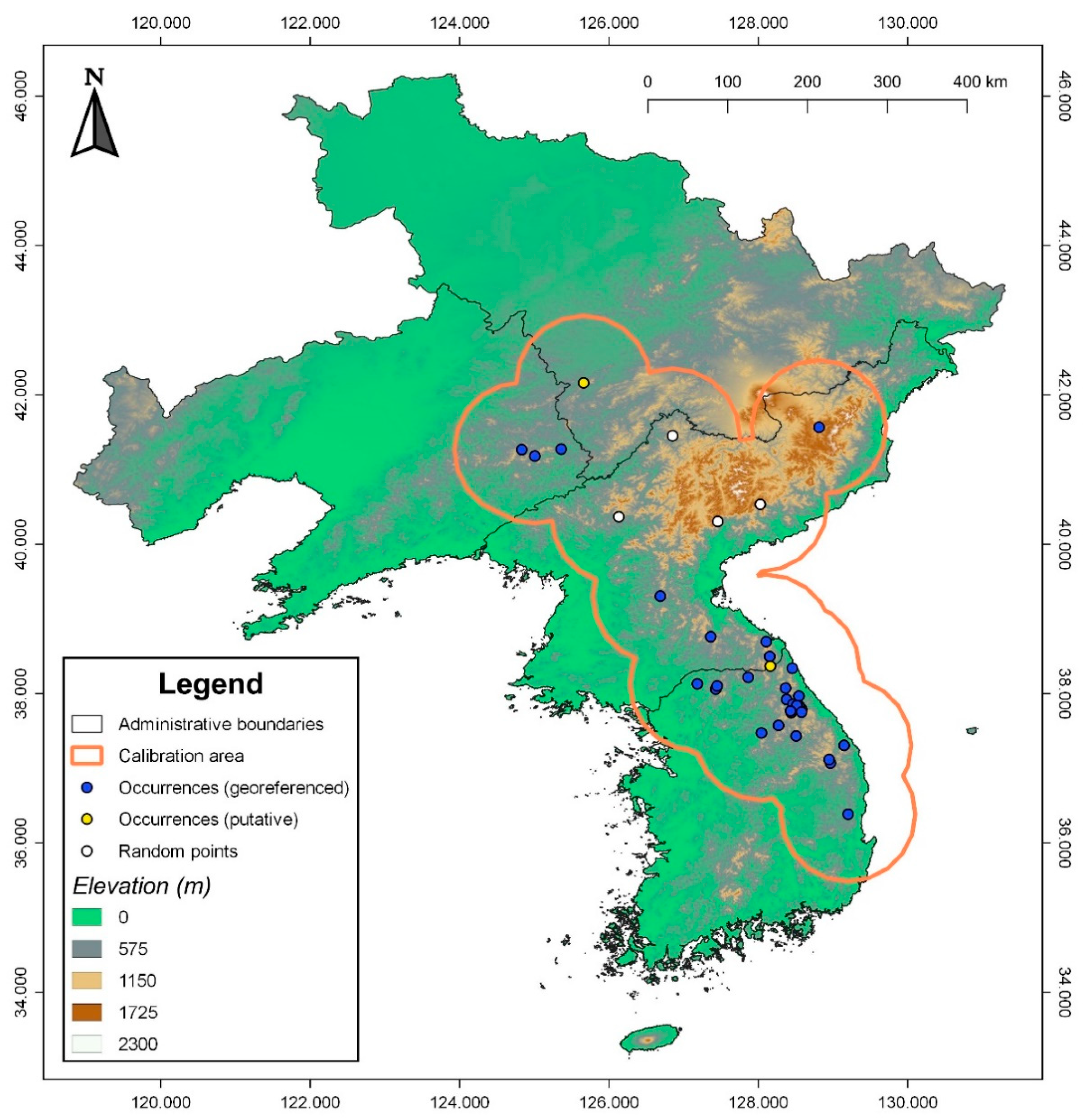
Figure 2.
The study area and model calibration area used in this study. The blue dots represent georeferenced occurrence points used for modelling, yellow dots represent putative datapoints that were only used to generate a density raster for bias correction, and the white dots are randomly generated points between georeferenced and putative datapoints between the Democratic People’s Republic of Korea, Liaoning, and Jilin to define a non-disjunct model calibration area. All downstream analyses on the extent of occurrence and threat status assessments were based on this calibration area.
Figure 2.
The study area and model calibration area used in this study. The blue dots represent georeferenced occurrence points used for modelling, yellow dots represent putative datapoints that were only used to generate a density raster for bias correction, and the white dots are randomly generated points between georeferenced and putative datapoints between the Democratic People’s Republic of Korea, Liaoning, and Jilin to define a non-disjunct model calibration area. All downstream analyses on the extent of occurrence and threat status assessments were based on this calibration area.

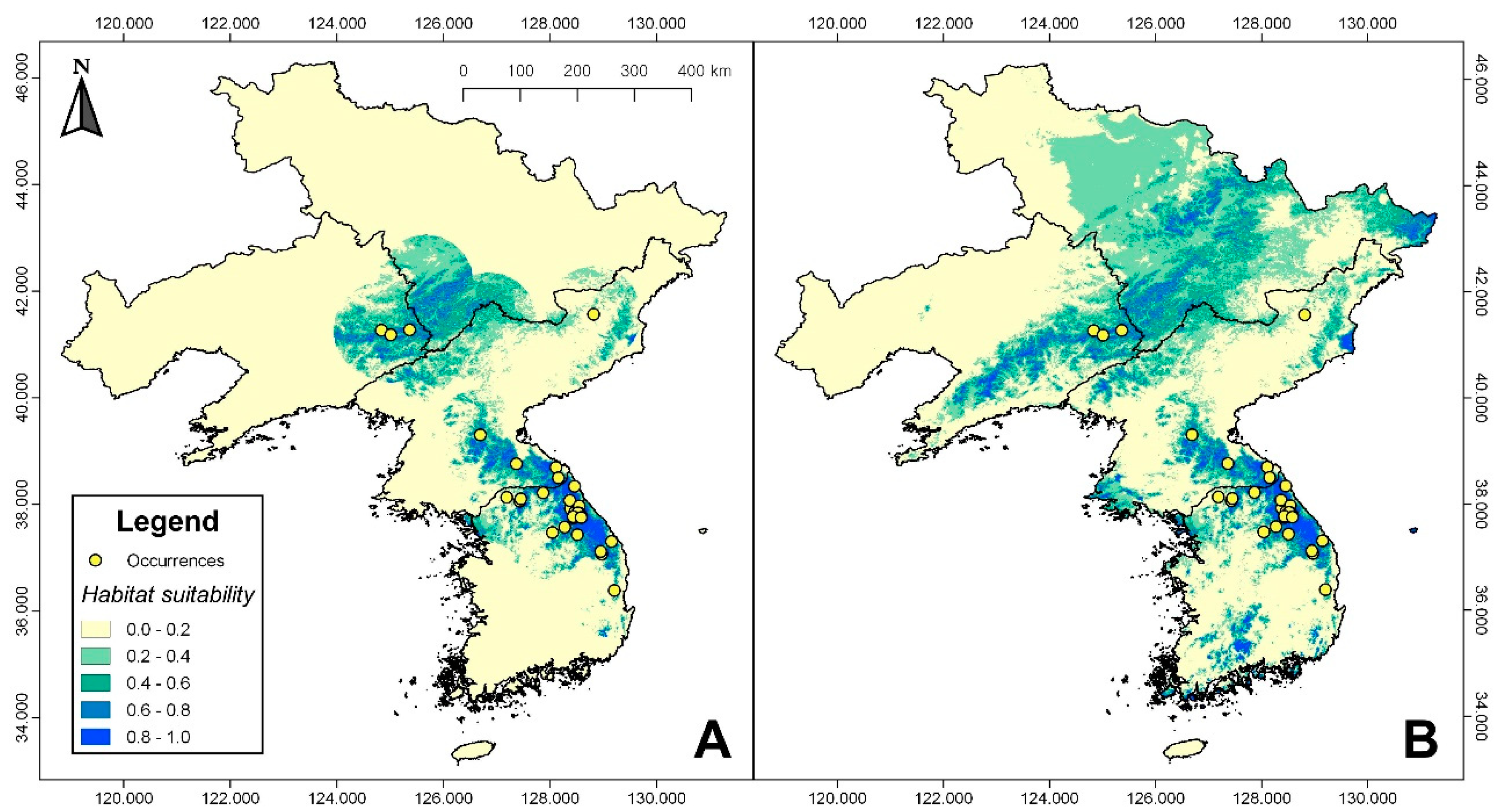
Figure 3.
Maxent habitat suitability models (area under the receiver operating characteristic curve (AUC) = 0.85 ± 0.06; True Skill Statistic (TSS) = 0.63 ± 0.03) generated within the calibration area (A) and projected to the entire study area (B). The species is restricted to the Baekdu Mountain Range, with areas of high suitability (>0.58) mostly located in eastern Liaoning province, western Jilin province, and eastern Korean Peninsula. The projected model indicates a considerably broad area of suitable, but unsampled habitat suggests the potential existence of new populations.
Figure 3.
Maxent habitat suitability models (area under the receiver operating characteristic curve (AUC) = 0.85 ± 0.06; True Skill Statistic (TSS) = 0.63 ± 0.03) generated within the calibration area (A) and projected to the entire study area (B). The species is restricted to the Baekdu Mountain Range, with areas of high suitability (>0.58) mostly located in eastern Liaoning province, western Jilin province, and eastern Korean Peninsula. The projected model indicates a considerably broad area of suitable, but unsampled habitat suggests the potential existence of new populations.
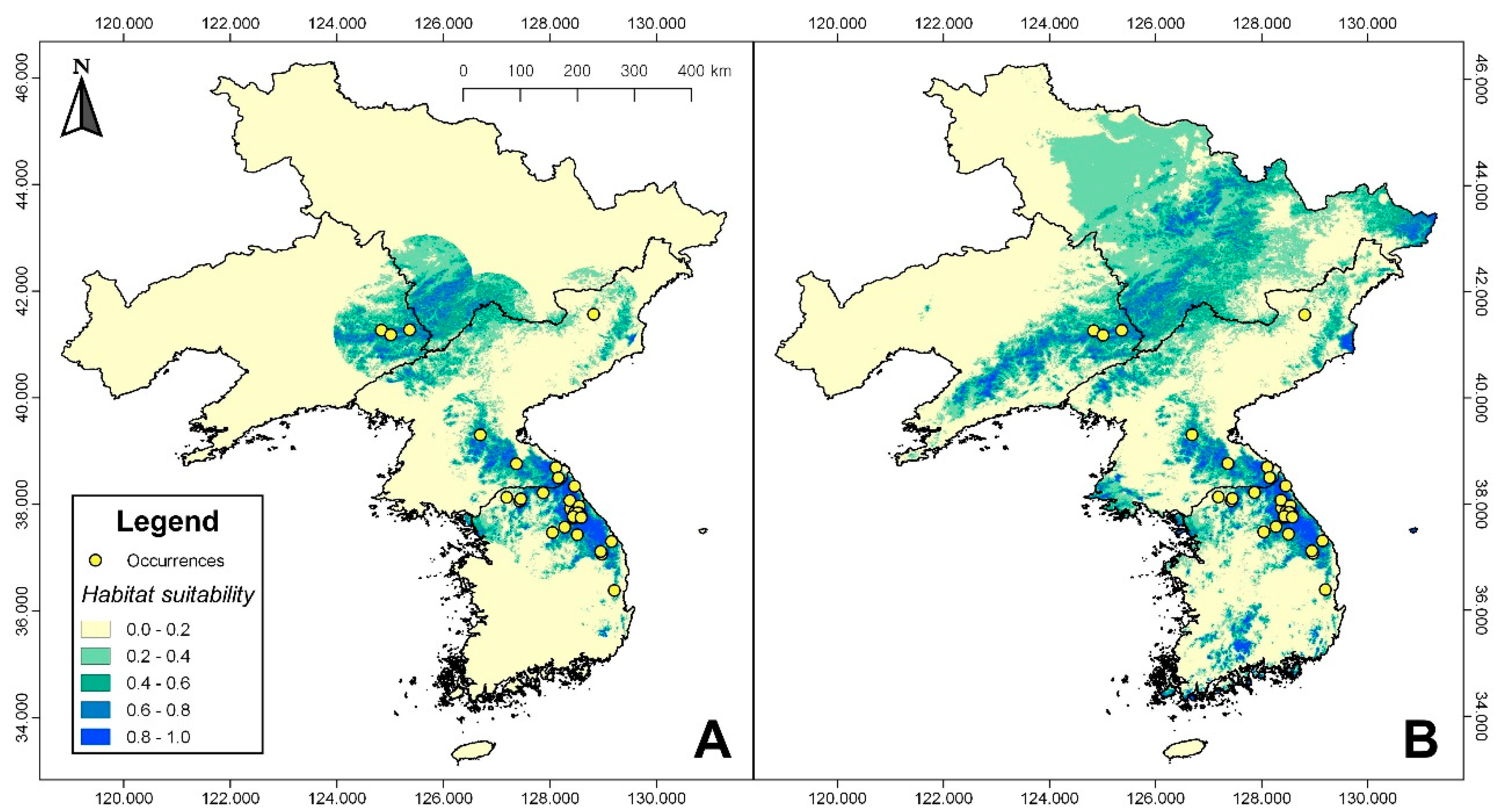
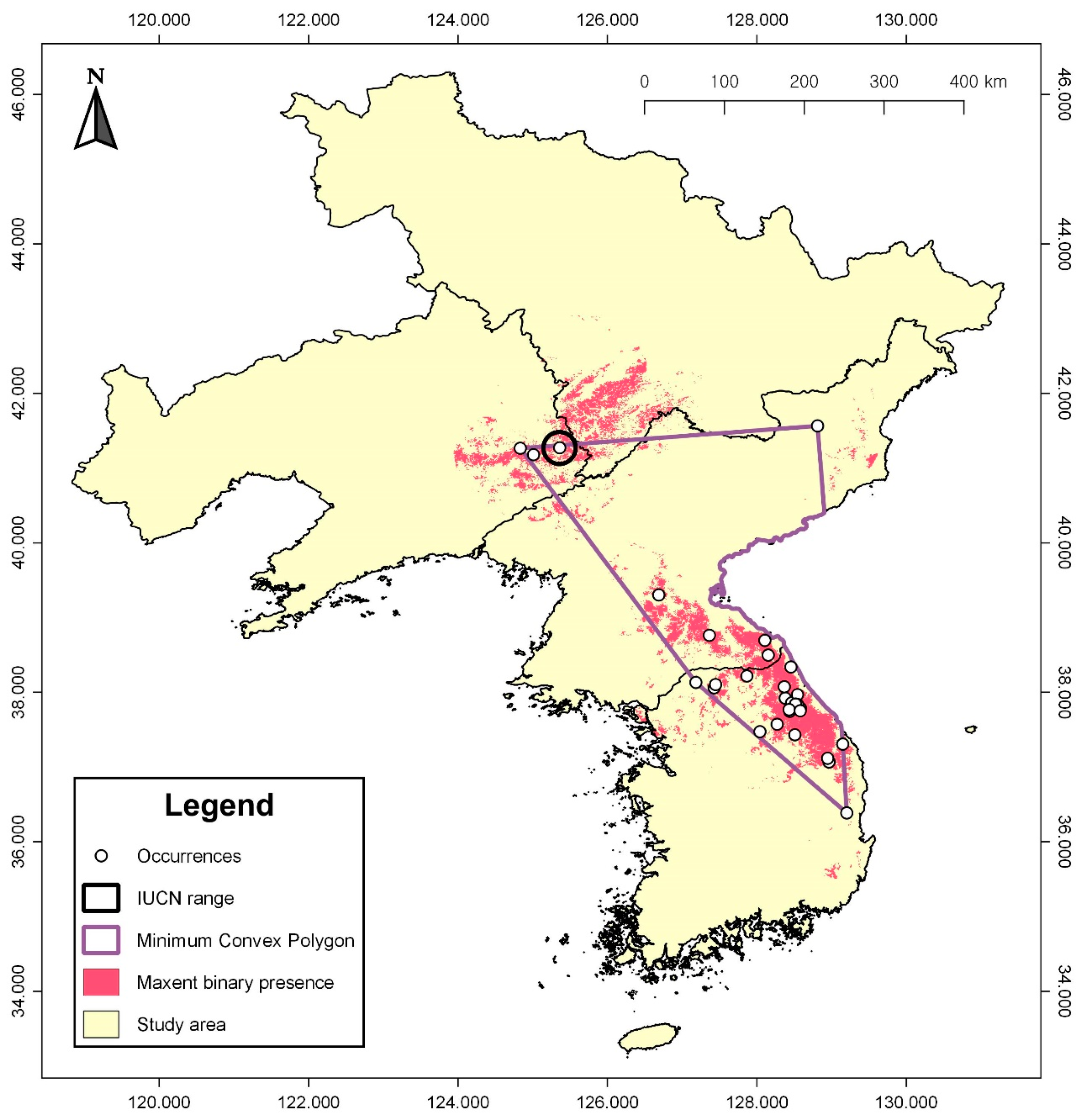
Figure 4.
Ranges for Scincella huanrenensis. The range shaded in red was computed from a minimum convex polygon (MCP) restricted to land area and the Maxent binary presence map. The range size estimated from the MCP (87,648.1 km2) was considerably broader than the one based on Maxent binary map (26,721.9 km2). This discrepancy is likely due to poor sampling in Jilin and Democratic People’s Republic of Korea. Also, the range “gap” between Liaoning-Jilin and the Democratic People’s Republic of Korea likely reflects poor sampling in these regions.
Figure 4.
Ranges for Scincella huanrenensis. The range shaded in red was computed from a minimum convex polygon (MCP) restricted to land area and the Maxent binary presence map. The range size estimated from the MCP (87,648.1 km2) was considerably broader than the one based on Maxent binary map (26,721.9 km2). This discrepancy is likely due to poor sampling in Jilin and Democratic People’s Republic of Korea. Also, the range “gap” between Liaoning-Jilin and the Democratic People’s Republic of Korea likely reflects poor sampling in these regions.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Occurrence dataset used for the analyses. Each coordinate is a known point for the presence of Scincella huanrenensis. Occurrence points with asterisks (*) represent putative datapoints used to create a bias layer but not used in the habitat suitability model itself. The voucher numbers for “EWNHM-ANIMAL” 6493, 6494, 6495, and 6496 are historical specimens collected from a single location and deposited at the Ewha Womans University Natural History Museum (EWNHM). “Workshop” is a Pers. Comm. at a workshop on amphibian conservation in the Democratic People’s Republic of Korea. NIE stands for the 4th National Ecosystem Survey conducted by the National Institute of Ecology in the Republic of Korea. Links and relevant references for the NIE, iNaturalist and GBIF datasets are provided in the text. All occurrence points have been rounded to two digits to decrease the risk of poaching, and each set of coordinates is different, although not necessarily visible in this table due to rounding.
Table 1.
Occurrence dataset used for the analyses. Each coordinate is a known point for the presence of Scincella huanrenensis. Occurrence points with asterisks (*) represent putative datapoints used to create a bias layer but not used in the habitat suitability model itself. The voucher numbers for “EWNHM-ANIMAL” 6493, 6494, 6495, and 6496 are historical specimens collected from a single location and deposited at the Ewha Womans University Natural History Museum (EWNHM). “Workshop” is a Pers. Comm. at a workshop on amphibian conservation in the Democratic People’s Republic of Korea. NIE stands for the 4th National Ecosystem Survey conducted by the National Institute of Ecology in the Republic of Korea. Links and relevant references for the NIE, iNaturalist and GBIF datasets are provided in the text. All occurrence points have been rounded to two digits to decrease the risk of poaching, and each set of coordinates is different, although not necessarily visible in this table due to rounding.
| Origin | Latitude (° N) | Longitude (° E) |
|---|---|---|
| Field survey | 38.07 | 128.36 |
| Field survey | 37.43 | 128.50 |
| Field survey | 36.38 | 129.20 |
| Field survey | 37.83 | 128.54 |
| Field survey | 37.83 | 128.53 |
| Field survey | 36.38 | 129.20 |
| Field survey | 37.76 | 128.57 |
| Field survey | 37.92 | 128.37 |
| Field survey | 38.13 | 127.18 |
| Field survey | 38.06 | 127.42 |
| Field survey | 38.10 | 127.44 |
| Field survey | 37.06 | 128.96 |
| Field survey | 37.74 | 128.43 |
| Field survey | 37.76 | 128.57 |
| Field survey | 37.85 | 128.46 |
| Field survey | 37.11 | 128.94 |
| Field survey | 36.38 | 129.20 |
| Field survey | 38.07 | 128.36 |
| Field survey | 37.57 | 128.27 |
| Field survey | 37.78 | 128.58 |
| Field survey | 37.47 | 128.04 |
| EWNHM-ANIMAL | 37.96 | 128.54 |
| iNaturalist | 37.74 | 128.43 |
| iNaturalist | 37.83 | 128.54 |
| iNaturalist | 37.83 | 128.54 |
| iNaturalist | 37.84 | 128.51 |
| GBIF | 37.76 | 128.43 |
| GBIF | 37.75 | 128.58 |
| NIE | 38.21 | 127.86 |
| NIE | 38.33 | 128.45 |
| NIE | 37.30 | 129.14 |
| Dong 2005 | 41.26 | 124.83 |
| Zhao and Huang, 1982 | 41.18 | 125.01 |
| Dong 2008 | 41.26 | 124.83 |
| Bobrov 1999 * | 42.16 | 125.66 |
| Workshop * | 38.36 | 128.16 |
| Won 1971/Kim and Han 2009 | 38.76 | 127.36 |
| Won 1971/Kim and Han 2009 | 39.30 | 126.68 |
| Won 1971/Kim and Han 2009 | 41.56 | 128.81 |
| Won 1971/Kim and Han 2009 | 38.69 | 128.10 |
| Won 1971/Kim and Han 2009 | 38.49 | 128.15 |
| IUCN | 41.27 | 125.36 |
Table 2.
Environmental variables used to build the habitat suitability model of Scincella huanrenensis in Maxent. The contribution of each environmental variable to the model is listed in percentage of contribution and permutation importance.
Table 2.
Environmental variables used to build the habitat suitability model of Scincella huanrenensis in Maxent. The contribution of each environmental variable to the model is listed in percentage of contribution and permutation importance.
| Variables | Contribution (%) | Permutation Importance |
|---|---|---|
| Elevation | 27 | 30.3 |
| Isothermality (bio3) | 21.7 | 14.9 |
| Precipitation of driest quarter (bio17) | 21 | 41.7 |
| Deciduous broadleaf forest cover | 16.7 | 1.5 |
| Annual precipitation (bio12) | 7.9 | 9.5 |
| Mixed forest cover | 4.5 | 1.6 |
| Needleleaf forest cover | 1.1 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shin, Y.; Messenger, K.R.; Koo, K.S.; Lee, S.C.; Hou, M.; Borzée, A. How Threatened Is Scincella huanrenensis? An Update on Threats and Trends. Conservation 2021, 1, 58-72. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation1010005
AMA Style
Shin Y, Messenger KR, Koo KS, Lee SC, Hou M, Borzée A. How Threatened Is Scincella huanrenensis? An Update on Threats and Trends. Conservation. 2021; 1(1):58-72. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation1010005
Chicago/Turabian StyleShin, Yucheol, Kevin R. Messenger, Kyo Soung Koo, Sang Cheol Lee, Mian Hou, and Amaël Borzée. 2021. "How Threatened Is Scincella huanrenensis? An Update on Threats and Trends" Conservation 1, no. 1: 58-72. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation1010005