
Growing Tomato under Protected Cultivation Conditions: Overall Effects on Productivity, Nutritional Yield, and Pest Incidences




Abstract
:1. Introduction
2. Materials and Methods
2.1. Location
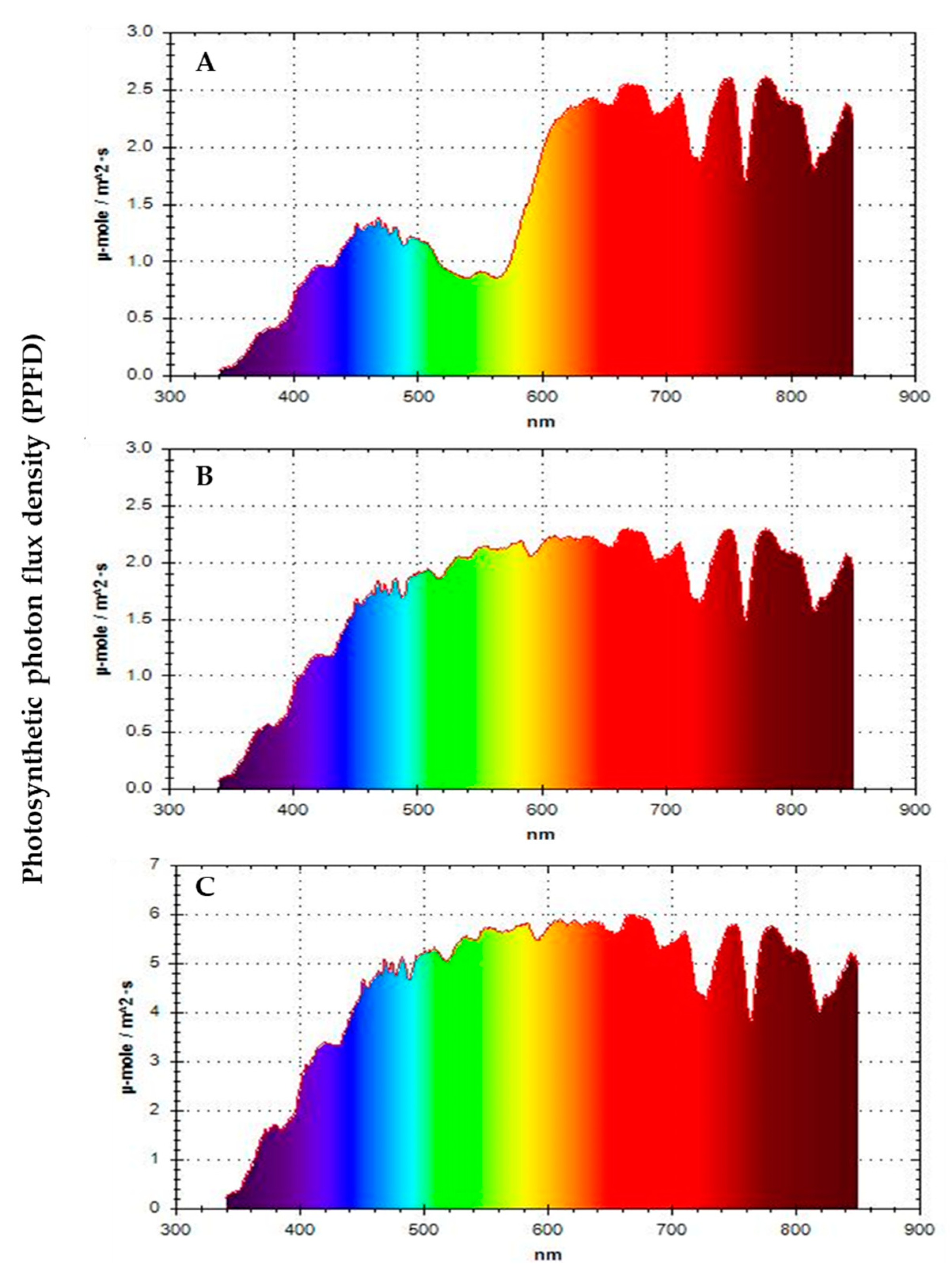
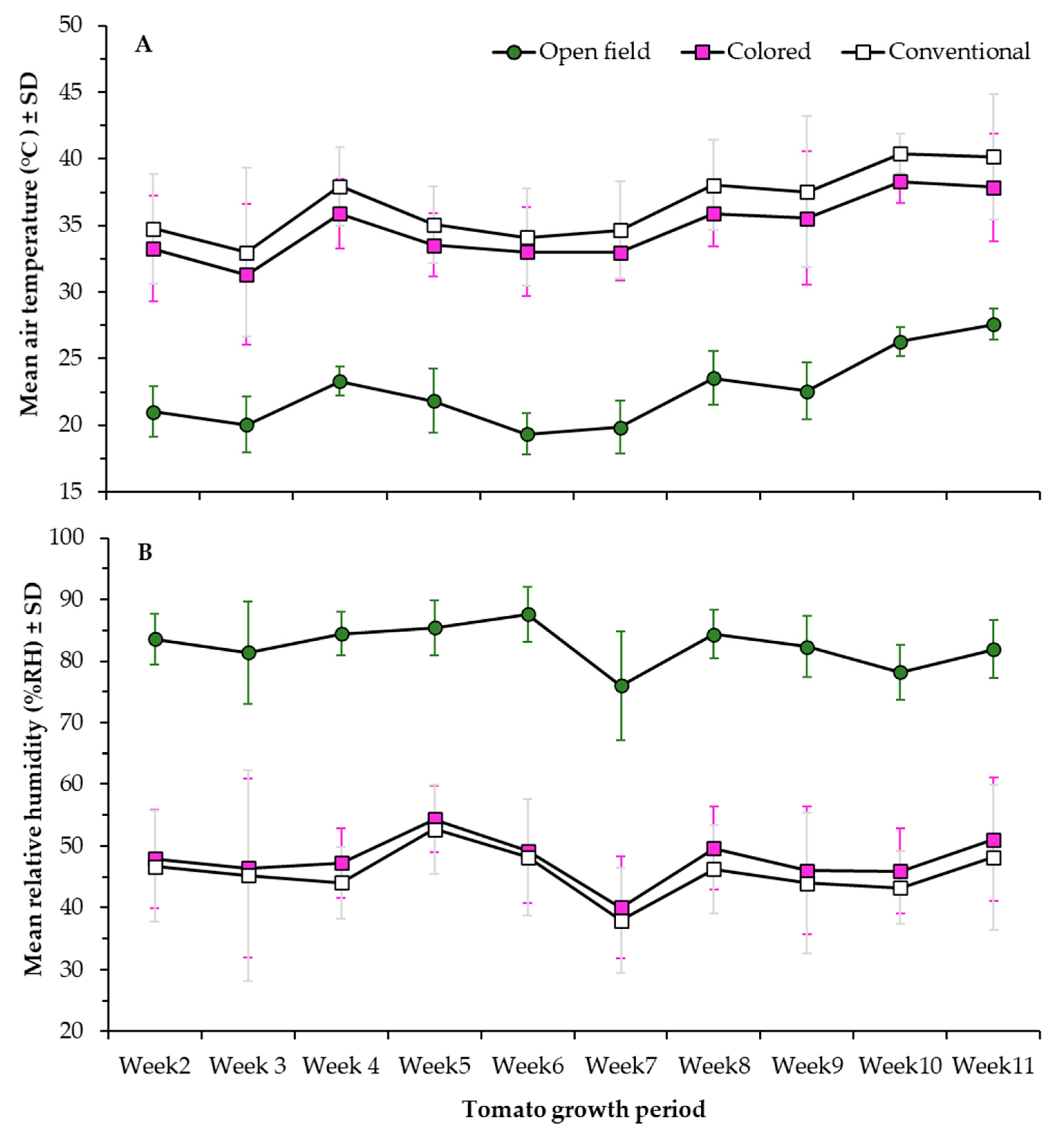
2.2. Treatments and Data Collection
2.3. Nutritional Analysis
2.3.1. Lycopene and β-Carotene
2.3.2. Vitamin C
2.4. Monitoring of Insect Pests
2.5. Data Analysis
3. Results
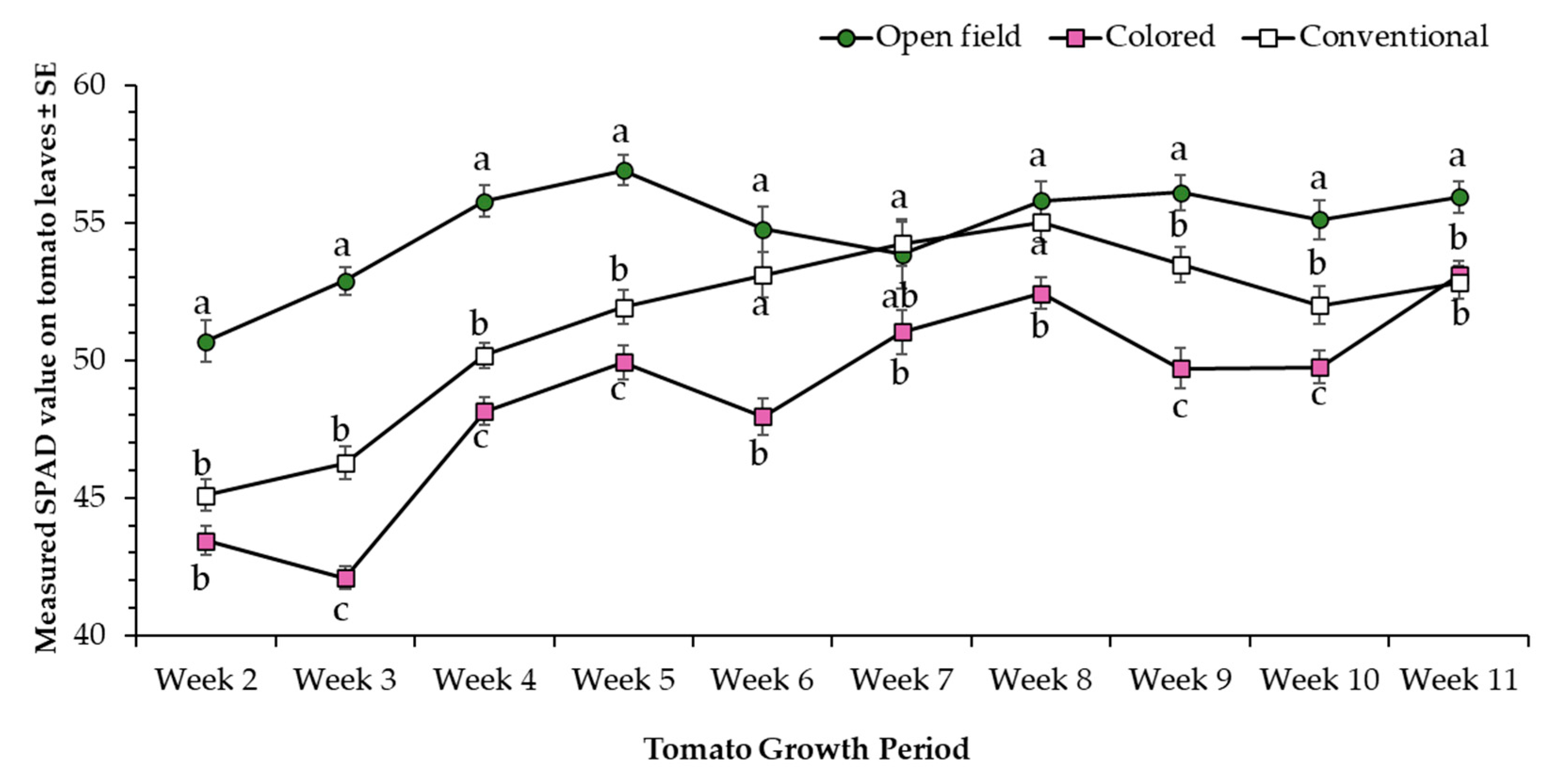
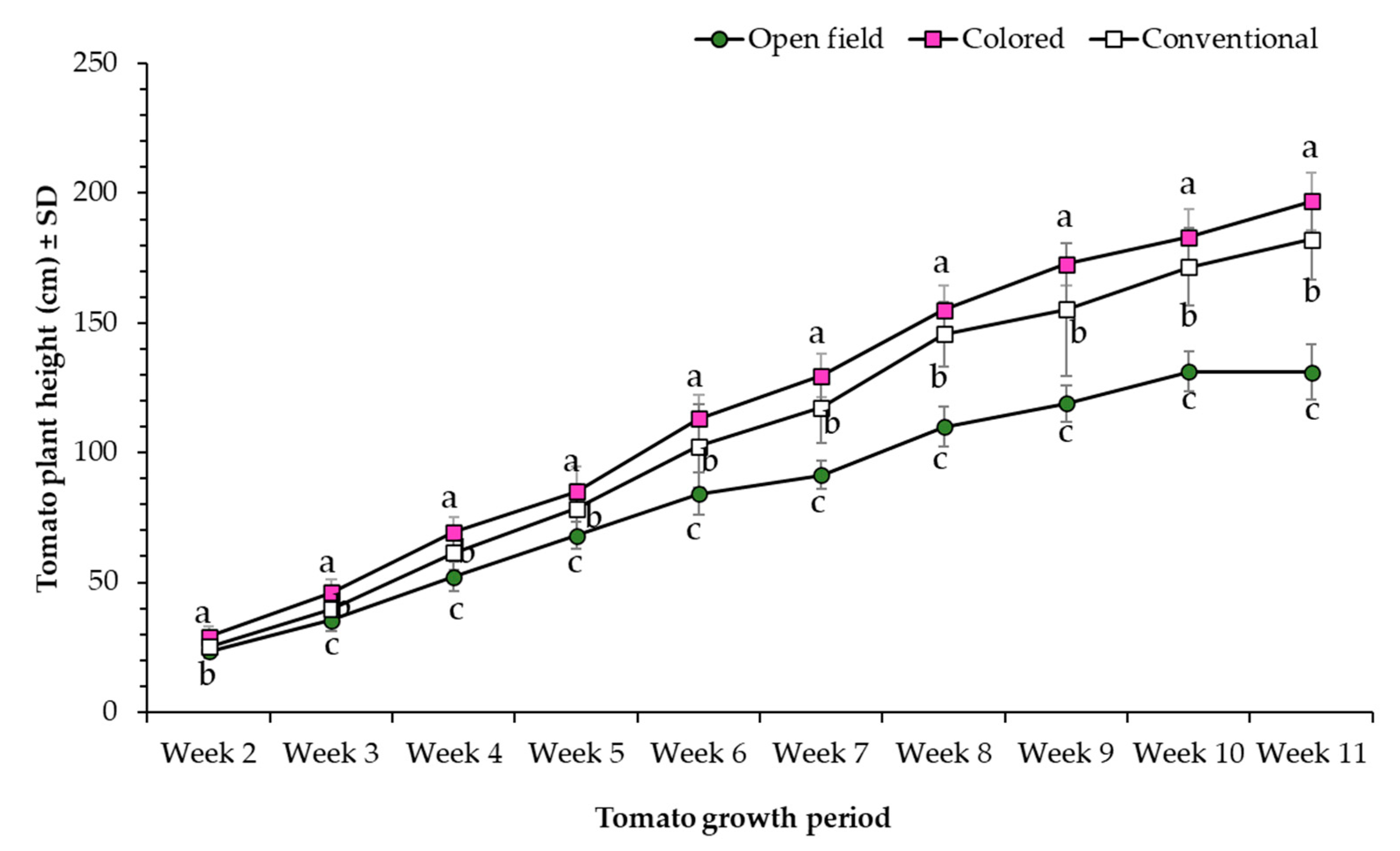
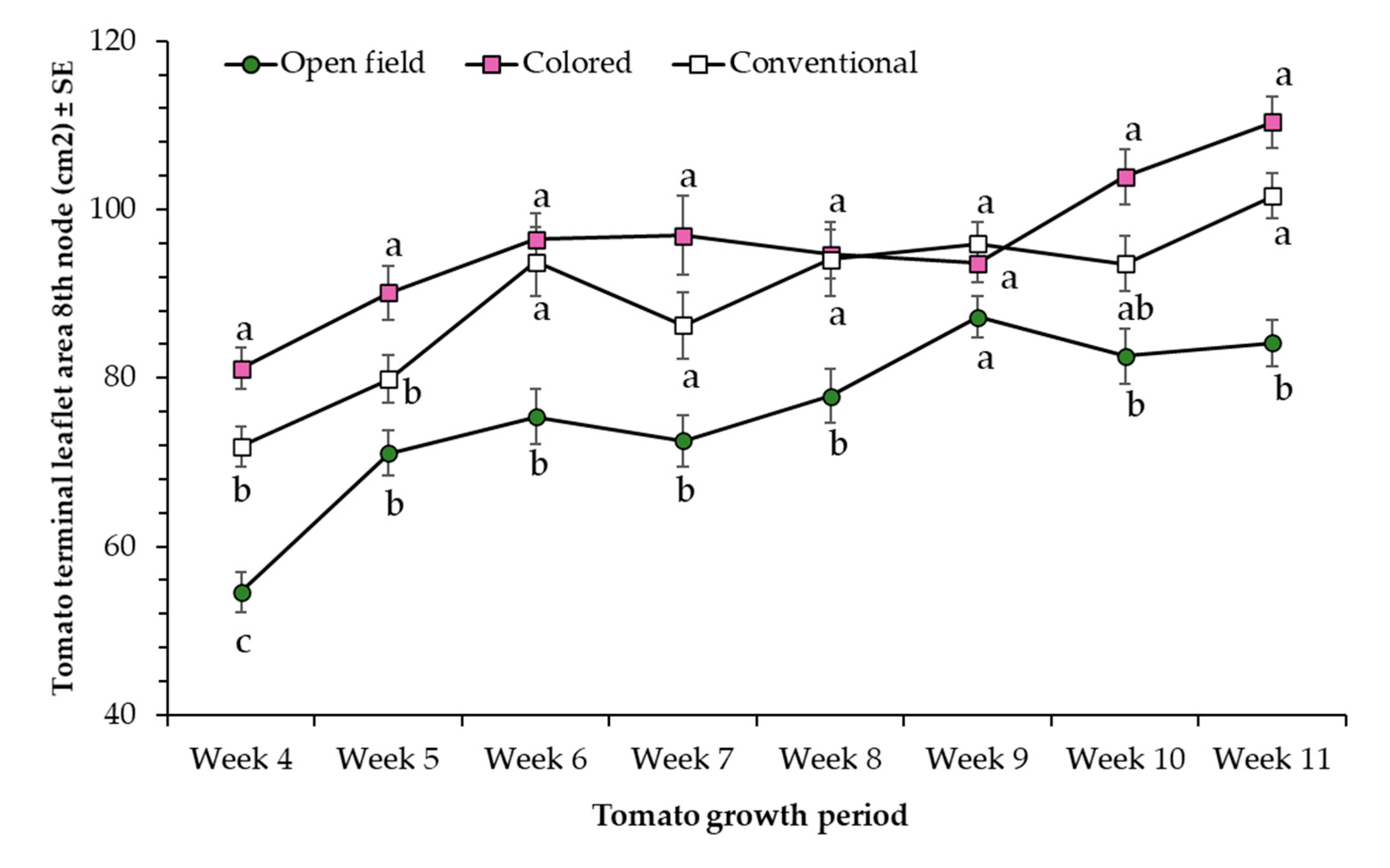
3.1. Morphometric Parameters
3.2. Productivity and Nutritional Yield
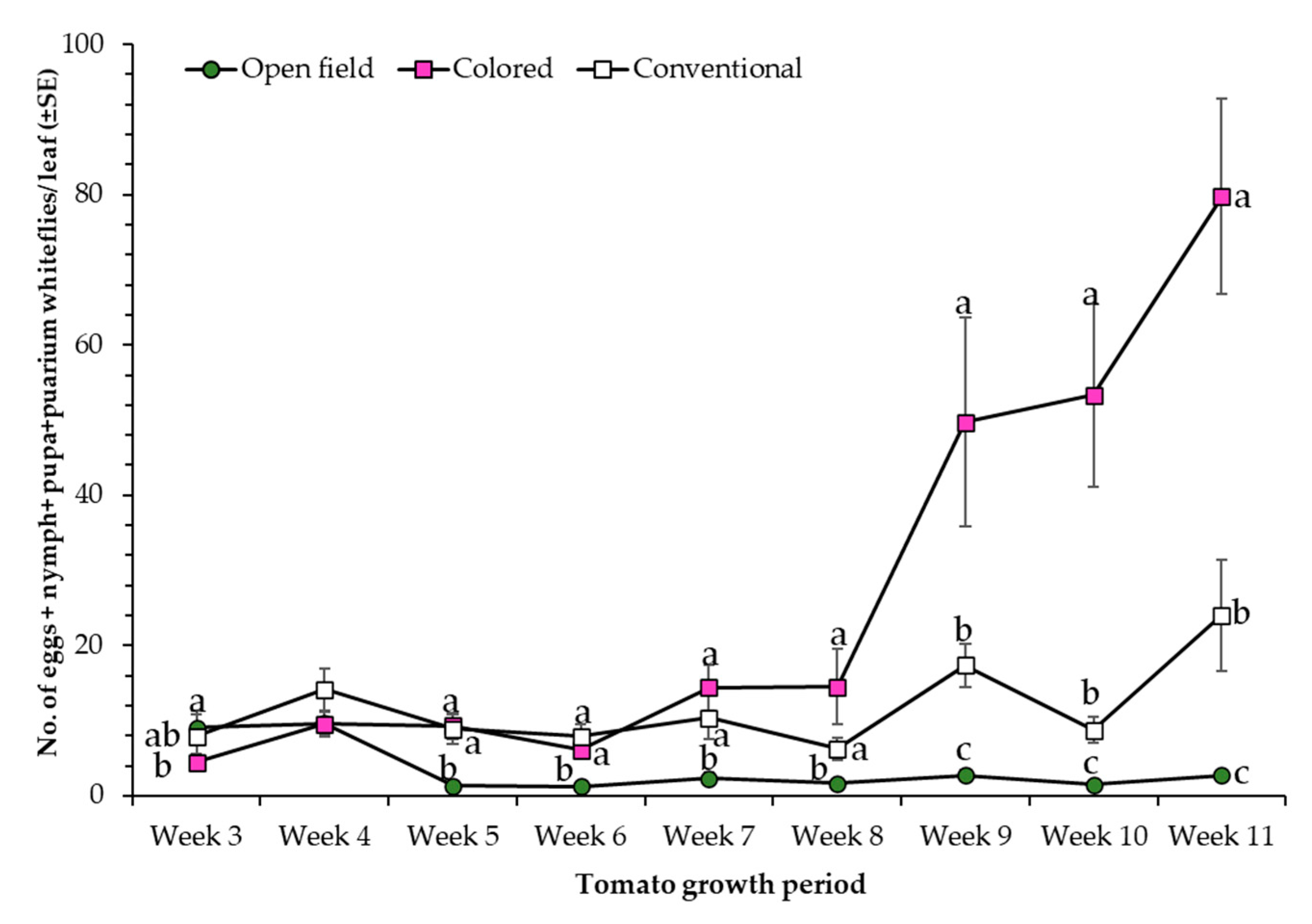
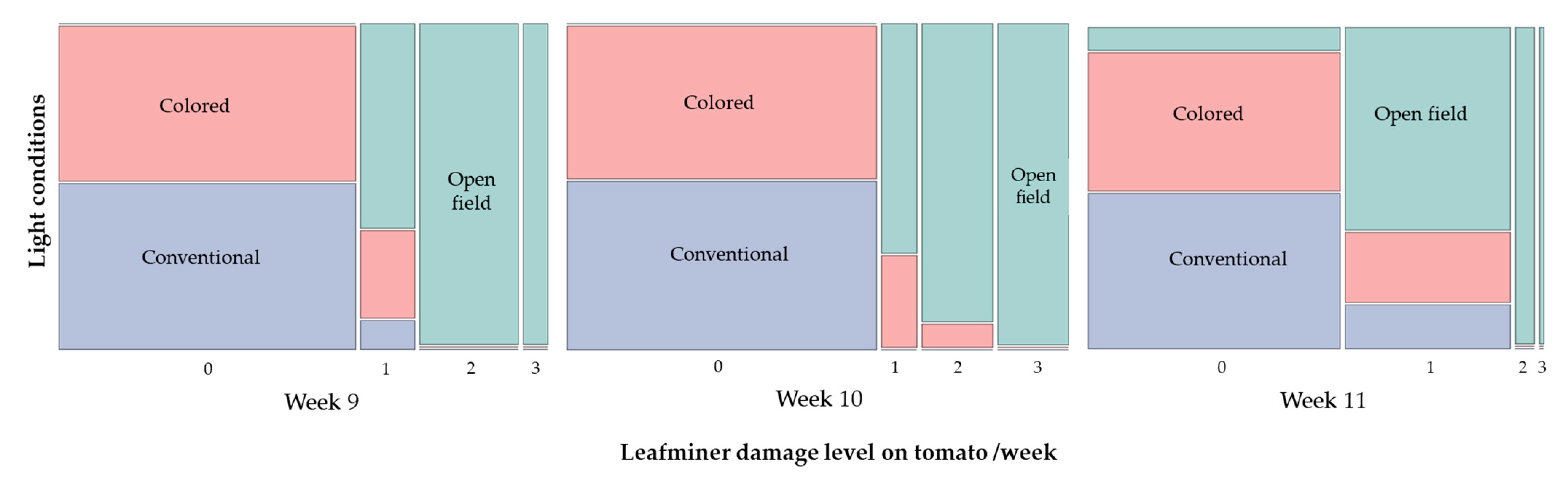
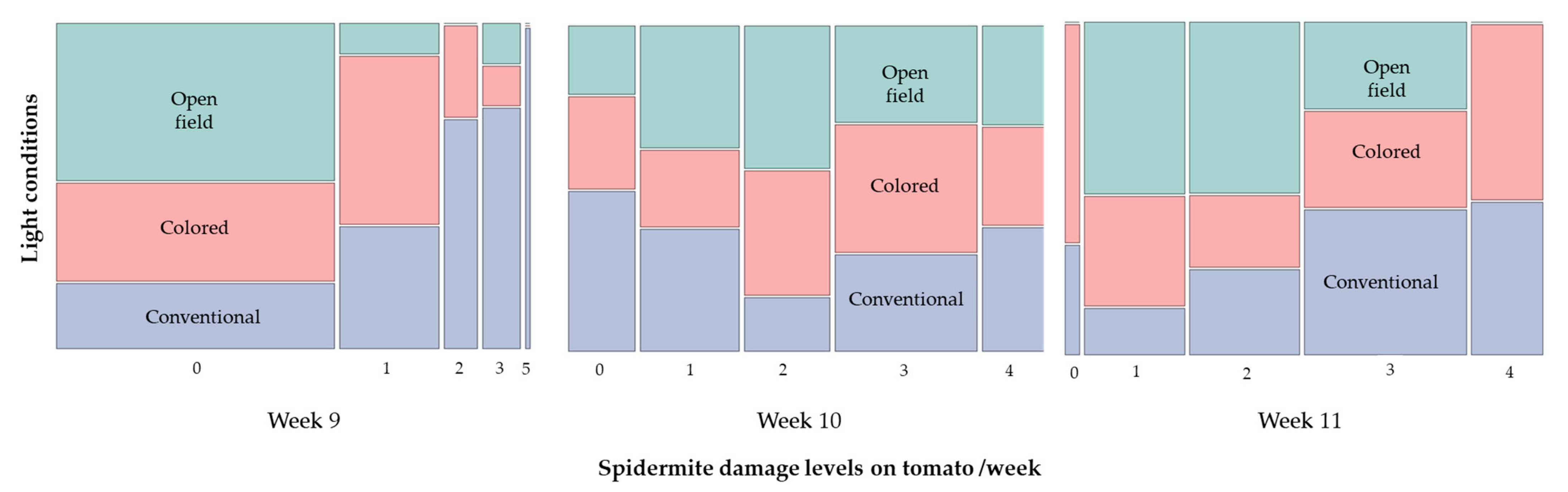
3.3. Monitoring of Insect Pests
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa, J.M.; Heuvelink, E. The Global Tomato Industry. In Tomatoes; CABI Publishing: Wallingford, UK, 2018; pp. 276–313. [Google Scholar]
- FAO. Crops and Livestock Products. Available online: http://www.fao.org/faostat/en/#data/QCL (accessed on 30 July 2021).
- Rajapakse, N.C.; Shahak, Y. Light-quality manipulation by horticulture industry. In Light and Plant Development. Annal Plant Reviews; Whitelam, G.C., Halliday, K.J., Eds.; Blackwell Publishing: Oxford, UK, 2007; Volume 30, pp. 290–312. [Google Scholar]
- Arsovski, A.A.; Galstyan, A.; Guseman, J.M.; Nemhauser, J.L. Photomorphogenesis. Arab. Book 2012, 10, e0147. [Google Scholar] [CrossRef] [Green Version]
- Bentsink, L.; Koornneef, M. Seed Dormancy and Germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, E.R.; Benítez, M.; Corvera-Poiré, A.; Chaos-Cador, A.; de Folter, S.; Gamboa de Buen, A.; Garay-Arroyo, A.; García-Ponce, B.; Jaimes-Miranda, F.; Pérez-Ruiz, R.V.; et al. Flower Development. Arab. Book 2010, 8, e0127. [Google Scholar] [CrossRef] [Green Version]
- Pedmale, U.V.; Celaya, R.B.; Liscum, E. Phototropism: Mechanism and Outcomes. Arab. Book 2010, 8, e0125. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Deng, X.W. Phytochrome Signaling Mechanism. Arab. Book 2004, 3, e0074. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The Cryptochrome Blue Light Receptors. Arab. Book 2010, 8, e0135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, I.; Pokorny, R.; Byrdin, M.; Hoang, N.; Ritz, T.; Brettel, K.; Essen, L.O.; van der Horst, G.T.; Batschauer, A.; Ahmad, M. The cryptochromes: Blue light photoreceptors in plants and animals. Annu. Rev. Plant. Biol. 2011, 62, 335–364. [Google Scholar] [CrossRef]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schafer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Shahak, Y.; Gussakovsky, E.E.; Gal, E.; Ganelevin, R. Colornets: Crop protection and light-quality manipulation in one technology. Acta Hortic. 2004, 659, 143–151. [Google Scholar] [CrossRef]
- Kotilainen, T.; Robson, T.M.; Hernández, R. Light quality characterization under climate screens and shade nets for controlled-environment agriculture. PLoS ONE 2018, 13, e0199628. [Google Scholar] [CrossRef] [PubMed]
- Ilić, Z.S.; Fallik, E. Light quality manipulation improves vegetable quality at harvest and postharvest: A review. Environ. Exp. Bot. 2017, 139, 79–90. [Google Scholar] [CrossRef]
- Ilić, S.Z.; Milenković, L.; Šunić, L.; Fallik, E. Effect of coloured shade-nets on plant leaf parameters and tomato fruit quality. J. Sci. Food Agric. 2015, 95, 2660–2667. [Google Scholar] [CrossRef]
- Selahle, M.K.; Sivakumar, D.; Soundy, P. Effect of photo-selective nettings on postharvest quality and bioactive compounds in selected tomato cultivars. J. Sci. Food Agric. 2014, 94, 2187–2195. [Google Scholar] [CrossRef]
- Díaz-Pérez, J.C. Bell pepper (Capsicum annuum L.) crop as affected by shade level: Fruit yield, quality, and postharvest attributes, and incidence of phytophthora blight (caused by Phytophthora capsici Leon.). HortScience 2014, 49, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Süß, A.; Danner, M.; Obster, C.; Locherer, M.; Hank, T.; Richter, K. Measuring leaf chlorophyll content with the Konica Minolta SPAD-502Plus—Theory, Measurement, Problems, Interpretation. EnMAP Field Guides Tech. Rep. GFZ Data Serv. 2015. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. Harvest Plus Handbook for Carotenoids Analysis; Harvest Plus Technical Monograph 2; International Food Policy Research Institute (IFPRI): Washington, DC, USA; International Center for Tropical Agriculture (CIAT): Cali, Colombia, 2004; p. 58. [Google Scholar]
- Pelletier, O. Vitamin C (L-ascorbic and dehydro-L-ascorbic acids). In Methods of Vitamin Assay, 4th ed.; Augustin, J., Klein, B.P., Becker, D.A., Venugopal, P.B., Eds.; Wiley: New York, NY, USA, 1985; pp. 303–347. [Google Scholar]
- Nihoul, P.; Van Impe, G.; Hance, T. Characterizing indices of damage to tomato by the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae) to achieve biological control. J. Hortic. Sci. 1991, 66, 643–648. [Google Scholar] [CrossRef]
- Jiang, C.; Johkan, M.; Hohjo, M.; Tsukagoshi, S.; Maruo, T. A correlation analysis on chlorophyll content and SPAD value in tomato leaves. Hort. Res. 2017, 71, 37–42. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Šunić, L.; Manojlović, M. Color shade nets improve vegetables quality at harvest and maintain quality during storage. Contemp. Agric. 2018, 67, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Nemeskéri, E.; Neményi, A.; Bőcs, A.; Pék, Z.; Helyes, L. Physiological factors and their relationship with the productivity of processing tomato under different water supplies. Water 2019, 11, 586. [Google Scholar] [CrossRef] [Green Version]
- Boardman, N.K. Comparative photosynthesis of sun and shade plants. Ann. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Berry, S.; Uddin, M. Effect of high temperature on fruit set in tomato cultivars and selected germplasm. HortScience 1988, 23, 606–608. [Google Scholar]
- Peet, M.M.; Willits, D.; Gardner, R. Response of ovule development and post-pollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. J. Exp. Bot. 1997, 48, 101–111. [Google Scholar] [CrossRef]
- Ayankojo, I.T.; Morgan, K.T. Increasing air temperatures and its effects on growth and productivity of tomato in South Florida. Plants. 2020, 9, 1245. [Google Scholar] [CrossRef]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and shade-avoidance responses in plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Schrager-Lavelle, A.; Herrera, L.A.; Maloof, J.N. Tomato phyE is required for shade avoidance in the absence of phyB1 and phyB2. Front. Plant Sci. 2016, 7, 1275. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Whitelam, G. Phytochrome, a family of photoreceptors with multiple physiological roles. Plant Cell Environ. 1990, 13, 695–707. [Google Scholar] [CrossRef]
- Nozue, K.; Tat, A.V.; Devisetty, U.K.; Robinson, M.; Mumbach, M.R.; Ichihashi, Y.; Lekkala, S.; Maloof, J.N. Shade avoidance components and pathways in adult plants revealed by phenotypic profiling. PLoS Genet. 2015, 11, e1004953. [Google Scholar] [CrossRef] [Green Version]
- de Wit, M.; George, G.M.; Ince, Y.C.; Dankwa-Egli, B.; Hersch, M.; Zeeman, S.C.; Fankhauser, C. Changes in resource partitioning between and within organs support growth adjustment to neighbor proximity in Brassicaceae seedlings. Proc. Natl. Acad. Sci. USA 2018, 115, E9953–E9961. [Google Scholar] [CrossRef] [Green Version]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [Green Version]
- Gent, M.P.N. Effect of degree and duration of shade on quality of greenhouse tomato. HortScience 2007, 42, 514–520. [Google Scholar] [CrossRef]
- García-Valverde, V.; Navarro-Gonzáles, I.; García-Alonso, J.; Periago, J.M. Antioxidant bioactive compounds in selected industrial processing and fresh consumption tomato cultivars. Food Bioprod. Technol. 2013, 6, 391–402. [Google Scholar] [CrossRef]
- Lee, G.H.; Bunn, J.M.; Han, Y.J.; Christenbury, G.D. Ripening characteristics of light irradiation tomatoes. J. Food Sci. 1997, 62, 138–140. [Google Scholar] [CrossRef]
- Alba, R.; Cordonnier-Pratt, C.; Pratt, L.H. Fruit-localized phytochromes regulate lycopene accumulation independently of ethylene production in tomato. Plant Physiol. 2000, 123, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Brandt, S.; Pék, Z.; Barna, E.; Lugasi, A.; Helyes, L. Lycopene content and colour of ripening tomatoes as affected by environmental conditions. J. Sci. Food Agric. 2006, 86, 568–572. [Google Scholar] [CrossRef]
- Kaur, S.; Srinivasan, R.; Cheema, D.; Lal, T.; Ghai, T.; Chadha, M. Monitoring of major pests on cucumber, sweet pepper and tomato under net-house conditions in Punjab, India. Pest. Manag. Hortic. Ecosyst. 2010, 16, 148–155. [Google Scholar]
- Saidi, M.; Gogo, E.O.; Itulya, F.M.; Martin, T.; Ngouajio, M. Microclimate modification using eco-friendly nets and floating row covers improves tomato (Lycopersicon esculentum) yield and quality for small holder farmers in East Africa. Agric. Sci. 2013, 4, 577. [Google Scholar] [CrossRef] [Green Version]
- Nordey, T.; Basset-Mens, C.; De Bon, H.; Martin, T.; Déletré, E.; Simon, S.; Parrot, L.; Despretz, H.; Huat, J.; Biard, Y.; et al. Protected cultivation of vegetable crops in sub-Saharan Africa: Limits and prospects for smallholders. A review. Agron. Sustain. Dev. 2017, 37, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.E.; Tao, Y.; Chory, J.; Ballare, C.L. Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proc. Natl. Acad. Sci. USA 2009, 106, 4935–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.A.; Kearney, E.E.; Hastings, A.P.; Ramsey, T.E. Attenuation of the jasmonate burst, plant defensive traits, and resistance to specialist monarch caterpillars on shaded common milkweed (Asclepias syriaca). J. Chem. Ecol. 2012, 38, 893–901. [Google Scholar] [CrossRef]
- de Wit, M.; Spoel, S.H.; Sanchez-Perez, G.F.; Gommers, C.M.M.; Pieterse, C.M.J.; Voesenek, L.A.; Pierik, R. Perception of low red: Far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J. 2013, 75, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Li, L. Hormonal regulation in shade avoidance. Front. Plant Sci. 2017, 8, 1527. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Open Field | Colored Net House | Conventional Net House |
|---|---|---|---|
| Material | -- | Polyethylene | Polyethylene |
| Radiance range (W⋅m−2) | 336.5–458.7 | 96.9–192.5 | 124.3–228.9 |
| PPFD range (μmol cm−2 s−1) | 1544.9–2304.1 | 457.9–1019.8 | 573.2–1153.8 |
| Luminance range (lux) | 82,520–86,590 | 19,559–24,232 | 32,248–41,780 |
| Red (R) range % | 37.5–38.9 | 52.1–53.2 | 38.5–39.2 |
| Green (G) range % | 35.9–36.0 | 23.9–24.1 | 36.1–36.2 |
| Blue (B) range % | 26.0–27.1 | 23.2–24.7 | 25.3–26.1 |
| R/FR range % | 123.0–127.2 | 117.6–122.8 | 124.0–126.4 |
| B/R range % | 66.9–72.2 | 43.9–47.5 | 64.7–67.8 |
| Treatment | N | Yield (t/ha) | Lycopene (g/ha) | ß-Carotene (g/ha) | Vitamin C (g/ha) |
|---|---|---|---|---|---|
| Open field | 4 | 11.6 ± 2.4 | 645.7 ± 133.4 b | 92.6 ± 19.1 | 3911.6 ± 807.9 |
| Colored net house | 4 | 16.1 ± 4.0 | 1846.6 ± 464.5 a | 116.2 ± 29.2 | 5796.9 ± 1458.2 |
| Conventional net house | 4 | 15.7 ± 4.3 | 1041.6 ± 285.8 b | 130.2 ± 35.7 | 4228.2 ± 1160.1 |
| F 2,11 | 1.76 | 12.67 | 1.69 | 2.87 | |
| Pr > F | 0.2510 | 0.0070 | 0.2616 | 0.1332 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotelo-Cardona, P.; Lin, M.-Y.; Srinivasan, R. Growing Tomato under Protected Cultivation Conditions: Overall Effects on Productivity, Nutritional Yield, and Pest Incidences. Crops 2021, 1, 97-110. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020010
Sotelo-Cardona P, Lin M-Y, Srinivasan R. Growing Tomato under Protected Cultivation Conditions: Overall Effects on Productivity, Nutritional Yield, and Pest Incidences. Crops. 2021; 1(2):97-110. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020010
Chicago/Turabian StyleSotelo-Cardona, Paola, Mei-Ying Lin, and Ramasamy Srinivasan. 2021. "Growing Tomato under Protected Cultivation Conditions: Overall Effects on Productivity, Nutritional Yield, and Pest Incidences" Crops 1, no. 2: 97-110. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020010